

Abstract
Objectives
Psychobiological research with adolescent populations tends to focus on negative mood, stress, and psychopathology, but the role of positive emotions is insufficiently understood. The current study examines the relative contributions of both negative and positive affective experiences to the basal activity of the hypothalamic-pituitary-adrenal (HPA) axis, measured by levels of cortisol across the waking day.
Methods
A sample of 315 ethnically and racially diverse high school students (Mage = 17.1, 73% female) completed a multiple-day naturalistic salivary cortisol protocol twice over a 5-year period. Along with each saliva sample, youth provided diary reports of their current mood states. Principal components analysis revealed four factors: high arousal positive affect (PA), low arousal PA, high arousal negative affect (NA), and low arousal NA.
Results
Multilevel growth curve models suggested that greater high arousal PA was associated with adaptive patterns of HPA activity: steeper cortisol slope from waking to bedtime and lower evening cortisol, independent of NA. In addition, increases in high arousal PA over the 5-year follow-up period were associated with a steepening of the diurnal cortisol slope (β = -.038, p = .009; negative values indicate the decrease of cortisol throughout the day) and lower evening cortisol levels (β = -.661, p = .027) based on within-person fixed effect regression analysis.
Conclusion
This study shows that high arousal positive affect, such as feeling alert and active, is associated with a steeper decline in cortisol throughout the day. Low arousal positive emotions did not display this relationship.
Keywords: adolescent development, circumplex model of affect, diurnal cortisol, HPA axis
Psychobiological research tends to focus on negative mood, stress, and psychopathology, with relatively little emphasis on the role of positive emotions. This trend is evident in research on the hypothalamic-pituitary-adrenal (HPA) axis, one of the key biological stress systems thought to mediate the connections between psychological experiences and health outcomes (1). There is now convincing evidence that negative social and affective experiences such as anger, sadness, loneliness, depressed mood, and perceived stress are associated with dysregulated patterns of cortisol secretion (2-5), the major hormonal output of the HPA axis. However, much less is known about the role of positive affective experiences in relation to cortisol, especially during adolescence. Although traditionally described as a vulnerable period for stress and emotional turmoil, recent research and theory has now reframed adolescence as a time of heightened arousal and emotional intensity (6, 7). Therefore, psychobiological approaches to adolescence would be enhanced by studying both negative and positive expressions of affect and arousal.
Cortisol follows a strong circadian rhythm. Typically, cortisol levels are high upon waking, increase to a peak about 30-45 minutes after waking (the cortisol awakening response, CAR), and then decline throughout the remainder of the day (8, 9). The rate of decline in cortisol from waking to bedtime (cortisol slope) represents an important marker of diurnal HPA activity, with flatter slopes indicating a slower decline in cortisol from waking to bedtime. There is empirical evidence that flattened diurnal cortisol slopes may be implicated in risk for diseases, including depression and heart disease, and faster progression of cancer (10-13). Therefore, diurnal changes in cortisol activity represent meaningful individual differences in stress biology that may play a role in future health (14).
Accumulating evidence from medical and social sciences suggests that negative affect (NA) and positive affect (PA) both make important and independent contributions to health outcomes, which underlines the need for a revised model of the role of affect in the stress process (15, 16). The current evidence regarding PA and HPA activity remains limited and inconsistent. For instance, Steptoe and colleagues found that PA (measured as happiness only, averaged over two days) was associated with lower total salivary cortisol across the day, independent of age, gender, socioeconomic status, smoking, and NA (17, 18). However, this finding has not been replicated across other adult samples (19, 20) and has never been examined in an adolescent sample. Similarly, although there is some empirical support that PA is associated with a steeper cortisol slope in adulthood (21, 22), other studies have failed to find a direct association between PA and cortisol slope (23-25) or even found that PA was associated with a flatter diurnal slope (19). None of the research to date has taken into account level of arousal, an important dimension introduced by the circumplex model of affect (26).
The circumplex model of affect suggests that every affective experience is the consequence of both valence and arousal, two independent neurophysiological systems that together can be used to represent a particular emotional experience (26, 27). In this model, affect exists on two axes: one describing positive versus negative valence, and the other describing high versus low activation. Therefore, the circumplex model allows the prediction of differential effects of high arousal PA (e.g., excited, active), low arousal PA (e.g., calm, placid), high arousal NA (e.g., fearful, hostile), and low arousal NA (e.g., drowsy, dull). This distinction across valence and arousal may be especially important when studying the physiology of the HPA axis during adolescence, a developmental period associated with alterations in both stress physiology as well as alterations in the neurobiology of affect, and in particular, the neurobiology of positive affect (28, 29).
There are biologically-based changes in neural systems of emotions at the beginning of adolescence, which contribute to a natural inclination towards strong emotions, replete with intensity and arousal (7). The impact of strong negative emotions on health, stress, and well-being has received significant attention, particularly the sharp rise in clinical depression and anxiety disorders during this developmental period (32, 33) and the relationship between negative mood and HPA dysfunction (2, 4, 34). However, current research and theory on positive youth development underlines the equally important role of positive impacts (35, 36). Beyond risk, adolescents seek excitement, thrills, and activity – positive feelings characterized by high arousal and intensity that youth experience in their daily lives (7). The current study examines the contribution of affect (positive and negative valence), activation (high and low arousal), and intensity (strong emotions) to adolescent HPA axis activity, both across individuals and within-individuals over time.
Design and Methods
Participants
Data for the current study were collected as part of the Youth Emotion Project, a two site longitudinal investigation examining risk factors for mood and anxiety disorders in late adolescence (37-42). Participants were drawn from two ethnically and socioeconomically diverse high schools in suburban Chicago and Los Angeles. Students in their junior year of high school were screened and selected to participate based on their neuroticism score from the Eysenck Personality Questionnaire – Revised (EPQ-R-N) (43), a personality variable that increases the risk for subsequent development of mood and anxiety disorders (44, 45). Those scoring in the top third of this measure were oversampled, comprising 59% of the final group of 627 who consented to be part of the longitudinal study and completed the baseline assessment across three consecutive cohorts. Two thirds of the participants were randomly selected for the cortisol study subsample. Of those invited, 350 youth (71%) agreed to participate and completed the cortisol task. Exclusion criteria for the current analyses were use of corticosteroid-based medications (N=11), or antipsychotic medications (N=3). Additionally, twenty participants were excluded due to insufficient cortisol data (i.e., fewer than 50% of all samples and/or missing all wakeup or bedtime samples). The final analytic sample included 315 adolescents (230 female) with a mean age of 17.05 (SD = .38) at baseline.
The greater proportion of females over males is accounted for by the fact that females are, on average, higher on the neuroticism personality trait (46). Although use of this high-neuroticism sample limits generalizability, it also provides a larger range and more normal distribution of PA for both males and females, which usually has ceiling effects in the general population (i.e. positive moods are negatively skewed, with most peoples' responses clustering at the high end). We examined the differences between those in the top third of the neuroticism measure (i.e., high neuroticism risk, N = 192) compared to the rest of the sample on key demographic and cortisol parameters. Results revealed that youth in the high risk group did not differ from the rest of the sample based on age, gender, racial/ethnic background, socioeconomic status, waking cortisol, cortisol awakening response, bedtime cortisol, or diurnal cortisol slope.
Procedure
As part of the larger investigation, all youth completed the Structured Clinical Interview for DSM–IV-TR (47), interviews for chronic and episodic stress, and a series of questionnaires to measure additional health and demographic characteristics every year. In addition, the cortisol study subsample participated in a diary study utilizing a modified experience sampling method (ESM) (48) protocol six times a day for three consecutive weekdays (18 samples total). Adolescents were asked to provide saliva samples and diary reports at wakeup, 40-minutes after wakeup, and immediately prior to bedtime (to model the cortisol diurnal rhythm). Additionally, adolescents wore wristwatches that beeped at semi-random moments to prompt three additional sampling times across the day (approximately 3, 8, and 12 hours after participants' typical wakeup times).
Along with every saliva sample, youth were asked to report, in paper diaries, where they were, who they were with, and what they were thinking and feeling at the time. Diary reports also asked youth specific questions regarding sleep and health-related behaviors occurring in the past hour that may influence cortisol levels. All procedures were reviewed and approved by Institutional Review Boards at Northwestern University and the University of California at Los Angeles.
Measures
Cortisol
At each sampling point, participants expelled a small, passive drool saliva sample through a straw into a sterile 2 mL polypropylene cryogenic vial. They were asked to report the exact time of each sample, and to store the sample in the refrigerator as soon as possible after completion. Participants were asked not to eat or drink in the 30 minutes prior to each sample, and for the unanticipated beeps, were asked to record whether or not they had consumed food or beverages in the hour prior to that beep. Completed samples (labeled with exact date and time) and all other study materials were returned in one packet through a school drop box or by regular postal mail. Once returned to the laboratory, samples were refrigerated at -20 degrees Celsius until they were sent by courier on dry ice to Biochemisches Labor at the University of Trier, Germany to be assayed.
Assays were conducted in duplicate using a time-resolved immunoassay with fluorometric detection (DELFIA) (49). Intra-assay coefficients of variation (CVs) were between 4.0% and 6.7%, and inter-assay CVs ranged from 7.1% to 9.0%. To correct for a strong positive skew in the cortisol distribution, cortisol values were natural logarithmically transformed prior to analysis. The transformation substantially reduced the non-normality of the data and all analyses include robust standard errors, which can compensate for minor deviations from normality.
Positive and negative affect
We combined ESM measures across all three days of data collection as our measure of typical affective state. Aggregation of ESM measures is thought to provide a more valid estimate of typical affective states than questionnaire-based reporting of typical affect (50). Notably, in a study that examined both ESM and survey methods to capture PA, effects on neuroendocrine and cardiovascular activity were substantially stronger when positive affect was assessed by aggregating ESM samples than with questionnaire measures (25).
Participants were asked to indicate in their diary reports how much they felt each of the following mood states at the time of each sampling occasion: happy, nervous, lonely, active, frustrated, alert, worried, relaxed, irritable, stressed, sad, and cheerful. Each mood state was rated on a scale of 0 (not at all) to 3 (very much). Four affect scales were created based on a principal components analysis (PCA) of all diary-reported moods states, with orthogonal rotation. PCA weights were used to construct scales such that the resulting uncorrelated factors could be included in a single model. The four affect scales align with the circumplex model of affect (26, 51): PA-high arousal (active, alert; ρ = .700), PA-low arousal (happy, cheerful, relaxed; α = .839), NA-high arousal (nervous, frustrated, worried, irritable, stressed; α = .906), and NA-low arousal (sad, lonely; ρ = .802). Each component had an eigenvalue > 1 and together explain 78% of the variance (Table S1, Supplemental Digital Content 1). We also created a quadratic version of each affect scale by squaring scale values, for use in modeling the impact of high intensity moods (i.e., strong emotions).
Covariates
Basal cortisol levels may be influenced by a number of health and demographic factors including sleep, nicotine, oral contraceptive use, race/ethnicity, socioeconomic status (SES), and major depressive disorder (MDD) (40, 52-56). Each of these factors were therefore measured and included as covariates in the final models. In the daily diaries, participants' recorded their wake time each day. Participants also responded to health and demographic questionnaires that assessed their typical hours of sleep, use of oral contraceptives (females only), nicotine use, race/ethnicity, and family SES (computed from parents' average level of education). Recent MDD (current or within the last 3 months) was assessed using the Structured Clinical Interview for DSM-IV-TR Axis I Disorders. Finally, we included an indicator for neuroticism risk, the sampling variable, which was measured by the EPQ-R-N.
Analytic Plan
Multilevel growth models
A three-level growth curve analysis was run to model the diurnal rhythm of cortisol, to examine person-level predictors of that rhythm, and to adjust for the nonindependence of observations associated with nested data. The model included momentary salivary cortisol samples (Level 1) nested within day (Level 2), nested within person (Level 3). Based on prior work (2, 57), three time of day indicators were included in Level 1 to capture the key elements of the cortisol diurnal rhythm: a dummy variable representing the CAR (to model the increase in cortisol from waking to 40 minutes after waking), time of day (to model the decline of cortisol across the day), and a time of day squared parameter (to model the curvilinear shape of the rhythm). Time was centered at waking, and represented hours since waking, which allows the intercept to be interpreted as the waking value. In a separate set of models, time was re-centered at bedtime, representing hours until bedtime, such that the intercept could be interpreted as bedtime cortisol level and we could examine associations between our affect variables and bedtime cortisol levels.
Daily wake time was entered at Level 2 to adjust for any impact of time of waking on cortisol rhythms each day, and person-level predictors and covariates were entered at Level 3 to predict differences in basal cortisol rhythms estimated across all three study days. The affect factors (both linear and quadratic) were included in Level 3 to determine whether high and low arousal PA predicts wakeup cortisol, the CAR, cortisol slopes across the day, and bedtime cortisol levels. Importantly, high and low arousal NA were entered into the model at the same time as the PA variables, to test the unique effect of each affect factor on cortisol parameters.
| (1.1) |
| (1.2) |
| (1.3) |
Longitudinal models
All participants in the first wave of cortisol sampling (Time 1) were asked to complete another wave of cortisol sampling data collection using identical proceduresfive years after their baseline measures (Time 2). Therefore, a subset of youth (N=166; 53%) provided ESM and cortisol data at two time points over five years. To bolster our cross-sectional findings with longitudinal models of within-person change, in addition to the multilevel growth models described above, we also measured within-person changes in cortisol (e.g., cortisol slope, CAR) as a function of within-person changes in high and low arousal PA and NA over the 5-year time period, using an econometric approach called fixed-effects regression. The fixed effect model for panel data utilizes the following equation:
| (2.0) |
where Δ indicates the change in variables taking place between Time 1 and Time 2 and μi represents person-level characteristics that are constant over time. Time-varying unexplained variation is represented by ε. In the estimation stage, all stable characteristics drop out of the model (i.e., a time-invariant characteristic cannot cause such a change, because it is constant for each person). As a result, we can estimate the effect of PA and NA on cortisol only because affect changes over time.
This econometric model strengthens causal claims because it controls for the effects of time-invariant variables, reducing bias from potential selection effects driven by individual characteristics (58). In other words, the fixed-effects model controls for all time-invariant differences between the individuals, both observed (e.g., race/ethnicity or gender) and unobserved (e.g., culture or early childhood factors), so the estimated coefficients of the fixed-effects models are not biased because of omitted time-invariant characteristics. The only omitted variables that threaten a fixed effect design are those variables that change within individuals over time. In an attempt to reduce this bias, our analyses included key time-varying predictors of cortisol including wake time, typical sleep duration, and nicotine use.
Results
At Time 1, youth, on average, woke up at 6:49AM (SD = 6.81 minutes) and reported 7.15 hours of sleep per night (SD = .90). Fourteen youth in the sample met criteria for recent MDD, 17 students reported being a regular smoker and 26 females in the sample were using oral contraceptives. Details on cortisol parameters and demographic covariates are presented in Table 1, both for the overall sample and, for illustrative purposes, those at the extreme ends of each affect variable (i.e., those in the highest quartile). On average, the diurnal cortisol slopes across the day are negative (with a higher waking value and a lower bedtime value), such that a positive coefficient is indicative of “flatter” slopes, and a negative coefficient indicates steeper slopes (see Figure 1).
Table 1. Descriptive statistics for sample, presented by high and low arousal positive affect (PA) and negative affect (NA).
| Overall N=315 |
PA High Arousal N=79 |
PA Low Arousal N=79 |
NA High Arousal N=79 |
NA Low Arousal N=80 |
||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
| ||||||||||
| N | (%) | N | (%) | N | (%) | N | (%) | N | (%) | |
|
|
||||||||||
| Male | 85 | 27.0 | 24 | 30.4 | 24 | 30.4 | 12 | 15.2 | 18 | 22.5 |
| Race/ethnicity | ||||||||||
| White | 150 | 47.6 | 34 | 43.0 | 40 | 50.6 | 48 | 60.8 | 36 | 45.0 |
| Black | 33 | 10.5 | 11 | 13.9 | 10 | 12.7 | 7 | 8.9 | 10 | 12.5 |
| Asian/Pacific Islander | 18 | 5.7 | 5 | 6.3 | 2 | 2.5 | 2 | 2.5 | 4 | 5.0 |
| Latino | 59 | 18.7 | 19 | 24.1 | 17 | 21.5 | 6 | 7.6 | 9 | 11.3 |
| Multi-ethnic/ multi-racial | 38 | 12.1 | 8 | 10.1 | 8 | 10.1 | 11 | 13.9 | 12 | 15.0 |
| Other race/ethnicity | 17 | 5.4 | 2 | 2.5 | 2 | 2.5 | 5 | 6.3 | 9 | 11.3 |
| Regular nicotine use | 17 | 5.4 | 1 | 1.3 | 2 | 2.5 | 4 | 5.1 | 6 | 7.5 |
| Oral contraceptive use | 26 | 8.3 | 7 | 8.9 | 5 | 6.3 | 15 | 19.0 | 13 | 16.3 |
|
|
||||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
|
|
||||||||||
| Average hours of sleep | 7.150 | .90 | 7.125 | 1.01 | 7.316 | .94 | 6.998 | .83 | 6.954 | .93 |
| Neuroticism score | 11.840 | 61.0 | 11.34 | 4.44 | 9.843 | 5.18 | 13.700 | 3.22 | 13.984 | 2.95 |
| Positive Affect | ||||||||||
| Active | 1.045 | .47 | 1.599 | .35 | 1.372 | .49 | .983 | .42 | 0.984 | .45 |
| Alert | 1.352 | .61 | 2.068 | .31 | 1.768 | .53 | 1.382 | .54 | 1.239 | .61 |
| Happy | 1.612 | .56 | 1.982 | .51 | 2.249 | .34 | 1.456 | .41 | 1.417 | .48 |
| Cheerful | 1.106 | .62 | 1.612 | .59 | 1.799 | .49 | .972 | .51 | 0.966 | .54 |
| Relaxed | 1.432 | .57 | 1.714 | .58 | 2.052 | .44 | 1.241 | .46 | 1.217 | .52 |
| Negative Affect | ||||||||||
| Nervous | 0.422 | .47 | .377 | .45 | .308 | .43 | 0.993 | .53 | 0.737 | .62 |
| Frustrated | 0.777 | .51 | .688 | .55 | .533 | .43 | 1.375 | .40 | 1.148 | .53 |
| Worried | 0.623 | .54 | .501 | .53 | .402 | .49 | 1.328 | .44 | 1.040 | .59 |
| Irritable | 0.705 | .53 | .593 | .53 | .494 | .44 | 1.294 | .48 | 1.055 | .55 |
| Stressed | 0.928 | .68 | .803 | .68 | .581 | .57 | 1.710 | .52 | 1.478 | .65 |
| Sad | 0.368 | .44 | .306 | .44 | .234 | .37 | 0.721 | .51 | 0.955 | .44 |
| Lonely | 0.382 | .51 | .301 | .44 | .249 | .46 | 0.755 | .62 | 1.053 | .54 |
| Cortisol parameters | ||||||||||
| Wakeup cortisol | .417 | .21 | .435 | .23 | .414 | .25 | .402 | .19 | .399 | .18 |
| Bedtime cortisol | .107 | .15 | .097 | .13 | .118 | .23 | .133 | .23 | .144 | .22 |
| Cortisol awakening response (CAR) | .160 | .29 | .103 | .31 | .157 | .29 | .164 | .30 | .170 | .30 |
| Cortisol slope | -.107 | .05 | -.115 | .05 | -.107 | .05 | -.104 | .06 | -.094 | .06 |
Note. Raw cortisol values (μg/dl) and high/low positive and negative affect groups representing the highest quartile are presented for descriptive purposes only. Log transformed cortisol values and continuous affect measures are used in all analyses.
Figure 1. Average cortisol rhythms across the waking day by affect and arousal.
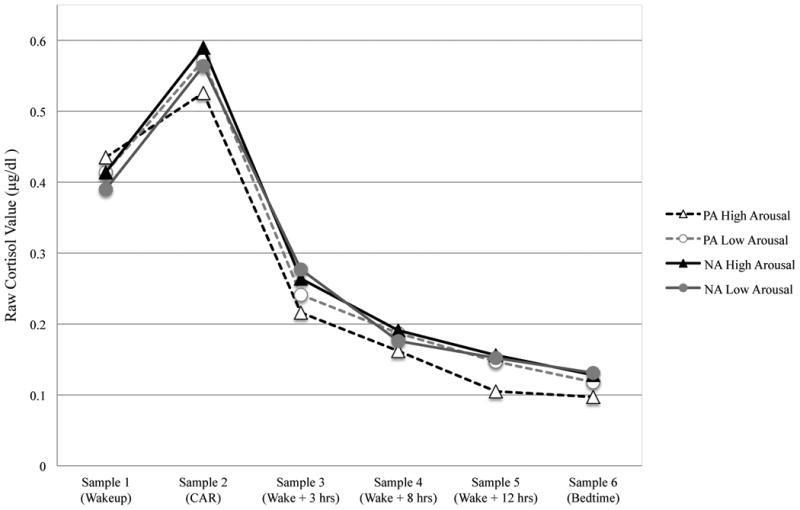
Diurnal cortisol analyses
We observed a number of findings based on the health and sociodemographic covariates, as reported in previous research on this sample (40, 41, 52). In particular, male gender was associated with lower wakeup levels of cortisol (γ = -.195, p = .001) and a smaller CAR (γ = -.253, p = .003). Black race was associated with lower wakeup levels of cortisol (γ = -.176, p = .036), higher bedtime levels of cortisol (γ = .433, p = .006), and a flatter slope across the waking day (γ =.038, p <.001). Greater total hours of sleep, on the other hand, were associated with higher cortisol at waking (γ = .064, p = .025) and a steeper slope across the day (γ = -.007, p = .032).
Next, we examined whether high and low arousal PA and/or NA were associated with diurnal cortisol parameters in this three-level multilevel growth curve analysis (Table 2). There was a marginally significant relationship between high arousal PA and lower levels of cortisol at bedtime (γ = -.067, p = .10), with a significant, negative quadratic term (γ = -.090, p = .007), illustrating that the effect of high arousal PA on lower bedtime cortisol increased at higher intensity levels (i.e., for stronger positive emotions). For the diurnal cortisol slope, the quadratic term was also significant and negative (γ = -.004 p = .037); the linear term was not significant (γ = -.002, p = .270), suggesting that only values at the high end of the high arousal PA scale were associated with a steeper cortisol slope. High arousal PA was not significantly associated with waking levels of cortisol or the CAR. There were also no statistically significant relationships between low arousal PA with any of the cortisol parameters.
Table 2. Three-Level Growth Model Examining the Effect of PA and NA on Log Transformed Cortisol (μg/dl). N = 315.
| Estimation of fixed effects | Coefficient | SE |
|---|---|---|
| Waking cortisol | ||
| Intercept | -1.160** | .027 |
| High arousal PA | -.030 | .023 |
| High arousal PA2 | -.017 | .017 |
| Low arousal PA | .007 | .026 |
| Low arousal PA2 | -.016 | .020 |
| High arousal NA | -.055† | .034 |
| High arousal NA2 | .042* | .019 |
| Low arousal NA | -.075* | .041 |
| Low arousal NA2 | .024 | .014 |
| Cortisol slope | ||
| Intercept | -.147** | .008 |
| High arousal PA | -.002 | .003 |
| High arousal PA2 | -.004* | .002 |
| Low arousal PA | .004 | .003 |
| Low arousal PA2 | -.001 | .002 |
| High arousal NA | .003 | .004 |
| High arousal NA2 | .001 | .002 |
| Low arousal NA | .011** | .004 |
| Low arousal NA2 | -.002 | .002 |
| Bedtime cortisol | ||
| Intercept | -2.939** | .041 |
| High arousal PA | -.067† | .041 |
| High arousal PA2 | -.090** | .030 |
| Low arousal PA | .073 | .041 |
| Low arousal PA2 | -.027 | .031 |
| High arousal NA | -.017 | .053 |
| High arousal NA2 | .052 | .031 |
| Low arousal NA | .106† | .059 |
| Low arousal NA2 | -.019 | .024 |
|
| ||
| Variance explained | Pseudo R2 | |
|
| ||
| Wakeup cortisol | .267 | |
| Cortisol slope | .252 | |
| Bedtime cortisol | .194 | |
Note. Covariates include a time since waking squared variable to capture the quadratic curvilinear components of change in cortisol across the day and a dummy variable indicating the CAR (Level 1); daily wake time (Level 2); and race/ethnicity, average hours sleep, oral contraceptives use, nicotine use, recent MDD, and neuroticism risk (Level 3). The estimates for bedtime cortisol were measured in a separate HLM model with time centered at bedtime.
Multiply cortisol values (μg/dL) by 27.59 to obtain nmol/L.
p < 0.01,
p < 0.05,
p < 0.1
High arousal NA was marginally associated with lower cortisol at waking (γ = -.055, p = .091), with a significant, positive quadratic term (γ = .042, p = .030), suggesting that at very high levels of high arousal NA, the relationship between high arousal NA and waking cortisol reverses. Since these scales are standardized, it can be interpreted that starting at high intensity levels of the scale (at least 1.3 SDs above the mean), high arousal NA is associated with higher levels of waking cortisol. Low arousal NA (sad, lonely) was associated with lower cortisol upon waking (γ = -.075, p = .038), a flatter slope across the waking day (γ = .011, p = .007), and marginally associated with higher levels of cortisol at bedtime (γ = .106, p = .079).
Within-person change over five years
Overall, most participants showed a decrease in morning cortisol level (average within-person decrease of .163 μg/dl) across five years, from Time 1 to Time 2. However, changes in bedtime cortisol level (.016 μg/dl lower at Time 2), CAR (.023 μg/dl lower at Time 2), and cortisol slope (.005 μg/dl flatter at Time 2) were modest. There were also small mean within-person changes in affect – with overall increases in both high and low arousal PA and overall decreases in both high and low arousal NA – from Time 1 to Time 2 (Table S2, Supplemental Digital Content 2).
The fixed effects models, examining change in affect and cortisol over a 5-year time period, are presented in Table 3. The estimated effect is interpreted as the effect of a within-person change in high and low arousal PA or NA on change in cortisol rhythms over time, controlling for both measured and unmeasured time-invariant covariates. High arousal PA was the only affect factor to predict within-person changes in cortisol. Estimates of the coefficients in Models 1-2 show a significant association between high arousal PA and the diurnal cortisol slope (β = -.038, p = .009) and bedtime cortisol (β = -.661, p = .027) over the 5 year time period. That is, an increase in high arousal PA was associated with a steepening of the diurnal cortisol slope and lowering of evening cortisol over time, controlling for all time-invariant covariates.
Table 3. Fixed Effect Regression Examining Within Person Change in Average PA and NA on Within Person Change in Log Transformed Cortisol (μg/dl) from Time 1 to Time 2. N = 166.
| Model 1 Diurnal Cortisol Slope |
Model 2 Bedtime Cortisol |
|||
|---|---|---|---|---|
|
| ||||
| β | SE | β | SE | |
| Affect variables | ||||
| PA – high arousal | -.038** | .015 | -.661* | .295 |
| PA – low arousal | .021 | .015 | .111 | .305 |
| NA – high arousal | -.009 | .105 | -.387 | .380 |
| NA – low arousal | .021 | .019 | .508 | .347 |
| Health covariates | ||||
| Mean wake time | -.009* | .004 | -.209** | .077 |
| Average hours of sleep | -.005 | .005 | -.153 | .105 |
| Nicotine use | .053** | .019 | 1.131** | .358 |
| Oral contraceptive use | .021* | .010 | .213 | .173 |
|
| ||||
| R2 within = .160 | R2 within = .208 | |||
Note. Multiply cortisol values (μg/dL) by 27.59 to obtain nmol/L.
p < 0.01,
p < 0.05,
p < 0.1
Discussion
Past research has focused on HPA dysregulation (e.g., hypo- or hyper-cortisolemia) as markers of poor mental and physical health outcomes. For instance, many researchers have examined changes in normal HPA axis activity as possible mechanisms to explain the sharp rise in clinical depression and anxiety disorders during adolescence (32, 33). Because cortisol is commonly known as a “stress hormone,” few studies have attempted to link variations in diurnal cortisol to positive psychological experiences. The current study addresses this gap in the literature by examining the relationship between multiple measures of positive affect and salubrious HPA functioning in a diverse sample of high school students.
The most novel finding from the current study was that high arousal positive affect was associated with healthy patterns of cortisol secretion across the day. Youth with elevated, high intensity active and alert moods had a steeper decline in cortisol than their less energetic peers. Additional analyses uncovered that the steeper decline was primarily driven by lower evening cortisol, which could be a reflection of more efficient negative feedback control of the HPA axis. The inability to suppress cortisol at night, leading to prolonged glucocorticoid exposure, is a hypothesized mechanism linking flat cortisol slopes to a variety of poor health outcomes including obesity (59), depression (60), and all-cause mortality (12). On the other hand, a steep diurnal pattern of cortisol secretion across the day (high on awakening, moderate CAR with rapid recovery, and low in the evening, defining a steep slope across the day from waking to bedtime) could prove to be a useful indicator of good health (3, 57, 61). High arousal PA may therefore promote or sustain healthy psychobiological trajectories across adolescent development.
Importantly, our analyses control for a number of potentially confounding variables, including wake time and sleep duration. Previous research has reported a reciprocal relationship between sleep and basal patterns of cortisol across the day, with more hours of sleep predicting a steeper cortisol slope, and higher waking cortisol predicting more sleep and lower fatigue the following day (2, 52). Furthermore, both laboratory and epidemiological data have shown that shorter sleep duration and poor sleep quality are associated with a flatter cortisol slope and increased evening levels (62-64). The current study suggests that active and alert moods predict diurnal cortisol across the day, above and beyond these sleep-specific predictors of cortisol activity.
An additional methodological contribution of the study is the use of fixed effects estimation. This technique is commonly used in economics and has more recently been applied in developmental and health research to reduce the impact of confounding by time-invariant factors, such as the unmeasured characteristics of the individual. In these within-person analyses, youth who experienced increases in high arousal positive affect between adolescence and young adulthood also demonstrated steepening cortisol slope and lowering bedtime cortisol across the same time period.
Given that both high arousal (e.g., anger, stress) and low arousal (e.g., sad, depressed) negative emotions have been linked to measures of cortisol activity in the past (2, 4), it is often assumed that valence, rather than arousal, is the most important emotional predictor of cortisol activity. The current study, however, highlights the importance of arousal, particularly high intensity feelings of arousal, when studying the physiological mechanisms linking PA and HPA axis activity in adolescents. Strong emotions may be particularly salient for youth, relative to both younger children and adults, due to neurobiological changes that occur at the beginning of adolescence. Research on adolescent brain development suggests differential development of bottom-up limbic systems, implicated in incentive and emotional processing, to prefrontal top-down control systems during this developmental period. This imbalance model proposes that differential developmental timing of these regions (i.e., less top-down regulation) accounts for heightened emotionality during adolescence ([30, 31]; see also [65] for a critical review of this hypothesis).
Although high and low arousal positive emotions often co-occur, these different expressions of emotion may have distinct psychobiological profiles. We also found that low arousal NA (but not high arousal NA) during adolescence was associated with lower waking cortisol and a flatter cortisol slope across the day, a finding that is consistent with past research (4). Further, although moderate levels of high arousal NA during adolescence may be associated with lower cortisol upon waking, youth at the extreme levels of high arousal NA show a reversal in this pattern (i.e., higher waking cortisol). The associations between NA and basal cortisol, however, were not replicated in the longitudinal analyses. Future research is needed to uncover the role of emotional arousal and intensity in diurnal cortisol in adult samples, which may help elucidate inconsistencies across past research studies.
Because measuring PA was not an original goal of the larger study from which these data were drawn, we were limited by the type and quantity of arousal-specific emotions we could measure. Future research should seek to examine a wider range of arousal-specific emotions such as: excitement, vigor, pep, strength, and exuberance. The current study is also limited in terms of generalizability (i.e., higher proportion of female and neurotic youth) and causality (i.e., unknown causal direction). The greater proportion of females over males is accounted for by the fact that females usually score higher than males on neuroticism, the personality trait that was oversampled at baseline (46), and females were more likely to agree to participate in the study if invited. A few previous studies have found differences in diurnal cortisol rhythms in adult subjects high in neuroticism as compared to those with low neuroticism (66, 67), while others have not (68, 69). Importantly, neuroticism risk did not significantly predict any diurnal cortisol parameters in the current analyses, nor did it change point estimates when it was included as a covariate. Finally, although many studies conclude that stressful or emotional experiences are responsible for alterations in diurnal cortisol activity, causal directions are not yet firmly established (70). While emotional experience may influence cortisol levels, it is also possible that cortisol could influence perceived arousal level and perceived emotional experience (3).
Despite these limitations, the current study underlines the importance of examining both valence and arousal to uncover the complex psychobiology of youth emotions, and highlights the role of positive affective experiences in diurnal cortisol activity. High arousal positive affect may be a particularly salient influence of the HPA axis during adolescence, a developmental period characterized by periodic heightened emotional intensity and a proclivity towards excitement and activity.
Supplementary Material
Acknowledgments
Sources of Funding: This research was conducted with the support of a William T. Grant Foundation Scholars Award (Emma K. Adam, Principal Investigator), NIMH 1 R01 MH65651-01 (Michelle G. Craske, Principal Investigator), NIHM R01 MH65652 (Richard E. Zinbarg and Susan Mineka, Principal Investigators), and funding from both the American Association of University Women (AAUW American Fellowship) and the Society for Research in Child Development (SRCD SECC Dissertation Research Funding Award) to the first author (Lindsay T. Hoyt). The granting agencies involved had no further role in the study design, data collection, analysis and interpretation of the data, in the writing of the manuscript or the decision to submit the article for publication.
Abbreviations
- CAR
cortisol awakening response
- HPA
hypothalamic-pituitary-adrenal
- MDD
major depressive disorder
- NA
negative affect
- PA
positive affect
Footnotes
Conflicts of Interest: None of the authors have any conflicts of interest to declare with respect to this manuscript.
Contributor Information
Lindsay T. Hoyt, Robert Wood Johnson Foundation Health and Society Scholar, University of California at San Francisco and Berkeley.
Michelle G. Craske, Department of Psychology, University of California, Los Angeles.
Susan Mineka, Department of Psychology, Northwestern University.
Emma K. Adam, Program on Human Development and Social Policy, School of Education and Social Policy, Faculty Fellow, Institute for Policy Research, Northwestern University.
References
- 1.Cohen S, Rodriguez M. Pathways linking affective disturbances and physical disorders. Health Psychol. 1995;14:374. doi: 10.1037//0278-6133.14.5.374. [DOI] [PubMed] [Google Scholar]
- 2.Adam EK. Transactions among adolescent trait and state emotion and diurnal and momentary cortisol activity in naturalistic settings. Psychoneuroendocrinology. 2006;31:664–79. doi: 10.1016/j.psyneuen.2006.01.010. [DOI] [PubMed] [Google Scholar]
- 3.Adam EK, Hawkley LC, Kudielka BM, Cacioppo JT. Day-to-day dynamics of experience–cortisol associations in a population-based sample of older adults. PNAS. 2006;103:17058–63. doi: 10.1073/pnas.0605053103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Doane LD, Adam EK. Loneliness and cortisol: Momentary, day-to-day, and trait associations. Psychoneuroendocrinology. 2010;35:430–41. doi: 10.1016/j.psyneuen.2009.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.van Eck M, Berkhof H, Nicolson N, Sulon J. The effects of perceived stress, traits, mood states, and stressful daily events on salivary cortisol. Psychosom Med. 1996;58:447–58. doi: 10.1097/00006842-199609000-00007. [DOI] [PubMed] [Google Scholar]
- 6.Steinberg L. Risk taking in adolescence new perspectives from brain and behavioral science. Curr Dir Psychol Sci. 2007;16:55–9. [Google Scholar]
- 7.Dahl RE. Adolescent brain development: A period of vulnerabilities and opportunities. Keynote address. Ann N Y Acad Sci. 2004;1021:1–22. doi: 10.1196/annals.1308.001. [DOI] [PubMed] [Google Scholar]
- 8.Pruessner J, Wolf O, Hellhammer D, Buske-Kirschbaum A, Von Auer K, Jobst S, Kaspers F, Kirschbaum C. Free cortisol levels after awakening: a reliable biological marker for the assessment of adrenocortical activity. Life Sci. 1997;61:2539–49. doi: 10.1016/s0024-3205(97)01008-4. [DOI] [PubMed] [Google Scholar]
- 9.Kirschbaum C, Hellhammer DH. Salivary cortisol in psychobiological research: A overview. Neuropsychobiology. 1989 doi: 10.1159/000118611. [DOI] [PubMed] [Google Scholar]
- 10.Matthews K, Schwartz J, Cohen S, Seeman T. Diurnal cortisol decline is related to coronary calcification: CARDIA study. Psychosom Med. 2006;68:657–61. doi: 10.1097/01.psy.0000244071.42939.0e. [DOI] [PubMed] [Google Scholar]
- 11.Sephton SE, Sapolsky RM, Kraemer HC, Spiegel D. Diurnal cortisol rhythm as a predictor of breast cancer survival. J Natl Cancer Inst. 2000;92:994–1000. doi: 10.1093/jnci/92.12.994. [DOI] [PubMed] [Google Scholar]
- 12.Kumari M, Shipley M, Stafford M, Kivimaki M. Association of diurnal patterns in salivary cortisol with all-cause and cardiovascular mortality: findings from the Whitehall II study. J Clin Endocrinol Metab. 2011;96:1478–85. doi: 10.1210/jc.2010-2137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rosmond R, Wallerius S, Wanger P, Martin L, Holm G, Björntorp P. A 5-year follow-up study of disease incidence in men with an abnormal hormone pattern. Journal of internal medicine. 2003;254:386–90. doi: 10.1046/j.1365-2796.2003.01205.x. [DOI] [PubMed] [Google Scholar]
- 14.Adam EK, Kumari M. Assessing salivary cortisol in large-scale, epidemiological research. Psychoneuroendocrinology. 2009;34:1423–36. doi: 10.1016/j.psyneuen.2009.06.011. [DOI] [PubMed] [Google Scholar]
- 15.Folkman S. The case for positive emotions in the stress process. Anxiety, Stress & Coping. 2008;21:3–14. doi: 10.1080/10615800701740457. [DOI] [PubMed] [Google Scholar]
- 16.Chida Y, Steptoe A. Positive psychological well-being and mortality: A quantitative review of prospective observational studies. Psychosom Med. 2008;70:741–56. doi: 10.1097/PSY.0b013e31818105ba. [DOI] [PubMed] [Google Scholar]
- 17.Steptoe A, Wardle J. Positive affect and biological function in everyday life. Neurobiol Aging. 2005;26:108–12. doi: 10.1016/j.neurobiolaging.2005.08.016. [DOI] [PubMed] [Google Scholar]
- 18.Steptoe A, O'Donnell K, Badrick E, Kumari M, Marmot M. Neuroendocrine and inflammatory factors associated with positive affect in healthy men and women. Am J Epidemiol. 2008;167:96. doi: 10.1093/aje/kwm252. [DOI] [PubMed] [Google Scholar]
- 19.Polk D, Cohen S, Doyle W, Skoner D, Kirschbaum C. State and trait affect as predictors of salivary cortisol in healthy adults. Psychoneuroendocrinology. 2005;30:261–72. doi: 10.1016/j.psyneuen.2004.08.004. [DOI] [PubMed] [Google Scholar]
- 20.Jacobs N, Myin-Germeys I, Derom C, Delespaul P, van Os J, Nicolson N. A momentary assessment study of the relationship between affective and adrenocortical stress responses in daily life. Biol Psychol. 2007;74:60–6. doi: 10.1016/j.biopsycho.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 21.Lai JC, Evans PD, Ng SH, Chong AM, Siu OT, Chan CL, Ho SM, Ho RT, Chan P, Chan CC. Optimism, positive affectivity, and salivary cortisol. Br J Health Psychol. 2005;10:467–84. doi: 10.1348/135910705X26083. [DOI] [PubMed] [Google Scholar]
- 22.Ryff CD, Singer BH, Love GD. Positive health: connecting well-being with biology. Philosophical Transactions of the Royal Society B: Biological Sciences. 2004;359:1383–94. doi: 10.1098/rstb.2004.1521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Moskowitz J, Epel E. Benefit finding and diurnal cortisol slope in maternal caregivers: A moderating role for positive emotion. The Journal of Positive Psychology. 2006;1:83–91. [Google Scholar]
- 24.Smyth JM, Ockenfels MC, Gorin AA, Catley D, Porter LS, Kirschbaum C, Hellhammer DH, Stone AA. Individual differences in the diurnal cycle of cortisol. Psychoneuroendocrinology. 1997;22:89–105. doi: 10.1016/s0306-4530(96)00039-x. [DOI] [PubMed] [Google Scholar]
- 25.Steptoe A, Gibson E, Hamer M, Wardle J. Neuroendocrine and cardiovascular correlates of positive affect measured by ecological momentary assessment and by questionnaire. Psychoneuroendocrinology. 2007;32:56–64. doi: 10.1016/j.psyneuen.2006.10.001. [DOI] [PubMed] [Google Scholar]
- 26.Russell JA. A circumplex model of affect. J Pers Soc Psychol. 1980;39:1161–78. [Google Scholar]
- 27.Watson D, Tellegen A. Toward a consensual structure of mood. Psychol Bull. 1985;98:219–35. doi: 10.1037//0033-2909.98.2.219. [DOI] [PubMed] [Google Scholar]
- 28.Dahl RE, Hariri AR. Lessons from G. Stanley Hall: Connecting new research in biological sciences to the study of adolescent development. J Res Adolesc. 2005;15:367–82. [Google Scholar]
- 29.Spear LP. Rewards, aversions and affect in adolescence: Emerging convergences across laboratory animal and human data. Dev Cogn Neurosci. 2011;1:390. doi: 10.1016/j.dcn.2011.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Casey B, Jones RM, Hare TA. The adolescent brain. Ann N Y Acad Sci. 2008;1124:111–26. doi: 10.1196/annals.1440.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Casey B, Jones RM, Levita L, Libby V, Pattwell SS, Ruberry EJ, Soliman F, Somerville LH. The storm and stress of adolescence: Insights from human imaging and mouse genetics. Dev Psychobiol. 2010;52:225–35. doi: 10.1002/dev.20447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hankin BL, Abramson LY, Moffitt TE, Silva PA, McGee R, Angell KE. Development of depression from preadolescence to young adulthood: Emerging gender differences in a 10-year longitudinal study. J Abnorm Psychol. 1998;107:128–40. doi: 10.1037//0021-843x.107.1.128. [DOI] [PubMed] [Google Scholar]
- 33.Lewinsohn PM, Rohde P, Seeley JR. Major depressive disorder in older adolescents: prevalence, risk factors, and clinical implications. Clin Psychol Rev. 1998;18:765–94. doi: 10.1016/s0272-7358(98)00010-5. [DOI] [PubMed] [Google Scholar]
- 34.Romeo RD. Adolescence: a central event in shaping stress reactivity. Dev Psychobiol. 2010;52:244–53. doi: 10.1002/dev.20437. [DOI] [PubMed] [Google Scholar]
- 35.Lerner RM, Dowling EM, Anderson PM. Positive youth development: Thriving as the basis of personhood and civil society. Appl Dev Sci. 2003;7:172–80. doi: 10.1002/yd.14. [DOI] [PubMed] [Google Scholar]
- 36.Arnett JJ. Adolescent storm and stress, reconsidered. Am Psychol. 1999;54:317–26. doi: 10.1037//0003-066x.54.5.317. [DOI] [PubMed] [Google Scholar]
- 37.Zinbarg RE, Mineka S, Craske MG, Griffith JW, Sutton J, Rose RD, Nazarian M, Mor N, Waters AM. The Northwestern-UCLA youth emotion project: Associations of cognitive vulnerabilities, neuroticism and gender with past diagnoses of emotional disorders in adolescents. Behav Res Ther. 2010;48:347–58. doi: 10.1016/j.brat.2009.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Adam EK, Vrshek-Schallhorn S, Kendall AD, Mineka S, Zinbarg RE, Craske MG. Prospective associations between the cortisol awakening response and first onsets of anxiety disorders over a six-year follow-up. Psychoneuroendocrinology. 2014;44:47–59. doi: 10.1016/j.psyneuen.2014.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Adam EK, Doane LD, Zinbarg RE, Mineka S, Craske MG, Griffith JW. Prospective prediction of major depressive disorder from cortisol awakening responses in adolescence. Psychoneuroendocrinology. 2010;35:921–31. doi: 10.1016/j.psyneuen.2009.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Doane LD, Mineka S, Zinbarg RE, Craske M, Griffith JW, Adam EK. Are flatter diurnal cortisol rhythms associated with major depression and anxiety disorders in late adolescence? The role of life stress and daily negative emotion Dev Psychopathol. 2013;25:629–42. doi: 10.1017/S0954579413000060. [DOI] [PubMed] [Google Scholar]
- 41.DeSantis AS, Adam EK, Doane LD, Mineka S, Zinbarg RE, Craske MG. Racial/ethnic differences in cortisol diurnal rhythms in a community sample of adolescents. J Adolesc Health. 2007;41:3–13. doi: 10.1016/j.jadohealth.2007.03.006. [DOI] [PubMed] [Google Scholar]
- 42.Vrshek-Schallhorn S, Doane L, Mineka S, Zinbarg R, Craske M, Adam E. The cortisol awakening response predicts major depression: predictive stability over a 4-year follow-up and effect of depression history. Psychol Med. 2013;43:483–93. doi: 10.1017/S0033291712001213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Eysenck H, Eysenck S. Manual of the Eysenck Personality Questionnaire (adult and junior) Longon: Hodder & Stoughton; 1975. [Google Scholar]
- 44.Clark LA, Watson D, Mineka S. Temperament, personality, and the mood and anxiety disorders. J Abnorm Psychol. 1994;103:103. [PubMed] [Google Scholar]
- 45.Klein DN, Durbin CE, Shankman SA. Personality and mood disorders. In: Gotlib IH, Hammen CL, editors. Handbook of depression. 2 ed. New York: Guilford Press; 2009. pp. 93–112. [Google Scholar]
- 46.Costa P, Jr, Terracciano A, McCrae RR. Gender differences in personality traits across cultures: robust and surprising findings. J Pers Soc Psychol. 2001;81:322. doi: 10.1037/0022-3514.81.2.322. [DOI] [PubMed] [Google Scholar]
- 47.First MB, Spitzer RL, Gibbon M, Williams JBW. Structured clinical interview for DSM-IV-TR Axis I disorders, research version, non-patient edition (SCID-I/NP) New York: Biometrics Research, New York State Psychiatric Institute; 2002. [Google Scholar]
- 48.Csikszentmihalyi M, Larson R. Validity and reliability of the experience-sampling method. J Nerv Ment Dis. 1987;175:526. doi: 10.1097/00005053-198709000-00004. [DOI] [PubMed] [Google Scholar]
- 49.Dressendörfer RA, Kirschbaum C, Rohde W, Stahl F, Strasburger CJ. Synthesis of a cortisol-biotin conjugate and evaluation as a tracer in an immunoassay for salivary cortisol measurement. J Steroid Biochem Mol Biol. 1992;43:683–92. doi: 10.1016/0960-0760(92)90294-s. [DOI] [PubMed] [Google Scholar]
- 50.Schwarz N. Self-reports: How the questions shape the answers. Am Psychol. 1999;54:93. [Google Scholar]
- 51.Posner J, Russell JA, Peterson BS. The circumplex model of affect: An integrative approach to affective neuroscience, cognitive development, and psychopathology. Dev Psychopathol. 2005;17:715–34. doi: 10.1017/S0954579405050340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zeiders KH, Doane LD, Adam EK. Reciprocal relations between objectively measured sleep patterns and diurnal cortisol rhythms in late adolescence. J Adolesc Health. 2011;48:566–71. doi: 10.1016/j.jadohealth.2010.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kirschbaum C, Kudielka BM, Gaab J, Schommer NC, Hellhammer DH. Impact of gender, menstrual cycle phase, and oral contraceptives on the activity of the hypothalamus-pituitary-adrenal axis. Psychosom Med. 1999;61:154–62. doi: 10.1097/00006842-199903000-00006. [DOI] [PubMed] [Google Scholar]
- 54.Cohen S, Schwartz JE, Epel E, Kirschbaum C, Sidney S, Seeman T. Socioeconomic status, race, and diurnal cortisol decline in the Coronary Artery Risk Development in Young Adults (CARDIA) Study. Psychosom Med. 2006;68:41–50. doi: 10.1097/01.psy.0000195967.51768.ea. [DOI] [PubMed] [Google Scholar]
- 55.Badrick E, Kirschbaum C, Kumari M. The relationship between smoking status and cortisol secretion. J Clin Endocrinol Metab. 2007;92:819–24. doi: 10.1210/jc.2006-2155. [DOI] [PubMed] [Google Scholar]
- 56.Rohleder N, Kirschbaum C. The hypothalamic–pituitary–adrenal (HPA) axis in habitual smokers. Int J Psychophysiol. 2006;59:236–43. doi: 10.1016/j.ijpsycho.2005.10.012. [DOI] [PubMed] [Google Scholar]
- 57.Adam EK, Gunnar MR. Relationship functioning and home and work demands predict individual differences in diurnal cortisol patterns in women. Psychoneuroendocrinology. 2001;26:189–208. doi: 10.1016/s0306-4530(00)00045-7. [DOI] [PubMed] [Google Scholar]
- 58.Wooldridge J. Introductory econometrics: A modern approach. Cincinnati, OH: South-Western College Publishing; 2003. [Google Scholar]
- 59.Mattsson C, Reynolds RM, Simonyte K, Olsson T, Walker BR. Combined receptor antagonist stimulation of the HPA axis test identifies impaired negative feedback sensitivity to cortisol in obese men. J Clin Endocrinol Metab. 2009;94:1347–52. doi: 10.1210/jc.2008-2054. [DOI] [PubMed] [Google Scholar]
- 60.Gunnar MR, Vazquez DM. Low cortisol and a flattening of expected daytime rhythm: potential indices of risk in human development. Dev Psychopathol. 2001;13:515–38. doi: 10.1017/s0954579401003066. [DOI] [PubMed] [Google Scholar]
- 61.Huppert FA. Positive emotions and cognition: Developmental, neuroscience and health perspectives. In: Forgas JP, editor. Affect in Social Thinking and Behavior. New York, NY: Psychology Press; 2006. pp. 235–52. [Google Scholar]
- 62.Buckley TM, Schatzberg AF. On the interactions of the hypothalamic-pituitary-adrenal (HPA) axis and sleep: normal HPA axis activity and circadian rhythm, exemplary sleep disorders. J Clin Endocrinol Metab. 2005;90:3106–14. doi: 10.1210/jc.2004-1056. [DOI] [PubMed] [Google Scholar]
- 63.Leproult R, Copinschi G, Buxton O, Van Cauter E. Sleep loss results in an elevation of cortisol levels the next evening. Sleep. 1997;10:856–70. [PubMed] [Google Scholar]
- 64.Kumari M, Badrick E, Ferrie J, Perski A, Marmot M, Chandola T. Self-reported sleep duration and sleep disturbance are independently associated with cortisol secretion in the Whitehall II study. J Clin Endocrinol Metab. 2009;94:4801–9. doi: 10.1210/jc.2009-0555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Pfeifer JH, Allen NB. Arrested development? Reconsidering dual-systems models of brain function in adolescence and disorders. Trends Cogn Sci. 2012;16:322–9. doi: 10.1016/j.tics.2012.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Gerritsen L, Geerlings MI, Bremmer MA, Beekman AT, Deeg DJ, Penninx BW, Comijs HC. Personality characteristics and hypothalamic-pituitary-adrenal axis regulation in older persons. The American Journal of Geriatric Psychiatry. 2009;17:1077–84. doi: 10.1097/JGP.0b013e3181bd1be6. [DOI] [PubMed] [Google Scholar]
- 67.Nater UM, Hoppmann C, Klumb PL. Neuroticism and conscientiousness are associated with cortisol diurnal profiles in adults—role of positive and negative affect. Psychoneuroendocrinology. 2010;35:1573–7. doi: 10.1016/j.psyneuen.2010.02.017. [DOI] [PubMed] [Google Scholar]
- 68.Kirschbaum C, Bartussek D, Strasburger C. Cortisol responses to psychological stress and correlations with personality traits. Pers Individ Dif. 1992;13:1353–7. [Google Scholar]
- 69.Schommer N, Kudielka B, Hellhammer D, Kirschbaum C. No evidence for a close relationship between personality traits and circadian cortisol rhythm or a single cortisol stress response. Psychol Rep. 1999;84:840–2. doi: 10.2466/pr0.1999.84.3.840. [DOI] [PubMed] [Google Scholar]
- 70.Adam EK. Momentary emotion and cortisol levels in the everyday lives of working parents. In: Schneider B, Waite L, editors. Being together, working apart: Dual career families and the work-life balance. Cambridge: Cambridge University Press; 2005. pp. 105–34. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.