

Abstract
AIM: To investigate the antifibrotic effects of peginterferon-alpha 2b and taurine on oxidative stress markers and hepatocellular apoptosis.
METHODS: Sixty rats with CCl4-induced liver fibrosis were divided into 4 groups (n = 15). Group 1 was left for spontaneous recovery (SR). Groups 2-4 received peginterferon-alpha 2b, taurine, and their combination, respectively, for four weeks. Histological fibrosis scores, histomorphometric analysis, tissue hydroxyproline, tissue MDA, GPx and SOD activities were determined. Activated stellate cells and hepatocellular apoptosis were also evaluated.
RESULTS: The degree of fibrosis decreased in all treatment groups compared to spontaneous recovery group. Taurine alone and in combination with peginterferon-alpha 2b reduced oxidative stress markers, but peginterferon-alpha 2b alone did not. Apoptotic hepatocytes and activated stellate cells were higher in groups 2-4 than in group 1. Combined taurine and peginterferon-alpha 2b further reduced fibrosis and increased activated stellate cell apoptosis, but could not improve oxidative stress more than taurine alone.
CONCLUSION: Peginterferon-alpha 2b exerts anti-fibrotic effects on rat liver fibrosis. It seems ineffective against oxidative stress in vivo. Peginterferon-alpha 2b in combination with taurine seems to be an antifibrotic strategy.
Keywords: Liver fibrosis, Liver cirrhosis, Pegylated interferons, Taurine, Oxidative stress, Hepatic stelate cells, Apoptosis, Malondialdehyde, Glutathione peroxidase, Superoxide dismutase
INTRODUCTION
Liver fibrosis, an early stage of cirrhosis, occurs as a response to chronic injury from many causes, and is characterized by overproduction and diminished degradation of extracellular matrix proteins in the sinusoids[1,2]. Excessive matrix proteins are synthesized primarily by hepatic stellate cells (HSCs), which activate and acquire a myofibroblast like phenotype in damaged areas of the liver.
Oxidative stress (OS) is associated with liver fibrosis as evidenced both in experimental[3-6] and in clinical studies[7]. Therefore, antioxidant treatment approaches have been studied in a series of experiments. In vitro investigations and studies on animals have revealed beneficial effects of antioxidants in terms of reduction in collagen production and matrix accumulation in the liver[8-11], though, significant improvements in the degree of fibrosis were not found in the clinical setting[12].
Interferons (IFNs) are a family of cytokines with pleiotropic effects on target cells, including induction of an anti-viral state inhibition of cell growth and modulation of the immune response[13]. IFN-alpha (2a and 2b) is the hallmark of treatment in viral hepatitis, which is a major cause of liver cirrhosis and hepatocellular cancer worldwide. Standard IFNs are inferior to the pegylated interferons (peginterferon) in terms of tolerability and attaining sustained response rates in patients with viral hepatitis[14-16]. On the other hand, IFNs display in vitro and in vivo antifibrotic actions with IFN-alpha being the least potent[17-21]. It was reported that the antifibrotic effects of IFN-alpha in rats and patients with hepatitis C virus (HCV) infection are independent from their antiviral activity, i.e., it reduces the degree of fibrosis even when there is no sufficient antiviral response[10,22]. It was also reported that IFN-alpha decreases the production of reactive oxygen species (ROS) in stimulated hepatocytes and HSCs[23], and inhibits OS in patients with HCV infection[24]. However, the activity of recombinant IFN-alpha against oxidative damage has not been confirmed in a rat model of short-term liver injury[10]. It was reported that peginterferons increase the rates of sustained response in patients with viral hepatitis[15,25,26], but their antifibrotic behavior has not been examined widely.
Taurine is a potent antioxidant which prevents DNA damage at concentrations normally present in cells[27]. It was reported that taurine prevents experimental diabetic neuropathy[28], lead-induced oxidative damage[29], CCl4-induced OS[30] as well as experimental liver fibrosis[6,11] through antioxidant mechanisms.
Reversibility of liver fibrosis has been defined in animals and proposed to be associated with loss of activated HSCs mainly through apoptosis in the liver[31,32]. It is also known that the degree of fibrosis decreases after successful treatment of viral hepatitis or haemochromatosis, cessation of alcohol, and biliary decompression[33]. Therefore, enhancement of spontaneous regression capacity in the liver may contribute to etiological treatment providing faster and better reconstitution of liver functions. In this work, we studied the effects of peginterferon-alpha 2b, taurine and their combination on hepatic histology, activation and apoptosis of HSCs, and OS parameters in rat liver fibrosis. Given the evidence of high regeneration capacity in the liver, efficacy of both agents was compared with spontaneous recovery (SR).
MATERIALS AND METHODS
Animals and study protocol
One hundred male Sprague-Dawley rats (250-400 g) were kept at 20-25°C in a 12 h light/12 h dark cycle with free access to food and water. The animals were injected s.c with 0.2 mL/100 g of CCl4 (dissolved 1:1 in sterile olive oil) for two consecutive days per week for 12 wk. Development of fibrosis was confirmed by killing three animals every 6 wk. A total of 60 animals survived the induction period and were divided into four groups (n = 15). Treatment protocols were as follows: group 1 was left for SR for 4 wk without any treatment, group 2 received 1.5 μg/kg peginterferon-alpha 2b per week[32], group 3 was treated with 1200 mg/kg taurine[11] intraperitoneally each day, and group 4 was treated with a combination of these two agents at the same doses. At the end of 4 wk of treatment the animals were killed under i.p. pentobarbital (Nembutal, Abbott) anesthesia, and their livers were rapidly removed. Liver samples were processed for histological and biochemical studies. The experiments, approved by the Institutional Animal Use and Care Committee of the Gulhane Medical Academy, Ankara, Turkey, were performed according to the criteria of the National Institutes of Health guidelines for the care and handling of animals.
Assessment of fibrosis
Histological scoring: Liver samples were fixed in 4% buffered formaldehyde, embedded in paraffin, and stained with hematoxylin-eosin. Masson’s trichrome staining was performed for extracellular matrix evaluation. Histologic evaluation was performed twice by two pathologists blinded to the protocol on five low-power fields per slide. Modified Ishak scoring system was used with minor changes in order to define the degree of fibrosis[34] (Table 1).
Table 1.
Histological scoring model
| 0 | No fibrosis or minimal fibrous expansion of occasional portal areas |
| 1 | Moderate fibrous expansion of some portal areas with short fibrous septae |
| 2 | Moderate fibrous expansion of most portal areas, occasional portal-to-portal bridging |
| 3 | None or minimal expansion of portal areas, porto-portal fibrosis, incomplete bridging nodules |
| 4 | Marked portal fibrosis, portal to portal fibrosis, occasional micronodules |
| 5 | Mild to moderate fibrous expansion in portal areas, micronodules, incomplete cirrhosis |
| 6 | Marked fibrosis in portal areas, macro- and micronodules with thick fibrous septae, complete cirrhosis |
Histomorphometric analysis: Percentage of fibrosis was determined by a computer system composed of the following components: an IBM compatible computer running Microsoft Windows NT 4.0 service pack 6 operating system, a light microscope with motorized stage (Zeiss axioscope, Gottingen, Germany), frame grabber card (Matrox Meteor, Matrox imaging Dorval, Quebec, Canada), a digital camera attached to the microscope (Sony AVT Horn 3 CCD, Tokyo), a 21 inch color video monitor (Philips, Chungli, Taiwan), and a special image analysis software (Karl Zeiss KS 400 version 3.0 for Windows, Zeiss, Gottingen, Germany). With the aid of KS 400 for Windows, a software using a semiautomatic, interactive "macro" (a batch file containing all of the instructions to accomplish a predetermined set of measurements) was prepared.
Measurement was started with selection of the area of interest under a plan 2.5X objective. Then, this area was scanned automatically as a consecutive rectangular area under a plan 20X objective. The fibrotic areas and parenchyma were counted separately and summed up by the software at the end of each measurement. The results were expressed as the percentage of fibrosis.
Measurement of hydroxyproline: Liver tissue hydroxy-proline, which represents a reliable indicator of tissue collagen content in the liver, was quantified as previously described[35]. Small modifications in the duration of incubation of liver tissues in Ehrlich’s solution (35 min at 60°C instead of 25 min at 60°C) and dissolving hydroxyproline in the homogenates by concentrated HCl at 110°C were applied.
Measurement of oxidative stress
Liver tissue malondialdehyde (MDA), glutathione peroxidase (GPx) and superoxide dismutase (SOD) measurements were performed as described previously[36-38] to determine the oxidative status in the liver specimens. MDA levels were expressed as nmol/g tissue. GPx and SOD activities were expressed as U/g tissue dye.
Immunohistochemical tests
Detection of activated stellate cells: Alpha-smooth muscle actin (alpha-SMA) immunostaining was used to detect and count the activated HSCs[39]. For this purpose, the tissue samples were stained for monoclonal rat anti-alpha-SMA (Sigma) with lbeled streptavidin-biotin (LSAB, Dako) complex method as described elsewhere[31], and analyzed by two blinded observers for the number of apoptotic HSCs in 30 random high-power fields in each specimen.
Detection of apoptotic hepatocytes and hepatic stellate cells: Hepatocytes and activated HSCs undergoing apoptosis were detected by terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling (TUNEL) staining[11,31] using a commercial ready-to-use kit (In situ cell death detection kit, AP, Roche Diagnostics, Germany) as previously described[32]. Slides were analyzed by two blinded observers who counted the number of TUNEL positive hepatocytes and HSCs in 30 random high-power fields from each specimen.
Statistical analysis
Parametric results were expressed as mean ± SE, and the significance of differences was assessed by one-way ANOVA and Tukey’s HSD as post hoc. Nonparametric results (histological scores) were expressed as median and percentiles (25%-75%) and assessed by Mann-Whitney U test. Correlation analysis was assessed with Pearson Correlation procedure. The differences were accepted as statistically significant when P value was less than 0.05.
RESULTS
Effects on fibrosis
Histological scores: Peginterferon-alpha 2b (group 2), taurine (group 3) and their combination (group 4) reduced fibrosis scores when compared to SR group (group 1) (P < 0.01) (Table 2). Fibrosis scores were similar in groups 2-4. Combination treatment did not reduce fibrosis scores compared to individual treatments. The histological sections of fibrotic rat livers from four groups are shown in Figure 1.
Table 2.
Results of fibrosis, oxidative stress parameters, activated HSC counts and apoptotic cells in groups 1-4
| Spontaneous recovery (n = 15) | Peginterferon-alpha (n = 15) | Taurine (n = 15) | Peginterferon-alpha plus Taurine (n = 15) | |
| Histological score | 5 (5-6) | 3 (2-4) | 2 (1-3) | 3 (2-4) |
| Percentage fibrosis (%) | 14.3 ± 0.8 | 7.9 ± 0.5 | 7.9 ± 0.6 | 6.1 ± 0.3 |
| Hydroxyproline (μg/g wet weight) | 1.03 ± 0.07 | 0.58 ± 0.04 | 0.64 ± 0.03 | 0.32 ± 0.04 |
| MDA (nmol/g tissue) | 47.8 ± 1.0 | 43.3 ± 1.8 | 20.2 ± 0.8 | 18.4 ± 0.6 |
| GPx (U/g tissue) | 128 ± 3 | 168 ± 6 | 271 ± 13 | 270 ± 11 |
| SOD (U/g tissue) | 343 ± 5 | 385 ± 8 | 644 ± 14 | 619 ± 19 |
| alpha-SMA (+) cell count | 120.0 ± 9.4 | 59.6 ± 2.7 | 46.7 ± 3.5 | 32.6 ± 3.0 |
| Apoptotic hepatocyte count | 19.1 ± 1.0 | 31.3 ± 2.6 | 34.8 ± 3.9 | 43.9 ± 4.0 |
| Apoptotic activated HSC count | 4.6 ± 0.5 | 14.1 ± 1.5 | 11.0 ± 0.8 | 21.5 ± 2.0 |
MDA: Malondialdehyde; GPx: Glutathione peroxidase; SOD: Superoxide dismutase; alpha-SMA: alpha-smooth muscle actin.
Figure 1.

Sections of fibrotic rat livers from four groups. A: Mild-moderate fibrous-expansion in portal areas and several micronodules in group 1 (Masson’s trichrome stain, × 100); B: Moderate expansion of portal areas and porto-portal fibrosis in group 2 (Masson’s trichrome stain, × 200); C: Moderate fibrous expansion of most portal areas and porto-portal bridging in group 3 (Masson’s trichrome stain, × 200); D: Mild fibrous expansion of some portal areas in group 4 (Masson’s trichrome stain, × 200).
Histomorphometry: The percentage of fibrotic areas was lower in groups 2-4, in line with histological scores and hydroxyproline measurement than in group 1 (P < 0.01), whereas it was similar in individual treatment groups. Combined treatment reduced percent fibrosis significantly when compared to peginterferon-alpha 2b or taurine alone (P < 0.05) (Table 2).
Hydroxyproline: Liver tissue hydroxyproline decreased in groups 2-4 when compared to group 1 (P < 0.01) (Table 2). Hydroxyproline contents were similar in groups 2 and 3. Combination treatment reduced tissue hydroxyproline more significantly than individual treatments (P < 0.05 and P < 0.01 for combination vs peginterferon-alpha 2b and taurine, respectively).
Effects on oxidative stress
Compared to group 1, liver tissue MDA levels decreased, and GPx and SOD activities increased in groups 3 and 4 (P < 0.01), but not in group 2 (Table 2). Similar results were also obtained in groups 3 and 4 when compared to group 2 (P < 0.01). Decrease in MDA and increase in GPx and SOD activities in groups 3 and 4 were comparable.
Correlation analysis
We evaluated the predictive value of histological fibrosis scores or OS markers regarding hydroxyproline content and found that the amount of liver tissue hydroxyproline content correlated with the percentage of fibrosis and histological score as well as tissue MDA, GPx and SOD (P < 0.05).
Effects on the number of alpha-smooth muscle actin positive HSCs
The number of alpha-SMA (+) HSCs was lower in groups 2-4 than in group 1 (P < 0.01). Groups 2 and 3 had similar alpha-SMA (+) HSC counts (P > 0.05) (Table 2). The number of alpha-SMA (+) HSCs was lower in group 4 than in group 2 (P < 0.05), whereas no difference was found between groups 4 and 3 (P > 0.05).
Effects on the number of apoptotic hepatocytes and activated hepatic stellate cells
TUNEL positive hepatocytes were comparable in groups 2 and 3 (Table 2). The number of hepatocytes undergoing apoptosis was higher in groups 2-4 than in group 1 (P < 0.05, respectively). The apoptotic hepatocyte number was also higher in group 4 than in group 2 (P < 0.05). There was no difference between groups 3 and 4.
The number of alpha-SMA (+) apoptotic HSCs (dually stained with anti-alpha-SMA and TUNEL technique) was increased in groups 2-4 in comparison with group 1 (P < 0.05). The sinusoidal lining as well as activated and apoptotic stellate cells are shown in Figure 2. Combined treatment increased activated HSC apoptosis more significantly than either peginterferon-alpha 2b or taurine alone (P < 0.01). Apoptotic HSC count was similar in groups 2 and 3.
Figure 2.
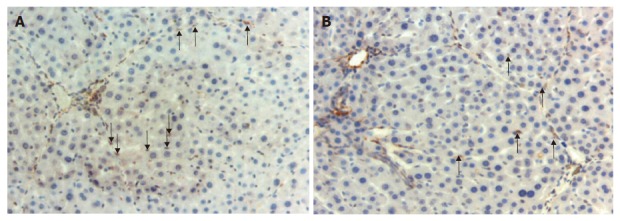
Activated and apoptotic hepatic stellate cells. A: α-SMA (+) stellate cells in the sinusoids in a section from group 3 (arrows) (× 200); B: Apoptotic activated stellate cells positively stained with TUNEL technique in the sample from group 2 (arrows) (× 200).
The cause(s) and therapeutic alternatives for liver fibrosis are shown in Figure 3.
Figure 3.
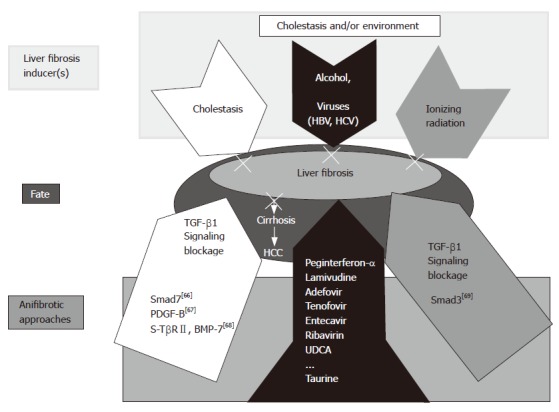
Working hypothesis: cause(s) and therapeutic alternatives for liver fibrosis. In the present study, a combined peginterferon-α plus taurine treatment for fibrotic livers was applied successfully. Though different etiologies are responsible for chronic liver diseases, viruses and alcohol prevail worldwide. Our results can represent a therapeutic option to prevent progression of the underlying disease to fibrosis in the liver. Other experimental models are also presented including TGF-β1 signaling blockage by Smad7 overexpression[66], PDGF-B[67], and a soluble receptor TGF-β2 (TβRII) and BMP-7[68]. Liver fibrosis affected by ionizing radiation can be inhibited by Smad knock out [Smad3 (-)][69].
DISCUSSION
In the present investigation, peginterferon-alpha 2b and taurine improved experimental liver fibrosis in an equal pattern, and their combination yielded further benefits. Improvement of fibrosis in the liver was accompanied with decrease in OS as well as increase in the number of activated HSCs undergoing apoptosis. This study was designed to investigate the therapeutic efficacy, but not the preventive effects of peginterferon-alpha 2b and taurine in chronically injured liver.
Involvement of OS in activation of HSCs and initiation of liver fibrosis has been well described[40,41]. Moreover, an increased state of oxidative activity[24,42-44] and decreased antioxidant levels[45] have also been shown in patients with chronic HCV infection and cirrhosis. Upon injury, major reactive oxygen intermediates (ROIs) and aldehydic end products of lipid peroxidation, which are released by injured hepatocytes and activated inflammatory cells such as neutrophils, macrophages and Kupffer cells, deteriorate cellular signal transduction as well as proliferative and functional responses[46]. Macrophages, known to activate with a variety of stimuli including exposure to abnormally deposited collagenous matrix[47-49], have recently been shown to play an important role in both fibrogenesis and its resolution in the liver. It was reported that ROIs released by these cells can limit activation of cytotoxic lymphocytes by IFN-alpha[50], suggestive of usefulness of antioxidants, such as taurine, in enhancing immunostimulatory efficiency of this agent.
In chronic liver diseases, an imbalance occurs between the antioxidant defenses and OS, which may result in cytotoxicity, apoptotic cell death and fibrosis[51]. It was reported that several antioxidants can prevent activation of HSCs, decrease collagen synthesis and improve fibrosis in the liver of animals[9-11,52]. However, the results provided by clinical trials with antioxidants are less impressive[53], in particular, when histological changes are concerned[7,12,54]. Nevertheless, marked improvements in OS parameters reported in experimental studies indicate that these agents work in the liver. But, to get clearer results on fibrosis, optimal starting time, dose and duration of the treatment should be fully established. Of note, more potent antioxidants and combined treatment approaches with agents with proven antifibrotic activity might also be more useful.
Taurine is a potent antioxidant which has been shown to improve liver histology in experimental conditions mainly through decreasing the production of ROS[6,11]. The present study confirms the previous findings that showed taurine’s ability to exert an antioxidant activity at the tissue level in fibrotic rat livers. Therefore, histological recovery in the taurine-treated group in the present study could be attributed to the decrease in OS. Indeed, increased levels of OS markers exist not only in the early stages of liver diseases but also in advanced states such as cirrhosis[43]. Moreover, although experimental studies have revealed controversial results[55], it is possible that natural defense mechanisms against oxidant injury may fail to prevent progression of the disease further. Thus, attempts to decrease OS even in the advanced stages of liver diseases could be beneficial. However, Look et al[12] reported that decreases in markers of OS is not accompanied with histological improvement, though the duration of treatment in their study was relatively short for adaptation of antioxidant systems.
Since there is evidence that supports the existence of an increased oxidative status in patients with chronic viral hepatitis[42,43], several authors have studied the effects of IFN treatment on the parameters of OS. In dimethylnitrosamine-induced liver fibrosis, recombinant IFN-alpha was found to exert no significant antioxidant activity[10]. In contrast, Lu et al[23] have reported in vitro antioxidant effects of IFN-alpha on isolated rat hepatocytes and HSCs. Though the exact mode of antifibrotic action of IFN-alpha has not been fully elucidated to date, a so called direct antioxidant effect may also be involved in the mechanism of its antifibrotic features. However, the present in vivo investigation also confirms that peginterferon-alpha 2b does not display any effect on markers of OS in the liver.
Our results indicate that combination of peginterferon-alpha 2b with taurine could result in significant improvement in the degree of fibrosis, but could not enhance the resultant antioxidant efficacy. Several experimental and clinical studies partly confirming our results have previously addressed the biochemical and histological changes after treatment with IFN-alpha combined with antioxidants. Addition of N-acetylcysteine could not result in further decrease in liver collagen content in dimethyl nitrosamine-induced rat fibrosis[10]. In a pilot study, similar results were also found in patients with HCV infection treated with standard IFN-alpha plus antioxidants. But, no further decrease was observed in the serum markers of OS when compared to IFN-alpha alone[12]. However, a decrease in OS correlated with fibrosis grade has been reported in IFN-alpha plus antioxidant-treated hepatitis C patients[24]. These contradictory reports suggest that more reliable data are needed regarding the clinical expression of IFN effects over oxidative damage. Moreover, as yet there is no clinical study on combination of peginterferon-alpha 2b with an antioxidant.
It was reported that liver fibrosis can regress spon-taneously mainly through increased rate of apoptosis of activated HSCs[31]. Therefore, inducing apoptosis of activated HSCs in the liver might be an effective treatment approach. It has been shown that taurine acts as an antiapoptotic agent in some types of cells under different conditions[56,57]. However, no effect on isolated HSCs[58] and even in vivo proapoptotic effects[11] were reported. Also in the present study, taurine induced apoptosis of hepatocytes and activated HSCs in the resolution period of liver fibrosis, which possibly resulted from the decrease in the degree of OS. ROIs released from resident liver cells and activated inflammatory cells like macrophages are potential inducers of apoptosis in the healing liver. While severe OS results in rapid cell necrosis, less severe OS has been suggested to cause liver cell apoptosis[51], which is a desired and a proposed condition in recovery of liver fibrosis[31]. Therefore, reducing OS may prevent liver cell loss by necrosis, but induce programmed death of phenotypically changed cells in the liver.
There is also a delicate interaction of ROIs with nitric oxide (NO), which is produced by liver cells, including HSCs, as well as other cell types such as macrophages in conditions with chronic inflammation[59]. NO can reduce liver injury by deceasing generation of OH[60] and lipid peroxidation products[61] with a so-called antioxidant activity. However, in the states of increased OS where high levels of ROIs are present, NO can interact with ROIs to produce cytotoxic compounds[61]. Similarly, increased levels of NO can lead to apoptosis of several cell types in the presence of superoxide anions that react with NO to produce peroxynitrite, which seems to be an inducer of apoptosis[59,62]. It was reported that NO synthesis is up-regulated in vitro in IFN-alpha treated monocytes through induction of nitric oxide synthase[63]. It was also reported that IFN-treated HCV patients have significantly higher nitric oxide synthase activity in their mononuclear cells, and serum nitrite and nitrate levels are increased in IFN-alpha responder patients[64]. Moreover, though activated HSCs play an in vitro direct antiapoptotic role in liver diseases[65], IFN-alpha-associated increase in NO concentration might induce apoptosis of these cells. The increased number of hepatocytes and HSCs undergoing apoptosis in peginterferon-alpha 2b and taurine-treated animals as well as in the combined treatment group in our study seems to be associated with multiple, but as yet unidentified mechanisms.
In conclusion, the results of this study indicate that peginterferons, like the conventional IFNs, effectively reduce fibrosis in a rat model of liver fibrosis. But, peginteferon-alpha 2b has no antioxidant action. The reduction in OS is accompanied with an increase in activated HSC apoptosis. The additive antifibrotic effects of peginterferons with taurine may have clinical implications.
Footnotes
Supported by the Gulhane School of Medicine Research Council (AR-02-15)
S- Editor Liu Y L- Editor Wang XL E- Editor Liu Y
References
- 1.Prosser CC, Yen RD, Wu J. Molecular therapy for hepatic injury and fibrosis: where are we? World J Gastroenterol. 2006;12:509–515. doi: 10.3748/wjg.v12.i4.509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Thomson RK, Arthur MJ. Mechanisms of liver cell damage and repair. Eur J Gastroenterol Hepatol. 1999;11:949–955. doi: 10.1097/00042737-199909000-00002. [DOI] [PubMed] [Google Scholar]
- 3.Parola M, Pinzani M, Casini A, Albano E, Poli G, Gentilini A, Gentilini P, Dianzani MU. Stimulation of lipid peroxidation or 4-hydroxynonenal treatment increases procollagen alpha 1 (I) gene expression in human liver fat-storing cells. Biochem Biophys Res Commun. 1993;194:1044–1050. doi: 10.1006/bbrc.1993.1927. [DOI] [PubMed] [Google Scholar]
- 4.Britton RS, Bacon BR. Role of free radicals in liver diseases and hepatic fibrosis. Hepatogastroenterology. 1994;41:343–348. [PubMed] [Google Scholar]
- 5.Svegliati Baroni G, D'Ambrosio L, Ferretti G, Casini A, Di Sario A, Salzano R, Ridolfi F, Saccomanno S, Jezequel AM, Benedetti A. Fibrogenic effect of oxidative stress on rat hepatic stellate cells. Hepatology. 1998;27:720–726. doi: 10.1002/hep.510270313. [DOI] [PubMed] [Google Scholar]
- 6.Miyazaki T, Karube M, Matsuzaki Y, Ikegami T, Doy M, Tanaka N, Bouscarel B. Taurine inhibits oxidative damage and prevents fibrosis in carbon tetrachloride-induced hepatic fibrosis. J Hepatol. 2005;43:117–125. doi: 10.1016/j.jhep.2005.01.033. [DOI] [PubMed] [Google Scholar]
- 7.Houglum K, Venkataramani A, Lyche K, Chojkier M. A pilot study of the effects of d-alpha-tocopherol on hepatic stellate cell activation in chronic hepatitis C. Gastroenterology. 1997;113:1069–1073. doi: 10.1053/gast.1997.v113.pm9322499. [DOI] [PubMed] [Google Scholar]
- 8.Lee KS, Buck M, Houglum K, Chojkier M. Activation of hepatic stellate cells by TGF alpha and collagen type I is mediated by oxidative stress through c-myb expression. J Clin Invest. 1995;96:2461–2468. doi: 10.1172/JCI118304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Soylu AR, Aydogdu N, Basaran UN, Altaner S, Tarcin O, Gedik N, Umit H, Tezel A, Dokmeci G, Baloglu H, et al. Antioxidants vitamin E and C attenuate hepatic fibrosis in biliary-obstructed rats. World J Gastroenterol. 2006;12:6835–6841. doi: 10.3748/wjg.v12.i42.6835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Vendemiale G, Grattagliano I, Caruso ML, Serviddio G, Valentini AM, Pirrelli M, Altomare E. Increased oxidative stress in dimethylnitrosamine-induced liver fibrosis in the rat: effect of N-acetylcysteine and interferon-alpha. Toxicol Appl Pharmacol. 2001;175:130–139. doi: 10.1006/taap.2001.9234. [DOI] [PubMed] [Google Scholar]
- 11.Refik Mas M, Comert B, Oncu K, Vural SA, Akay C, Tasci I, Ozkomur E, Serdar M, Mas N, Alcigir G, et al. The effect of taurine treatment on oxidative stress in experimental liver fibrosis. Hepatol Res. 2004;28:207–215. doi: 10.1016/j.hepres.2003.11.012. [DOI] [PubMed] [Google Scholar]
- 12.Look MP, Gerard A, Rao GS, Sudhop T, Fischer HP, Sauerbruch T, Spengler U. Interferon/antioxidant combination therapy for chronic hepatitis C--a controlled pilot trial. Antiviral Res. 1999;43:113–122. doi: 10.1016/s0166-3542(99)00041-8. [DOI] [PubMed] [Google Scholar]
- 13.Borden EC. Interferons--expanding therapeutic roles. N Engl J Med. 1992;326:1491–1493. doi: 10.1056/NEJM199205283262209. [DOI] [PubMed] [Google Scholar]
- 14.von Wagner M, Huber M, Berg T, Hinrichsen H, Rasenack J, Heintges T, Bergk A, Bernsmeier C, Häussinger D, Herrmann E, et al. Peginterferon-alpha-2a (40KD) and ribavirin for 16 or 24 weeks in patients with genotype 2 or 3 chronic hepatitis C. Gastroenterology. 2005;129:522–527. doi: 10.1016/j.gastro.2005.05.008. [DOI] [PubMed] [Google Scholar]
- 15.Glue P, Fang JW, Rouzier-Panis R, Raffanel C, Sabo R, Gupta SK, Salfi M, Jacobs S. Pegylated interferon-alpha2b: pharmacokinetics, pharmacodynamics, safety, and preliminary efficacy data. Hepatitis C Intervention Therapy Group. Clin Pharmacol Ther. 2000;68:556–567. doi: 10.1067/mcp.2000.110973. [DOI] [PubMed] [Google Scholar]
- 16.Manns MP, McHutchison JG, Gordon SC, Rustgi VK, Shiffman M, Reindollar R, Goodman ZD, Koury K, Ling M, Albrecht JK. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: a randomised trial. Lancet. 2001;358:958–965. doi: 10.1016/s0140-6736(01)06102-5. [DOI] [PubMed] [Google Scholar]
- 17.Chang XM, Chang Y, Jia A. Effects of interferon-alpha on expression of hepatic stellate cell and transforming growth factor-beta1 and alpha-smooth muscle actin in rats with hepatic fibrosis. World J Gastroenterol. 2005;11:2634–2636. doi: 10.3748/wjg.v11.i17.2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shen H, Zhang M, Minuk GY, Gong Y. Different effects of rat interferon alpha, beta and gamma on rat hepatic stellate cell proliferation and activation. BMC Cell Biol. 2002;3:9. doi: 10.1186/1471-2121-3-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guido M, Rugge M, Chemello L, Leandro G, Fattovich G, Giustina G, Cassaro M, Alberti A. Liver stellate cells in chronic viral hepatitis: the effect of interferon therapy. J Hepatol. 1996;24:301–307. doi: 10.1016/s0168-8278(96)80008-0. [DOI] [PubMed] [Google Scholar]
- 20.Capra F, Casaril M, Gabrielli GB, Tognella P, Rizzi A, Dolci L, Colombari R, Mezzelani P, Corrocher R, De Sandre G. alpha-Interferon in the treatment of chronic viral hepatitis: effects on fibrogenesis serum markers. J Hepatol. 1993;18:112–118. doi: 10.1016/s0168-8278(05)80018-2. [DOI] [PubMed] [Google Scholar]
- 21.Hiramatsu N, Hayashi N, Kasahara A, Hagiwara H, Takehara T, Haruna Y, Naito M, Fusamoto H, Kamada T. Improvement of liver fibrosis in chronic hepatitis C patients treated with natural interferon alpha. J Hepatol. 1995;22:135–142. doi: 10.1016/0168-8278(95)80420-x. [DOI] [PubMed] [Google Scholar]
- 22.Yagura M, Murai S, Kojima H, Tokita H, Kamitsukasa H, Harada H. Changes of liver fibrosis in chronic hepatitis C patients with no response to interferon-alpha therapy: including quantitative assessment by a morphometric method. J Gastroenterol. 2000;35:105–111. doi: 10.1007/s005350050022. [DOI] [PubMed] [Google Scholar]
- 23.Lu G, Shimizu I, Cui X, Itonaga M, Tamaki K, Fukuno H, Inoue H, Honda H, Ito S. Interferon-alpha enhances biological defense activities against oxidative stress in cultured rat hepatocytes and hepatic stellate cells. J Med Invest. 2002;49:172–181. [PubMed] [Google Scholar]
- 24.Serejo F, Emerit I, Filipe PM, Fernandes AC, Costa MA, Freitas JP, de Moura MC. Oxidative stress in chronic hepatitis C: the effect of interferon therapy and correlation with pathological features. Can J Gastroenterol. 2003;17:644–650. doi: 10.1155/2003/710693. [DOI] [PubMed] [Google Scholar]
- 25.Abergel A, Darcha C, Chevallier M, Ughetto S, Henquell C, Pol S, de Ledinghen V, Canva V, Bronowicki JP, Tran A, et al. Histological response in patients treated by interferon plus ribavirin for hepatitis C virus-related severe fibrosis. Eur J Gastroenterol Hepatol. 2004;16:1219–1227. doi: 10.1097/00042737-200411000-00022. [DOI] [PubMed] [Google Scholar]
- 26.Lindsay KL, Trepo C, Heintges T, Shiffman ML, Gordon SC, Hoefs JC, Schiff ER, Goodman ZD, Laughlin M, Yao R, et al. A randomized, double-blind trial comparing pegylated interferon alfa-2b to interferon alfa-2b as initial treatment for chronic hepatitis C. Hepatology. 2001;34:395–403. doi: 10.1053/jhep.2001.26371. [DOI] [PubMed] [Google Scholar]
- 27.Messina SA, Dawson R. Attenuation of oxidative damage to DNA by taurine and taurine analogs. Adv Exp Med Biol. 2000;483:355–367. doi: 10.1007/0-306-46838-7_40. [DOI] [PubMed] [Google Scholar]
- 28.Obrosova IG, Fathallah L, Stevens MJ. Taurine counteracts oxidative stress and nerve growth factor deficit in early experimental diabetic neuropathy. Exp Neurol. 2001;172:211–219. doi: 10.1006/exnr.2001.7789. [DOI] [PubMed] [Google Scholar]
- 29.Gürer H, Ozgünes H, Saygin E, Ercal N. Antioxidant effect of taurine against lead-induced oxidative stress. Arch Environ Contam Toxicol. 2001;41:397–402. doi: 10.1007/s002440010265. [DOI] [PubMed] [Google Scholar]
- 30.Vohra BP, Hui X. Taurine protects against carbon tetrachloride toxicity in the cultured neurons and in vivo. Arch Physiol Biochem. 2001;109:90–94. doi: 10.1076/apab.109.1.90.4287. [DOI] [PubMed] [Google Scholar]
- 31.Iredale JP, Benyon RC, Pickering J, McCullen M, Northrop M, Pawley S, Hovell C, Arthur MJ. Mechanisms of spontaneous resolution of rat liver fibrosis. Hepatic stellate cell apoptosis and reduced hepatic expression of metalloproteinase inhibitors. J Clin Invest. 1998;102:538–549. doi: 10.1172/JCI1018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tasci I, Mas MR, Vural SA, Comert B, Alcigir G, Serdar M, Mas N, Isik AT, Ates Y. Rat liver fibrosis regresses better with pegylated interferon alpha2b and ursodeoxycholic acid treatments than spontaneous recovery. Liver Int. 2006;26:261–268. doi: 10.1111/j.1478-3231.2005.01210.x. [DOI] [PubMed] [Google Scholar]
- 33.Bonis PA, Friedman SL, Kaplan MM. Is liver fibrosis reversible? N Engl J Med. 2001;344:452–454. doi: 10.1056/NEJM200102083440610. [DOI] [PubMed] [Google Scholar]
- 34.Ishak K, Baptista A, Bianchi L, Callea F, De Groote J, Gudat F, Denk H, Desmet V, Korb G, MacSween RN. Histological grading and staging of chronic hepatitis. J Hepatol. 1995;22:696–699. doi: 10.1016/0168-8278(95)80226-6. [DOI] [PubMed] [Google Scholar]
- 35.Bergman I, Loxley R. Two improved and simplified methods for the spectrophotometric determination of hydroxyproline. Anal Chem. 1963;35:1961–1965. [Google Scholar]
- 36.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–358. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 37.Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med. 1967;70:158–169. [PubMed] [Google Scholar]
- 38.Aydin A, Orhan H, Sayal A, Ozata M, Sahin G, Işimer A. Oxidative stress and nitric oxide related parameters in type II diabetes mellitus: effects of glycemic control. Clin Biochem. 2001;34:65–70. doi: 10.1016/s0009-9120(00)00199-5. [DOI] [PubMed] [Google Scholar]
- 39.Enzan H, Himeno H, Iwamura S, Saibara T, Onishi S, Yamamoto Y, Hara H. Immunohistochemical identification of Ito cells and their myofibroblastic transformation in adult human liver. Virchows Arch. 1994;424:249–256. doi: 10.1007/BF00194608. [DOI] [PubMed] [Google Scholar]
- 40.Svegliati-Baroni G, Saccomanno S, van Goor H, Jansen P, Benedetti A, Moshage H. Involvement of reactive oxygen species and nitric oxide radicals in activation and proliferation of rat hepatic stellate cells. Liver. 2001;21:1–12. doi: 10.1034/j.1600-0676.2001.210101.x. [DOI] [PubMed] [Google Scholar]
- 41.Galli A, Svegliati-Baroni G, Ceni E, Milani S, Ridolfi F, Salzano R, Tarocchi M, Grappone C, Pellegrini G, Benedetti A, et al. Oxidative stress stimulates proliferation and invasiveness of hepatic stellate cells via a MMP2-mediated mechanism. Hepatology. 2005;41:1074–1084. doi: 10.1002/hep.20683. [DOI] [PubMed] [Google Scholar]
- 42.Yamamoto Y, Yamashita S. Plasma ubiquinone to ubiquinol ratio in patients with hepatitis, cirrhosis, and hepatoma, and in patients treated with percutaneous transluminal coronary reperfusion. Biofactors. 1999;9:241–246. doi: 10.1002/biof.5520090219. [DOI] [PubMed] [Google Scholar]
- 43.Bianchi G, Marchesini G, Fabbri A, Ronchi M, Chianese R, Grossi G. Lipoperoxide plasma levels in patients with liver cirrhosis. Hepatogastroenterology. 1997;44:784–788. [PubMed] [Google Scholar]
- 44.Emerit I, Serejo F, Filipe P, Alaoui Youssefi A, Fernandes A, Costa A, Freitas J, Ramalho F, Baptista A, Carneiro de Moura M. Clastogenic factors as biomarkers of oxidative stress in chronic hepatitis C. Digestion. 2000;62:200–207. doi: 10.1159/000007814. [DOI] [PubMed] [Google Scholar]
- 45.Bandara P, George J, McCaughan G, Naidoo D, Lux O, Salonikas C, Kench J, Byth K, Farrell GC. Antioxidant levels in peripheral blood, disease activity and fibrotic stage in chronic hepatitis C. Liver Int. 2005;25:518–526. doi: 10.1111/j.1478-3231.2005.01049.x. [DOI] [PubMed] [Google Scholar]
- 46.Parola M, Robino G. Oxidative stress-related molecules and liver fibrosis. J Hepatol. 2001;35:297–306. doi: 10.1016/s0168-8278(01)00142-8. [DOI] [PubMed] [Google Scholar]
- 47.Duffield JS, Forbes SJ, Constandinou CM, Clay S, Partolina M, Vuthoori S, Wu S, Lang R, Iredale JP. Selective depletion of macrophages reveals distinct, opposing roles during liver injury and repair. J Clin Invest. 2005;115:56–65. doi: 10.1172/JCI22675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhang-Hoover J, Sutton A, van Rooijen N, Stein-Streilein J. A critical role for alveolar macrophages in elicitation of pulmonary immune fibrosis. Immunology. 2000;101:501–511. doi: 10.1046/j.1365-2567.2000.00143.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tsunawaki S, Sporn M, Ding A, Nathan C. Deactivation of macrophages by transforming growth factor-beta. Nature. 1988;334:260–262. doi: 10.1038/334260a0. [DOI] [PubMed] [Google Scholar]
- 50.Hansson M, Romero A, Thorén F, Hermodsson S, Hellstrand K. Activation of cytotoxic lymphocytes by interferon-alpha: role of oxygen radical-producing mononuclear phagocytes. J Leukoc Biol. 2004;76:1207–1213. doi: 10.1189/jlb.0204113. [DOI] [PubMed] [Google Scholar]
- 51.Kaplowitz N. Mechanisms of liver cell injury. J Hepatol. 2000;32:39–47. doi: 10.1016/s0168-8278(00)80414-6. [DOI] [PubMed] [Google Scholar]
- 52.Kanter M, Coskun O, Budancamanak M. Hepatoprotective effects of Nigella sativa L and Urtica dioica L on lipid peroxidation, antioxidant enzyme systems and liver enzymes in carbon tetrachloride-treated rats. World J Gastroenterol. 2005;11:6684–6688. doi: 10.3748/wjg.v11.i42.6684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Idéo G, Bellobuono A, Tempini S, Mondazzi L, Airoldi A, Benetti G, Bissoli F, Cestari C, Colombo E, Del Poggio P, et al. Antioxidant drugs combined with alpha-interferon in chronic hepatitis C not responsive to alpha-interferon alone: a randomized, multicentre study. Eur J Gastroenterol Hepatol. 1999;11:1203–1207. doi: 10.1097/00042737-199911000-00003. [DOI] [PubMed] [Google Scholar]
- 54.Parés A, Planas R, Torres M, Caballería J, Viver JM, Acero D, Panés J, Rigau J, Santos J, Rodés J. Effects of silymarin in alcoholic patients with cirrhosis of the liver: results of a controlled, double-blind, randomized and multicenter trial. J Hepatol. 1998;28:615–621. doi: 10.1016/s0168-8278(98)80285-7. [DOI] [PubMed] [Google Scholar]
- 55.Maher JJ, Saito JM, Neuschwander-Tetri BA. Glutathione regulation in rat hepatic stellate cells. Comparative studies in primary culture and in liver injury in vivo. Biochem Pharmacol. 1997;53:637–641. doi: 10.1016/s0006-2952(96)00865-9. [DOI] [PubMed] [Google Scholar]
- 56.Wu QD, Wang JH, Fennessy F, Redmond HP, Bouchier-Hayes D. Taurine prevents high-glucose-induced human vascular endothelial cell apoptosis. Am J Physiol. 1999;277:C1229–C1238. doi: 10.1152/ajpcell.1999.277.6.C1229. [DOI] [PubMed] [Google Scholar]
- 57.Alfieri RR, Cavazzoni A, Petronini PG, Bonelli MA, Caccamo AE, Borghetti AF, Wheeler KP. Compatible osmolytes modulate the response of porcine endothelial cells to hypertonicity and protect them from apoptosis. J Physiol. 2002;540:499–508. doi: 10.1113/jphysiol.2001.013395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chen YX, Zhang XR, Xie WF, Li S. Effects of taurine on proliferation and apoptosis of hepatic stellate cells in vitro. Hepatobiliary Pancreat Dis Int. 2004;3:106–109. [PubMed] [Google Scholar]
- 59.Clemens MG. Nitric oxide in liver injury. Hepatology. 1999;30:1–5. doi: 10.1002/hep.510300148. [DOI] [PubMed] [Google Scholar]
- 60.Li J, Billiar TR. Nitric Oxide. IV. Determinants of nitric oxide protection and toxicity in liver. Am J Physiol. 1999;276:G1069–G1073. doi: 10.1152/ajpgi.1999.276.5.G1069. [DOI] [PubMed] [Google Scholar]
- 61.Grisham MB, Jourd'Heuil D, Wink DA. Nitric oxide. I. Physiological chemistry of nitric oxide and its metabolites: implications in inflammation. Am J Physiol. 1999;276:G315–G321. doi: 10.1152/ajpgi.1999.276.2.G315. [DOI] [PubMed] [Google Scholar]
- 62.Heigold S, Sers C, Bechtel W, Ivanovas B, Schäfer R, Bauer G. Nitric oxide mediates apoptosis induction selectively in transformed fibroblasts compared to nontransformed fibroblasts. Carcinogenesis. 2002;23:929–941. doi: 10.1093/carcin/23.6.929. [DOI] [PubMed] [Google Scholar]
- 63.Sharara AI, Perkins DJ, Misukonis MA, Chan SU, Dominitz JA, Weinberg JB. Interferon (IFN)-alpha activation of human blood mononuclear cells in vitro and in vivo for nitric oxide synthase (NOS) type 2 mRNA and protein expression: possible relationship of induced NOS2 to the anti-hepatitis C effects of IFN-alpha in vivo. J Exp Med. 1997;186:1495–1502. doi: 10.1084/jem.186.9.1495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hokari A, Zeniya M, Esumi H, Ishikawa T, Kurasima Y, Toda G. Role of nitric oxide (NO) in interferon-alpha therapy for hepatitis C. J Infect. 2005;51:47–53. doi: 10.1016/j.jinf.2004.08.025. [DOI] [PubMed] [Google Scholar]
- 65.Saile B, Eisenbach C, El-Armouche H, Neubauer K, Ramadori G. Antiapoptotic effect of interferon-alpha on hepatic stellate cells (HSC): a novel pathway of IFN-alpha signal transduction via Janus kinase 2 (JAK2) and caspase-8. Eur J Cell Biol. 2003;82:31–41. doi: 10.1078/0171-9335-00285. [DOI] [PubMed] [Google Scholar]
- 66.Dooley S, Hamzavi J, Breitkopf K, Wiercinska E, Said HM, Lorenzen J, Ten Dijke P, Gressner AM. Smad7 prevents activation of hepatic stellate cells and liver fibrosis in rats. Gastroenterology. 2003;125:178–191. doi: 10.1016/s0016-5085(03)00666-8. [DOI] [PubMed] [Google Scholar]
- 67.Borkham-Kamphorst E, Stoll D, Gressner AM, Weiskirchen R. Antisense strategy against PDGF B-chain proves effective in preventing experimental liver fibrogenesis. Biochem Biophys Res Commun. 2004;321:413–423. doi: 10.1016/j.bbrc.2004.06.153. [DOI] [PubMed] [Google Scholar]
- 68.Breitkopf K, Haas S, Wiercinska E, Singer MV, Dooley S. Anti-TGF-beta strategies for the treatment of chronic liver disease. Alcohol Clin Exp Res. 2005;29:121S–131S. doi: 10.1097/01.alc.0000189284.98684.22. [DOI] [PubMed] [Google Scholar]
- 69.Flanders KC. Smad3 as a mediator of the fibrotic response. Int J Exp Pathol. 2004;85:47–64. doi: 10.1111/j.0959-9673.2004.00377.x. [DOI] [PMC free article] [PubMed] [Google Scholar]