

Abstract
Analysis of an intrinsically disordered protein (IDP) reveals an underlying multifunnel structure for the energy landscape. We suggest that such ‘intrinsically disordered’ landscapes, with a number of very different competing low-energy structures, are likely to characterise IDPs, and provide a useful way to address their properties. In particular, IDPs are present in many cellular protein interaction networks, and several questions arise regarding how they bind to partners. Are conformations resembling the bound structure selected for binding, or does further folding occur on binding the partner in a induced-fit fashion? We focus on the p53 upregulated modulator of apoptosis (PUMA) protein, which adopts an  -helical conformation when bound to its partner, and is involved in the activation of apoptosis. Recent experimental evidence shows that folding is not necessary for binding, and supports an induced-fit mechanism. Using a variety of computational approaches we deduce the molecular mechanism behind the instability of the PUMA peptide as a helix in isolation. We find significant barriers between partially folded states and the helix. Our results show that the favoured conformations are molten-globule like, stabilised by charged and hydrophobic contacts, with structures resembling the bound state relatively unpopulated in equilibrium.
-helical conformation when bound to its partner, and is involved in the activation of apoptosis. Recent experimental evidence shows that folding is not necessary for binding, and supports an induced-fit mechanism. Using a variety of computational approaches we deduce the molecular mechanism behind the instability of the PUMA peptide as a helix in isolation. We find significant barriers between partially folded states and the helix. Our results show that the favoured conformations are molten-globule like, stabilised by charged and hydrophobic contacts, with structures resembling the bound state relatively unpopulated in equilibrium.
Intrinsically disordered proteins (IDPs) defy the textbook structure-function paradigm, according to which a protein folds into a single and unique 3D structure to accomplish its physiological function. With their flexibility and inherent plasticity, IDPs play an important role in protein-protein interactions1 in many cellular processes, such as signal transduction and gene expression2, and therefore represent an important challenge in structural biology3. The presence of intrinsically disordered regions in cancer-associated proteins has highlighted their implication in human diseases4, as exemplified by p53 5 and HPV6. Amyloid aggregation and pathological assembly of IDPs also characterise neurodegenerative diseases, for example, the A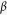 peptide in Alzheimer’s7.
peptide in Alzheimer’s7.
Although unsuitable for high resolution X-ray crystallography, increasing efforts have been invested in the last decade in many other experimental techniques to characterise the conformational ensemble of IDPs, such as NMR8,9, SAXS5,10, and mass spectrometry11.
As major components of protein-protein interaction networks, the disordered nature of an IDP is advantageous in many ways12, including fast association with alternative partners, highlighting the prevalence of IDPs in cell signalling pathways. From a structural point of view, disordered proteins are noteworthy in undergoing a disorder-to-order transition upon binding to a partner, referred to as the ‘coupled folding and binding mechanism’13,14. This binding mechanism rules out a lock-and-key scenario, highlighting a key question: does binding occur through conformational selection or induced-fit? In the first case the binding partner selects a conformation closely related to the IDP-bound conformation, although not necessarily most populated in the unbound ensemble. In the latter case, the binding partner induces structure and folding of the disordered protein upon contact. These two scenarios are not mutually exclusive, but could be system dependent, and molecular simulation has proved to be a crucial tool in this challenging field. Monte Carlo simulations have recently shown how two different IDPs bind to the same partner in a similar manner15. Multicanonical molecular dynamics simulations were performed to enhance sampling, and identified a cooperative induced-fit and conformational selection procedure for the binding of a 15-residue IDP16. A sophisticated multistate G -model was recently applied to show that the NCBD disordered protein binds two different partners, mainly through an induced-fit recognition mechanism, although the authors do not exclude an alternative conformational selection pathway for binding17.
-model was recently applied to show that the NCBD disordered protein binds two different partners, mainly through an induced-fit recognition mechanism, although the authors do not exclude an alternative conformational selection pathway for binding17.
Here we focus on the PUMA protein (p53 upregulated modulator of apoptosis), belonging to the Bcl-2 homology 3 (BH3)-only subclass of proteins, and leading to the activation of apoptosis when bound to its partner the antiapoptotic protein Mcl-118. The region of PUMA that binds Mcl-1 is called the BH3 region; this motif is intrinsically disordered and adopts a helical structure when bound to its globular folded partner19. Recent biophysical studies have demonstrated that PUMA associates rapidly and tightly to its partner Mcl-120. Upon introducing proline substitutions in the peptide sequence21 (thus breaking helical structure), association rate constants with Mcl-1 are mainly unaffected, suggesting that particular residual helical structure does not seem to be essential for binding. Using rapid-mixing stopped flow, Rogers and coworkers22 have recently shown that neither folding nor specific interactions are required for association, suggesting that conformational selection for binding is unlikely. Thus, it seems that the IDP PUMA binds its partner in an induced-fit fashion. From a structural perspective, why then isn’t the alpha-helical conformation of PUMA stable in isolation?
To answer this question, we employ an array of computational methods, ranging from molecular dynamics to creation of a kinetic transition network23,24,25 using geometry optimisation. Our results show that indeed isolated PUMA is not stable as a contiguous  -helix and the conformational ensemble is populated by structures with mainly low to medium helicity content, in agreement with experiment20. Visualizing the energy landscape of PUMA in isolation reveals a frustrated26,27 multifunneled structure with no dominant energy minimum and molten globule-like structures at the bottom of the funnels. Most importantly, we show that (i) the barriers to folding for an
-helix and the conformational ensemble is populated by structures with mainly low to medium helicity content, in agreement with experiment20. Visualizing the energy landscape of PUMA in isolation reveals a frustrated26,27 multifunneled structure with no dominant energy minimum and molten globule-like structures at the bottom of the funnels. Most importantly, we show that (i) the barriers to folding for an  -helix from the most populated conformations are large, (ii) unfolding is orders of magnitude faster than folding, and (iii) the driving forces to unfolding of the contiguous helix are mainly electrostatic in nature, but not necessarily residue-specific. These results help to explain the instability of this IDP when isolated, as well as the induced fit mechanism for binding determined by experiment22.
-helix from the most populated conformations are large, (ii) unfolding is orders of magnitude faster than folding, and (iii) the driving forces to unfolding of the contiguous helix are mainly electrostatic in nature, but not necessarily residue-specific. These results help to explain the instability of this IDP when isolated, as well as the induced fit mechanism for binding determined by experiment22.
Diverse electrostatic and hydrophobic interactions are formed within the partially folded structures of PUMA, leading to the presence of the multiple partially folded conformations, and form the basis for the intrinsically disordered behaviour and the corresponding ‘intrinsically disordered energy landscape’, which seems likely to characterise such proteins. This structure provides a useful way to address emergent observable properties from the energy landscape perspective.
Results
The
 -helical conformation of the PUMA peptide unfolds in isolation
-helical conformation of the PUMA peptide unfolds in isolation
To obtain some initial insight into the 32-residue PUMA peptide when isolated, we performed 600 ns molecular dynamics simulations starting from the contiguous helical structure at 280 and 300 K in explicit solvent (Fig. 1). At 280 K, the percentage of residual helicity reaches 40% after 15 ns and fluctuates between 40 and 80% over a time scale of 200 ns. At 300 K, the helicity rapidly decreases to 35% after 20 ns. Over the last 100 ns of the MD simulations, the average helicity values are 32 and 30% at 280 and 300 K, respectively (Fig. 1A,B). To monitor the similarity to the contiguous helical PUMA, we calculated the backbone root mean square deviation (RMSD) and our results show a rapid increase to more than 3 Å after only 17 ns at 280 K. At 600 ns, we observe relatively large values for the RMSD, ranging from 10 to 12 Å in all simulations, following the trends of helical percentage (Fig. 1C). In each case, the structures become more collapsed, and their radius of gyration decreases from about 16 Å to around 10 Å (Supplementary Figure 1A). This increase in the compaction of the isolated protein is evident from calculating distances between two residues located at the N- and C-terminal regions, which decrease from high (over 35 Å) to low (under 5 Å) values (Supplementary Figure 1B). From these results it is clear that the helix is not stable by itself, consistent with the induced-fit scenario identified experimentally20,21,22.
Figure 1.
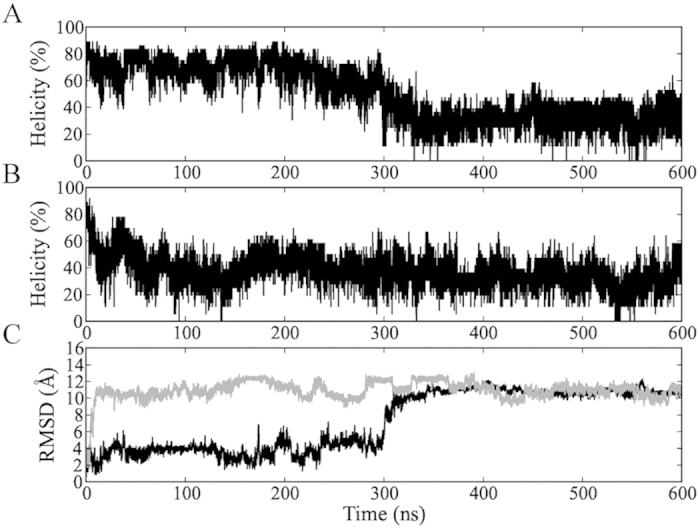
(A, B) The percentage of residual helicity and (C) backbone RMSD with respect to the minimised  -helix conformation of PUMA, where the MD starting points are the helix at 280 (A and black lines in C) and 300 K (B and grey lines in C).
-helix conformation of PUMA, where the MD starting points are the helix at 280 (A and black lines in C) and 300 K (B and grey lines in C).
Conventional molecular dynamics simulations are prone to kinetic trapping, and to enhance the conformational sampling of the helical PUMA peptide in isolation, we performed 140 ns of replica exchange molecular dynamics (REMD)28 using 20 temperature replicas ranging from 223.7 to 650 K in implicit solvent, resulting in a total simulation time of 2.8  s. The first 30 ns of the simulation were excluded from the analysis to allow for equilibration. REMD simulations were deemed to have converged when the heat capacity and residual helicity values calculated in the time intervals 30 to 85 and 85 to 140 ns showed similar behaviour (Supplementary Figure 2). A heat capacity peak appears at 313 K, where the residual helicity is about 24%.
s. The first 30 ns of the simulation were excluded from the analysis to allow for equilibration. REMD simulations were deemed to have converged when the heat capacity and residual helicity values calculated in the time intervals 30 to 85 and 85 to 140 ns showed similar behaviour (Supplementary Figure 2). A heat capacity peak appears at 313 K, where the residual helicity is about 24%.
At 280 K the average helicity in our simulations is 27.8% and that calculated with the AGADIR predictor29, used by Rogers and coworkers to design point mutations in the PUMA sequence21, is 28%. These values are consistent with CD experiments suggesting about 20% helicity at 298 K20. Calibrating simulation temperatures and experimental ones is a difficult task, especially when force fields are used with an implicit solvent representation. The temperatures used for this comparison are not exactly the same, but we believe that 280 K approximates best the experimental room temperature, in view of the heat capacity curve obtained from our simulations (Supplementary Figure 2) suggesting that at 298 K the system is too close to the melting regime. Although slightly over-estimating the helicity with respect to experiment, the force field reproduces the experimental range of values with no significant differences. These results are supported by the comparison of different force fields using NMR scalar coupling data, which suggest that the AMBER representation we have used has the best agreement with experimental data among those tested30. We calculated the helical propensities at a residue level at 280 K for the last 110 ns of the REMD simulations and compared the values obtained with AGADIR29 (Fig. 2). Overall, the two profiles agree in the general trends of helicity, where the highest helical content is present mainly in the C-terminal region. The largest discrepancies appear at positions A150 and Q151, but the  -helical percentages for these residues are still elevated (
-helical percentages for these residues are still elevated ( 30%).
30%).
Figure 2.
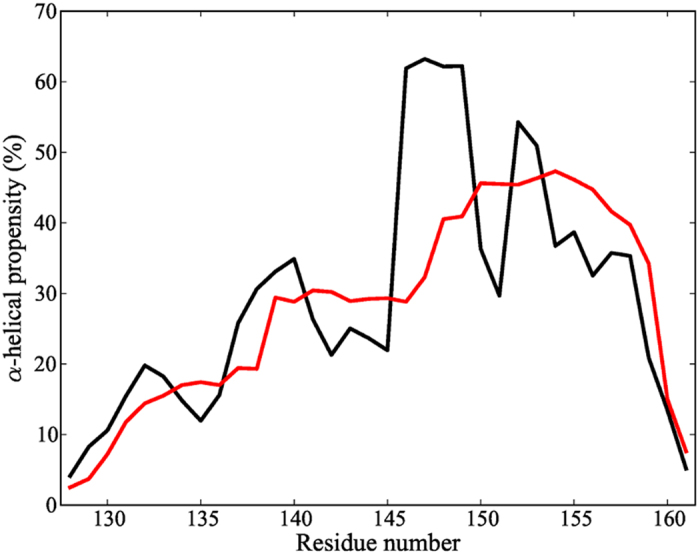
Helical propensities at the residue level for the PUMA sequence calculated from REMD simulations (black lines) and from the AGADIR29 helical propensity predictor (red lines).
A free energy surface for the PUMA peptide was constructed at 280 K, using as order parameters the fraction of residual helicity and the radius of gyration (Fig. 3). The lowest free energy minimum is rather broad and corresponds to a range of fractional helicity between 0.2 and 0.4, with a radius of gyration of about 10 Å. The other low-lying free energy minimum corresponds to a more extended structure, with a radius of gyration of around 12 Å, but covering the same range of helicity. It is important to note that regions with residual helicity greater than 0.6 exhibit relatively high free energies, and correspond to values around 15 to 16 Å for the radius of gyration (close to the contiguous helix), confirming that the helical structure of the PUMA peptide is unfavourable.
Figure 3.
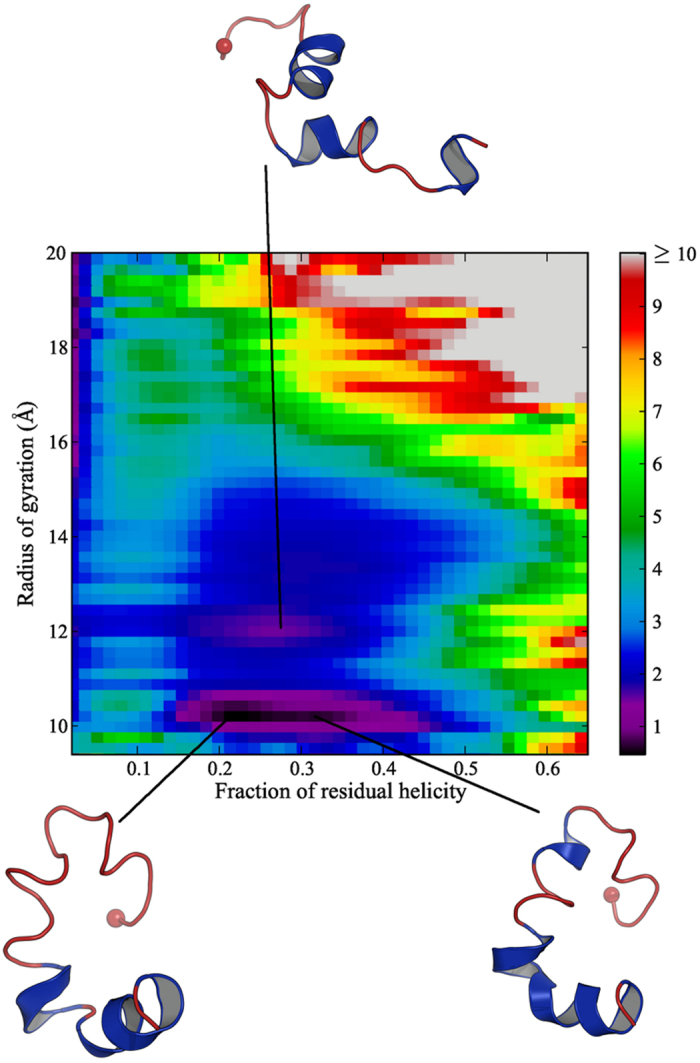
Free energy surface at 280 K for the PUMA peptide in terms of fraction of residual helicity and radius of gyration. The free energy scale in kcal/mol is shown on the right. Typical conformations for the minima are represented, where the Cα of the N-terminal residue Val128 is indicated by a sphere.
Visualising the conformational ensemble of the IDP using discrete path sampling
We used the discrete path sampling (DPS) approach31 to analyse the underlying potential energy landscape for PUMA. The DPS technique is based on geometry optimisation and efficiently produces stationary point databases corresponding to a kinetic transition network23,24,25. To extract relevant partially folded PUMA structures, clustering analysis was performed on the trajectory at 280 K, based on dihedral angles, which groups structures that display similar secondary structure content. We considered the ten most populated structures at 280 K, which in total account for 45% of the analysed simulation time frames, as starting points for building a connected database, together with the helical structure of PUMA. The combined stationary point database from DPS calculations is visualised using a potential energy disconnectivity graph32,33 in Fig. 4 and consists of a total of 205,677 minima and 191,392 transition states. The colouring in this figure corresponds to the percentage of helicity for each minimum in the database. At the bottom of the funnels, the helicity percentages are between 20 and 30%; the lowest and highest helicity values are not favourable, and such structures are located at branches near the top of the graph. Frustration26,27 in the disconnectivity graph corresponds to low-lying morphologies separated by high barriers, and illustrates the diversity of competing conformations, with no single dominant structure. The corresponding free energy disconnectivity graph is illustrated in Supplementary Figure 3, and it displays the same structure as the potential energy landscape for the relevant temperature range. The observed difference between the 2D free energy surface calculated from the REMD simulations and the disconnectivity graph obtained using the DPS technique occurs because some partially folded states are incorrectly lumped together on projection to obtain a low-dimensional representation. Such projections of configuration space do not generally preserve the kinetics, often producing artifically smooth surfaces that do not reflect the barriers properly, thus masking the complexity of the landscape25,34,35,36. In contrast, a kinetic transition network can faithfully represent the barriers25. Here we note that for complex biomolecular pathways a suitable progress coordinate may not exist. Such conformational transitions are therefore ideal targets for DPS, which only requires specification of product and reactant states.
Figure 4.

Potential energy disconnectivity graph constructed from the most populated structures obtained in the REMD simulations at 280 K. The colouring of the branches corresponds to the  -helical percentage calculated for each minimum in the database, as defined in the key.
-helical percentage calculated for each minimum in the database, as defined in the key.
The multifunnel structure seems likely to be associated with the intrinsically disordered nature of the protein. Hence the disconnectivity graph representation provides a visual explanation for the induced fit mechanism of binding. In particular, it is clear that the contiguous helix conformation of PUMA (i) does not occupy a low energy funnel and (ii) the barriers to unfolding are much lower than the ones to fold into the helix. In fact, the funnel corresponding to the contiguous helix is relatively narrow and underpopulated with respect to the other funnels. The most populated structure at 280 K (12.4%) corresponds to the largest funnel in terms of local minima, although it competes with low energy funnels related to the other most populated structures.
Such multifunnel energy landscapes have been extensively studied for atomic and molecular clusters37, where they provide benchmarks for global optimisation, enhanced sampling schemes to address broken ergodicity38,39,40,41, and rare event dynamics31,42,43, corresponding to changes of morphology. The competing funnels lead to multiple relaxation time scales and features in the heat capacity profile25, and correspond to glassy behaviour in systems with an exponentially large number of low-lying amorphous states44. Consistent with the present results, a multifunneled energy landscape has previously been characterised for an amyloid peptide derived from the disordered domain of the yeast prion protein Sup35. As for PUMA, this system exhibits competition between alternative conformations, which are 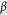 -sheet structures for the amyloid45. Hence, we suggest that IDPs are likely to correspond to intrinsically disordered, multifunneled energy landscapes. We propose to test this hypothesis for other IDPs in future work.
-sheet structures for the amyloid45. Hence, we suggest that IDPs are likely to correspond to intrinsically disordered, multifunneled energy landscapes. We propose to test this hypothesis for other IDPs in future work.
We calculated the phenomenological rate constants for the unfolding and folding from the contiguous helix (conformation A) and the most populated structure at 280 K (conformation B), using graph transformation46. The comparison of these rates indicate that folding is slow with respect to unfolding, indeed the estimate for 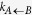 is 0.27 × 10−20 s−1 (probably a weak lower bound) whereas
is 0.27 × 10−20 s−1 (probably a weak lower bound) whereas 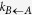 is over 10.5 s−1. We also calculated the rates for conversion of the helix to the other most populated structures (Table 1). These transitions are also several orders of magnitude faster than the predicted folding rates of each of these structures to the helix. The numerical values cannot be compared directly with binding rate constants, since the partner MCL-1 is absent in these calculations, and it is more appropriate to interpret the relative rather than the absolute rates. The particularly slow folding rate obtained for the helical conformation supports the lack of evidence for a conformational selection for binding22 and suggests that the presence of the partner may accelerate this transition.
is over 10.5 s−1. We also calculated the rates for conversion of the helix to the other most populated structures (Table 1). These transitions are also several orders of magnitude faster than the predicted folding rates of each of these structures to the helix. The numerical values cannot be compared directly with binding rate constants, since the partner MCL-1 is absent in these calculations, and it is more appropriate to interpret the relative rather than the absolute rates. The particularly slow folding rate obtained for the helical conformation supports the lack of evidence for a conformational selection for binding22 and suggests that the presence of the partner may accelerate this transition.
Table 1. Estimated rate contants (in s−1) at 280 K for the unfolding and folding from the contiguous
 -helical conformation of the PUMA peptide to each of the 10 most populated structures from the REMD simulations, and their respective populations. The values for folding should be considered as lower bounds.
-helical conformation of the PUMA peptide to each of the 10 most populated structures from the REMD simulations, and their respective populations. The values for folding should be considered as lower bounds.
Rate constants between the initial partially folded minima used to build the energy landscape of PUMA were calculated using the same procedure (Supplementary Table S1). Apart from the first two conformations, which are similar in structure, (Cα-RMSD of 1.7 Å), the transitions are relatively slow between the partially helical states. Although no experimental results for these interconversion rates between partially folded states for the free PUMA protein have been reported so far, it is known that conformational fluctuations in IDPs can be slowed by the presence of residual structure47. Indeed, experimental studies on an analogue of the BPTI show slow interconversion rates between two partially folded conformations with different degrees of order48. NMR relaxation experiments for the compact molten globule NCBD domain of transcription factor CBP reported the presence of slow conformational fluctuations in this IDP49. A theoretical study on an archetypal IDP sequence revealed slow interconversion between distinct conformations of the peptide in water50.
To understand the molecular mechanism of unfolding, we calculated the electrostatic and van der Waals components of the potential energy for the successive minimum-transition state-minimum stationary points in the discrete path31 leading from the helix to the most populated structure at 280 K (Fig. 5). This pathway is mainly downhill in energy, and the most important forces driving the unfolding are electrostatic (Fig. 5B), with a gain of approximately 400 kcal/mol on forming a more globular structure. The corresponding change in the van der Waals energy is negligible in comparison, with only about 10 kcal/mol differece between the two end points (Fig. 5C). Hence it seems that electrostatics drive the unfolding and determine the instability of the contiguous helical form.
Figure 5.
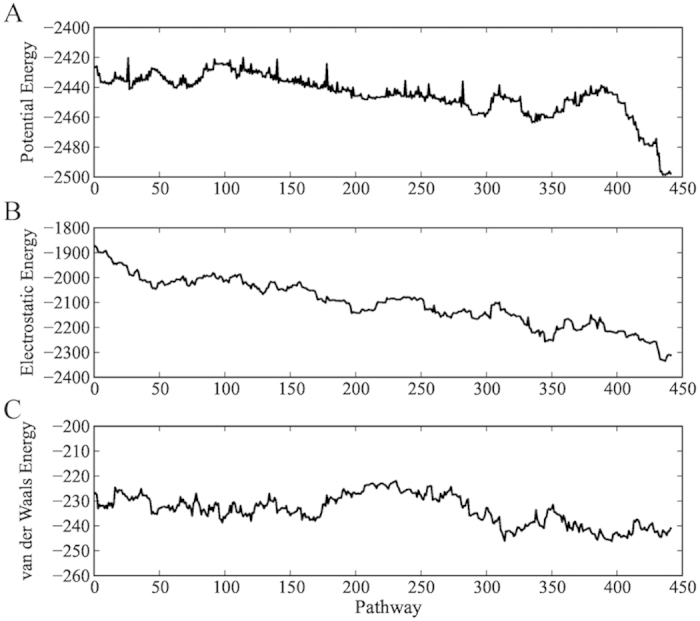
Energies in kcal/mol along the fastest path for the transition between the contiguous helical conformation of PUMA and the partially folded structure corresponding to the most populated conformation at 280 K. The potential energy (A) of each starting point is decomposed into (B) electrostatic and (C) van der Waals components.
Structural characterisation of the partially folded ensemble
To provide a more detailed understanding of the electrostatic characteristics of PUMA at the residue level, we first analysed the hydrogen-bond network, defined using a distance cutoff of 3 Å and a maximum deviation from a linear angle of 40°. All the hydrogen bonds present for more than 40% of the simulation time considered for the analysis are associated with side chains of charged residues. The four most prevalent hydrogen bonds involve residues D146 with R154 and R155, R142 and E132, and R143 and E159. Looking at the positions of these charged residues in the structure of the most populated cluster (Fig. 6A), we see that they are located around the center of the U-shaped PUMA conformation.
Figure 6.

(A) Most populated structure at 280 K, where the hydrogen bonds with an occurrence greater than 50% of the simulation time are included for the analysis. Residues implicated in the bonds are represented by sticks, the N-terminal Cα of the PUMA sequence is indicated by a sphere. (B) Contact maps calculated for the structures for the four most populated clusters.
We then performed a structural analysis of the PUMA protein and calculated contact maps for the structures in the four most populated clusters (Fig. 6B). Interactions between charged and hydrophobic residues are observed. For example, the strongest hydrophobic contacts involve I137, G138 and A139 with L141, and M144 with L141. Significant interactions between charged residues occur for E132-R135, R143-D146 and R156-E158. These hydrophobic and charged interactions are also the most important ones observed in the contact map calculated for all the structures observed at 280 K (Supplementary Figure 4), where the number of the most populated hydrophobic and charged contacts lie within the same sequence ranges of 8 and 11 contacts, respectively. However, in the most populated cluster, it seems that the charged interactions are more prevalent than the hydrophobic ones, with 24 most populated compared to 10. Although some contacts are similar, different interaction patterns are observed for each cluster (Fig. 6B). This result could explain the diversity in the PUMA structures and hence the absence of one dominant single fold for the protein in the absence of binding partners, as suggested when looking at the population of each cluster at the same temperature (Table 1) and by the multifunneled character of the energy landscape.
Mutations of charged residues to alanine (for instance R143) do not destabilize the binding of PUMA to MCL-121. Although we cannot infer any details of the binding process, we can suggest explanations for this observation. It is conceivable that the disruption of just one charged residue does not perturb the electrostatic network much (Fig. 6A), as other charged residues may replace the missing contributions.
To characterise the effect of this substitution, we mutated one of the residues, R143, involved in the electrostatic network into an alanine in the structure of the most populated cluster at 280 K. The mutated structure was considered as a starting point for basin-hopping global optimisation using the GMIN program51. Here, we used rotation of side chain groups to explore the configuration space and hence investigate the effects of this mutation on the electrostatic network of the wild-type geometry. Overall, the resulting structures display the same pattern of charged interactions. The RMSD calculated using carbon atoms of the side chains is 0.6 and 1 Å for the six residues discussed previously (E132, R142, D146, R154, R155, E159) in the R143A mutant. This result again reflects the non-specific character of the electrostatic network formed within the PUMA protein.
Analysis of amino acid composition for IDPs has indeed shown that they typically contain numerous charged residues52,53, and the alternative electrostatic networks that can be formed in addition to hydrophobic contacts may contribute to the variety of molten-globule-like conformations observed in the PUMA protein when unbound, and hence its intrinsically disordered nature. Hydrophobic contacts could explain the overall preservation of helicity when varying the ionic strength, as determined experimentally20. This globule-like phase in the conformational space of IDPs has been linked to the net charge per residue54 and the charge patterning in the sequence55. In strong polyampholytic IDPs, with an elevated fraction of charged residues as for PUMA, the distribution of the charges in the sequence will influence the conformational properties of the protein. Das and Pappu55 have quantified this effect using a patterning variable termed  , which ranges from 0 to 1, for a very symmetric distribution of charges and thus hairpin-like structures due to long-range electrostatic interactions between residues. For PUMA,
, which ranges from 0 to 1, for a very symmetric distribution of charges and thus hairpin-like structures due to long-range electrostatic interactions between residues. For PUMA,  is about 0.18 according to the webserver CIDER56, which correlates well with the partially folded states identified in our simulations.
is about 0.18 according to the webserver CIDER56, which correlates well with the partially folded states identified in our simulations.
Discussion
Experimental evidence regarding the binding of the intrinsically disorered protein PUMA to its partner, involved in subsequent activation of apoptosis in cells, suggests an induced fit mechanism21,22. These recent results show that no specific residual helicity or particular fold seem to be required for binding. In the present contribution, we have used a combination of computational methods to understand the behaviour in the absence of the binding partner to explain the induced fit mechanism. Our results show that the contiguous helical conformation of the protein is unstable, and the protein conformation ensemble is mostly molten globule with low to medium residual helicity, in agreement with CD results20. Most importantly, our aim was to understand the molecular mechanism that drives the destabilisation of the helix in isolation.
Visualisation of the corresponding energy landscape reveals a multifunnel structure, with a number of alternative low-lying morphologies separated by high energy barriers. The funnel representing the helical structure is clearly unfavourable with respect to the partially folded structures, explaining why conformational selection for this protein is unlikely and has not been observed experimentally22. The calculated rate constants corresponding to the transition from the PUMA helix to partially folded molten-globule-like states indicate a fast unfolding relative to folding, governed mainly by electrostatic forces, while the reverse process is predicted to be much slower. We have also shown that interactions between both charged and hydrophobic residues contribute to the stability of the partially folded states, and thus to the structure of the landscape and its emergent properties.
Mutations of single charged residues into alanine do not perturb either association or dissociation of PUMA to its partner MCL-121, and we believe the reason is that the electrostatic network involves several charged amino acids, so mutation of one of these components to alanine can be compensated by another nearby charged residue.
We suggest that the multiple funnel structure is likely to characterise the landscapes of intrinsically disordered proteins, and this hypothesis will be tested in future work. The characterisation of such intrinsically disordered energy landscapes may provide a general approach to understand and calculate the properties of such systems.
Methods
Structure preparation
To compare our results with the studies of Rogers and coworkers20, we chose to work with the sequence 128-161 of the PUMA protein (Uniprot Q99ML1). The  -helical structure of the peptide was built using the PyMol software and is very similar to the NMR structure of the peptide (residues 130-156) in the bound state with MCL-1 (pdb id: 2ROC)19, with an RMSD of 1 Å. Molecular dynamics simulations were carried out using the AMBER99SB57 force field via the AMBER12 package58. NMR structural and relaxation data of the A
-helical structure of the peptide was built using the PyMol software and is very similar to the NMR structure of the peptide (residues 130-156) in the bound state with MCL-1 (pdb id: 2ROC)19, with an RMSD of 1 Å. Molecular dynamics simulations were carried out using the AMBER99SB57 force field via the AMBER12 package58. NMR structural and relaxation data of the A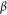 peptide were obtained using this force field in good agreement with experimental data, justifying its use for IDPs59. The N-terminus and the C-terminus were acetylated and amidated respectively. To eliminate steric clashes between atoms, a minimisation consisting of 500 steepest-descent steps was performed, followed by 500 steps of conjugate gradient, until the root mean square (RMS) gradient of the potential energy reached 10−4 kcal mol−1Å−1.
peptide were obtained using this force field in good agreement with experimental data, justifying its use for IDPs59. The N-terminus and the C-terminus were acetylated and amidated respectively. To eliminate steric clashes between atoms, a minimisation consisting of 500 steepest-descent steps was performed, followed by 500 steps of conjugate gradient, until the root mean square (RMS) gradient of the potential energy reached 10−4 kcal mol−1Å−1.
Dynamics Simulations in Explicit Solvent
The initial helical structure was solvated with TIP3P60 water molecules in a box of dimension 330 nm3, with periodic boundary conditions applied for all molecular dynamics (MD) simulations. The SHAKE algorithm61 was applied to constrain bonds involving hydrogen atoms, and a time step of 2 fs was applied. The temperatures used in the MD simulations were 280 and 300 K, and a Langevin thermostat62 was used with a collision frequency of 2 ps−1. An initial minimisation with position restraints using a force constant of 500 kcal mol−1Å−2 on the PUMA protein was performed using 500 and 1,500 steepest-descent and conjugate gradient steps, respectively, in order to locally equilibrate the water and ions. Another minimisation was performed without any restraints on the protein for 1,000 steepest-descent steps followed by 3,000 conjugate gradient steps. Both minimisations were continued until the RMS gradient of the potential energy reached 10−4 kcal mol−1Å−1. Next, the solvent was heated to the required temperature for 20 ps with moderate restraints on the protein using a force constant of 10 kcal mol−1 Å−2. To allow the solvent density to equilibrate, a 500 ps NPT simulation was then performed with isotropic position scaling to a reference pressure of 1 bar. Finally, the production simulations using NVT conditions were run for a total of 600 ns at each temperature.
Replica Exchange Molecular Dynamics
Replica exchange molecular dynamics (REMD)28 simulations were performed using the relaxed conformation of PUMA with 20 replicas at temperatures ranging from 223.7 to 650 K. The generalised Born model was used for an implicit solvent representation63. Temperatures were controlled using a Langevin thermostat with a collision frequency of 2.0 ps−1. The maximum interatomic distance for computing the effective Born radii was set to 25 Å, and no truncation was applied to the nonbonded interactions. Each simulation was equilibrated for 400 ps to reach the selected temperature for the REMD simulations. The production run consisted of a total of 140 ns for each replica, resulting in a total simulation time of 2.8  s. Exchanges were attempted every ps leading to an acceptance ratio of approximately 22%. The analysis was performed using the ptraj tools in the AMBER program, excluding the first 30 ns of the simulation for convergence purposes. The multistate Bennett acceptance ratio method (MBAR) of Shirts and Chodera64, a binless-equivalent to the weighted histogram analysis method65, was used to analyse the REMD trajectories. Clustering of the trajectories was performed on dihedral angles using the cluster tool in the MMTSB toolset66 with an angle cutoff of 40º. Secondary structure analysis was performed using the DSSP implementation in the AMBER trajectory analysis tools67.
s. Exchanges were attempted every ps leading to an acceptance ratio of approximately 22%. The analysis was performed using the ptraj tools in the AMBER program, excluding the first 30 ns of the simulation for convergence purposes. The multistate Bennett acceptance ratio method (MBAR) of Shirts and Chodera64, a binless-equivalent to the weighted histogram analysis method65, was used to analyse the REMD trajectories. Clustering of the trajectories was performed on dihedral angles using the cluster tool in the MMTSB toolset66 with an angle cutoff of 40º. Secondary structure analysis was performed using the DSSP implementation in the AMBER trajectory analysis tools67.
Exploring the potential energy landscape with basin-hopping
Exploration of the potential energy landscape was performed largely via basin-hopping68,51, an efficient strategy for global optimisation. The method consists of trial perturbations in configuration space, each followed by local energy minimisations, resulting in a set of configurational minima on the potential energy landscape. The acceptance criteria for a given trial move are determined by the energy difference between minima, as well as a user-specified temperature parameter. Because detailed balance is not required, the method affords great flexibility in choosing step sizes and temperature parameters that optimise sampling. As a result, basin-hopping can rapidly explore the potential energy landscape.
Basin-hopping was performed for the PUMA mutation R143A with 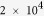 steps. The trial moves consisted of Cartesian displacements, pivot moves (i.e. rotations of the amino acid chain about a backbone dihedral angle), and rotations of R-group segments about various R-group bonds.
steps. The trial moves consisted of Cartesian displacements, pivot moves (i.e. rotations of the amino acid chain about a backbone dihedral angle), and rotations of R-group segments about various R-group bonds.
Discrete path sampling
After clustering the REMD trajectory at the desired temperature, pathways connecting the 10 most populated clusters of PUMA were calculated, along with the contiguous helical conformation of the protein. Transition state candidates were produced as initial guesses using the doubly-nudged69 elastic band algorithm70. These candidates were tightly converged using a hybrid eigenvector-following approach71. Local minima were obtained using a modified limited-memory Broyden-Fletcher-Golgfarb-Shano (L-BFGS) algorithm72. Once an initial connected path between the chosen end points was found, the stationary point database was grown by addition of all minima and transition states identified in successive connection attempts for pairs of minima already existing in the database73. This approach, discrete path sampling (DPS)31, is implemented in the program PATHSAMPLE, which organises parallel connection attempts using the OPTIM program.
The rate constant between the two selected end points can be expressed as the sum over all discrete paths between product and reactant states31,74. The discrete paths that make the largest contribution to this rate constant were calculated using the Dijkstra algorithm, and the highest potential energy barriers on this path were identified. The procedure SHORTCUT BARRIER45 implemented in PATHSAMPLE was used to refine the database, until the rate constants converged to within an order of magnitude. Artificial kinetic traps were eliminated using the UNTRAP procedure45, which attempts to connect minima close in distance to the product set but separated by high barriers. The resulting database consisted of 205,677 minima and 191,392 transition states and was visualised using disconnectivity graphs32,33. The database of minima obtained from the DPS method was used to calculate free energies using the harmonic superposition approximation75. Phenomenological two-state rate constants, denoted kAB and kBA for the transitions from B to A and A to B respectively, were calculated using the graph transformation approach74,76 from the complete database after regrouping self-consistently structures that were separated by free energy barriers below a certain threshold77 (3 kcal/mol in this work).
Author Contributions
Y.C., A.J.B. and D.C. performed the simulations and analyzed the data. Y.C., A.J.B., D.C. and D.J.W. discussed the results and wrote the manuscript.
Additional Information
How to cite this article: Chebaro, Y. et al. Intrinsically Disordered Energy Landscapes. Sci. Rep. 5, 10386; doi: 10.1038/srep10386 (2015).
Supplementary Material
Acknowledgments
The authors thank Prof. Jane Clarke, Dr. Chris Whittleston, Dr. Joanne Carr, Dr. Iskra Staneva and Dr. David de Sancho for helpful discussions. Y.C. and A.J.B. acknowledge funding from the EPSRC grant number EP/I001352/1, D.C. gratefully acknowledges the Cambridge Commonwealth European and International Trust for financial support and D.J.W. the ERC for an Advanced Grant.
References
- Dosztányi Z., Chen J., Dunker A. K., Simon I. & Tompa P. Disorder and sequence repeats in hub proteins and their implications for network evolution. J. Proteome. Res. 5, 2985–2995 (2006). [DOI] [PubMed] [Google Scholar]
- Uversky V. N., Oldfield C. J. & Dunker A. K. Showing your id: intrinsic disorder as an id for recognition, regulation and cell signaling. J. Mol. Recognit. 18, 343–384 (2005). [DOI] [PubMed] [Google Scholar]
- Tompa P. Unstructural biology coming of age. Curr. Opin. Struct. Biol. 21, 419–425 (2011). [DOI] [PubMed] [Google Scholar]
- Iakoucheva L. M., Brown C. J., Lawson J. D., Obradović Z. & Dunker A. K. Intrinsic disorder in cell-signaling and cancer-associated proteins.J. Mol. Biol. 323, 573–584 (2002). [DOI] [PubMed] [Google Scholar]
- Wells M. et al. Structure of tumor suppressor p53 and its intrinsically disordered n-terminal transactivation domain. Proc. Natl. Acad. Sci. USA. 105, 5762–5767 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uversky V. N., Roman A., Oldfield C. J. & Dunker A. K. Protein intrinsic disorder and human papillomaviruses: increased amount of disorder in e6 and e7 oncoproteins from high risk hpvs. J. Proteome. Res. 5, 1829–1842 (2006). [DOI] [PubMed] [Google Scholar]
- Knowles T. P. J., Vendruscolo M. & Dobson C. M. The amyloid state and its association with protein misfolding diseases. Nat. Rev. Mol. Cell. Biol. 15, 384–396 (2014). [DOI] [PubMed] [Google Scholar]
- Jensen M. R., Ruigrok R. W. H. & Blackledge M. Describing intrinsically disordered proteins at atomic resolution by NMR. Curr. Opin. Struct. Biol. 23, 426–435 (2013). [DOI] [PubMed] [Google Scholar]
- Camilloni C., Simone A. D., Vranken W. F. & Vendruscolo M. Determination of secondary structure populations in disordered states of proteins using nuclear magnetic resonance chemical shifts. Biochemistry 51, 2224–2231 (2012). [DOI] [PubMed] [Google Scholar]
- Bernadó P. & Svergun D. I. Structural analysis of intrinsically disordered proteins by small-angle x-ray scattering. Mol. Biosyst. 8, 151–167 (2012). [DOI] [PubMed] [Google Scholar]
- Jurneczko E. et al. Intrinsic disorder in proteins: a challenge for (un)structural biology met by ion mobility-mass spectrometry. Biochem. Soc. Trans. 40, 1021–1026 (2012). [DOI] [PubMed] [Google Scholar]
- Mittag T., Kay L. E. & Forman-Kay J. D. Protein dynamics and conformational disorder in molecular recognition. J. Mol. Recognit 23, 105–116 (2010). [DOI] [PubMed] [Google Scholar]
- Haynes C. et al. Intrinsic disorder is a common feature of hub proteins from four eukaryotic interactomes. PloS. Comput Biol. 2, e100 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright P. E. & Dyson H. J. Linking folding and binding. Curr. Opin. Struct. Biol. 19, 31–38 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staneva I., Huang Y., Liu Z. & Wallin S. Binding of two intrinsically disordered peptides to a multi-specific protein: a combined monte carlo and molecular dynamics study. PloS. Comput. Biol. 8, e1002682 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higo J., Nishimura Y. & Nakamura H. A free-energy landscape for coupled folding and binding of an intrinsically disordered protein in explicit solvent from detailed all-atom computations. J. Am. Chem. Soc. 133, 10448–10458 (2011). [DOI] [PubMed] [Google Scholar]
- Knott M. & Best R. B. Discriminating binding mechanisms of an intrinsically disordered protein via a multi-state coarse-grained model. J. Chem. Phys. 140, 175102 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roulston A., Muller W. J. & Shore G. C. Bim, puma, and the achilles’ heel of oncogene addiction. Sci. Signal 6, pe12 (2013). [DOI] [PubMed] [Google Scholar]
- Day C. L. et al. Structure of the bh3 domains from the p53-inducible bh3-only proteins noxa and puma in complex with mcl-1. J. Mol. Biol. 380, 958–971 (2008). [DOI] [PubMed] [Google Scholar]
- Rogers J. M., Steward A. & Clarke J. Folding and binding of an intrinsically disordered protein: fast, but not ‘diffusion-limited’. J. Am. Chem. Soc. 135, 1415–1422 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers J. M., Wong C. T. & Clarke J. Coupled folding and binding of the disordered protein puma does not require particular residual structure. J. Am. Chem. Soc. 136, 5197–5200 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers J. M. et al. Interplay between partner and ligand facilitates the folding and binding of an intrinsically disordered protein. Proc. Natl. Acad. Sci. USA. 111, 15420–15425 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noé F. & Fischer S. Transition networks for modeling the kinetics of conformational change in macromolecules. Curr. Op. Struct. Biol. 18, 154–162 (2008). [DOI] [PubMed] [Google Scholar]
- Prada-Gracia D., Gómez-Gardenes J., Echenique P. & Fernando F. Exploring the free energy landscape: From dynamics to networks and back. PloS. Comput Biol. 5, e1000415 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wales D. J. Energy landscapes: Some new horizons . Curr. Op. Struct. Biol. 20, 3–10 (2010). [DOI] [PubMed] [Google Scholar]
- Bryngelson J. D., Onuchic J. N., Socci N. D. & Wolynes P. G. Funnels, pathways, and the energy landscape of protein folding: a synthesis. Proteins 21, 167–195 (1995). [DOI] [PubMed] [Google Scholar]
- Onuchic J. N., Luthey-Schulten Z. & Wolynes P. G. Theory of protein folding: the energy landscape perspective. Annu Rev. Phys Chem. 48, 545–600 (1997). [DOI] [PubMed] [Google Scholar]
- Sugita Y. & Okamoto Y. Replica-exchange molecular dynamics method for protein folding. Chem. Phys. Lett. 314, 141–151 (1999). [Google Scholar]
- Muñoz V. & Serrano L. Elucidating the folding problem of helical peptides using empirical parameters. Nat. Struct. Biol. 1, 399–409 (1994). [DOI] [PubMed] [Google Scholar]
- Best R. B., Buchete N.-V. & Hummer G. Are current molecular dynamics force fields too helical? Biophys. J. 95, L07–L09 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wales D. J. Discrete path sampling. Mol. Phys. 100, 3285–3306 (2002). [Google Scholar]
- Becker O. M. & Karplus M. The topology of multidimensional potential energy surfaces: Theory and application to peptide structure and kinetics. J. Chem. Phys. 106, 1495–1515 (1997). [Google Scholar]
- Wales D. J., Miller M. A. & Walsh T. R. Archetypal energy landscapes. Nature 394, 758–760 (1998). [Google Scholar]
- Krivov S. V. & Karplus M. Hidden complexity of free energy surfaces for peptide (protein) folding. Proc. Nat. Acad. Sci. USA 101, 14766–14770 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krivov S. V. & Karplus M. One-dimensional free-energy profiles of complex systems: progress variables that preserve the barriers. J Phys Chem B 110, 12689–12698 (2006). [DOI] [PubMed] [Google Scholar]
- Wales D. J. & Salamon P. Observation time scale, free-energy landscapes, and molecular symmetry. Proc. Nat. Acad. Sci. USA. 111, 617–622 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doye J. P. K., Miller M. A. & Wales D. J. The double-funnel energy landscape of the 38-atom lennard-jones cluster. J. Chem. Phys. 110, 6896–6906 (1999). [Google Scholar]
- Neirotti J. P., Calvo F., Freeman D. L. & Doll J. D. Phase changes in 38 atom lennard-jones clusters. i: A parallel tempering study in the canonical ensemble. J. Chem. Phys. 112, 10340–10349 (2000). [Google Scholar]
- Sharapov V. A. & Mandelshtam V. A. Solid-solid structural transformations in lennard-jones clusters: Accurate simulations versus the harmonic superposition approximation. J. Phys. Chem. A. 111, 10284–10291 (2007). [DOI] [PubMed] [Google Scholar]
- Calvo F. Free-energy landscapes from adaptively biased methods: Application to quantum systems. Phys. Rev. E. 82, 046703 (2010). [DOI] [PubMed] [Google Scholar]
- Wales D. J. Surveying a complex potential energy landscape: Overcoming broken ergodicity using basin-sampling. Chem. Phys. Lett. 584, 1–9 (2013). [Google Scholar]
- Wales D. J. Some further applications of discrete path sampling to cluster isomerization. Mol. Phys. 102, 891–908 (2004). [Google Scholar]
- Picciani M., Athenes M., Kurchan J. & Tailleur J. Simulating structural transitions by direct transition current sampling: The example of lj 3 8. J. Chem. Phys. 135, 034108 (2011). [DOI] [PubMed] [Google Scholar]
- de Souza V. K. & Wales D. J. Energy landscapes for diffusion: Analysis of cage-breaking processes. J. Chem. Phys. 129, 164507 (2008). [DOI] [PubMed] [Google Scholar]
- Strodel B., Whittleston C. S. & Wales D. J. Thermodynamics and kinetics of aggregation for the gnnqqny peptide. J. Am. Chem. Soc. 129, 16005–16014 (2007). [DOI] [PubMed] [Google Scholar]
- Wales D. J. Calculating rate constants and committor probabilities for transition networks by graph transformation. J. Chem. Phys. 130, 204111 (2009). [DOI] [PubMed] [Google Scholar]
- Chen J. Towards the physical basis of how intrinsic disorder mediates protein function. Arch Biochem Biophys 524, 123–131 (2012). [DOI] [PubMed] [Google Scholar]
- Barbar E. NMR characterization of partially folded and unfolded conformational ensembles of proteins. Biopolymers 51, 191–207 (1999). [DOI] [PubMed] [Google Scholar]
- Ebert M.-O., Bae S.-H., Dyson H. J. & Wright P. E. NMR relaxation study of the complex formed between CBP and the activation domain of the nuclear hormone receptor coactivator ACTR. Biochemistry 47, 1299–1308 (2008). [DOI] [PubMed] [Google Scholar]
- Vitalis A., Wang X. & Pappu R. V. Quantitative characterization of intrinsic disorder in polyglutamine: insights from analysis based on polymer theories. Biophys J. 93, 1923–1937 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wales D. J. & Doye J. P. K. Global optimization by basin-hopping and the lowest energy structures of lennard-jones clusters containing up to 110 atoms. J. Phys. Chem. A 101, 5111–5116 (1997). [Google Scholar]
- Uversky V. N., Gillespie J. R. & Fink A. L. Why are “natively unfolded” proteins unstructured under physiologic conditions? Proteins 41, 415–427 (2000). [DOI] [PubMed] [Google Scholar]
- Campen A. et al. Top-idp-scale: a new amino acid scale measuring propensity for intrinsic disorder. Protein Pept. Lett. 15, 956–963 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mao A. H., Crick S. L., Vitalis A., Chicoine C. L. & Pappu R. V. Net charge per residue modulates conformational ensembles of intrinsically disordered proteins. Proc. Natl. Acad. Sci. U S. A 107, 8183–8188 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das R. K. & Pappu R. V. Conformations of intrinsically disordered proteins are influenced by linear sequence distributions of oppositely charged residues. Proc. Natl. Acad. Sci. USA 110, 13392–13397 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holehouse A. S., Ahad J., Das R. K. & Pappu R. V. Cider: Classification of intrinsically disordered ensemble regions. (2014) Available at: http://pappulab.wustl.edu/cider (Accessed: 10th November 2014).
- Hornak V. et al. Comparison of multiple amber force fields and development of improved protein backbone parameters. Proteins 65, 712–725 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Case D. et al. The Amber biomolecular simulation programs. J. Comp. Chem. 26, 1668–1688 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fawzi N. L. et al. Structure and dynamics of the abeta(21-30) peptide from the interplay of nmr experiments and molecular simulations. J. Am. Chem. Soc. 130, 6145–6158 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorgensen W., Chandrasekhar J., Madura J., Impey R. & Klein M. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 79, 926–935 (1983). [Google Scholar]
- Ryckaert J.-P., Ciccotti G. & Berendsen H. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 23, 327–341 (1977). [Google Scholar]
- Loncharich R. J., Brooks B. R. & Pastor R. W. Langevin dynamics of peptides: the frictional dependence of isomerization rates of n-acetylalanyl-n’-methylamide. Biopolymers 32, 523–535 (1992). [DOI] [PubMed] [Google Scholar]
- Onufriev A., Bashford D. & Case D. A. Exploring protein native states and large-scale conformational changes with a modified generalized born model. Proteins 55, 383–394 (2004). [DOI] [PubMed] [Google Scholar]
- Shirts M. R. & Chodera J. D. Statistically optimal analysis of samples from multiple equilibrium states. J Chem Phys 129, 124105 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S., Rosenberg J., Bouzida D., Swendsen R. & Kollman P. The weighted histogram analysis method for free-energy calculations on biomolecules. i. the method. J. Comp. Chem. 13, 1011–1021 (1992). [Google Scholar]
- Feig M., Karanicolas J. & Brooks C. L. III Mmtsb tool set. MMTSB NIH Research Resource, The Scripps Research Institute, (2001).
- Kabsch W. & Sander C. Dictionary of protein secondary structure: pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 22, 2577–2637 (1983). [DOI] [PubMed] [Google Scholar]
- Li Z. & Scheraga H. A. Monte carlo-minimization approach to the multiple-minima problem in protein folding. Proc. Natl. Acad. Sci. USA. 84, 6611 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trygubenko S. A. & Wales D. J. A doubly nudged elastic band method for finding transition states. J. Chem. Phys. 120, 2082–2094 (2004). [DOI] [PubMed] [Google Scholar]
- Henkelman G. & Jónsson H. A dimer method for finding saddle points on high dimensional potential surfaces using only first derivatives. J. Chem. Phys. 111, 7010–7022 (1999). [Google Scholar]
- Munro L. J. & Wales D. J. Defect migration in crystalline silicon. Phys. Rev. B 59, 3969–3980 (1999). [Google Scholar]
- Liu D. & Nocedal J. On the limited memory bfgs method for large scale optimization. Math Prog. 45, 503–528 (1989). [Google Scholar]
- Carr J. M., Trygubenko S. A. & Wales D. J. Finding pathways between distant local minima. J. Chem. Phys. 122, 234903 (2005). [DOI] [PubMed] [Google Scholar]
- Wales D. J. Energy landscapes: Calculating pathways and rates. Int. Rev. Phys. Chem. 25, 237–282 (2006). [Google Scholar]
- Strodel B. & Wales D. J. Free energy surfaces from an extended harmonic superposition approach and kinetics for alanine dipeptide. Chem. Phys. Lett. 466, 105–115 (2008). [Google Scholar]
- Trygubenko S. A. & Wales D. J. Graph transformation method for calculating waiting times in markov chains. J. Chem. Phys. 124, 234110 (2006). [DOI] [PubMed] [Google Scholar]
- Carr J. M. & Wales D. J. Folding pathways and rates for the three-stranded beta-sheet peptide beta3s using discrete path sampling. J. Phys. Chem. B 112, 8760–8769 (2008). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.