

Abstract
With the widespread ban on the use of antibiotics in swine feed, alternative measures need to be sought to maintain swine health and performance. Antimicrobial peptides (AMPs) are part of the nonspecific defense system and are natural antibiotics produced by plants, insects, mammalians, and micro-organisms as well as by chemical synthesis. Due to their broad microbicidal activity against various fungi, bacteria and enveloped viruses, AMPs are a potential alternative to conventional antibiotics for use in swine production. This review focuses on the structure and mechanism of action of AMPs, as well as their effects on performance, immune function and intestinal health in pigs. The aim is to provide support for the application of AMPs as feed additives replacing antibiotics in swine nutrition.
Keywords: Antimicrobial peptides, Antibiotics, Applications, Swine
Background
Antibiotics have been used in the swine industry for more than 50 years to improve growth and prevent infectious diseases. However, the misuse of antibiotics has caused many problems including the emergence of bacteria resistant to antibiotics and the potential of producing drug residues in meat products [1]. As a result, a global trend has emerged towards restriction of the inclusion of antibiotics in swine diets as a routine means of growth promotion. In response, a considerable amount of research has been focused on the development of alternatives to antibiotics to maintain swine performance and health.
Antimicrobial peptides (AMPs) are one of the most widely researched alternatives to conventional antibiotics. AMPs are potent, broad spectrum antibiotics which have been demonstrated to kill gram-negative and gram-positive bacteria, mycobacteria, viruses, fungi and even transformed or cancerous cells while having no effect on the cells of treated animals [2]. In recent years, studies on AMPs and their applications have become one of the hot spots in the areas of agricultural science, biology, medicine, and physiology as well as having potential applications in medicine and the food industry.
Supplementation with various antimicrobial peptides has been reported to have positive effects on performance, nutrient digestibility, the intestinal microflora, intestinal morphology and immune function in pigs [3-5]. This article provides an overview of AMPs, their categories and structure, mechanism of action and their potential applications in swine production.
Structure and categories of antimicrobial peptides
AMPs are oligopeptides with a variable composition of amino acids and amino acid number (typically 6 to 100 amino acids). Based on the different sources, AMPs are divided into mammalian AMPs (e.g. defensin), amphibian AMPs (e.g. magainins), insect AMPs (e.g. cecropin), plant AMPs (e.g. thionin), and microbial AMPs (e.g. gramicidin and nisin). Based on their biological activities, AMPs can also be divided into antiviral peptides (e.g. defensins, and NP-1), antibacterial peptides (e.g. nisin, and pyrrhocoricin), antifungal peptides, and antiparasitic peptides [6].
AMPs are small, positively charged, amphipathic molecules which possess both hydrophobic and hydrophilic regions. Based on their secondary structure, AMPs are characterized as one of four types including α-helical, β-sheet (due to the presence of 2 or more disulfide bonds), alpha-beta and non-alpha-beta structure [7]. Because they consist solely of amino acids, it is very easy to modify the structure of AMPs. Chemical synthesis or recombinant expression systems can be used to produce fully synthetic peptides [6].
Mechanism of antimicrobial activity of antimicrobial peptides
It has been suggested that the interaction and action of AMPs with target cells depends on the two factors: the cell surface which is the classic and large acting mechanism and the amino acid composition of AMPs [8]. Furthermore, researchers have found that there have two main kinds of AMPs namely membrane-active AMPs and intracellular-active AMPs (Figure 1).
Figure 1.
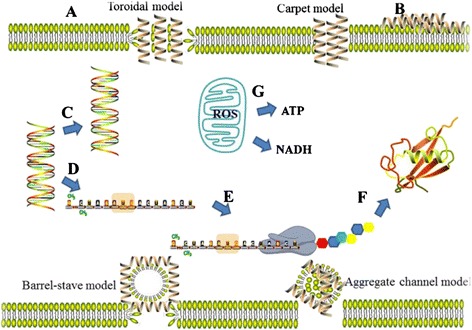
The diverse mechanistic modes of action for antimicrobial peptides. The figure means two main kinds of AMPs namely membrane-active AMPs and intracellular-active AMPs. The mechanism of membrane-active AMPs includes the “barrel-stave”, “toroidal”, “carpet” and “aggregate channel” models (A); B-G means the mechanism of intracellular-active AMPs. A, membrane activity contains Toroidal model, Carpet model, Barrel-stave model and Aggregate channel model; B, Inhibition of enzymes necessary for linking of cell wall structural proteins; C, Inhibition of DNA synthesis; D, Inhibition of RNA synthesis; E, Inhibition of ribosomal function and protein synthesis; F, Blocking of chaperone proteins necessary for proper folding of proteins; G, inhibition of cellular respiration and induction of ROS formation and damage of mitochondrial cell membrane integrity and efflux of ATP and NADH.
Membrane-active antimicrobial peptides
In order to explain membrane disruption by AMPs, researchers have proposed many models including the “barrel-stave”, “toroidal”, “carpet” and “aggregate channel” models. Early in 1977, Ehrenstein and Lecar [9] proposed the “barrel-stave model” which suggests that peptides directly insert into the lipid core of the target membrane to form trans-membrane pores. In the “toroidal model”, peptide molecules are inserted into the membrane forming a bundle, inducing the lipid monolayers to continuously bend through the pore [9]. The “carpet model”, suggests that AMPs use a detergent-like action to cover the membrane surface in order to affect its architecture [10,11]. In the “aggregate channel model”, the peptides insert into the membrane and then cluster into unstructured aggregates that span the membrane. These aggregates are proposed to have water molecules associated with them providing channels for leakage of ions and possibly larger molecules through the membrane [12] (Figure 1).
Intracellular-active antimicrobial peptides
Cell membrane permeabilization by AMPs was thought to be the primary mechanism of killing. However, there is increasing evidence to prove that some AMPs can interact with an array of intracellular targets including DNA, RNA and protein to kill their target cells, but not damage the cell membrane. AMPs can directly prevent DNA, RNA and protein synthesis, as well as cell wall synthesis and proteases of microbes by means of direct penetration and endocystosis to enter the cells [13] (Figure 1). For example, PR-39 from pig intestines can act like a proteolytic agent and suppress protein and DNA synthesis to kill bacteria [14], while seminaplasmin can inhibit RNA polymerase and stop RNA synthesis completely at very low concentrations [15].
The applications of antimicrobial peptides in swine nutrition
Some AMPs, including antimicrobial peptide A3, P5, colicin E1, cecropin AD, and cipB-lactoferricin-lactoferrampin (cipB-LFC-LFA) have been shown to have beneficial effects on performance, nutrient digestibility, intestinal morphology as well as intestinal and fecal microflora. Their activities are summarized in Table 1.
Table 1.
Summary of studies showing the applications of AMPs in swine nutrition
| Antimicrobial peptide and dose | Animal and treatments | Application effects | References |
|---|---|---|---|
| Antimicrobial peptide-A3 (AMP-A3); 60 or 90 mg/kg | Weanling piglets fed with basal diet + 1.5 g apramycin/kg diet and basal diet supplemented with 0, 60 and 90 mg AMP-A3/kg diet in 2 phases (d 0–14 post-weaning: 14.28 MJ/kg ME and 15.5 g/kg lysine; d 15–28 post-weaning: 14.11 MJ/kg ME and 13.5 g/kg lysine) | Has beneficial effects on performance, total tract apparent digestibility of nutrients, intestinal morphology and intestinal and fecal microflora | Yoon et al. 2012 [19] |
| Antimicrobial peptide-P5 (AMP-P5); 40 or 60 mg/kg | Weanling piglets fed with basal diet, basal diet + 1.5 g/kg apramycin, basal diet + 40 mg/kg AMP-P5 and basal diet + 60 mg/kg AMP-P5 in 2 phases (d 0–14 post-weaning: 14.28 MJ/kg ME and 15.5 g/kg lysine; d 15–28 post-weaning: 14.11 MJ/kg ME and 13.5 g/kg lysine) | Improves the performance and apparent total tract digestibility of nutrients and reduces coliforms | Yoon et al. 2013 [4] |
| Synthetic antimicrobial peptide-A3 or P5 (AMP-A3 and P5); 60 mg AMP-A3 or 60 mg AMP-P5/kg | Weanling piglets fed with basal diet, basal diet + 150 mg/kg avilamycin, basal diet + 60 mg/kg AMP-A3 and basal diet + 60 mg/kg AMP-P5 for 28 days | Improves the performance, nutrient digestibility, intestinal morphology and to reduces pathogenic bacteria | Yoon et al. 2014 [20] |
| Antimicrobial peptide colicin E1; 11 or 16.5 mg/kg | Weaned pigs fed with diets containing 0, 11, or 16.5 mg colicin E1/kg diet and were orally inoculated with 1 x 109 CFU of each of two F18-positive E. coli strains | Improves the performance, reduces incidence of postweaning diarrhe | Cutler et al. 2007 [21] |
| Antimicrobial peptide cecropin AD; 400 mg/kg | Weaned barrows fed with basal diet or similar diets supplemented with antibiotics (100 mg/kg kitasamycin plus 800 mg/kg colistin sulfate) or 400 mg/kg cecropin AD and were orally challenged with 109 CFU/mL of E. coli K88 | Enhances pig performance through increasing immune status and nitrogen and energy retention as well as reducing intestinal pathogens | Wu et al. 2012 [27] |
| cipB-lactoferricin-lactoferrampin (cipB-LFC-LFA); 100 mg/kg | Weanling piglets were challenged with enterotoxigenic E. coli and randomly assigned to four treatment groups fed a maize–soyabean meal diet containing either no addition, cipB at 100 mg/kg, cipB-LFC-LFA at 100 mg/kg or colistin sulfate at 100 mg/kg for 3 weeks | Improves performance through an antibacterial effect, the regulation of immune function, improvement of the absorption of Fe and a reduction in the incidence of diarrhea | Tang et al. 2009 [5] |
| Recombinant Lactoferrampin-Lactoferricin; 100 mg/kg | Weanling piglets fed with basal diet, basal diet + 0.1 g /kg lactoferrampin-lactoferricin and basal diet + 0.1 g /kg chlortetracycline for 21 days | Improves performance and affects serum parameters | Tang et al. 2012 [17] |
| Composite antimicrobial peptides (CAP, consist mainly of antibacterial lactoferrin peptides, along with plant defensins and active yeast); 400 mg/kg | Weanling piglets fed with basal diet, basal diet + 0.4% CAP, basal diet + 4 mg/kg deoxynivalenol, and basal diet + 4 ppm deoxynivalenol + 0.4% CAP for 30 days | Improves feed efficiency, immune function, and antioxidation capacity and alleviates organ damage | Xiao et al. 2013 [29] |
| Composite antimicrobial peptides (CAP, consist mainly of antibacterial lactoferrin peptides, along with plant defensins and active yeast); 400 mg/kg | Weanling piglets fed with basal diet, basal diet + 0.4% CAP, basal diet + 4 mg/kg deoxynivalenol, and basal diet + 4 ppm deoxynivalenol + 0.4% CAP for 30 days | Improves intestinal morphology and intestinal epithelial cell proliferation and protein synthesis; May repair the intestinal injury induced by DON | Xiao et al. 2013 [30] |
| Composite antimicrobial peptides (CAP, consist mainly of antibacterial lactoferrin peptides, along with plant defensins and active yeast); 400 mg/kg | Weanling piglets fed with basal diet, basal diet + 0.4% CAP, basal diet + 4 mg/kg deoxynivalenol, and basal diet + 4 ppm deoxynivalenol + 0.4% CAP for 30 days | Attenuate the metabolic disturbances in AA, lipid, and energy metabolism induced by DON. | Xiao et al. 2015 [31] |
| A mixture of lactoferrin, cecropin, defensin, and plectasin | Pigs fed with basal diet, basal diet + 2.0 g/kg of AMPs and basal diet +3.0 g/kg of AMPs for 32 days | Improves performance, reduces the incidence of diarrhea, and increases the survival rate of weaned pigs | Xiong et al. 2014 [32] |
Antimicrobial peptides can promote the performance of pigs
Antimicrobial lactoferrin peptides are one of the most prevalent AMPs used in swine nutrition. It has been demonstrated that dietary supplementation with recombinant lactoferrampin-lactoferricin (produced by the Institute of Subtropical Agriculture, Chinese Academy of Science which obtained through the expression of the lactoferampin-lactoferricin gene in the expression host P. pastoris (KM71) XS10 [16]) increased the final body weight and the average daily gain (ADG) of piglets by 13.3 and 29.3%, respectively while decreasing feed conversion by 11.5% [16,17]. Tang et al. [16,17] showed that piglets supplemented with cipB-LFC-LFA had higher ADG and ADFI than pigs fed control diets A mixture of AMPs including lactoferrin, cecropin, defensin, and plectasin was shown to enhance ADG, ADFI and G:F on 5 farms [18]. Growth promoting effects of the antimicrobial peptides A3 and P5 were also observed [4]. Increasing the levels of dietary AMP-A3 from 0 to 90 mg/kg in diets linearly improved ADG [19] while dietary supplementation with 60 mg/kg AMP-P5 increased ADG, ADFI and G:F [4], but the effects of AMP-A3 or P5 did not surpass that of a positive control treatment supplemented with 150 mg/kg avilamycin [20].
Dietary inclusion of Colicin E1 had a significant effect on pig performance in that pigs fed the control diet gained an average of 380 g, while pigs receiving 11 and 16.5 mg Colicin E1 per kg of diet gained 540 and 940 g, respectively [21]. However, the joint use of antibacterial peptide and Zn-Met did not show any synergistic effects on pig performance [22].
The effects of AMPs on performance can be explained on the basis of their antimicrobial activity. For example, Colicins E1 and N have been shown to inhibit the activities of E. coli strains that caused post-weaning diarrhea and edema disease in pigs [23]. The improvement in performance can also be related to improvements in nutrient digestibility [4,20,24]. Yoon et al. [4,19,20] found that pigs diets supplemented with AMP-A3 or P5 showed an increase in the apparent total tract digestibility of dry matter, crude protein and gross energy.
Antimicrobial peptides can enhance the immune status of pigs
AMPs are important components of the host’s defense system and are effector molecules of innate immunity with direct antimicrobial and immune mediator function [2,25,26]. Tang et al. [5] found that dietary supplementation with cipB–lactoferricin–lactoferrampin increased serum IgA and IgG but reduced serum IgM. The researchers from National Feed Engineering Technology Research Center (Beijing, China) prepared antimicrobial peptide cecropin AD using cecropin A and cecropin D isolated from the silkworm Hyalophora cecropia and added it to weaned piglets challenged with E. coli [27]. The results show that cecropin AD could increase levels of secretory IgA in jejunum and serum IgA, IgG, interleukin-1β and interleukin-6 [27]. AMPs can influence the adaptive immune system, either directly or indirectly via alteration of the gut microflora [5]. This was confirmed by results showing that dietary AMP-A3 or P5 decreased fecal Clostridium spp. and coliforms, as well as decreasing ileal and cecal total anaerobic bacteria, Clostridium spp. and coliforms [20].
Antimicrobial peptides can improve the intestinal health of pigs
A toxin produced by pathogenic bacteria in the gut can cause inflammation of the intestinal mucosa and diarrhea associated with morphological changes in the small intestine, such as shortening of the villi and an increase in crypt depth [28]. The antibacterial action of AMPs provides an effective support for normal intestinal morphology and function. Tang et al. [5] found that lactoferrampin-lactoferricin increased the height of the villi in the jejunum and ileum as well as the villus height: crypt depth ratio in the jejunum and ileum, which may be related to the fact that LFC-LFA can decrease the concentration of E. coli and increase lactobacilli and bifidobacteria in the gut. Similar results were observed in pigs following AMP-A3 [19] or cecropin AD [27] treatment. In addition, dietary supplementation with AMPs induced lower serum D-lactate concentrations [17] that increased intestinal permeability and enhanced the efficiency of absorption and utilization of nutrients.
Antimicrobial peptides alleviate the toxic effects of deoxynivalenol (DON) in pigs
Recently, we found that AMPs played a protective effect in piglets challenged with DON [29]. The composite antimicrobial peptide GLAM®180# used in our studies contains antibacterial lactoferrin peptides, plant defensins and active yeast and these three bioactive components have been shown to have a positive effect on growth and health of animals. Feeding 0.4% GLAM®180# to piglets challenged with diets containing 4 mg/kg DON improved overall feed efficiency (Table 2), promoted blood circulation, alleviated organ damage, and reduced DON toxicity [29].
Table 2.
Effects of composite antimicrobial peptides on the performance of piglets (12–26 kg) challenged with deoxynivalenol 1,2
| Item | Diets | SEM | P -value | |||
|---|---|---|---|---|---|---|
| NC | CAP | DON | DON + CAP | |||
| ADG, kg | ||||||
| Day 0 to 15 | 0.32 | 0.24 | 0.30 | 0.30 | 0.01 | 0.09 |
| Day 15 to 30 | 0.64a | 0.66a | 0.40b | 0.48b | 0.02 | <0.01 |
| Day 0 to 30 | 0.48a | 0.45a | 0.35b | 0.39b | 0.01 | <0.01 |
| ADFI, kg | ||||||
| Day 0 to 15 | 0.67a | 0.56b | 0.68a | 0.63ab | 0.02 | 0.02 |
| Day 15 to 30 | 1.33a | 1.10b | 1.07b | 1.04b | 0.03 | <0.01 |
| Day 0 to 30 | 1.00a | 0.83b | 0.87b | 0.84b | 0.02 | <0.01 |
| G:F | ||||||
| Day 0 to 15 | 0.47 | 0.42 | 0.44 | 0.49 | 0.02 | 0.54 |
| Day 15 to 30 | 0.49b | 0.61a | 0.38c | 0.47bc | 0.02 | <0.01 |
| Day 0 to 30 | 0.48ab | 0.55a | 0.40b | 0.47a | 0.01 | <0.01 |
Xiao et al. 2013 [29].
1NC: Basal diet. CAP: Basal diet + 0.4% composite antimicrobial peptide, DON: Basal diet + 4 mg/kg DON, DON + CAP: Basal diet + 4 mg/kg DON + 0.4% composite antimicrobial peptide.
2 n = 7. a-c Values with different letters within the same row are significantly different (P < 0.05).
As indicators of intestinal morphology and function, the serum D-lactate and diamine oxidase content were lower but the villous height/crypt depth (Table 3) and the proliferating cell nuclear antigen (PCNA) labeling indexes in the jejunum and ileum were greater in piglets fed DON + GLAM®180# treatments than those in the DON treatment alone. In addition, GLAM®180# increased the protein levels of phosphorylated Akt, mTOR and 4E-binding protein 1 in the jejunum of piglets. The results indicate that GLAM®180# improved intestinal morphology and promoted intestinal epithelial cell proliferation and protein synthesis [30]. The combined results of 1H-NMR and LC-MS/MS showed the serum concentrations of HDL, unsaturated lipids, proline, citrate and fumarate were greater while those of glycoprotein, urea, TMAO, glycine and lactate were lower, in the DON + CAP group compared to those in the DON group, which indicated GLAM®180# could attenuate the metabolic disturbances in AA, lipid, and energy metabolism induced by DON [31]. The application of AMPs in DON challenged piglets demonstrates that GLAM®180# can alleviate the toxic effect of DON on pigs.
Table 3.
Effects of composite antimicrobial peptides on the jejunal and ileal morphology of piglets challenged with d eoxynivalenol 1,2
| Item | Diets | SEM | P -value | |||
|---|---|---|---|---|---|---|
| NC | CAP | DON | DON + CAP | |||
| Jejunum | ||||||
| Villus height, μm | 240.01 | 239.08 | 197.56 | 221.87 | 7.05 | 0.23 |
| Crypt depth, μm | 126.39ab | 117.03b | 145.14a | 110.48b | 6.88 | 0.04 |
| Villus height: Crypt depth | 1.92 | 2.06 | 1.39 | 2.05 | 0.11 | 0.08 |
| Goblet cell number | 11.50 | 11.00 | 17.67 | 12.00 | 1.20 | 0.26 |
| Lymphocyte number | 195.00b | 198.50b | 256.33a | 204.50b | 9.26 | <0.01 |
| Ileum | ||||||
| Villus height, μm | 263.20a | 240.15a | 170.98b | 185.08b | 10.31 | <0.01 |
| Crypt depth, μm | 117.73 | 120.05 | 109.45 | 104.73 | 4.36 | 0.56 |
| Villus height: Crypt depth | 2.32a | 2.06ab | 1.57c | 1.81ab | 0.09 | 0.04 |
| Goblet cell number | 22.33 | 16.00 | 19.00 | 18.50 | 1.36 | 0.38 |
| Lymphocyte number | 181.25c | 154.75d | 232.00a | 204.00b | 7.70 | <0.01 |
Xiao et al. 2013 [30].
1NC: Basal diet. CAP: Basal diet + 0.4% composite antimicrobial peptide, DON: Basal diet + 4 mg/kg DON, DON + CAP: Basal diet + 4 mg/kg DON + 0.4% composite antimicrobial peptide.
2 n = 7. a-d Values with different letters within the same row are significantly different (P < 0.05).
Conclusions
Due to their broad spectrum of activity against several species of bacteria, fungi, protozoa, and enveloped virus, AMPs show beneficial effects on performance, nutrient digestibility, intestinal morphology as well as intestinal and fecal microflora in pigs. With the development of technology, the cost of addition of AMPs is gradually reduced, especially in swine production. Although most AMPs did not provide equal effects to that of antibiotics in swine nutrition, they have considerable potential as an alternative for antibiotics in rations fed to swine.
Acknowledgement
This work was supported by the National Natural Science Foundation of China (No. 31330075; 31372326).
Footnotes
Competing interests
The authors declare they have no competing interests.
Authors’ contributions
HX, FYS, MMW, WKR, BET, and YLY collected papers and drafted the manuscript. HX, FYS, and BET co-wrote the paper: All authors read and approved the final manuscript.
Authors’ information
Hao xiao and Fangyuan Shao are joint first authors.
Contributor Information
Hao Xiao, Email: 380585550@qq.com.
Fangyuan Shao, Email: shaogz2011@sina.com.
Miaomiao Wu, Email: miao1988613@tom.com.
Wenkai Ren, Email: renwenkai19@126.com.
Xia Xiong, Email: xx@isa.ac.cn.
Bie Tan, Email: bietan@isa.ac.cn.
Yulong Yin, Email: yinyulong@isa.ac.cn.
References
- 1.Diez-Gonzalez F. Applications of bacteriocins in livestock. Curr Issues Intest Microbiol. 2007;8:15–23. [PubMed] [Google Scholar]
- 2.Reddy KV, Yedery RD, Aranha C. Antimicrobial peptides: premises and promises. Int J Antimicrob Agents. 2004;24:536–47. doi: 10.1016/j.ijantimicag.2004.09.005. [DOI] [PubMed] [Google Scholar]
- 3.Wang Y, Lu Z, Feng F, Zhu W, Guang H, Liu J, et al. Molecular cloning and characterization of novel cathelicidin-derived myeloid antimicrobial peptide from Phasianus colchicus. Dev Comp Immunol. 2011;35:314–22. doi: 10.1016/j.dci.2010.10.004. [DOI] [PubMed] [Google Scholar]
- 4.Yoon JH, Ingale SL, Kim JS, Kim KH, Lohakare J, Park YK, et al. Effects of dietary supplementation with antimicrobial peptide-P5 on growth performance, apparent total tract digestibility, faecal and intestinal microflora and intestinal morphology of weanling pigs. J Sci Food Agric. 2013;93:587–92. doi: 10.1002/jsfa.5840. [DOI] [PubMed] [Google Scholar]
- 5.Tang Z, Yin Y, Zhang Y, Huang R, Sun Z, Li T, et al. Effects of dietary supplementation with an expressed fusion peptide bovine lactoferricin-lactoferrampin on performance, immune function and intestinal mucosal morphology in piglets weaned at age 21 d. Br J Nutr. 2009;101:998–1005. doi: 10.1017/S0007114508055633. [DOI] [PubMed] [Google Scholar]
- 6.Bahar AA, Ren D. Antimicrobial peptides. Pharmaceuticals. 2013;6:1543–75. doi: 10.3390/ph6121543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang G. Improved methods for classification, prediction, and design of antimicrobial peptides. Methods Mol Biol. 2015;1268:43–66. doi: 10.1007/978-1-4939-2285-7_3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Guilhelmelli F, Vilela N, Albuquerque P, Derengowski Lda S, Silva-Pereira I, Kyaw CM. Antibiotic development challenges: the various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front Microbiol. 2013;4:353. doi: 10.3389/fmicb.2013.00353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yang L, Harroun TA, Weiss TM, Ding L, Huang HW. Barrel-stave model or toroidal model? A case study on melittin pores. Biophys J. 2001;81:1475–85. doi: 10.1016/S0006-3495(01)75802-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pouny Y, Rapaport D, Mor A, Nicolas P, Shai Y. Interaction of antimicrobial dermaseptin and its fluorescently labeled analogues with phospholipid membranes. Biochemistry. 1992;31:12416–23. doi: 10.1021/bi00164a017. [DOI] [PubMed] [Google Scholar]
- 11.Rotem S, Mor A. Antimicrobial peptide mimics for improved therapeutic properties. Biochim Biophys Acta. 2009;1788:1582–92. doi: 10.1016/j.bbamem.2008.10.020. [DOI] [PubMed] [Google Scholar]
- 12.Li Y, Xiang Q, Zhang Q, Huang Y, Su Z. Overview on the recent study of antimicrobial peptides: origins, functions, relative mechanisms and application. Peptides. 2012;37:207–15. doi: 10.1016/j.peptides.2012.07.001. [DOI] [PubMed] [Google Scholar]
- 13.Mayor S, Pagano RE. Pathways of clathrin-independent endocytosis. Nat Rev Mol Cell Biol. 2007;8:603–12. doi: 10.1038/nrm2216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Boman HG, Agerberth B, Boman A. Mechanisms of action on Escherichia coli of cecropin P1 and PR-39, two antibacterial peptides from pig intestine. Infect Immun. 1993;61:2978–84. doi: 10.1128/iai.61.7.2978-2984.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Scheit KH, Reddy ES, Bhargava PM. Seminaplasmin is a potent inhibitor of E. coli RNA polymerase in vivo. Nature. 1979;279:728–31. doi: 10.1038/279728a0. [DOI] [PubMed] [Google Scholar]
- 16.Tang XS, Tang ZR, Wang SP, Feng ZM, Zhou D, Li TJ, et al. Expression, purification, and antibacterial activity of bovine lactoferrampin-lactoferricin in Pichia pastoris. Appl Biochem Biotechnol. 2012;166:640–51. doi: 10.1007/s12010-011-9455-0. [DOI] [PubMed] [Google Scholar]
- 17.Tang X, Fatufe AA, Yin YL, Tang ZR, Wang SP, Liu ZQ, et al. Dietary supplementation with recombinant lactoferrampin-lactoferricin improves growth performance and affects serum parameters in piglets. J Anim Vet Adv. 2012;11:2548–55. doi: 10.3923/javaa.2012.2548.2555. [DOI] [Google Scholar]
- 18.Xiong X, Yang HS, Li L, Wang YF, Huang RL, Li FN, et al. Effects of antimicrobial peptides in nursery diets on growth performance of pigs reared on five different farms. Livestock Sci. 2014;167:206–10. doi: 10.1016/j.livsci.2014.04.024. [DOI] [Google Scholar]
- 19.Yoon JH, Ingale SL, Kim JS, Kim KH, Lee SH, Park YK, et al. Effects of dietary supplementation of antimicrobial peptide-A3 on growth performance, nutrient digestibility, intestinal and fecal microflora and intestinal morphology in weanling pigs. Anim Feed Sci Technol. 2012;177:98–107. doi: 10.1016/j.anifeedsci.2012.06.009. [DOI] [Google Scholar]
- 20.Yoon JH, Ingale SL, Kim JS, Kim KH, Lee SH, Park YK, et al. Effects of dietary supplementation of synthetic antimicrobial peptide-A3 and P5 on growth performance, apparent total tract digestibility of nutrients, fecal and intestinal microflora and intestinal morphology in weanling pigs. Livestock Sci. 2014;159:53–60. doi: 10.1016/j.livsci.2013.10.025. [DOI] [Google Scholar]
- 21.Cutler SA, Lonergan SM, Cornick N, Johnson AK, Stahl CH. Dietary inclusion of colicin e1 is effective in preventing postweaning diarrhea caused by F18-positive Escherichia coli in pigs. Antimicrob Agents Chemother. 2007;51:3830–5. doi: 10.1128/AAC.00360-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang JH, Wu CC, Feng J. Effect of dietary antibacterial peptide and zinc-methionine on performance and serum biochemical parameters in piglets. Czech J Anim Sci. 2011;56:30–6. [Google Scholar]
- 23.Stahl CH, Callaway TR, Lincoln LM, Lonergan SM, Genovese KJ. Inhibitory activities of colicins against Escherichia coli strains responsible for postweaning diarrhea and edema disease in swine. Antimicrob Agents Chemother. 2004;48:3119–21. doi: 10.1128/AAC.48.8.3119-3121.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dubos RJ. Studies on a bactericidal agent extracted from a soil bacillus: Ii. Protective effect of the bactericidal agent against experimental pneumococcus infections in mice. J Exp Med. 1939;70:11–7. doi: 10.1084/jem.70.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Peters BM, Shirtliff ME, Jabra-Rizk MA. Antimicrobial peptides: primeval molecules or future drugs? PLoS Pathog. 2010;6:e1001067. doi: 10.1371/journal.ppat.1001067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Beisswenger C, Bals R. Functions of antimicrobial peptides in host defense and immunity. Curr Protein Pept Sci. 2005;6:255–64. doi: 10.2174/1389203054065428. [DOI] [PubMed] [Google Scholar]
- 27.Wu S, Zhang F, Huang Z, Liu H, Xie C, Zhang J, et al. Effects of the antimicrobial peptide cecropin AD on performance and intestinal health in weaned piglets challenged with Escherichia coli. Peptides. 2012;35:225–30. doi: 10.1016/j.peptides.2012.03.030. [DOI] [PubMed] [Google Scholar]
- 28.Swidsinski A, Ladhoff A, Pernthaler A, Swidsinski S, Loening-Baucke V, Ortner M, et al. Mucosal flora in inflammatory bowel disease. Gastroenterology. 2002;122:44–54. doi: 10.1053/gast.2002.30294. [DOI] [PubMed] [Google Scholar]
- 29.Xiao H, Wu MM, Tan BE, Yin YL, Li TJ, Xiao DF, et al. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: I. Growth performance, immune function, and antioxidation capacity. J Anim Sci. 2013;91:4772–80. doi: 10.2527/jas.2013-6426. [DOI] [PubMed] [Google Scholar]
- 30.Xiao H, Tan BE, Wu MM, Yin YL, Li TJ, Yuan DX, et al. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: II. Intestinal morphology and function. J Anim Sci. 2013;91:4750–6. doi: 10.2527/jas.2013-6427. [DOI] [PubMed] [Google Scholar]
- 31.Xiao H, Xiao H, Wu MM, Tan B, Li T, Ren WK, et al. Metabolic profiles in the response to supplementation with composite antimicrobial peptides in piglets challenged with deoxynivalenol. J Anim Sci. 2015;93:1114–23. doi: 10.2527/jas.2014-8229. [DOI] [PubMed] [Google Scholar]