

Abstract
Leishmania amazonensis (L. amazonensis) infection can cause severe local and diffuse injuries in humans, a condition clinically known as American cutaneous leishmaniasis (ACL). Currently, the therapeutic approach for ACL is based on Glucantime, which shows high toxicity and poor effectiveness. Therefore, ACL remains a neglected disease with limited options for treatment. Herein, the in vitro antiprotozoal effect and mechanisms of the diterpene kaurenoic acid [ent-kaur-16-en-19-oic acid] (KA) against L. amazonensis were investigated. KA exhibited a direct antileishmanial effect on L. amazonensis promastigotes. Importantly, KA also reduced the intracellular number of amastigote forms and percentage of infected peritoneal macrophages of BALB/c mice. Mechanistically, KA treatment reestablished the production of nitric oxide (NO) in a constitutive NO synthase- (cNOS-) dependent manner, subverting the NO-depleting escape mechanism of L. amazonensis. Furthermore, KA induced increased production of IL-1β and expression of the inflammasome-activating component NLRP12. These findings demonstrate the leishmanicidal capability of KA against L. amazonensis in macrophage culture by triggering a NLRP12/IL-1β/cNOS/NO mechanism.
1. Introduction
American cutaneous leishmaniasis (ACL) is a devastating illness caused by the protozoa Leishmania spp. ACL displays distinct clinical manifestations depending on both the parasite strain and the capability of the host to mount an effective immune response. Therefore, this disease may clinically appear in the host as cutaneous, mucocutaneous, or diffuse forms [1].
The treatment of ACL is based on a highly toxic chemotherapy with the antimonials sodium stibogluconate (Pentostam) and antimonate N-methyl-glucamine (Glucantime). In case of lack of response, second-line drugs such as amphotericin B or pentamidines are used [2]. However, these drugs frequently exhibit high toxicity, which has been related to its restricted use and resistance resulting in more restrictions in chemotherapy [2–5]. These considerations reveal the urgency to develop new therapeutic agents for the treatment of this disease.
Therefore, the search for more effective and less toxic chemotherapeutic agents for the treatment of ACL is increasing. There are various reported studies of synthetic compounds and natural products as potential sources of leishmanicidal activity [6, 7]. An interesting molecule towards this aim is kaurenoic acid [ent-kaur-16-en-19-oic acid] (KA), a diterpene obtained from various Brazilian plants [8, 9]. This molecule has been reported as showing a wide variety of biological activities such as antiprotozoal [10, 11], antimicrobial [12], antinociceptive [13], vasorelaxant, hypotensive [14, 15], and anti-inflammatory [13, 16, 17] activities. Moreover, some immunomodulatory properties of KA have been reported in these models.
The antiprotozoal activity of this diterpene involves its direct action in altering cell membrane integrity and mitochondrial membrane depolarization in promastigote and amastigote forms of L. amazonensis [18] and epimastigote forms of Trypanosoma cruzi [19], respectively. This direct activity of KA against the protozoa is not enough to reflect its overall potential as a therapeutic leishmanicidal drug, since Leishmania spp. parasites replicate intracellularly in macrophages and have several escape mechanisms against microbicidal molecules that are not dependent on the direct action of antileishmanial drugs [20–23].
Members of the Nod-like receptor (NLR) family of proteins have emerged as important innate immune sensors of pathogen-associated molecular patterns (PAMPS) and damage-associated molecular patterns (DAMPS) [24]. NLRs are the key components of the inflammasome that regulate the maturation of the potent inflammatory cytokine interleukin- (IL-) 1β [25].
Accordingly, Lima-Junior et al. found that the NLRP-3 inflammasome is engaged in the response against L. amazonensis restricting parasite replication. Additionally, IL-1β seems to be important for host resistance to infection by inducible (i) NOS-mediated production of NO, a major host defense mechanism against Leishmania spp. [26].
Taking into account the above-mentioned evidence, KA was evaluated for its in vitro effect on susceptible macrophages from BALB/c mice infected with L. amazonensis promastigote forms. Accordingly, we performed in vitro assays to investigate the direct effect of KA on parasites as well as its modulatory action on Leishmania-infected macrophages. We further investigated a putative mechanism of action of this compound as a modulator of proinflammatory molecules such as oxygen reactive species, NO, cytokines, and inflammasome.
2. Materials and Methods
2.1. Parasite
L. amazonensis (MHOM/BR/1989/166MJO) was used in promastigote forms, kept in culture medium 199 (Invitrogen-GIBCO) supplemented with 10% fetal bovine serum (Invitrogen-GIBCO), 1 M Hepes, 0.1% human urine, 0.1% L-glutamine, 10 U/mL penicillin and 10 μg/mL streptomycin (Invitrogen-GIBCO), and 10% sodium bicarbonate (complete medium for promastigotes—CMP). Cell cultures were incubated at 25°C in 25 cm2 flasks.
2.2. Animals
Female BALB/c mice weighing approximately 25–30 g and aged 6–8 weeks were obtained from Fundação Osvaldo Cruz, FIOCRUZ, Curitiba, Brazil. Mice were kept under pathogen-free conditions and used according to protocols approved by the Ethics Committee of the State University of Londrina (protocol number 33064/2012.42). Every effort was made to minimize the number of animals used and their suffering.
2.3. Plant Material
KA used in this paper was obtained from Sphagneticola trilobata. The crude extract was obtained from dried roots, which were pulverized and extracted with dichloromethane and partitioned with n-hexane and ethyl acetate; all solvents were dried under reduced pressure. The hexane fraction was subjected to vacuum liquid chromatography (VLC) by increasing gradient polarity. The second fraction produced an amorphous compound (200 mg), which was washed with cold methanol and analyzed by high performance liquid chromatography (HPLC) methods, yielding 96% purity. The identification was performed by 1H and 13 C nuclear magnetic resonance (NMR), electron impact mass spectrometry (EIMS), and comparison with literature data [27]. The stock solution of KA was dissolved in 2% dimethyl sulfoxide (DMSO) (Invitrogen-Gibco). However, DMSO concentration did not exceed 0.2% in all experiments.
2.4. Viability of Promastigotes
The viability of L. amazonensis promastigote forms treated with KA was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as previously described [28]. Promastigote forms (106/100 μL) were incubated with different concentrations of KA (10, 30, 50, 70, and 90 μM) or with KA solvent (0.2% DMSO) and maintained in culture for 24, 48, and 72 h at 25°C. Thereafter, 10 μL of MTT (5 mg/mL) were added, followed by incubation for an additional 4 h at 24°C. The MTT formazan product was diluted with 300 μL of DMSO, transferring to 96-well plates and measured in a spectrophotometer with absorbance determined at 550 nm. The results were expressed as percentage MTT reduction relative to the control group calculated as the following formula: (viable promastigotes)% = (OD of drug-treated sample/OD of untreated sample) × 100.
2.5. Cell Proliferation Kinetics
Promastigote forms (106/mL) incubated in CMP were treated with different concentrations of KA (10, 30, 50, 70, and 90 μM) or with KA solvent (0.2% DMSO) and cultured for 5 days at 25°C. Promastigotes were counted in a Neubauer chamber after 24, 48, 72, and 120 h.
2.6. Phagocytic Assay
Macrophages (5 × 105/mL) were obtained from the peritoneal cavity by the injection of 2 mL of RPMI 1640 culture medium (Invitrogen-GIBCO) supplemented with 10% fetal bovine serum (Invitrogen-GIBCO) and cultured on 24-well plates containing 13 mm diameter glass coverslips. Cells were preincubated with 200 μL of RPMI medium for 2 h for adherence and incubated with promastigote forms (5 : 1) for 2 h. KA (50, 70 or 90 μM) or medium was added, followed by 24 h incubation at 37°C and 5% CO2. The cells were stained with Giemsa to establish the phagocytic index of infection (by percentage) and the parasites/macrophage (mean). The supernatant was utilized to measure the levels of malondialdehyde (MDA), total antioxidant capacity of plasma (TRAP), nitric oxide (NO), and cytokines.
2.7. Measurement of the Total Antioxidant Capacity of Samples (Trapping Antioxidant Parameter (TRAP))
Samples (50 μL of supernatant with cells) obtained from the phagocytic assay were analyzed as previously described by Repetto et al. [29], by using the chemiluminescence-based method. Soluble vitamin E (Trolox) was employed as a standard antioxidant. The chemiluminescence curves were obtained using the Glomax luminometer (Promega), and the results were expressed in nM Trolox.
2.8. Measurement of Malondialdehyde Levels (MDA) by High Performance Chromatography
MDA levels were determined as resultant of oxidative stress occurrence, which causes lipid peroxidation and the production of this metabolite. Accordingly, we used HPLC as previously described by Victorino et al. [30], with slight modifications. The analyses were conducted with an Alliance e2695 HPLC (Waters, Milford, MA, USA) equipped with a SecurityGuard ODS-C18 (4 × 3.0 mm, Phenomenex), C18 reverse phase column (Eclipse XDB-C18; 4.6 × 250 mm, 5 μm, Agilent), and a photodiode array detector (Photodiode Array Detector (PDA), 2998). Analyses were conducted using Empower 2 software (Waters, Milford, MA, USA). MDA standards were prepared using 1,1,3,3-tetraethoxypropane (TEP). Aliquots containing 250 μL of cells + supernatants were deproteinized by adding 20% trichloroacetic acid and reacted with 1 mL of thiobarbituric acid. The mobile phase was 70% 10 mM KH2PO4 buffer, pH 7.0, and 40% HPLC-grade methanol. Readings were obtained at 532 nm, following an 8 min isocratic flow at the rate of 1 mL/min. The results were expressed in nM MDA.
2.9. Determination of Nitrite Levels as Estimate of NO Production
The determination of nitrite in supernatants collected from the phagocytic tests was used to measure the concentration of nitric oxide (NO) according to Panis et al. [31], with some modifications. Briefly, the supernatant aliquots were deproteinized by adding 50 μL of 75 mM ZnSO4 and 70 μL of NaOH and shaking and centrifuging for 5 min at 10,000 rpm and 25°C. The supernatant was recovered and diluted in glycine buffer (45 g/L, pH 9.7). Cadmium granules were rinsed with distilled sterile water and added to a 5 mM CuSO4 in glycine-NaOH buffer (15 g/L, pH 9.7), followed by 5 min incubation, and the copper-coated cadmium granules were used within 10 min. Activated granules were added to glycine buffer-diluted supernatant and the suspension stirred for 10 min. Aliquots of 200 μL were recovered in appropriate tubes for nitrite determination, and the same volume of Griess reagent was added. After 10 min incubation at room temperature, tubes were centrifuged at 10,000 rpm for 2 min at 25°C and added to 96-well microplates in triplicate. A calibration curve was prepared using dilutions of NaNO2, and the absorbance was determined at 550 nm in a microplate reader.
2.10. Cytokine Determination
The supernatants obtained from the phagocytic assay were used to determine the levels of IL-1β, IL-12, TNF-α, IFN-γ, TGF-β, and IL-10 using eBioscience commercial kits capture enzyme-linked immune sorbent assay (ELISA) (San Diego, CA, USA). According to the manufacturer's instructions, absorbance was read at 450 nm using a spectrophotometer and the results are expressed in pg/mL based on a standard curve. The sensitivity of the test was 8 pg/mL for IL-1β, TNF-α, and TGF-β, 15 pg/mL for IL-12 and IFN-γ, and 32 pg/mL for IL-10.
2.11. Immunocytochemical Labeling for NLRP12 and iNOS
Immunocytochemistry of NLRP12 and inducible nitric oxide synthase (iNOS) was performed on coverslip-adherent cells (cells prepared according to the protocol described in the phagocytic assay) using the labeled streptavidin biotin method with the LSAB kit (DAKO Japan, Kyoto, Japan) without microwave treatment. The coverslips were incubated with 10% Triton X-100 for 1 h, washed 3 times with PBS, and treated for 40 min at room temperature with 10% BSA. In addition, coverslips were incubated overnight at 4°C with the primary antibody (anti-NLRP12 rabbit polyclonal antibody diluted 1 : 300 (Abcam, catalog number ab93113) and anti-iNOS rabbit monoclonal antibody diluted 1 : 200 (BD Biosciences, catalog number 610599)). After secondary antibody treatment (2 h, room temperature), horseradish peroxidase activity was visualized by treatment with H2O2 and 3,3′-diaminobenzidine (DAB) for 5 min. In the last step, the sections were weakly counterstained with Harry's hematoxylin (Merck). For each case, negative controls were performed by omitting the primary antibody. Intensity and localization of immunoreactivity against primary antibody used were examined in all coverslips using a photomicroscope (Olympus BX41, Olympus Optical Co., Ltd., Tokyo, Japan). Color photomicrographs of representative areas (×40 objective lens) were digitally acquired for image analysis. For determining a semiquantitative scoring, images were evaluated by using the color deconvolution tool from Image J software (NIH, USA). Pixels were categorized as previously described by Chatterjee et al. [32] as strong positive (3+), positive (2+), weak positive (1+), and negative (0).
2.12. cNOS Inhibition Assay
Peritoneal macrophages were challenged with L. amazonensis and treated with KA as described in the phagocytic assay method. Before the treatment with KA, the cells were incubated with 20 μM NG-nitro-L-arginine methyl ester (L-NAME) for 2 h at 36°C and 5% CO2 [33], aiming to cause pharmacological blockage of constitutive NOS. The supernatants were utilized to measure NO levels (as previously described).
2.13. Statistical Analysis
Three independent experiments were performed, each with triplicate datasets. Data were expressed as mean ± standard error of the mean. Data were analyzed using the GraphPad Prism statistical software (GraphPad Software, Inc., USA, 500.288). Significant differences between the treatments were determined by one-way ANOVA, followed by Tukey's test for multiple comparisons. P < 0.05 was considered statistically significant.
3. Results
3.1. Kaurenoic Acid Exerts Leishmanicidal Effect against Promastigote and Amastigote Forms of L. Amazonensis
In the first set of experiments, the antileishmanial effect of KA was investigated against the promastigote forms of L. amazonensis. We observed that KA at concentrations of 50, 70, and 90 μM reduced promastigote viability according to an MTT assay of 24 h by 30, 31, and 34%, respectively (Figure 1(a)), the reduction was maintained for 72 h. We also observed 28.2, 45.8, and 51.5% decrease in the proliferation of the promastigote forms at concentrations of 50, 70, and 90 μM, respectively, after 120 h with 24 h of pretreatment (Figure 1(b)). Therefore, we chose testing all concentrations of KA for 24 h treatment of macrophages.
Figure 1.

Kaurenoic acid has a leishmanicidal effect against promastigote and amastigote forms of L. amazonensis. MTT assay in promastigote forms of L. amazonensis treated with kaurenoic acid (10, 30, 50, 70, and 90 μM) or 0.2% DMSO for 24, 48, and 72 h (Panel (a)). Proliferation kinetics of L. amazonensis promastigote forms after pretreatment for 24 h with kaurenoic acid (10, 30, 50, 70, and 90 μM) or 0.2% DMSO for 24, 48, 72, and 120 h (Panel (b)). Percentage of infected macrophages (Panel (c)) and number of amastigotes per macrophage after 24 h of incubation with kaurenoic acid (50, 70, and 90 μM) (Panel (d)). Data represent mean ± SEM of three independent experiments (promastigotes) and six independent experiments (amastigotes). (∗Significantly different from control (P < 0.05 compared with control group promastigotes in culture medium) (one-way ANOVA followed by Tukey's test).)
In attempt to verify if KA could enhance the leishmanicidal capacity of macrophages, we initially challenged these cells with promastigote forms of L. amazonensis for 2 h, for phagocytosis. Afterwards, the cells were treated with KA (at concentrations ranging from 50 to 90 μM) for 24 h. Macrophages and amastigotes were counted to establish the phagocytic index, indicating the extent of infection as the number of parasites per macrophage.
Regarding the percentage of infected macrophages, 50, 70, and 90 μM KA, respectively, caused a decrease of 26.6, 25.6, and 28.4% of infected macrophages when compared to the untreated infected macrophages (Figure 1(c)). Moreover, the mean number of amastigotes per macrophage was significantly decreased at the concentrations of 70 and 90 μM, by 21.5 and 20.3%, respectively (Figure 1(d)).
3.2. Effect of Kaurenoic Acid on Macrophages Is Not Associated with ROS Production
In order to assess the involvement of KA in modulating the respiratory burst of macrophages during the Leishmania challenge, we measured the oxidative stress status of these cells by quantifying its total antioxidant capacity (TRAP) and MDA formation.
There was an increase in TRAP of macrophages infected with L. amazonensis treated with 50 and 70 μM KA, while TRAP was reduced by 90 μM KA compared to the infected control (Figure 2(a)). The MDA level was reduced at all KA concentrations with significant difference at 50 μM KA (Figure 2(b)).
Figure 2.

The effect of kaurenoic acid on macrophages is not associated with oxidative stress generation. Total antioxidant capacity (TRAP) (Panel (a)) and MDA levels (Panel (b)) were evaluated as markers of the oxidative status in supernatant or macrophages infected with L. amazonensis and treated with kaurenoic acid (50, 70, and 90 μM) for 24 h. Data represent the mean ± SEM of three independent experiments. (∗Significantly different from infected cells (P < 0.05)) (one-way ANOVA followed by Tukey's test).
3.3. Kaurenoic Acid Upregulates NO Levels in a cNOS-Dependent Mechanism
Concerning NO levels, our results showed that untreated macrophages infected with L. amazonensis displayed decreased levels of NO. Interestingly, the treatment with KA reestablished baseline NO levels at all concentrations tested (Figure 3(a)). Thus, we next investigated the enzymatic pathway involved in KA-induced NO production.
Figure 3.

Kaurenoic acid exerted its effect by upregulating NO levels in a cNOS-dependent mechanism. NO levels (Panel (a)); immunocytochemistry scoring for inducible nitric oxide synthase (iNOS). Peritoneal macrophages infected with L. amazonensis and treated with kaurenoic acid (50, 70, and 90 μM) for 24 h (Panel (b)). Determination of NO in peritoneal macrophages infected with L. amazonensis and blocked with 20 μM L-NAME and treated with kaurenoic acid (50, 70, and 90 μM) for 24 h (Panel (c)). Data represent the mean ± SEM of three independent experiments. (∗Significantly different from infected cells (P < 0.05). #Significantly different from control cells (P < 0.05) (one-way ANOVA followed by Tukey's test).)
The expression of iNOS was assessed by immunocytochemistry. Our data showed that KA did not alter iNOS expression (Figure 3(b)).
Aiming to investigate the involvement of cNOS, we pretreated macrophages with the preferential cNOS inhibitor L-NAME. After the Leishmania challenge, macrophages were pretreated with L-NAME and then incubated with KA, resulting in a substantial reduction in NO production only at 90 μM, indicating that the augmented NO previously observed at 90 μM KA was probably dependent on cNOS activity (Figure 3(c)).
3.4. Kaurenoic Acid Promotes the Production of Active IL-1β in Macrophages Infected with L. amazonensis
In order to determine the immunomodulatory action of KA on cytokines in infected macrophages, we measured the levels of IL-12, TNF-α, IFN-γ, TGF-β, IL-10, and IL-1β. We observed that the production of IL-12, TNF-α, IFN-γ, TGF-β, and IL-10 was not significantly different between the KA-treated groups and the control group (Figure 4). On the other hand, 70 and 90 μM KA augmented the levels of IL-1β (Figure 4(f)).
Figure 4.

Kaurenoic acid promotes the production of active IL-1β in macrophages infected with L. amazonensis. Mapping the cytokine profiling produced in vitro by macrophages infected with L. amazonensis and treated with kaurenoic acid (50, 70, and 90 μM) for 24 h determined by ELISA. IL-12 production (Panel (a)), TNF-α production (Panel (b)), IFN-γ production (Panel (c)), TGF-β production (Panel (d)), IL-10 production (Panel (e)), and IL-1β production (Panel (f)). Data represent the mean ± SEM of three independent experiments. (∗Significantly different from infected cells (P < 0.05) (one-way ANOVA followed by Tukey's test).)
3.5. Kaurenoic Acid Upregulates NLRP12 Expression in Macrophages Infected with L. amazonensis
In this present study, the activation and participation of the inflammasome during the immune response to infection by intracellular pathogen were investigated. The augmented levels of IL-1β induced during KA treatment of infected macrophages, combined with the lack of information about this complex in ACL, led us to investigate the role of NLRP12, one member of the subfamily of NLRP innate receptors. As shown in Figure 5, KA at 90 μM was able to upregulate NLRP12 expression in macrophages infected with L. amazonensis. These results indicate that the stimulation of macrophages with KA triggered the overexpression of NLRP12, with consequent activation of IL-1β.
Figure 5.
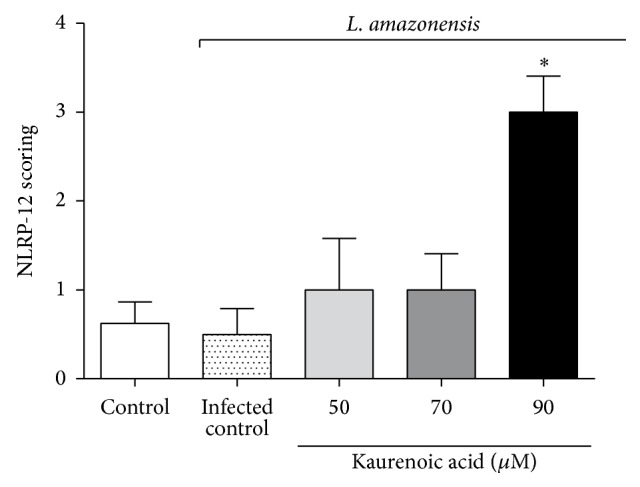
Kaurenoic acid upregulated the NLRP12 expression in macrophages infected with L. amazonensis. Immunocytochemistry scoring for NLRP12 in macrophages infected with L. amazonensis and treated with kaurenoic acid (50, 70, and 90 μM) for 24 h. Data represent the mean ± SEM of three independent experiments. (∗Significantly different from infected cells (P < 0.05). (One-way ANOVA followed by Tukey's test).)
4. Discussion
The success of chemotherapy in ACL is mainly dependent on two factors: the microbicidal activity of the drug and the protective immune response triggered in the host during the treatment.
In the present study, we evaluated the therapeutic potential of KA treatment, which directly inhibited the viability and proliferation of L. amazonensis promastigote forms (Figure 1(a)). Previous in vitro studies demonstrated that KA has direct antileishmanial activity against promastigote and amastigote forms of L. amazonensis [19, 34, 35], and the main proposed mechanism was related to mitochondrial membrane depolarization in the protozoan. However, the immunomodulatory activity of KA in the macrophage experimental leishmaniasis model still remained to be determined.
Our results indicated that infected macrophages treated with KA were more effective during the leishmanicidal response against the intracellular forms of L. amazonensis (Figures 1(b) and 1(c)). The solvent (0.2% DMSO) did not affect the viability and proliferation of L. amazonensis promastigotes (Figures 1(a) and 1(b)). These findings suggested that this diterpene was able to reverse the downregulation of the killer machinery of macrophages caused by Leishmania infection [20, 36].
In order to elucidate the microbicidal effects of KA, we investigated the main leishmanicidal molecules produced by macrophages. The results showed that KA treatment was not able to enhance the oxidative burst of infected macrophages. On the other hand, even though L. amazonensis is capable of depleting NO levels [20], the results showed that treatment with KA was able to restore the levels of this microbicidal molecule (Figure 3(a)), but no alteration in the expression of iNOS was found.
In fact, NO is the main antileishmanial molecule produced in the early macrophage response against intracellular parasites. Besides iNOS, cNOS is also an important route for NO production [37, 38]. Some studies have demonstrated that KA induces cNOS-dependent activity in the disease context [13, 14], but the involvement of a cNOS mechanism for KA in parasitic infections has not been well elucidated.
Studies have demonstrated the importance of innate immune response-triggered cytokine production during the early stages of experimental leishmaniasis and have shown that some cytokines may drive the clinical manifestation of ACL by modulating resistance or susceptibility to infection [39]. IL-1β, IL-12, TNF-α, and IFN-γ are essential cytokines for the development of an effective immune response against Leishmania spp., leading to the activation of macrophages and promoting the microbicidal effects against this parasite [23, 26, 40, 41].
In the present experimental conditions, KA was able to increase the production of IL-1β despite of showing no effect on other cytokines evaluated, suggesting a selective effect of KA on IL-1β production/maturation system. IL-1β is a proinflammatory cytokine that becomes active after its cleavage by the inflammasome complex [42], and when active, this cytokine helps the activation of macrophages by enhancing the response against pathogens. In fact, a recent study showed that IL-1β was associated with resistance to L. amazonensis, L. braziliensis, and L. major infections. IL-1β maturation is dependent on the inflammasome NLRP3/ASC/caspase-1 complex [26]. In addition to NLRP3, which is endogenously expressed during Leishmania infection, NLRP12 is an important NLR involved in the inflammatory response against parasites such as Trypanosoma cruzi [43]. In this sense, the present study also addressed whether the expression of NLRP12 increased during Leishmania infection of macrophages to determine if KA induces IL-1β production/maturation by a previously unrecognized mechanism.
Our findings showed that there was no induction of NLRP12 in macrophages during Leishmania infection. On the other hand, KA at 90 μM upregulated the expression of NLRP12 in infected macrophages, thus explaining the increased production of IL-1β induced by KA. These data are also in line with the increased cNOS-dependent production of NO, since it has been shown that during Leishmania infection a NLRP3/ASC/caspase-1/IL-1β/cNOS/NO pathway is triggered to kill this parasite. In the present study, the data suggest that KA triggers a NLRP12/IL-1β/cNOS/NO killing mechanism during Leishmania infection of macrophages from BALB/c mice. Importantly, KA-induced the expression of a NLR (NLRP12) that is not endogenously activated to kill Leishmania, thereby upregulating unused endogenous mechanisms valuable to protect the host against Leishmania infection with additive effects for other NLRs such as NLRP3, as observed in T. cruzi infection [43]. Therefore, KA seems to unequivocally provide additional protective mechanisms against Leishmania infection. It is also possible that KA triggers similar mechanisms in other parasitic diseases, which remains to be determined.
In conclusion, the present study demonstrated that kaurenoic acid has therapeutic potential as a pharmacological approach against Leishmania infection. The mechanism of action of kaurenoic acid depends, at least in part, on triggering the NLRP12/IL-1β/cNOS/NO leishmanicidal pathway. Therefore, KA merits further preclinical and clinical studies as a possible therapy for Leishmania infection.
Acknowledgments
The authors gratefully acknowledge Juliano Bordignon for supplying the animals used in this paper. This study was supported by Conselho Nacional de Pesquisa (CNPq, Brazil), Coordenadoria de Aperfeiçoamento Pessoal de Nível Superior (CAPES, Brazil), and Fundação Araucária by Governo do Estado do Paraná (Brazil). Dr. A. Leyva helped with the English editing of the paper.
Conflict of Interests
The authors have no conflict of interests to declare.
Authors' Contribution
Milena Menegazzo Miranda, Carolina Panis, Suelen Santos da Silva, and Juliana Aparecida Macri equally contributed to this study.
References
- 1.Kaye P., Scott P. Leishmaniasis: complexity at the host-pathogen interface. Nature Reviews Microbiology. 2011;9(8):604–615. doi: 10.1038/nrmicro2608. [DOI] [PubMed] [Google Scholar]
- 2.Ministério da Saúde. Manual for Surveillance of American Tegumentary Leishmaniasis. 2nd. Brasília, Brazil: Ministério da Saúde, Secretaria de Vigilância em Saúde, Editora do Ministério da Saúde; 2010. (Série A Normas e Manuais Técnicos). [Google Scholar]
- 3.Goto H., Lindoso J. A. L. Current diagnosis and treatment of cutaneous and mucocutaneous leishmaniasis. Expert Review of Anti-Infective Therapy. 2010;8(4):419–433. doi: 10.1586/eri.10.19. [DOI] [PubMed] [Google Scholar]
- 4.Silva-López R. E. Proteases de Leishmania: novos alvos para o desenvolvimento racional de fármacos. Química Nova. 2010;33(7):1541–1548. doi: 10.1590/s0100-40422010000700022. [DOI] [Google Scholar]
- 5.Oliveira L. F., Schubach A. O., Martins M. M., et al. Systematic review of the adverse effects of cutaneous leishmaniasis treatment in the New World. Acta Tropica. 2011;118(2):87–96. doi: 10.1016/j.actatropica.2011.02.007. [DOI] [PubMed] [Google Scholar]
- 6.Tiuman T. S., Santos A. O., Ueda-Nakamura T., Filho B. P. D., Nakamura C. V. Recent advances in leishmaniasis treatment. International Journal of Infectious Diseases. 2011;15(8):e525–e532. doi: 10.1016/j.ijid.2011.03.021. [DOI] [PubMed] [Google Scholar]
- 7.da Silva S. S., Thomé G. D. S., Cataneo A. H. D., et al. Brazilian propolis antileishmanial and immunomodulatory effects. Evidence-Based Complementary and Alternative Medicine. 2013;2013:7. doi: 10.1155/2013/673058.673058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Baccarin T., Czepula A. I., Ferreira R. A., Lucinda-Silva R. M. Análise morfoanatômica das partes aéreas de Wedelia paludosa DC. (Acmela brasiliensis, Sphagneticola trilobata), Asteraceae. Revista Brasileira de Farmacognosia. 2009;19:612–616. [Google Scholar]
- 9.Batista R., Brandão G. C., Braga F. C., Oliveira A. B. Cytotoxicity of Wedelia paludosa D.C. extracts and constituents. Brazilian Journal of Pharmacognosy. 2009;19(1):36–40. doi: 10.1590/s0102-695x2009000100009. [DOI] [Google Scholar]
- 10.Batista R., Chiari E., de Oliveira A. B. Trypanosomicidal kaurane diterpenes from Wedelia paludosa . Planta Medica. 1999;65(3):283–284. doi: 10.1055/s-2006-960781. [DOI] [PubMed] [Google Scholar]
- 11.Batista R., Humberto J. L., Chiari E., de Oliveira A. B. Synthesis and trypanocidal activity of ent-kaurane glycosides. Bioorganic and Medicinal Chemistry. 2007;15(1):381–391. doi: 10.1016/j.bmc.2006.09.048. [DOI] [PubMed] [Google Scholar]
- 12.Wilkens M., Alarcón C., Urzúa A., Mendoza L. Characterization of the bactericidal activity of the natural diterpene kaurenoic acid. Planta Medica. 2002;68(5):452–454. doi: 10.1055/s-2002-32086. [DOI] [PubMed] [Google Scholar]
- 13.Mizokami S. S., Arakawa N. S., Ambrosio S. R., et al. Kaurenoic acid from Sphagneticola trilobata inhibits inflammatory pain: effect on cytokine production and activation of the NO-cyclic GMP-protein kinase G-ATP-sensitive potassium channel signaling pathway. Journal of Natural Products. 2012;75(5):896–904. doi: 10.1021/np200989t. [DOI] [PubMed] [Google Scholar]
- 14.Tirapelli C. R., Ambrosio S. R., da Costa F. B., Coutinho S. T., de Oliveira D. C. R., de Oliveira A. M. Analysis of the mechanisms underlying the vasorelaxant action of kaurenoic acid in the isolated rat aorta. European Journal of Pharmacology. 2004;492(2-3):233–241. doi: 10.1016/j.ejphar.2004.04.003. [DOI] [PubMed] [Google Scholar]
- 15.Tirapelli C. R., Ambrosio S. R., de Oliveira A. M., Tostes R. C. Hypotensive action of naturally occurring diterpenes: a therapeutic promise for the treatment of hypertension. Fitoterapia. 2010;81(7):690–702. doi: 10.1016/j.fitote.2010.05.018. [DOI] [PubMed] [Google Scholar]
- 16.Lim H., Jung H. A., Choi J. S., Kim Y. S., Kang S. S., Kim H. P. Anti-inflammatory activity of the constituents of the roots of Aralia continentalis . Archives of Pharmacal Research. 2009;32(9):1237–1243. doi: 10.1007/s12272-009-1909-3. [DOI] [PubMed] [Google Scholar]
- 17.Choi R. J., Shin E. M., Jung H. A., Choi J. S., Kim Y. S. Inhibitory effects of kaurenoic acid from Aralia continentalis on LPS-induced inflammatory response in RAW264.7 macrophages. Phytomedicine. 2011;18(8-9):677–682. doi: 10.1016/j.phymed.2010.11.010. [DOI] [PubMed] [Google Scholar]
- 18.Izumi E., Ueda-Nakamura T., Veiga V. F., Pinto A. C., Nakamura C. V. Terpenes from copaifera demonstrated in vitro antiparasitic and synergic activity. Journal of Medicinal Chemistry. 2012;55(7):2994–3001. doi: 10.1021/jm201451h. [DOI] [PubMed] [Google Scholar]
- 19.dos Santos A. O., Izumi E., Ueda-Nakamura T., Dias-Filho B. P., da Veiga-Júnior V. F., Nakamura C. V. Antileishmanial activity of diterpene acids in copaiba oil. Memórias do Instituto Oswaldo Cruz. 2013;108(1):59–64. doi: 10.1590/s0074-02762013000100010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Van Assche T., Deschacht M., Da Luz R. A. I., Maes L., Cos P. Leishmania-macrophage interactions: insights into the redox biology. Free Radical Biology and Medicine. 2011;51(2):337–351. doi: 10.1016/j.freeradbiomed.2011.05.011. [DOI] [PubMed] [Google Scholar]
- 21.Faria M. S., Reis F. C. G., Lima A. P. C. A. Toll-like receptors in Leishmania infections: guardians or promoters? Journal of Parasitology Research. 2012;2012:12. doi: 10.1155/2012/930257.930257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Handman E., Bullen D. V. Interaction of Leishmania with the host macrophage. Trends in Parasitology. 2002;18(8):332–334. doi: 10.1016/S1471-4922(02)02352-8. [DOI] [PubMed] [Google Scholar]
- 23.Sacks D., Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nature Reviews Immunology. 2002;2(11):845–858. doi: 10.1038/nri933. [DOI] [PubMed] [Google Scholar]
- 24.Krishnaswamy J. K., Chu T., Eisenbarth S. C. Beyond pattern recognition: NOD-like receptors in dendritic cells. Trends in Immunology. 2013;34(5):224–233. doi: 10.1016/j.it.2012.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Takahashi M. NLRP3 inflammasome as a novel player in myocardial infarction. International Heart Journal. 2014;55(2):101–105. doi: 10.1536/ihj.13-388. [DOI] [PubMed] [Google Scholar]
- 26.Lima-Junior D. S., Costa D. L., Carregaro V., et al. Inflammasome-derived IL-1β production induces nitric oxide-mediated resistance to Leishmania. Nature Medicine. 2013;19(7):909–915. doi: 10.1038/nm.3221. [DOI] [PubMed] [Google Scholar]
- 27.da Costa F. B., Albuquerque S., Vichnewski W. Diterpenes and synthetic derivatives from Viguiera aspillioides with trypanomicidal activity. Planta Medica. 1996;62(6):557–559. doi: 10.1055/s-2006-957971. [DOI] [PubMed] [Google Scholar]
- 28.Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of Immunological Methods. 1983;65(1-2):55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 29.Repetto M., Reides C., Gomez Carretero M. L., Costa M., Griemberg G., Llesuy S. Oxidative stress in blood of HIV infected patients. Clinica Chimica Acta. 1996;255(2):107–117. doi: 10.1016/0009-8981(96)06394-2. [DOI] [PubMed] [Google Scholar]
- 30.Victorino V. J., Panis C., Campos F. C., et al. Decreased oxidant profile and increased antioxidant capacity in naturally postmenopausal women. Age. 2013;35(4):1411–1421. doi: 10.1007/s11357-012-9431-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Panis C., Herrera A. C. S. A., Victorino V. J., et al. Oxidative stress and hematological profiles of advanced breast cancer patients subjected to paclitaxel or doxorubicin chemotherapy. Breast Cancer Research and Treatment. 2012;133(1):89–97. doi: 10.1007/s10549-011-1693-x. [DOI] [PubMed] [Google Scholar]
- 32.Chatterjee S., Malhotra R., Varghese F., et al. Quantitative immunohistochemical analysis reveals association between sodium iodide symporter and estrogen receptor expression in breast cancer. PLoS ONE. 2013;8(1) doi: 10.1371/journal.pone.0054055.e54055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee K.-S., Lee D.-K., Jeoung D., et al. Differential effects of substrate-analogue inhibitors on nitric oxide synthase dimerization. Biochemical and Biophysical Research Communications. 2012;418(1):49–55. doi: 10.1016/j.bbrc.2011.12.123. [DOI] [PubMed] [Google Scholar]
- 34.Santos A. O., Ueda-Nakamura T., Filho B. P. D., Junior V. F. V., Pinto A. C., Nakamura C. V. Effect of Brazilian copaiba oils on Leishmania amazonensis . Journal of Ethnopharmacology. 2008;120(2):204–208. doi: 10.1016/j.jep.2008.08.007. [DOI] [PubMed] [Google Scholar]
- 35.dos Santos A. O., Ueda-Nakamura T., Filho B. P. D., Junior V. F. D. V., Nakamura C. V. Copaiba oil: an alternative to development of new drugs against leishmaniasis. Evidence-Based Complementary and Alternative Medicine. 2012;2012:7. doi: 10.1155/2012/898419.898419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Balestieri F. M., Queiroz A. R., Scavone C., Costa V. M., Barral-Netto M., Abrahamsohn Ide A. Leishmania (L.) amazonensis-induced inhibition of nitric oxide synthesis in host macrophages. Microbes and Infection. 2002;4(1):23–29. doi: 10.1016/s1286-4579(01)01505-2. [DOI] [PubMed] [Google Scholar]
- 37.Huang Z., Hoffmann F. W., Fay J. D., et al. Stimulation of unprimed macrophages with immune complexes triggers a low output of nitric oxide by calcium-dependent neuronal nitric-oxide synthase. The Journal of Biological Chemistry. 2012;287(7):4492–4502. doi: 10.1074/jbc.m111.315598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Connelly L., Jacobs A. T., Palacios-Callender M., Moncada S., Hobbs A. J. Macrophage endothelial nitric-oxide synthase autoregulates cellular activation and pro-inflammatory protein expression. The Journal of Biological Chemistry. 2003;278(29):26480–26487. doi: 10.1074/jbc.m302238200. [DOI] [PubMed] [Google Scholar]
- 39.Nylen S., Gautam S. Immunological perspectives of leishmaniasis. Journal of Global Infectious Diseases. 2010;2:135–146. doi: 10.4103/0974-777X.62876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Babior B. M. Phagocytes and oxidative stress. American Journal of Medicine. 2000;109(1):33–44. doi: 10.1016/s0002-9343(00)00481-2. [DOI] [PubMed] [Google Scholar]
- 41.Cunningham A. C. Parasitic adaptive mechanisms in infection by Leishmania . Experimental and Molecular Pathology. 2002;72(2):132–141. doi: 10.1006/exmp.2002.2418. [DOI] [PubMed] [Google Scholar]
- 42.van de Veerdonk F. L., Netea M. G., Dinarello C. A., Joosten L. A. B. Inflammasome activation and IL-1β and IL-18 processing during infection. Trends in Immunology. 2011;32(3):110–116. doi: 10.1016/j.it.2011.01.003. [DOI] [PubMed] [Google Scholar]
- 43.Ataide M. A., Andrade W. A., Zamboni D. S., et al. Malaria-induced NLRP12/NLRP3-dependent caspase-1 activation mediates inflammation and hypersensitivity to bacterial superinfection. PLoS Pathogens. 2014;10(1) doi: 10.1371/journal.ppat.1003885.e1003885 [DOI] [PMC free article] [PubMed] [Google Scholar]