

Abstract
Despite the intensive research in the past decade on the microbial bioaccumulation of heavy metals, the significance of redox state for oxidative stress induction is not completely clarified. In the present study, we examined the effect of redox-active (copper and chromium) and redox-inactive (cadmium) metals on the changes in levels of oxidative stress biomarkers and antioxidant enzyme defence in Trichosporon cutaneum R57 cells. This filamentous yeast strain showed significant tolerance and bioaccumulation capability of heavy metals. Our findings indicated that the treatment by both redox-active and redox-inactive heavy metal induced oxidative stress events. Enhanced concentrations of Cu2+, Cr6+ and Cd2+ caused acceleration in the production of reactive oxygen species (ROS), increase in the level of oxidatively damaged proteins and accumulation of reserve carbohydrates (glycogen and trehalose). Cell response against heavy metal exposure also includes elevation in the activities of antioxidant enzymes, superoxide dismutase and catalase, which are key enzymes for directly scavenging of ROS. Despite the mentioned changes in the stress biomarkers, T. cutaneum did not show a significant growth diminution. Probably, activated antioxidant defence contributes to the yeast survival under conditions of heavy metal stress.
Keywords: filamentous yeast, oxidative stress, heavy metals, antioxidant defence, biomarkers of oxidative stress
Introduction
Metal ions are considered very important and at the same time very toxic for living organisms. They are some of the main pollutants in the environment. Heavy metals are present in soils as free or exchangeable metal ions, soluble metal complexes, organically bound metals, precipitated or insoluble compounds (oxides, carbonates and hydroxides).[1] Recently, microbial systems, like fungi, bacteria and algae, have been successfully used as adsorbing agents for removal of heavy metals.[2,3] Different species of Aspergillus, Pseudomonas, Sporophyticus, Bacillus, Phanerochaete, etc. have been reported as efficient reducers.[4] The response of microorganisms towards toxic heavy metals is critically important in the reclamation of polluted sites. Living organisms exposed environmentally to high metal concentrations follow various mechanisms to counter potential toxicity. Among the group of microorganisms used for bioremediation, yeasts are having a leading place.[5]
A possible consequence of heavy metal exposure is an increased production of reactive oxygen species (ROS) such as hydroxyl radical (HO·), superoxide radical (•O2 −) or hydrogen peroxide (H2O2) that could induce or exacerbate intracellular oxidative stress. These ROS may lead to the unspecific oxidation of proteins and membrane lipids or may cause DNA damage.[6] Defence mechanisms which counteract the impact of ROS, including enzyme and non-enzyme antioxidant systems, are found in all aerobic cells. Interaction between chemical elements, the level of oxidative stress and antioxidant defence play an important role in ecotoxicological response of microorganisms in polluted environments.[7]
The high potential of Trichosporon cutaneum strain R57 for heavy metal removal from contaminated waste water has been demonstrated in several previous investigations, showing significant tolerance and bioaccumulation capability for chromium (Cr), cadmium (Cd) and copper (Cu).[8,9] Our previous study has also shown high ability of the same yeast strain to grow in the media supplemented with high content of phenols and resistant to toxic chemicals, such as benzyl alcohol,[10,11] revealed that the capacity of the strain to sustain toxic concentrations of heavy metals in the medium often refers to its ability to accumulate harmful ions in the cells.[12] The question arises regarding the relationship between redox state of the metals and the oxidative stress induction in the yeast cells. Copper and chromium as redox-active metals generate ROS through redox cycling reactions.[13] Redox-inactive Cd impairs antioxidant defences, especially those involving thiol-containing antioxidants and enzymes.
Copper has a dual role in terms of regulating the life processes in the living organisms. As an essential trace element copper acts as a cofactor in multiple enzymes, including superoxide dismutase (SOD), ceruloplasmin, Cu monooxygenases, cytochrome c oxidase, etc. At the same time, copper is toxic to microorganisms and may lead to their death even within minutes of their exposure to copper.[14] The toxic effect may involve inhibition of growth,[15,16] substitution of essential ions and blocking of functional groups on proteins,[17] inactivation of enzymes,[18,19] disturbances of the metabolism,[20] alterations of membrane integrity and production of ROS.[16,21,22]
Chromium is a relatively abundant element in the Earth's crust. It represents an essential micronutrient for living organisms as a participant in the maintenance of normal carbohydrate metabolism in mammals and yeasts.[23] Moreover, it has also been suggested that Cr(III) is involved in the tertiary structure of proteins and in the conformation of cellular RNA and DNA.[24] At the same time, chromium pollution caused serious problems in many regions of the world. It is a transition metal, water soluble, enters living cells easily and is toxic and carcinogenic. Trivalent (Cr[III]) and hexavalent (Cr[VI]) compounds are thought to be the most biologically significant.[25] Inside living cells, the redox cycling of chromium species in different oxidation states generates ROS via the Fenton and Haber–Weiss reactions.[26]
Cadmium is a prevalent non-essential, redox-inactive, highly toxic metal. It is an important heavy metal pollutant. Cadmium shows high affinity towards functional groups of biomolecules, i.e., amino, carboxyl, phosphate and thiol groups,[27] and interferes with numerous biochemical and physiological processes like photosynthesis, respiration, plant–water relationships, nitrogen and protein metabolism, and nutrient uptake.[28] There are some evidences that cadmium-induced oxidative stress in Saccharomyces cerevisiae, since strains deficient in antioxidant defence enzymes have a high sensitivity to cadmium and cells grown in the absence of oxygen are more tolerant to cadmium.[29] The bacterial response to cadmium includes induction of expression of genes in many regulons, including genes involved in metal transport, DNA repair, the heat shock response and the oxidative stress response (see [29]).
Despite the great interest in the microbial metal bioaccumulation, the mechanism of oxidative stress induction by heavy metals with different redox status, particularly redox-inactive metals, is not fully understood.
Therefore, the aim of present paper was to evaluate and compare the effect of redox-active (copper and chromium ions) and redox-inactive (Cd) metals on induction of oxidative stress events in T. cutaneum R57 cells. To do this, we determined the growth, ROS production and oxidative damaged protein content in yeast cells exposed to different concentrations of metal ions for 6 h. In addition, the role of antioxidant enzymes SOD and catalase (CAT) in the cell response was investigated.
Materials and methods
Yeast strain and culture conditions
The T. cutaneum R57 strain was obtained from National Bank of Industrial Microbial and Cell Cultures, Bulgaria. The basidiomycete yeast strain of T. cutaneum R57 has been registered under N2414.[30]
The cultivation was performed as follows: 80 mL of seed medium was inoculated with 5 mL preculture at a concentration of 2 × 108 CFU/mL in 500 mL Erlenmeyer flasks, on a shaker (220 rpm) at 28 °C for 24 h. Then different concentrations of CdSO4 (1, 5 and 10 mmol/L), K2Cr2O7 (1, 5 and 10 mmol/L) and CuSO4.5H2O (0.5, 1 and 3 mmol/L) ions were added to the culture medium and cultivation continued for the next 6 h. These concentrations were chosen because they allow us to obtain enough biomass for bioaccumulation experiments.
Cell-free extract preparation
The cell-free extract was prepared as described earlier.[31] All steps were performed at 0–4 °C.
Enzyme activity determination
SOD activity was measured in cell-free extract by the nitro-blue tetrazolium (NBT) reduction method.[32] One unit of SOD activity was defined as the amount of SOD required for inhibition of the reduction of NBT by 50% (A560) and was expressed as units per mg protein (U/mg protein). Catalase was assayed by the method of Beers and Sizer,[33] in which the decomposition of H2O2 was analysed spectrophotometrically at 240 nm. One unit of catalase activity was defined as the amount of enzyme that decomposes 1 mmol H2O2 min−1 at an initial H2O2 concentration of 30 mmol/L at pH 7.0 and 25 °C. The specific activity is given as U/mg protein.
Determination of ROS
For measurement of •O2 − production rate, the method of SOD inhabitable reduction of cytochrome c was used.[34] A molar extinction coefficient of 2.11 × 104 was used to calculate the concentration of reduced cytochrome c.
For measurement of hydrogen peroxide production, the method of Pick and Mizel [35] was used. For calculations, a standard curve with H2O2 concentrations (from 5 to 50 mmol/L) was used.
Measurement of protein carbonyl content
Protein oxidative damage was measured spectrophotometrically as protein carbonyl content using the 2,4-dinitrophenylhydrazine (DNPH) binding assay,[36] slightly modified by Adachi and Ishii.[37] Following metal treatment, the cell-free extracts were incubated with DNPH for 1 h at 37 °C; proteins were precipitated in 10% cold trichloroacetic acid and washed with ethanol:ethylacetate (1:1), to remove excess DNPH and finally dissolved in 6 mol/L guanidine chloride, pH 2. The optimal density was measured at 380 nm, and the carbonyl content was calculated using a molar extinction coefficient of 21 (mmol/L)−1 cm−1, resulting in final measurement of nanomoles of DNPH incorporated (protein carbonyls) per mg of protein.
Determination of reserve carbohydrates
In order to determine glycogen and trehalose content, a procedure previously described by Becker [38] and Vandercammen et al. [39] and then modified by Parrou and Francois [40] was used. Soluble reducing sugars were determined by the Somogyi–Nelson method.[41]
Other analytical methods
Protein was estimated by the Lowry procedure [42] using crystalline bovine albumin as a standard.
Microbial growth was monitored by measuring the dry weight using Electronic Moisture Balance (KERN, Germany).
Results and discussion
Effect of metal ions on growth
The effects of heavy metals on yeast cell growth mainly depend on the mechanisms of metabolic or passive uptake of toxic ions into the cells. The highest concentrations of the metals used are chosen in our preliminary investigations as limiting for the organism survival (data not shown).
Growth of T. cutaneum R57 was studied in relation to 0–3 mmol/L CuSO4.5H2O exposure under submerged conditions (Figure 1(A)). Presence of copper ions in cultural medium induced yeast growth. Mycelia weight increased with rising of metal concentrations. The highest biomass production was detected at a concentration of 3 mmol/L (167% compared with the control).
Figure 1.
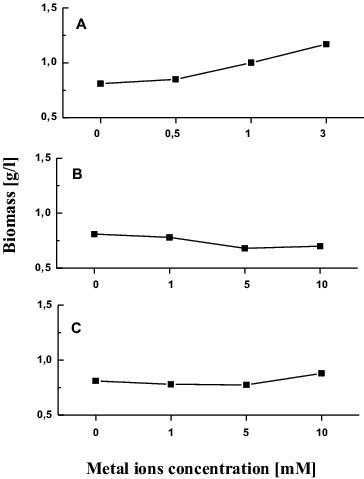
Growth of T. cutaneum R57 cells in the presence of different concentrations of metal ions (A) Cu, (B) Cr, and (C) Cd.
As shown in Figure 1(B), chromium concentrations of 5 and 10 mmol/L led to 16% less biomass production than the control. Slight reduction of the dry weight at 1 and 5 mmol/L was observed as a result of yeast cell treatment with cadmium ions. Exposure to the next concentration used (10 mmol/L Cd) caused only a minor increase (8%) in biomass yield compared to the control (Figure 1(C)).
Effect of metal stress on ROS generation
In the present experiments, all heavy metal concentrations applied to yeast culture induced oxidative stress events. It should be noted that the presence of copper, chromium or cadmium ions drastically changed ROS level in T. cutaneum R57 cells. Table 1 shows the effect of copper, chromium and cadmium ions on ·O2 − and H2O2 production in T. cutaneum R57 cells after 6 h of exposure to the metal ions. Elevation in ·O2 − content was found for all metal concentrations tested.
Table 1.
Increase in ROS generation in the intact cells of T. cutaneum R57 treated by enhanced concentrations of Cu, Cr and Cd ions.
| Cu |
Cr |
Cd |
||||
|---|---|---|---|---|---|---|
| Variants | ·O2− (nmol /mg d.w./h) | H2O2 (μmol /L.mg d.w./h) | ·O2− (nmol/mg d.w./h) | H2O2 (μmol/L.mg d.w./h) | ·O2− (nmol/mg d.w./h) | H2O2 (μmol/L.mg d.w./h) |
| Control | 0.63 | 13.1 | 0.60 | 15.20 | 0.69 | 14.00 |
| 0.5 mmol/L | 0.65 | 13.1 | Nd | Nd | Nd | Nd |
| 1 mmol/L | 0.89 | 22.4 | 0.90 | 33.9 | 0.86 | 20.80 |
| 3 mmol/L | 1.15 | 21.6 | Nd | Nd | Nd | Nd |
| 5 mmol/L | Nd | Nd | 1.58 | 37.5 | 1.20 | 22.00 |
| 10 mmol/L | Nd | Nd | 1.80 | 28.6 | 2.41 | 25.6 |
As is apparent from Table 1, the treatment by copper ion concentrations in the range from 0.5 to 3 mmol/L resulted in gradual increase in ·O2 − level above 0.5 mmol/L. The maximum response was achieved with 3 mmol/L (182% compared with the control).
Short-term exposure to 1 and 5 mmol/L chromium ions increased the ·O2 − level by about 1.5 and 2.7-fold, respectively, as compared to the control. The cells treated with 10 mmol/L showed extremely high ·O2 − levels (threefold higher than controls).
Cadmium ions had a similar effect on ·O2 − production. The level of ·O2 − increased depending on the metal concentrations. Maximum superoxide production was observed by the treatment with 10 mmol/L cadmium ions (349% in comparison with the control).
Metal treatment also caused an increase in H2O2 levels in concentration-dependent manner. A significant induction in H2O2 production was observed after 6 h of incubation with copper ions (1.6- and 1.7-fold higher in comparison with the control at concentrations of 1 and 3 mmol/L, respectively). Exposure to 1, 5 and 10 mmol/L chromium ions led to a significant increase in H2O2 levels compared to the control (2.2-, 2.5- and 1.9-fold, respectively). The same trend was shown for the Cd ions – 1.4-, 1.6- and 1.8-fold increase after exposure to 1, 5 and 10 mmol/L, respectively.
The current experiments indicated that metal treatment significantly increased production of ·O2 − and H2O2. This increase did not depend on the type of metals (redox-active or redox-inactive) but depended on their concentrations. Even a small increase of metal concentration in the cell led to ROS generation. Redox-active metals, such as copper and chromium, are prone to participate in the formation of ROS via a Fenton-like reaction.[25] Similar direct analyses of ROS content in microbial cells have not often been reported. Transition metal copper is one of the most potent elements catalysing Fenton's reaction. Copper treatment induced ROS generation in yeasts S. cerevisiae [43] and fungi Podospora anserina,[44] Humicola lutea [16] and aquatic hyphomycetes Varicosporium elodeae and Heliscus submersus.[45] Similar results have been reported for several plants [46] and human cells.[47,48] Chromium ion exposure also induced ROS generation in yeasts, fungi [49] and plants.[50] The redox-inactive metal cadmium affected ROS production in the model strain T. cutaneum R57 in the same way as copper and chromium. Cadmium, as the least representative member of the transition element group, does not induce production of ROS though a Fenton-like redox cycling mechanism, as is the case with copper and chromium.[51] However, this metal inhibits mainly complexes II and III of the electron transport chain and this inhibition induces ROS generation in the mitochondria. We found that cadmium induces ·O2 − and H2O2 about 3.5- and 1.9-fold, respectively, as compared to the control. This supports previous studies that suggested possible role of elevated ROS production in mediating Cd toxicity to bacteria,[52] yeast [53] and plants.[54]
Metal exposure caused protein oxidation
As a consequence of excessive ROS production, oxidative damages of proteins occurred.[55] Protein carbonylation, one of the most harmful irreversible oxidative protein modifications, is used as a biomarker of metal-induced oxidative stress. Exposure to elevated copper concentrations did not significantly affect carbonyl content with exception of 1 mmol/L when the carbonylated protein showed 42% higher level than in the control (Figure 2(A)).
Figure 2.
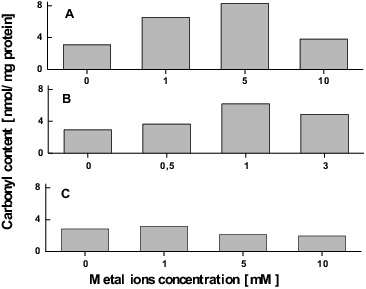
Protein oxidation induced in T. cutaneum R57 cells by Cu (A), Cr (B) and Cd (C) ions.
In contrast, treatment with enhanced chromium ion concentrations up to 5 mmol/L caused significant increase in the protein carbonyl content (about 2.3-fold compared with the control). It should be noted that the subsequent concentration (10 mmol/L) resulted in a sharp decrease in the amount of oxidative damaged protein (Figure 2(B)).
When the T. cutaneum R57 cells were treated with cadmium ions, the trends of protein carbonyl content showed a similar profile as after treatment with copper ions, and the highest level of damaged protein was found at 1 mmol/L concentration (Figure 2(C)).
The results mentioned above indicate that exposing T. cutaneum R57 to elevated concentrations of copper, chromium and cadmium ions inflicts oxidative damage on the intracellular proteins. It is worth noting that the redox-active metal ions (copper and chromium ions) demonstrated more drastic changes in oxidative damaged protein content. The majority of proteins that were oxidized over the course of the investigation demonstrated an increase in their relative abundance after exposure to 1 mmol/L CuSO4.5H2O, 5 mmol/L K2Cr2O7 and 1 mmol/L CdSO4. The subsequent decline in abundance after treatment with higher concentrations could be explained by enhancing both degradation of proteins by proteases and aggregation of heavily oxidized proteins (see [16]). Evidence for a positive correlation between increased levels of ROS and damaged proteins has been published for various microbial cells.[16]
Effect of stress on glycogen and trehalose content
The metal exposure of T. cutaneum R57 was accompanied by quantified changes in the reserve carbohydrates such as trehalose and glycogen. Level of reserve carbohydrates was influenced by all the metal ions added (Figure 3). We observed reduction of trehalose amount after 6 h of metal treatment. This reduction was the most significant in the variant treated with 10 mmol/L Cr (65% lower than the control) (Figure 3(B)).
Figure 3.
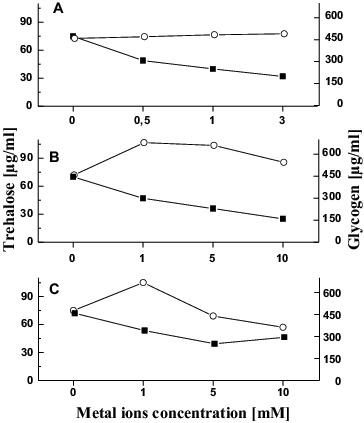
Changes in the glycogen (-o-) and trehalose (-▪-) level in T. cutaneum R57 cells treated with Cu (A), Cr (B) and Cd (C) ions.
The other carbohydrate glycogen showed a different behaviour. The glycogen amount in T. cutaneum R57 was not influenced after 6 h of copper treatment (Figure 3(A)).
We observed an increase in the level of glycogen in the other variants tested. Experiments with chromium ions showed a trend of enhancing this carbohydrate (above 48%) in variants with 1 and 5 mmol/L and then a slight decrease similar to the control level (Figure 3(B)). A similar trend is observed in variants with cadmium ions. After a 40% increase in glycogen level, a sharp decrease was observed at 10mmol/L Cd (Figure 3(C)).
As was mentioned above, the metal exposure of T. cutaneum R57 results in changes of the reserve carbohydrates such as trehalose and glycogen. Both compounds have different physiological effects and mode of action: trehalose might be a more general stress protectant and assists chaperones in controlling protein denaturation and renaturation, and glycogen is a storage carbohydrate.[56,57] Our results indicated that chromium and cadmium ion treatment led to a significant increase in glycogen content. These findings agree with earlier studies about the microbial response against heavy metal stress including chromium and cadmium ions.[21] In contrast, no changes were found in the amounts of glycogen in the cells treated with copper ions. Similar data has been reported by stress response in yeasts.[58] Furthermore, Arapaima gigas (a carnivorous fish) and the fry of common carp demonstrated a decrease in glycogen level under conditions of copper stress.[59,60] The possible explanation is a progressive glycogenolysis (breakdown of glycogen (n) to glucose-1-phosphate and glycogen (n − 1)) in the cells.[61]
In addition, the trehalose amount gradually decreased with increase in the metal ion concentration. These results could be due to the active metabolism of trehalose, its simultaneous synthesis and degradation during the period of stress.[62] Probably, after 6 h of exposure to metal stress neutral trehalase (enzyme responsible for trehalose degradation) exhibits higher activity than trehalose 6-phosphate phosphatase (enzyme responsible for its synthesis).
Metal effect on SOD and CAT activity
Fungi, like all aerobic organisms, have a set of defence mechanisms to deal with oxidative stress.[63] The mechanism of metal-induced formation of ROS is strongly modulated by the action of cellular antioxidants. The presence of metal ions stimulated enzyme antioxidant defence in T. cutaneum R57 cells. The results shown in Figure 4(A) demonstrate a gradual elevation in SOD activity after exposure to enhanced concentrations of copper ions. The maximum activity was observed at the concentration of 3 mmol/L when about 1.5-fold higher specific enzyme activity was achieved. While the treatment of yeast cells with 1 or 5 mmol/L chromium caused an insignificant increase in the level of SOD (19% compared to the control), the next concentration (10 mmol/L) resulted in a reduction of 22% compared with the highest enzyme activity (Figure 4(B)). The activity of SOD raised dose-dependent manner in a variant of cadmium ions treatment. The highest enzyme activity was observed at the concentration of 10 mmol/L (Figure 4(C)).
Figure 4.
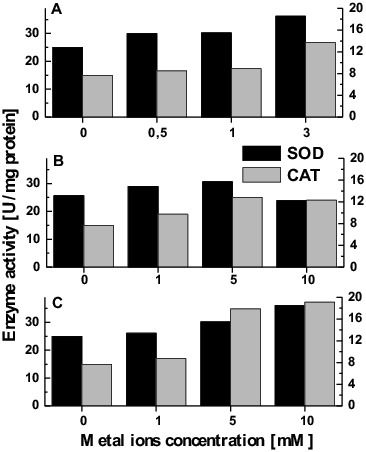
Antioxidant response of T. cutaneum R57 against metal-induced stress (A) Cu, (B) Cr, and (C) Cd.
As shown in Figure 4, the effect of metal treatment was more pronounced for CAT than for SOD. All of the metal ions tested increased CAT activity in a concentration-dependent manner. The maximum enzyme activity was 1.8-, 2.5- and 1.6-fold higher for copper, chromium and cadmium ions, respectively, than the activity in the corresponding control variant.
Enzymes, such as SOD and CAT, have been reported to be activated as a result of elevated ROS levels in several organisms exposed to heavy metal stress.[16,64] These enzymes are crucial for cellular detoxification, controlling the levels of superoxide anion radical and hydrogen peroxide.[63] Higher activities of SOD and CAT are associated with the induced resistance of the mycetes to different stress factors.[65,66] Our result indicated that all the metals used induced activation of the enzymes involved in antioxidant defence. Treatment of yeast cells with copper, chromium and cadmium ions showed a clear tendency of concentration-dependent stimulation of SOD and CAT activity.
Conclusion
In summary, our results provide additional confirmation for metal-mediated oxidative stress in filamentous yeasts. Both redox-active and non-redox-active metals caused oxidative stress events which included enhanced levels of oxidatively damaged proteins, changes of glycogen and trehalose levels, and activation of the antioxidant enzymes SOD and CAT. Despite the significant induction of antioxidant enzyme activity, copper exposure still has deleterious effects, probably mediated by the overloading of antioxidant defences.
Acknowledgements
The authors gratefully acknowledge support under the Bilateral Scientific Cooperation between the Bulgarian Academy of Sciences and the National Academy of Sciences of Ukraine (period 2013–2015).
Funding Statement
This work was supported [grant number BG051PO001-3.3.06-0059], financed by the European Social Fund and Operational Programme Human Resources Development (2007–2013) and co-financed by the Bulgarian Ministry of Education and Science.
References
- Leyval C, Turnau K, Haselwandter K. Effect of heavy metal pollution on mycorrhizal colonization and function: physiological, ecological and applied aspects. Mycorrhiza. 1997;7:139–153. [Google Scholar]
- Munoz R, Alvarez MT, Munoz A, Terrazas E, Guieysse B, Mattisasson B. Sequential removal of heavy metals ions and organic pollutants using an algal-bacterial consortium. Chemosphere. 2006;63:903–991. doi: 10.1016/j.chemosphere.2005.09.062. [DOI] [PubMed] [Google Scholar]
- Munoz R, Guieysse B. Algal-bacterial processes for the treatment of hazardous contaminants: a review. Water Res. 2006;40:2799–2815. doi: 10.1016/j.watres.2006.06.011. [DOI] [PubMed] [Google Scholar]
- Yan G, Viraraghavan T. Heavy-metal removal from aqueous solution by fungus Mucor rouxii. Water Res. 2003;37:4486–4496. doi: 10.1016/S0043-1354(03)00409-3. [DOI] [PubMed] [Google Scholar]
- Vasudevan P, Padmavathy V, Dhingra SC. Biosorption of heavy metal ions. Bioresour Technol. 2003;89:281–287. doi: 10.1016/s0960-8524(03)00067-1. [DOI] [PubMed] [Google Scholar]
- Fridovich I. Oxygen toxicity: a radical explanation. J Exp Biol. 1998;201:1203–1209. doi: 10.1242/jeb.201.8.1203. [DOI] [PubMed] [Google Scholar]
- Kamiński P, Kurhalyuk N, Szady-Grad M. Heavy metal-induced oxidative stress and changes in physiological process of free radicals in the blood of white stork (Ciconia ciconia) chicks in polluted areas. Polish J Environ Stud. 2007;16(4):555–562. [Google Scholar]
- Georgieva N, Peshev D, Rangelova N, Lazarova N. Effect of hexavalent chromium on growth of Trichosporon cutaneum r 57 . J Univ Chem Technol Metallurgy. 2011;46(3):293–298. [Google Scholar]
- Georgieva N. Growth of Trichosporon cutaneum R 57 in the presence of toxic concentration of cadmium and copper. Int J Agric Biol. 2008;10:325–328. [Google Scholar]
- Ivanova N, Yotova L. Biotransformation of furfural by yeast cells covalently bound to cellulose granules. Acta Biotechnol. 1993;13(3):79–82. [Google Scholar]
- Yotova L, Tzibranska I, Tileva F, Markx G, Georgieva N. Kinetics of the biodegradation of phenol in wastewaters from the chemical industry by covalently immobilized Trichosporon cutaneum cells. J Ind Microbiol Biotechnol. 2009;36(3):367–372. doi: 10.1007/s10295-008-0505-8. [DOI] [PubMed] [Google Scholar]
- Blackwell KJ, Singleton I, Tobin JM. Metal cation uptake by yeast: a review. Appl Microbiol Biotechnol. 1995;43:579–584. doi: 10.1007/BF00164757. [DOI] [PubMed] [Google Scholar]
- Ercal N, Gurer-Orhan H, Aykin-Burns N. Toxic metals and oxidative stress part 1: mechanisms involved in metal-induced oxidative damage. Curr Top Med Chem. 2001;1:529–539. doi: 10.2174/1568026013394831. [DOI] [PubMed] [Google Scholar]
- Espirito Santo C, Taudte N, Nies DH, Grass G. Contribution of copper ion resistance to survival of Esherichia coli on metallic copper surfaces. Appl Environ Microbiol. 2008;74:977–986. doi: 10.1128/AEM.01938-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillén Y, Machuca Á. The effect of copper on the growth of wood-rotting fungi and a blue-stain fungus. World J Microbiol Biotechnol. 2008;24(1):31–37. [Google Scholar]
- Krumova EZ, Pashova SB, Dolashka-Angelova PA, Stefanova T, Angelova MB. Biomarkers of oxidative stress in the fungal strain Humicola lutea under copper exposure. Process Biochem. 2009;44(3):288–295. [Google Scholar]
- Dupont CL, Grass G, Rensing C. Copper toxicity and the origin of bacterial resistance – new insights and applications. Metallomics. 2011;3(11):1109–1118. doi: 10.1039/c1mt00107h. [DOI] [PubMed] [Google Scholar]
- Gokhale NH, Cowan JA. Inactivation of human angiotensin converting enzyme by copper peptide complexes containing ATCUN motifs. Chem Commun (Cambridge) 2005;47(47):5916–5918. doi: 10.1039/b511081e. [DOI] [PubMed] [Google Scholar]
- Wang S, Teng S, Fan M. Interaction between heavy metals and aerobic granular sludge. In: Kumar Sarkar S, editor. Environmental management. Croatia: Sciyo; 2010. pp. 173–188. [Google Scholar]
- Chillappagari S, Seubert A, Trip H, Kuipers OP, Marahiel MA, Miethke M. Copper stress affects iron homeostasis by destabilizing iron–sulfur cluster formation in Bacillus subtilis . J Bacteriol. 2010;192(10):2512–2524. doi: 10.1128/JB.00058-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma SS, Dietz K-J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009;14(1):43–50. doi: 10.1016/j.tplants.2008.10.007. [DOI] [PubMed] [Google Scholar]
- Dávila Costa JS, Albarracín VH, Abate CM. Responses of environmental Amycolatopsis strains to copper stress. Ecotoxicol Environ Saf. 2011;74(7):2020–2028. doi: 10.1016/j.ecoenv.2011.06.017. [DOI] [PubMed] [Google Scholar]
- Debski B, Zalewski W, Gralak MA, Kosala TJ. Chromium–yeast supplementation of chicken broilers in an industrial farming system. Trace Elem Med Biol. 2004;18:47–51. doi: 10.1016/j.jtemb.2004.02.003. [DOI] [PubMed] [Google Scholar]
- Focardi S, Pepi M, Focardi SE. Microbial reduction of hexavalent chromium as a mechanism of detoxification and possible bioremediation applications. In: Chamy R, Rosenkranz F, editors. Agricultural and biological sciences: “Biodegradation – Life of Science”. 2013. pp. 321–347. [Google Scholar]
- Jomova K, Valko M. Advances in metal-induced oxidative stress and human disease. Toxicology. 2011;283:65–87. doi: 10.1016/j.tox.2011.03.001. [DOI] [PubMed] [Google Scholar]
- Lushchak VI. Oxidative stress as a component of transition metal toxicity in fish. In: Svensson EP, editor. Aquatic toxicology research focus. Hauppaug (NY): Nova Science Publishers Inc; 2008. pp. 1–29. [Google Scholar]
- Mendoza-Cozatl DG, Butko E, Springer F, Torpey JW, Komives EA, Kehr J, Schroeder JI. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation. Plant J. 2008;54:249–259. doi: 10.1111/j.1365-313X.2008.03410.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P, Wang WQ, Zhang GL, Kaminek M, Dobrev P, Xu J, Gruissem W. Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J Integr Plant Biol. 2010;52:653–669. doi: 10.1111/j.1744-7909.2010.00956.x. [DOI] [PubMed] [Google Scholar]
- Banjerdkij P, Vattanaviboon P, Mongkolsuk S. Exposure to cadmium elevates expression of genes in the OxyR and OhrR regulons and induces cross-resistance to peroxide killing treatment in Xanthomonas campestris . Appl Environ Microbiol. 2005;71(4):1843–1849. doi: 10.1128/AEM.71.4.1843-1849.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Georgieva N, Alexieva Zl. Selection and characterization of L-ethionine resistant mutants of Trichosporon cutaneum . Z Naturforsch. 2005;60c:657–660. doi: 10.1515/znc-2005-9-1001. [DOI] [PubMed] [Google Scholar]
- Angelova M, Genova L, Slokoska L, Pashova S. Effect of glucose on the superoxide dismutase production in fungal strain Humicola lutea. Can J Microbiol. 1995;41:978–983. doi: 10.1139/m95-136. [DOI] [PubMed] [Google Scholar]
- Beauchamp C, Fridovich I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal Biochem. 1971;44:276–287. doi: 10.1016/0003-2697(71)90370-8. [DOI] [PubMed] [Google Scholar]
- Beers RF, Sizer IW. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J Biol Chem. 1952;195:133–140. [PubMed] [Google Scholar]
- Hassan HM, Fridovich I. Intracellular production of superoxide radical and of hydrogen peroxide by redox active compounds. Arch Biochem Biophys. 1979;196:385–395. doi: 10.1016/0003-9861(79)90289-3. [DOI] [PubMed] [Google Scholar]
- Pick E, Mizel D. Rapid microassays for the measurement of superoxide and hydrogen peroxide production by macrophages in culture using an automatic enzyme immunoassay reader. J Immunol Methods. 1981;46:211–226. doi: 10.1016/0022-1759(81)90138-1. [DOI] [PubMed] [Google Scholar]
- Levine R, Garland D, Oliver C, Amici A, Climent I, Lenz A, Ahn B, Shaltiel S, Stadtman E. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymology. 1990;186:464–478. doi: 10.1016/0076-6879(90)86141-h. [DOI] [PubMed] [Google Scholar]
- Adachi H, Ishii N. Effects of tocotrienols on life span and protein carbonylation in Caenorhabditis elegans. J Gerontol A Biol Sci Med Sci. 2000;55:B280–B285. doi: 10.1093/gerona/55.6.b280. [DOI] [PubMed] [Google Scholar]
- Becker A. A method for glycogen determination in whole yeast cells. Anal Biochem. 1978;86:56–64. doi: 10.1016/0003-2697(78)90318-4. [DOI] [PubMed] [Google Scholar]
- Vandercammen A, Francois J, Her H. Characterization of trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in Saccharomyces cerevisiae . Eur J Biochem. 1989;182(5):613–620. doi: 10.1111/j.1432-1033.1989.tb14870.x. [DOI] [PubMed] [Google Scholar]
- Parrou JL, Francois J. A simplified procedure for a rapid and reliable assay of both glycogen and trehalose in whole yeast cells. Anal Biochem. 1997;248:186–188. doi: 10.1006/abio.1997.2138. [DOI] [PubMed] [Google Scholar]
- Somogyi M. Notes on sugar determination. J Biol Chem. 1952;195:19–23. [PubMed] [Google Scholar]
- Lowry OH, Rosenbrough HJ, Faar AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- Kumar A, John L, Maity S, Manchanda M, Sharma A, Saini N, Chakraborty K, Sengupta S. Converging evidence of mitochondrial dysfunction in a yeast model of homocysteine metabolism imbalance. J Biol Chem. 2011;286:21779–21795. doi: 10.1074/jbc.M111.228072. http://www.jbc.org/content/286/24/21779.full [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osiewacz HD, Stumpferl SW. Metabolism and aging in the filamentous fungus Podospora anserina . Arch Gerontol Geriatr. 2001;32:185–197. doi: 10.1016/s0167-4943(01)00096-6. [DOI] [PubMed] [Google Scholar]
- Azevedo MM, Carvalho A, Pascoal C, Rodrigues F, Cássi F. Responses of antioxidant defenses to Cu and Zn stress in two aquatic fungi. Sci Total Environ. 2007;377(2–3):233–243. doi: 10.1016/j.scitotenv.2007.02.027. [DOI] [PubMed] [Google Scholar]
- Lubaina AS, Meenu Krishnan VG, Murugan K. Induction of oxidative stress and antioxidative response mechanisms in Octoblepharum albidum hedw. A bryophyte under desiccation – rehydration stress. Indian J Plant. 2013;2(3):12–22. [Google Scholar]
- Liu P, Brown S, Goktug T, Channathodiyil P, Kannappan V, Hugnot J-P, Guichet P-O, Bian X, Armesilla AL, Darling JL, Wang W. Cytotoxic effect of disulfiram/copper on human glioblastoma cell lines and ALDH-positive cancer-stem-like cells. Br J Cancer. 2012;107:1488–1497. doi: 10.1038/bjc.2012.442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aileen Pogue I, Jones BM, Bhattacharjee S, Percy ME, Zhao Y, Lukiw WJ. Induced generation of ROS in human brain cells: detection using an isomeric mixture of 5- and 6-carboxy-2′,7′-dichlorofluorescein diacetate (carboxy-DCFDA) as a cell permeant tracer. Int J Mol Sci. 2012;13(8):9615–9626. doi: 10.3390/ijms13089615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poljsak B, Pòcsì I, Pesti M. Interference of chromium with cellular functions. In: Banfalvi G, editor. Cellular effects of heavy metals. London: Springer; 2011. pp. 59–86. [Google Scholar]
- Wang R, Gao F, Guo B-Q, Huang J-C, Wang L, Zhou Y-J. Short-term chromium-stress-induced alterations in the maize leaf proteome. Int J Mol Sci. 2013;14:11125–11144. doi: 10.3390/ijms140611125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stohs SJ, Bagchi D. Oxidative mechanisms in the toxicity of metal ions. Free Radic Biol Med. 1995;18:321–336. doi: 10.1016/0891-5849(94)00159-h. [DOI] [PubMed] [Google Scholar]
- Wang W, Lampi MA, Huang X-D, Gerhardt K, Dixon DG, Greenberg BM. Assessment of mixture toxicity of copper, cadmium, and phenanthrenequinone to the marine bacterium vibrio fischeri. Environ Toxicol. 2008;24(2):166–177. doi: 10.1002/tox.20411. [DOI] [PubMed] [Google Scholar]
- Amrita Nargund AM, Avery SV, Houghton JE. Cadmium induces a heterogeneous and caspase-dependent apoptotic response in Saccharomyces cerevisiae . Apoptosis. 2008;13(6):811–821. doi: 10.1007/s10495-008-0215-8. [DOI] [PubMed] [Google Scholar]
- Lin CY, Trinh NN, Fu S-F, Hsiung Y-C, Chia L-Ch, Lin C-W, Huang H-J. Comparison of early transcriptome responses to copper and cadmium in rice roots. Plant Mol Biol. 2013;81:507–522. doi: 10.1007/s11103-013-0020-9. [DOI] [PubMed] [Google Scholar]
- Cabiscol E, Piulats E, Echave P, Herrero E, Ros J. Oxidative stress promotes specific protein damage in Saccharomyces cerevisiae . J Biol Chem. 2000;275:27393–27398. doi: 10.1074/jbc.M003140200. [DOI] [PubMed] [Google Scholar]
- Francois J, Parrou JL. Reserve carbohydrates metabolism in the yeast Saccharomyces cerevisiae . FEMS Microbiol Rev. 2001;25:125–145. doi: 10.1111/j.1574-6976.2001.tb00574.x. [DOI] [PubMed] [Google Scholar]
- Westwater C, Balish E, Schofield D. Candida albicans-conditioned medium protects yeast cells from oxidative stress: a possible link between quorum sensing and oxidative stress resistance. Eukaryotic Cell. 2005;4(10):1654–1661. doi: 10.1128/EC.4.10.1654-1661.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gasch AP. The environmental stress response: a common yeast response to environmental stresses. In: Hohmann S, Mager P, editors. Yeast stress responses. Heidelberg: Springer-Verlag; 2002. pp. 11–70. [Google Scholar]
- Gomes LC. Physiological responses of pirarucu (Arapaima gigas) to acute handling stress. Acta Amazonica. [online] 2007;37(4):629–633. [Google Scholar]
- Sreenivasa Reddy A, Reddy MV, Radhakrishnaiah K. Impact of copper on the oxidative metabolism of the fry of common carp, Cyprinus carpio (Linn.) at different pH. J Environ Biol. 2008;29(5):721–724. [PubMed] [Google Scholar]
- Emad H, Naga AEL, Khalid M, Moselhy EL, Hamed MA. Toxicity of cadmium and copper and their effect on some biochemical parameters of marine fish, Mugil seheli. Egypt J Aquatic Res. 2005;31:60–71. [Google Scholar]
- Ocón A, Hampp R, Requena N. Trehalose turnover during abiotic stress in arbuscular mycorrhizal fungi. New Phytologist. 2007;174(4):879–891. doi: 10.1111/j.1469-8137.2007.02048.x. [DOI] [PubMed] [Google Scholar]
- Bai Z, Harvey LM, Mcneil B. Oxidative stress in submerged cultures of fungi. Crit Rev Biotechnol. 2003;23:267–302. doi: 10.1080/07388550390449294. [DOI] [PubMed] [Google Scholar]
- Romandini P, Tallandini L, Beltramini M, Salvato B, Manzano M, De Bertoldi M, Rocoo GP. Effects of copper and cadmium on growth, superoxide dismutase and catalase activities in different yeast strains. Comp Biochem Physiol. 1992;103C:255–262. doi: 10.1016/0742-8413(92)90004-q. [DOI] [PubMed] [Google Scholar]
- Belozerskaya T, Aslanidi K, Ivanova A, Gessler N, Egorova A, Karpenko Yu, Olishevskaya S. Characteristics of extremophylic fungi from chernobyl nuclear power plant. In: Mendez-Vilas A, editor. Current research, technology and education topics in applied microbiology and microbial biotechnology. Vol. 1. Badajoz (Spain): Formatex Research Center; 2010. pp. 88–94. [Google Scholar]
- Chattopadhyay MK, Raghu G, Sharma YVRK, Biju AR, Rajasekharan MV, Shivaji S. Increase in oxidative stress at low temperature in an antarctic bacterium. Curr Microbiol. 2011;62(2):544–546. doi: 10.1007/s00284-010-9742-y. [DOI] [PubMed] [Google Scholar]