

Abstract
The goal of this review is to integrate - in its two parts - the considerable amount of information that has accumulated during these recent years over the morphology, biology and functions of astrocytes - first part - and to illustrate the active role of these cells in pathophysiological processes implicated in various psychiatric and neurologic disorders – second part.
Keywords: Astrocytes, Reactive astrogliosis, Molecular mechanisms, Therapeutic targets
Introduction
Increasing research interest aroused by astrocytes over the past few years led to a dramatic evolution of the concept regarding their structure and function. Ubiquitously present in all regions of the central nervous system (CNS), astrocytes are specialized glial cells, providing structural and functional support for neurons.
Although considered for more than 100 years as a homogenous cell population, it is known today that glia encompasses various morphological entities that coexist; each of these populations are characterized by a particular molecular signature and specific functions related to their microenvironment. Moreover, dysfunctions of astrocytes might contribute to CNS pathological remodelling and disease [1].
Review
Short history
The concept of neuroglia, introduced by Rudolf Virchow in 1858, described a connective substance of the brain, represented most likely by “fibers and intercellular masses”. Otto Deiters, a German scientist, was the first who, in the second half of the 19th century, drew the astrocytes as stellate cells; later, Jacob Henle and Friedrich Merkel observed the network formed by the astrocytes processes within the grey matter [2]. Yet it was Camillo Golgi (1872) the first who detailed and described the morphology of glial cells by using the silver-chromate technique (a black staining reaction); he observed that some glial cells (known today as protoplasmic astrocytes) displayed endfeet on their processes, attached to the blood vessels. His theory postulated that there was a link between the morphology and function of astrocytes in the CNS; regarded as the “glue” of the brain, glial cells established an interconnection between vessels and parenchyma, therefore being responsible for metabolic exchanges. In 1893 Michael von Lenhossek contrived the term “astrocyte” that illustrated the morphology of these cells. The origin of this term arouse from a combination of the latin word for stars, astra, with the word for cell, cyte, thus a star-shaped cell [3]. Astrocytes were further classified into protoplasmic (found in the grey matter) and fibrous (within the white matter) [2–4].
At the beginning of the 20th century the morphological heterogeneity of the CNS glia was definitely set. However, only when Santiago Ramόn y Cajal (1913) has developed the gold chloride-sublimate staining technique, the first specific stain for astrocytes, this diversity was acknowledged. Cajal is considered the promoter of the future stem properties of neuroglia since, using this method, he proved that astrocytes originate from radial glia and undergo cell division in the adult brain. Numerous functions of astrocytes (e.g. neuronal nutrition and metabolism, nervous tissue homeostasis, brain cytoarchitecture, glial scar formation) were further determined, relying on Cajal’s histological research, rendering astrocytes essential brain “homeostatic cells” either in normal or pathologic conditions [5].
Yet, the gains regarding the functions of astrocytes were shadowed by the lack of adequate techniques that could have promoted them, versus neurons of which value was overstated by the neuronal doctrine [2].
Phylogenetic evolution
From the phylogenetical point of view, the organization of a centralized nervous system was marked by the appearance of astrocytes [5].
An interesting aspect is the constant augmentation the astrocytes/neurons ratio that parallelized the evolution of the brain (about 0.16 in nematodes to 0.33 in rodents, and reaching up to 1.65 astrocytes per neuron in the human cortex) [6]. It is considered that, in the human brain, to each neuron correspond 10 glial cells. In smaller creatures’ brain, the number of glial cells corresponding to a neuron is significantly reduced [7].
The primordial astrocytes performed a wide range of functions in the development of the nervous system. In nematodes, the astrocytes are not only involved in neuronal development, but also enable the sensory functions [5]. Moreover, the astrocytes’ performances improve with the evolutionary stages. For example, in arthropods glial cells fulfill an additional role, organizing the neurons in functional definite nervous centers [5]. In crustaceans, insects and cephalopods, even in some vertebrates (sharks), the astrocytes form the blood-brain barrier (BBB) or the hemolymph-brain barrier (HBB) isolating the nervous tissue from the rest of the body [5]. Primordial astrocytes also envelop the axons therefore being the predecessors of the myelin forming cells; the astroglial sheath of the axons improves the propagation of the action potential [5]. In higher vertebrates, astrocytes’ role in maintaining the BBB function is completed by the endothelial cells. Besides, in this stage of evolution, astrocytes specialize for the defensive function [5]. In humans, astrocytes achieve their greatest morphologic and functional complexity. For example, neocortex humans astrocytes compared to those of rodents, are 2.5 times larger, their processes are 10 times more numerous and they display particular histological features; the action potential velocity is also 4 times higher [7].
Stem cells and astrocytes differentiation
Initially astrocytes were identified due to their star-shaped morphology and presence of the glial fibrils. Nowadays these features are almost outdated.
The diversity of astrocytes is justified by two main factors: the heterogeneity of glial precursors and the various pathways of specific differentiation, both being influenced by the extracellular environment. Recent in vitro studies reported that growth factors levels activate in astrocytes the gene expression and regulate the transcription factors so that the subsets of progenitors are spontaneously engaged in different pathways of development [8]. During their differentiation, between the glial precursors and the microenvironment there is a mutual influence: cells secrete various soluble factors, and, on the other hand, the extracellular matrix (ECM) molecules (e.g. lectican and tenascins family) have the ability to stimulate or to inhibit cells proliferation, maturation and migration [9, 10]. Thus, in his study, Haas C. et al. in 2012, observed that by treating GRP in vitro with specific culture media, different astrocytic phenotypes were obtained (e.g. A2B5-/GFAP+ with a flat morphology fibroblast-like when treated with fetal bovine serum and A2B5+/GFAP+ star-shaped astrocytes when treated with both basic fibroblast growth factor (bFGF) and ciliary neurotrophic factor (CNTF) [8].
For example, if we consider a multipotent stem cell as a source of astrocytes, but initially, this cell has produced neuronal precursors, the turn towards glial differentiation implies a multi-step process. At first, a specific receptor on the surface of the multipotent stem cell modifies its structure to gain affinity for growth factors such as: fibroblast growth factor (FGF) and epidermal growth factor (EGF); then, the resulting glial precursor is subjected to the action of signalling molecules (e.g. CNTF, bone morphogenetic proteins (BMF) and EGF) that will control and continue its maturation [9, 10].
However, further research is needed in order to identify the heterogeneous subpopulations of astrocytes progenitors and accurately characterise them by new antigenic markers, physiological properties or molecular profiles [1].
At present, three distinct pools of glial progenitors have been described in the germinal niches of the cerebral cortex: a) radial cells of the ventricular zone b) postnatal glial progenitor cells of the subventricular zone and c) glial-restricted precursors (GRP) - also found in the embryonic spinal cord (see Table 1) [3, 8].
Table 1.
Ontogenetic astrocyte progenitor pools
| Radial glia | Postnatal glial progenitor cells | Glial restricted precursors | ||
|---|---|---|---|---|
| Origin | Neuroepithelial cells [1] | • Radial glia [11, 12] | Neuroepithelial cells skipping the radial glia stage [13, 14] | |
| • Dlx2 (distal-less homeobox 2) [3] | ||||
| • Local glial progenitors [1] | ||||
| Location | Ventricular zone [1] | • Subventricular zone | • Embryonic spinal cord [8] | • Optic nerve [8] |
| • Dorso-lateral subventricular zone | ||||
| • Marginal zone [1, 11, 12] | ||||
| Characteristics | Multipotential cells [11, 12] | • Multipotential cells | Tripotential cells [8] | • Bipotential cells O-2A, O-2A/OPC [8, 15] |
| • Bipotential cells [3] | ||||
| Roles | • Progenitors for neurons and glial cells | • Intermediate progenitors for astrocytes and oligodendrocytes [3] | • Promote neuroprotection | • Tumor genesis (oligoastrocytomas, multiform glioblastomas) [15] |
| • Guidance of neuronal migration [11, 12] | • Reduction of glial scar | |||
| • Formation and axonal growth [8] | ||||
| Type of resulting astrocytes | • Star shaped specialised cortical astrocytes | • Cortical astrocytes | • Self-renewal | • Astrocytes type 2 and oligodendrocytes (in vitro) |
| • White matter astrocytes | • Astrocytes types 1, 2 and | |||
| • Bergmann glia in the cerebellum [3, 16] | • Oligodendrocytes [3] | • Oligodendrocytes [8] | • Oligodendrocytes (in vivo) [8, 15] | |
The grey matter protoplasmic astrocytes are mostly generated by embryonic radial glia but also from the intermediate progenitors arisen from neonatal subventricular zones. Due to their different origin, the two populations of astrocytes will display different patterns of gene expression, which will enable potential different functions. The white matter fibrous astrocytes originate, instead, mainly from neonatal subventricular zone progenitors [1].
Astrocytes-like neural progenitors
An unexpected finding in the astrocyte research is the identification in the adult neurogenic zones - subventricular zone (SVZ) and subgranular zone (SGZ) - of a subtype of astrocytes considered to be the local stem cells. Regarded as mature astrocytes due to the expression of GFAP and glycogen granules, these cells unusually display features of both radial glia and neural progenitors (e.g. synaptic mediators’ release) [1].
It was demonstrated that the specific pro-neural genes (e.g. neurogenin-2 and Mash1) enable these astrocytes to regain their stem cells properties being able to differentiate into neurons [1]. Additionally, the embryonic extracellular matrix molecules present in the neurogenic niche are capable to maintain these cells’ “stemness” [1, 17].
In the adult SVZ and SGZ, two distinct population of neural progenitors (multipotent neural stem cells) express GFAP [1, 18–20]. The SVZ progenitors and give rise to neuroblasts which migrate to the olfactory bulb (to become olfactory interneurons) [1, 19–22]. GFAP-expressing cells found in the SVZ are also been referred to as astrocytes-like cells or B cells. From the histological point of view, these cells are irregular in shape, filling in the spaces between neighbouring cells; their cytoplasm is pale with few organelles (e.g. free ribosomes) but numerous intermediate filaments; the nuclei are also irregular due to the invaginations on their surface. There are significant differences between the two types of SVZ astrocytes. Type 1 (i.e. B1 cells) are larger, with euchromatic nuclei and are located in the proximity of the ependymal cells. Type 2 (i.e. B2 cells) are smaller, with hyperchromatic nuclei and are mostly adjacent to the striatal parenchyma. The SGZ neural progenitors generate newborn granular neurons [1].
Another type of stem cell which expresses GFAP can be found in the adult SVZ but it is not certain that these adult stem cells are, in fact, astrocytes. They have different molecular features, because they express nestin (an intermediate filament), that characterise only embryonic astrocytes, reactive astrocytes or neuroblasts and intermediate progenitors [1].
Considering the high plasticity of astrocytes, the GFAP expressing cells in the neurogenic niche can simultaneously behave as both astrocytic and neural stem cells [1].
Astrocytic markers and stains
Important advances in technologies to study the nervous tissue enabled the knowledge of astrocytes characteristics (see Table 2), Figures 1, 2 and 3. (All images presented in here, are microphotographs of human brain samples prelevated by autopsy in compliance with the Protocol elaborated by the Ethics Committee of “Iuliu Hatieganu” University of Medicine and Pharmacy Cluj-Napoca).
Table 2.
Astrocytic markers and stains
| Procedures | Characteristics | Results | Observation |
|---|---|---|---|
| Hematoxylin and eosin stain (H-E) [23] | Routine staining for basic morphology | Nuclear details | • Astrocytes are difficult to identify (nuclei: small, pale, ovoidal, euchromatic and centrally situated, are mimicking those of small neurons; cytoplasm and cellular processes are undifferentiated from those of neighbouring neurons) |
| Cytoplasm extracellular protein components | • The occasionally pericellular hallo (autolitic modification) impose a differential diagnosis with the oligodendrocytes [23] | ||
| Mallory’s (phosphotungstic acid – hematoxylin) stain [24] | Special stain | Astrocyte processes (deep blue) | |
| Orange-acridine stain [24] | Special stain | Cellular body | • Reveals the astrocytic hyperplasia, without the modification of the cytoplasm aspects [24] |
| Metallic impregnations [23] | Nuclei | • Reveals the cellular characteristic star- shaped aspect | |
| • Del Rio Hortega method | • Special technique with ammonia silver carbonate | Cytoplasm processes | • The abundant cytoplasm surrounding the nuclei differentiates the astrocytes from oligodendrocyte |
| • Ramon y Cajal method (see Figures 1 and 2) | • Special technique with gold chloride | • The fibrillar aspect of the cytoplasm is due to the material formed by the aggregation of GFAP intermediate filaments | |
| • Golgi stain | • Special technique with silver nitrate | • The vascular endfeet are easy to identify. | |
| • Protoplasmic astrocytes, due to their proximity to the blood vessels, are able to contact the vessel directly by their cell body | |||
| • The perivascular hallo is considered to be an artefact [23]. | |||
| Electron microscopy [24] | Cytoplasm intermediate GFAP | • Cytoplasm pale , with lack of organelles | |
| • The clear, perivascular spaces indicate excessive dilatation of astrocytic processes due to water imbibitions | |||
| • The ultrastructural resemblance between normal and well differentiated neoplastic astrocytes is one of the arguments against the use of this method for positive diagnosis of low grade glioma [24] | |||
| Immunohistochemistry | GFAP represents an integrator of the cellular space, but it is also implicated in complex cellular events, such as cytoskeleton reorganisation, myelination, cellular adhesion and several signalling pathways [23, 24]. | ||
| • GFAP (intracytoplasmic protein, with 50 Kda molecular weight, considered the major component of glial fibrils and a marker of astrocytic differentiation) [23, 24] (see Figure 3) | • Golden standard for the definition of astrocytes | Cell body | • Fibrillary astrocytes contain a massive amount of GFAP in their cell bodies and processes unlike protoplasmic astrocyte. |
| • There are different clones of antiGFAP antibodie, characteristic to the different research | Cell processes (positive immunostaining reaction: brown spots) | • Protoplasmic astrocytes are much larger than their GFAP-defined profiles due to the presence of numerous fine processes that are GFAP-negative | |
| • Laboratories (e.g. GF2 DAKO clone; Astro 1) [23, 24] | • In astrocytomas, along with the enhancement of malignity, the intracellular quantity of GFAP is progressively reduced; therefore the evaluation of GFAP immunohistochemical staining will enable the immunophenotypic characterisation of the investigated glial tumors and the confirmation of histopathological diagnosis | ||
| • Not all the cells in the CNS that express GFAP are astrocytes (e.g: astrocyte-like cells from the SVZ-derived from radial glia, ependymal cells) [1, 25, 26] | |||
| • GFAP has also been located in rat kidney glomeruli and peritubular fibroblasts [1, 27], Leydig cells of the testis [1, 28], skin keratinocytes [1, 29], osteocytes of bones, chondrocytes of epiglottis, bronchus [1, 30], and stellate-shaped cells of the pancreas and liver [1] | |||
| S100B (belongs to the S100 family of EF-band calcium binding proteins [1, 31]). | There are different clones of anti S100 antibodies, characteristic to the different research laboratories (e.g. MAB079, CBL410.) | Cell membrane | • Expressed by a subtype of mature astrocytes that ensheath blood vessels and by NG2-expressing astrocytes [1, 31] |
| Other astrocytic markers | |||
| • GLT-1 (the glutamate transporters GLAST) [6] | • GLT-1 is expressed by all astrocytes and provide punctuate staining [6] | ||
| • Human EAAT2 (excitatory amino acids, 1 and 2 for human brain) [6] | |||
| • Glycogen granules[6] | Cytoplasm | ||
| • Gglutamine synthase (GS)[1, 32–35] | GS- enzyme that catalyzes the conversion of ammonia and glutamate to glutamine | Cytoplasm | GS is expressed also by oligodendrocytes [1, 32–35] |
| Kir4.1 (inwardly rectifying K+ channels) [1, 36, 37] | Kir4.1 are only expressed by a subset of astrocytes [37] | ||
| • Aquaporin 4 channels[1, 38] | Cell processes | • Aquaporin 4 channels is localized in some parts of the astrocytic processes rendering identification of the whole cell difficult to interpret [38] | |
| • AldhL1 (aldehyde dehydrogenase 1 family, member L1) [1, 39]. | Genome | • All astrocytes | |
| Battery of tests[40]• GFAP-driven GFP (green fluorescent protein) expressionGFAPprotein expression, S100ß immunostaining | Combinatorial approach | • Nine different classes of astrocytes has been identified, that included Bergmann glia, ependymal glia, fibrous astrocytes, marginal glia, perivascular glia, protoplasmic astrocytes, radial glia, tanycytes and velate glia [3, 40] | |
| • GFAP expression glutamate response [41] | • Define the phenotype of an astrocyte population as (GFAP+/NG2-; T+/R-) which is distinct from NG2-glia (GFAP-/NG2+ T-/R+) [41] | ||
| Dye-filling techniques [6, 42](e.g. sharp electrode, patch clamp recordings, single cell electroporation) | Special techniques that identify cells recorded in situ after filling them with a dye present in a micro-electrode | Cell body | • This technique has the advantage that the cells to be studied can be preselected in living tissue [6, 42] |
| It is suplemented by use of presumed astrocyte- | Cell processes | • However, proteins and promoter activation are subjects to change. Hence one can have a GFAP(-) cell that one should call an astrocyte because it has these other properties [6, 42] | |
| Specific promoters to drive synthesis of fluorescent proteins | • Using these procedures the domain organisation of astrocytes has been demonstrated along with the fusiform morphology of astrocyte nucleus, both playing a possible role in pathology [3, 43, 44] | ||
| Transgenic techniques (use transgenic mice) [1] | Visualize fluorescent astrocytes | Cell body | • Mice specific for astrocytes express [1] |
| Cell processes | - GFP | ||
| - Enhanced GFP under the human GFAP promoter (hGFAP-GFP mice) | |||
| - GLT-1-GFP | |||
| - BLBP-dsRed2 |
Figure 1.
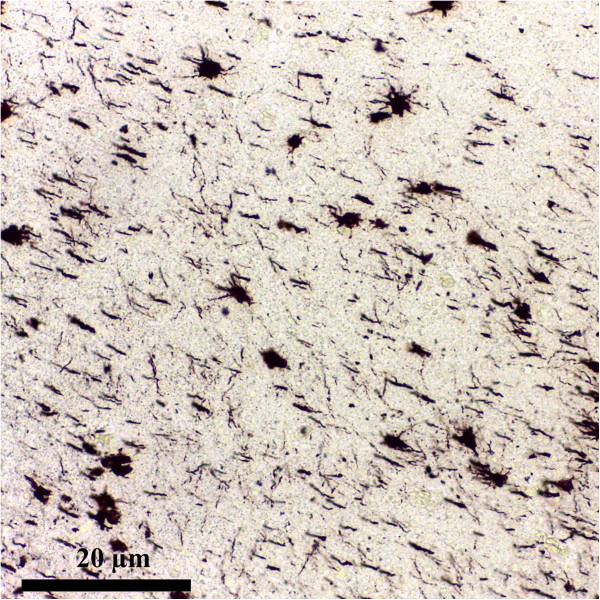
Astrocytes overview. Metalic impregnation Ramon Y Cajal Ob. 20x. Human brain (personal collection).
Figure 2.
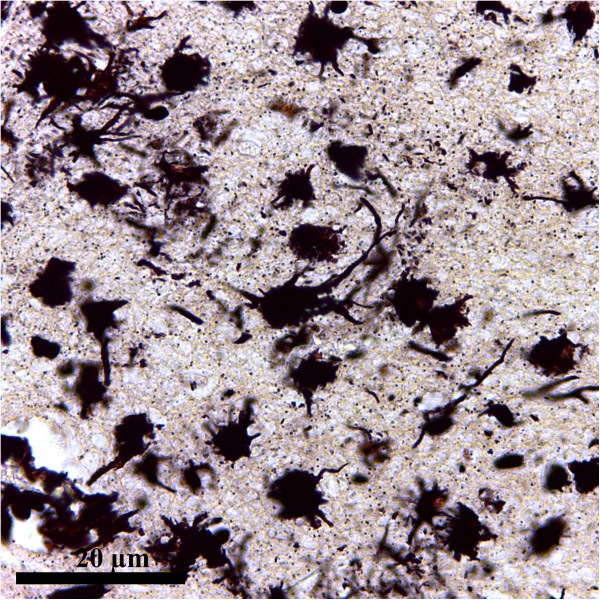
Astrocytes overview. Metalic impregnation Ramon Y Cajal Ob. 40x. Human brain (personal collection).
Figure 3.
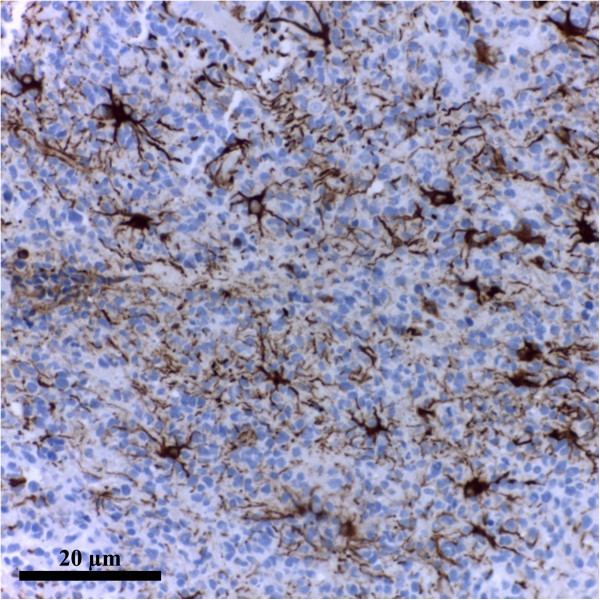
Astrocytes overview. GFAP Clone GF2 DAKO. Human brain. Ob. 20x. (personal collection).
For example, the grey matter protoplasmic astrocytes, are generated from embryonic radial glia and, to a lesser extent, from intermediate progenitors migrating from the neonatal subventricular zones. These two pathways of development will generate astrocytes with different patterns of gene expression and possibly different functions.
On the other hand, the white matter fibrous astrocytes are predominantly generated from neonatal subventricular zone progenitors [1].
Yet, it is important to recognize that subsets of progenitors will spontaneously differentiate in culture, as the intrinsic program of the cells modulates the process of cell division and differentiation together with culture conditions. Nevertheless, treatment of GRP cultures with fetal bovine serum (FBS) resulted in the production of A2B5-/GFAP + astrocytes with a fibroblastlike flat morphology, whereas exposure to basic fibroblast growth factor (bFGF) together with ciliary neurotrophic factor (CNTF) produced mostly process-bearing A2B5+/GFAP + astrocytes. Further research is needed to elucidate the identity of the different classes of intermediate progenitors or to obtain a clear antigenic signature of the lineage [8].
The development of astrocytes from a multipotent stem cell that prior to this has produced neuronal precursor cells, implies a specific differentiation via a multi-step process. The switch toward the glial differentiation is regulated by a change in receptor composition on the cell surface and responsiveness to fibroblast growth factor (FGF) and epidermal growth factor (EGF); futhermore, signaling molecules like CNTF, bone morphogenetic proteins (BMF), and EGF will continue to drive the glial precursor cell into the astroglial direction. However, the early astrocytes will interact with their microenvironment not only by releasing and responding to diverse soluble factors, but also expressing a wide range of extracellular matrix (ECM) molecules, as proteoglycans (lectican family) and tenascins. Lately it is considered that these ECM molecules have the ability to participate in glial development (e.g. the matrix protein Tenascin C (Tnc), proved to be an important regulator of astrocyte precursor cell proliferation, maturation and migration during spinal cord development) and those expressed by reactive astrocytes under pathophysiological conditions, are known to act mostly in an inhibitory fashion [9, 10].
Astrocytes as a source of stem cells
The most recent and exciting finding in the astrocyte field, which challenges the traditional definition of astrocyte itself, is the discovery that there is a subclass of mature astrocytes which represent the stem cells in the adult neurogenic zones. The GFAP-expressing stem cells have characteristics of embryonic radial glia and mature astrocytes, but display subtle differences and retain properties of neural progenitors. These stem cells act in concert with resident astrocytes to contribute to cell genesis and maintaining the neurogenic environment, the niche. Perhaps these cells are retained in a transitional stage between radial glia and astrocytes, due to the persistence of embryonic extracellular matrix molecules. This permissive environment in the neurogenic niche allows the retention of intrinsic genetic programs to maintain “stemness” [1, 17]. It was shown that, the proneural genes neurogenin-2 and Mash1 possess the ability to reprogram these astrocytes to stem cells that can generate neurons [1].
In the adult subventricular zone (SVZ) and subgranular zone (SGZ), two distinct population of neural progenitors (multipotent neural stem cells) express GFAP [1, 18–20] and give rise to neuroblasts that either migrate to the olfactory bulb (to become olfactory interneurons) [1, 19, 21, 22] or generate newborn granule neurons. GFAP-expressing cells of the SVZ have been termed SVZ astrocytes, astrocyte-like cells or B cells. The histology of these cells comprises irregular contours that filled the spaces between neighbouring cells, irregular nuclei with invaginations, and light cytoplasm with few free ribosomes. They also expressed abundant intermediate filaments. Differences were found between the two types of astrocyte-like cells. Type B1 cells are larger than type B2 cells and possess euchromatic nuclei; they are adjacent to ependymal cells. Type B2 cells are smaller with hyperchromatic nuclei and are mostly located at the interface with the striatal parenchyma [1].
Another type of stem cell which expresses GFAP can be found in the adult SVZ but it is questionable whether these adult stem cells really belong to the astrocyte family. They has different molecular features, because they express nestin (an intermediate filament), that characterise only embryonic astrocytes, reactive astrocytes or neuroblastes and intermediate progenitors [1].
In conclusion, there is much need for further studies to be conducted in an attempt of finding new antigenic markers, physiological properties or molecular profiles for a better definition of these varieties of stem cells and to answer to challenging question as the ability of every astrocyte to revert to stem cells given the right environment [1].
Astrocytic markers and stains
Many novel tools to study astrocytes were given by the technological advances over the past decades. From the early Golgi stains to immunostaining for glial fibrils, or the recent dye-filling techniques (e.g. sharp electrode, patch clamp recordings, single cell electroporation), and transgenic approaches to visualize fluorescent astrocytes, our understanding of astrocyte characteristics has dramatically evolved [1] (see Table 2), Figures 1, 2 and 3.
The morphological features and the close relationships with both neurons and capillaries are the most constant characteristics that can be used to define the astrocytic phenotype [3] (see Figure 4).
Figure 4.
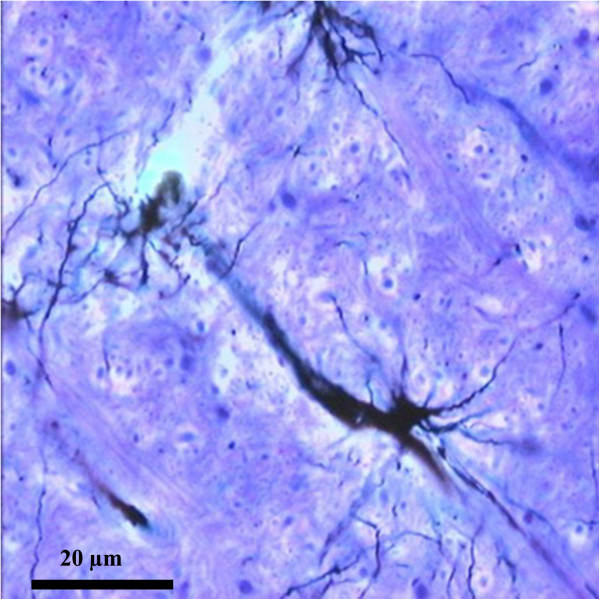
Protoplasmic astrocyte proximal to a blood vessel. Metallic impregnation Ramon Y Cajal. Ob. 20x. Human brain (personal collection).
Types and morphology
Two major classes of astrocytes were first described in the 19th century by using the Golgi staining, which revealed their distinct morphological pattern: the protoplasmic and fibrous astrocytes. Nowadays the classification of astrocytes into fibrous and protoplasmic is considered to be outdated [45]; their morphological diversity can be illustrated by specialised classes of astrocytes represented by: the cerebellar Bergmann and Fananas glia, the Müller glia of the retina, the pituicytes of the neurohypophysis and the interstitial cells of the epiphysis. Additionally, in humans and primates two novel subtypes of astrocytes have been described: interlaminar astrocytes and varicose projection astrocytes [3, 4, 46–49]. (see Table 3) Figures 5 and 6.
Table 3.
Types of astrocytes
| Types of astrocytes | Location | Morphology | Functions | Particularities |
|---|---|---|---|---|
| Protoplasmic astrocytes | Uniformly distributed within the grey matter [3] | Bushy appearance, with numerous short, branched, thick processes [50]. The cell body is ovoid or fusiform (see Figure 5) | • Form the blood–brain barrier | Their processes exhibit endfeet enveloping the synapses and the blood vessels [51]. The processes express |
| • Regulate the blood flow | ||||
| • Neuronal metabolism | • Receptors for neurotransmitters, cytokines, growth factors | |||
| • Implicated in the synapse function | • Transporters | |||
| • Fluid, ion, pH and transmitter homeostasis [45] | • Ion channels [7]. In rodents, there is minimal overlapping between the processes of the neighbouring astrocytes [43, 44, 52–54]. In humans, the superposition of the domains occupied by the astrocytes processes is augmented [3] | |||
| Fibrous astrocytes | Within the white matter, oriented longitudinally, along the nervous fibers bundles [1] | Star-shaped cells. Posses long, thin and straight processes [45] (see Figure 6) | Their endfeet processes envelop the nodes of Ranvier and the blood vessels [45] | |
| Interlaminar astrocytes | In the molecular 1st layer of the cerebral cortex, next to the pial surface | Spherical cell bodies and processes | Unknown Support the calcium wave propagation in humans [3] | Are found only in humans and primates. Their processes are included in the pial glial membrane, creating a thick network of GFAP fibers [46–49] |
| Varicose projection astrocytes | In the 5th and the 6th layers of the cerebral cortex | Exhibit 1 to 5 long processes (up to 1 mm in length), characterized by evenly (10 μm) spaced varicosities [3, 46] | Unknown | Were identified only in humans and chimpanzees. They are GFAP+ cells [3, 46] |
| Bergmann glia (epithelial glial cells) | In the Purkinje-cell and the granular layers of the cerebellar cortex | Posses long processes extending towards the molecular layer of the cerebellar cortex, in a fan-like arrangement, exhibiting pial vascular endfeet [23] | Implicated in synapse function: capable to interfere with synaptic transmission by communicating with neurons via the extracellular space, by modulating ion concentrations or transmitter levels in the synaptic cleft [23] | Display receptors with distinct biophysical and pharmacological features allowing them to sense the activity of synapses [23] |
| Fananas cells | In the molecular layer of the cerebellar cortex | Posses several short side processes with a characteristic feather-like arrangement [23] | ||
| Müller cells | In the 6th layer of the visual retina | Supportive cells: they form the inner and the outer limiting membranes | The limiting membranes consist of junctional complexes between the cellular processes of the Müller cells | |
| The outer membrane separates the external segment of the photoreceptor cells from the cell bodies and the outer membrane separates the retina from the vitrous body [23] | They have an intense metabolic activity and contain microfilaments and glycogen within their cytoplasm [23] | |||
| Pituicytes | In the neurohypophysis | Irregular in shape with many cytoplasmic processes extending in the proximity of the capillaries and surrounding the Herring bodies [24] | Their cytoplasm contains lipid droplets and pigment granules. | |
| They are immunoreactive for GFAP, vimentin and S100 protein [24] | ||||
| Inerstitial epiphysial cells | In the epiphysis | Exhibit cytoplasmic processes | Contain numerous filaments within their processes [23] |
Figure 5.

Protoplasmic astrocyte. Metallic impregnation Ramon Y Cajal Ob. 100 immersion. Human brain (personal collection).
Figure 6.
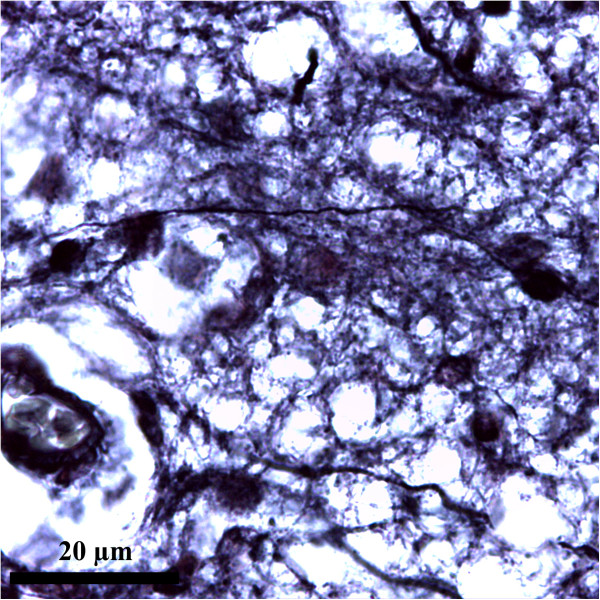
Fibrous astrocyte. Metallic impregnation Ramon Y Cajal Ob. 100 immersion. Human brain (personal collection).
The above presented heterogeneity of astrocytes could arise from separate lineages, plasticity of mature cells (motility and reactivity after injuries), or association of both factors [3, 54]. Methods of molecular biology, like time-lapse studies in slice culture, demonstrated the participation of astrocytes in synaptic remodelling, since the astrocytic processes are motile and enwrap active synapses [3, 55, 56].
It is well-known that mature astrocytes can exhibit forms of plasticity: motility and reactivity after injuries. Time lapse studies of astrocytes in acute slice and slice culture have shown that astrocyte processes act much like dendritic spines; they are frequently motile and contact active synapses [3, 55, 57], the role of this feature implying the synaptic remodelling.
Reactive astrocytes
Astrocytes become reactive notably after injuries, when the intermediate filament proteins (e.g. GFAP, vimentin, nestin) are upregulated, becoming larger and there is an alteration of the domain organization [58, 59].
The reactive morphological variants comprise two main categories: the individualised and the global reactive astrocytes. Individualized reactive astrocytes encompass several types: pilocytic astrocyte, gemistocytic astrocyte, type I and II Alzheimer astrocytes. The global reactive astrocytes are the characteristic feature of reactive astrogliosis (see Table 4) [60].
Table 4.
Individualized reactive astrocytes variants
| Individualized reactive astrocytes variants | Causes | Morphology | Particularities |
|---|---|---|---|
| Pilocytic astrocytes [23, 24] | • In mild and moderate injuries as individual form of reactive astrocytes | • Elongated, bipolar cell body | These cells contain the Rosenthal fibers (specific but inconstant eosinophilic, cork-screw shaped elements), representing an advanced stage of cellular degeneration in astrocytoma |
| • Astrocytoma | • Fusiform nuclei | ||
| • Thin and long hair-like GFAP+ processes | |||
| Gemistocytic astrocytes [23, 24] | • In mild and moderate injuries as individual form of reactive astrocytes | • Large, dilatated, oval cell body | The organelles are numerous and located in the central zone of the cell body. The glial filaments are also numerous and peripherally arranged, beneath the plasmalemma |
| • In gemistocytic astrocytoma as a characteristic feature of this tumors [23] | • Few thick cytoplasmic processes | ||
| • Abundant, deeply eosinophilic cytoplasm | |||
| • Polymorphic nuclei, frequently eccentrical. | |||
| Alzheimer type I astrocytes [23, 24] | • Progressive multifocal leuco-encephalopathy | • Enlarged cell body | |
| • Numerous nuclei | |||
| Alzheimer type II astrocytes [23, 24] | • Associated with high blood ammonia in hepatic encephalopathy | • Enlarged cell body | Ammonia taken up by astrocytes is converted to osmotically active glutamine, resulting in astrocytic swelling |
| • In Wilson disease | • Vesicular nuclei with one or more nucleoli |
Reactive astrogliosis, a hallmark of all forms of CNS injuries, is the result of a multi-step process involving gradates changes in astrocytes.
Histopathological examinations of human brain in various neurological conditions have provided different degrees of reactive astrogliosis. According to Sofroniew et al., the following categories of reactive astrogliosis can be identified: mild to moderate astrogliosis, severe astrogliosis and the glial scar [60].
Mild to moderate astrogliosis is a manifestation of various disorders (systemic viral and bacterial infections, non-penetrating trauma) and also found in the distant areas surrounding the focal cerebral lesions [60]. The changes associated with mild to moderate astrogliosis are reversible if the triggering mechanism has resolved. In this type of injuries, subtle alterations occur in the expression of molecules implicated in the cellular activity: cell structure, energy metabolism, intracellular signaling, membrane transporters and pumps [60].
Various functional categories of genes and molecules modulated by reactive astrocytes can be either upregulated or downregulated, depending on the trigger or the moment after the insult. As a result of these alterations, specific histopathological features can be identified in astrocytes (see Table 5).
Table 5.
Reactive astrogliosis
| Reactive astrogliois | Changes in astrocytes morphology | Changes in molecules expression | |
|---|---|---|---|
| Upregulated molecules | Upregulated or downregulated molecules | ||
| Mild to moderate astrogliosis | • Hypertrophy of cell body | • Structural elements: GFAP, nestin, vimentin | • Inflammatory cell regulators: cytokines, growth factors, glutathione |
| • Astrocytes processes are are numerous and thicker | • Transcriptional regulators: STAT3, NFκB, Rheb-m TOR, cAMP, Olig2, SOX9 [61–65]. | • Transporters and pumps: AQP4 and Na+/K+ transporters [61, 66–69] | |
| • Glutamate transporter [70–73] | |||
| • The non-overlapping domains of individual astrocytes are preserved | • Vascular regulators: PGE, NO [74, 75] | ||
| • Energy provision: lactate [76] | |||
| • Molecules implicated in synapse formation and | |||
| Severe astrogliosis and glial scar | • Intense hypertrophy of cell body | • Remodeling: thrombospondin and Complement C1q [77, 78] | |
| • Significant extension of processes | • Molecules implicated in oxidative stress and providing protection from oxidative stress: NO, NOS, SOD, Glutathione [67, 68, 79] | ||
| • Proliferation | |||
| • Overlapping of individual domains | |||
| • Substantial reorganization of tissue architecture [60] | |||
Severe diffuse reactive astrogliosis is characterised by permanent, pronounced and long-lasting changes and it is found in the areas surrounding severe focal lesions or infections, as well as in neurodegeneration [60].
Compact scar formation occurs in most severe injuries, such as overt tissue damage (e.g. penetrating or contusive trauma), inflammation initiated by invasive infections or abscesses, neoplasm and chronic neurodegeneration. Reactive astrogliosis reaches its highest level of activation: astrocytes undergo intense proliferation, and their long, branched processes overlap (see Table 5) [60].
Any cell type in the CNS (e.g. neurons, all types of glial cells, endothelial cells and leucocytes) is potentially able to release the molecular mediators of astrogliosis [60]. Signaling pathways and molecules implicated in mediating specific aspects of reactive astrogliosis include: STAT3 (signal transducer and activator of transcription 3), NFκB (nuclear factor kappa B), cAMP (cyclic adenosine monophosphate), all these inducing upregulation of structural molecules (e.g. GFAP, vimentin, nestin) [62, 63, 80]; moreover, STAT3 induces astrocyte hypertrophy, scar formation and exerts anti-inflammatory effect [62]; NFκB exerts pro-inflammatory effect [63, 81]; Olig2, Endothelin-1 induce astrocyte proliferation [65, 82].
The newly formed cells in the glial scar derive from different sources: mature astrocytes that re-enter the cell cycle, NG2 progenitors and ependymal cells progenitors.
Molecular mediators and triggers leading to proliferation include: cytokines and growth factors (e.g. IL (interleukin) 6, LIF (leukemia inhibitory factor), CNTF, IL1, IL10, TGFβ (transforming growth factor), TNFα (tumor necrosis factor), INFγ (interferon), [63, 69]), Toll-like receptor ligants [83], LPS (lipopolysaccharide), molecules of oxidative stress (e.g. NO (nitric oxide) and ROS (reactive oxygen species) [67]), modulators and neurotransmitters (e.g. noradrenalin and glutamate [84]), ischemia associated hypoxia and glucose deprivation [67], neurodegeneration associated amyloid- beta [85] and ATP (adenosine triphosphate) released by cell injury [86].
Reactive astrocytes interact with other cell types: fibromeningeal cells and NG-2 positive glia and are associated with a dense collagenous extracellular matrix to form complex glial scars [60, 62, 87]. The changes leading to scar formation persist even after the triggering factor has been removed [61, 62, 88, 89].
Recent evidence suggest that the glial scars might have a beneficial role, as they form narrow, dense and compact barriers; these barriers delimit the periphery of severe tissue damage, isolate the lesion, thus preventing inflammatory cells and infectious agents from spreading into the healthy parenchyma [90].
Conclusions
This part of the review is an insight into the morphology and biology of astrocytes, with an emphasis on the latest findings concerning the novel cell subtypes, the developmental lineages and their functions.
From the very first description of astrocytes in the 19th century, these cells concept has been at a standstill until recently when it remarkably progressed.
The variety of glial precursors, their ability to display stem cells features and different adult astrocytes morphology certify that the astrocytic phenotype is influenced by both the local cytoarchitecture and the functional requirements in specific brain areas.
Abbreviations
- AldhL1
Aldehyde dehydrogenase 1 family, member l1
- AQP4
Aquaporin 4
- ATP
Adenosine triphosphate
- BBB
Blood–brain barrier
- bFGF
Basic fibroblast growth factor
- BLBP
Brain lipid basic protein
- BMF
Bone Morphogenetic Proteins
- cAMP
Cyclic adenosine monophosphate
- CNS
Central nervous system
- CNTF
Ciliary neurotrophic factor
- Dlx2
Distal-less homeobox 2
- ECM
Extracellular matrix
- EGF
Epidermal growth factor
- EAAT
Excitatory amino-acid transporters
- FBS
Fetal bovine serum
- FGF
Fibroblast growth factor
- GLAST
Glutamate amino acid transporters
- GLT
Glutamate transporters
- GFAP
Glial fibrillary acidic protein
- GFP
Green fluorescent protein
- GRP
Glial-restricted precursors
- HBB
Hemolymph-brain barrier
- IL
Interleukin
- INF
Interferon
- Kir 4.1
Inwardly rectifying K + channels
- LIF
Leukemia inhibitory factor
- LPS
Lipopolysaccharide
- NFκB
Nuclear factor kappa B
- NO
Nitric oxide
- NOS
Nitric oxide synthase
- OLIG2
Oligodendrocyte transcription factor
- PGE
Prostaglandin E
- Rheb-m TOR
Ras homolog enriched in brain – mamalian target of rapamycin
- ROS
Reactive oxygen species
- SOD
Superoxide dismutase
- SOX9
Transcription factor SOX9
- STAT3
Signal transducer and activator of transcription 3
- SGZ
Subgranular zone
- SVZ
Subventricular zone
- TGF
Transforming growth factor
- Tnc
Tenascin C
- TNF
Tumor necrosis factor
- VZ
Ventricular zone.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
The images are part of the personal collection of the first author. Both authors contribute to the synthesis of the data from the actual scientific literature. All authors read and approved the final manuscript.
Contributor Information
Alina Simona Şovrea, Email: a_sovrea@yahoo.com.
Adina Bianca Boşca, Email: biancabosca@yahoo.com.
References
- 1.Wang DD, Bordey A. The astrocyte odyssey. Prog Neurobiol. 2008;86(4):342–367. doi: 10.1016/j.pneurobio.2008.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Parpura V, Verkhratsky A. Neuroglia at the crossroads of homoeostasis, metabolism and signalling: evolution of the concept. ASN Neuro. 2012;4(4):201–205. doi: 10.1042/AN20120019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Oberheim NA, Goldman SA, Nedergaard M. Heterogeneity of astrocytic form and function. Methods Mol Biol. 2012;814:23–45. doi: 10.1007/978-1-61779-452-0_3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Andriezen WL. The neuroglia elements in the human brain. Br Med J. 1893;2(1700):227–230. doi: 10.1136/bmj.2.1700.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Parpura V, Verkhratsky A. Astrocytes revisited: concise historic outlook on glutamate homeostasis and signaling. Croat Med J. 2012;53(6):518–528. doi: 10.3325/cmj.2012.53.518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kimelberg HK, Nedergaard M. Functions of astrocytes and their potential as therapeutic targets. Neurotherapeutics. 2010;7(4):338–353. doi: 10.1016/j.nurt.2010.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bélanger M, Magistretti PJ. The role of astroglia in neuroprotection. Dialogues Clin Neurosci. 2009;11(3):281–295. doi: 10.31887/DCNS.2009.11.3/mbelanger. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Haas C, Neuhuber B, Yamagami T, Rao M, Fischer I. Phenotypic analysis of astrocytes derived from glial restricted precursors and their impact on axon regeneration. Exp Neurol. 2012;233(2):717–732. doi: 10.1016/j.expneurol.2011.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kang W, Hébert JM. Signaling pathways in reactive astrocytes, a genetic perspective. Mol Neurobiol. 2011;43(3):147–154. doi: 10.1007/s12035-011-8163-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wiese S, Karus M, Faissner A. Astrocytes as a source for extracellular matrix molecules and cytokines. Front Pharmacol. 2012;3:120. doi: 10.3389/fphar.2012.00120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Malatesta P, Hartfuss E, Götz M. Isolation of radial glial cells by fluorescent-activated cell sorting reveals a neuronal lineage. Development. 2000;127(24):5253–5263. doi: 10.1242/dev.127.24.5253. [DOI] [PubMed] [Google Scholar]
- 12.Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 2001;409(6821):714–720. doi: 10.1038/35055553. [DOI] [PubMed] [Google Scholar]
- 13.Liu Y, Wu Y, Lee JC, Xue H, Pevny LH, Kaprielian Z, Rao MS. Oligodendrocyte and astrocyte development in rodents: an in situ and immunohistological analysis during embryonic development. Glia. 2002;40(1):25–43. doi: 10.1002/glia.10111. [DOI] [PubMed] [Google Scholar]
- 14.Cai J, Chen Y, Cai WH, Hurlock EC, Wu H, Kernie SG, Parada LF, Lu QR. A crucial role for Olig2 in white matter astrocyte development. Development. 2007;134(10):1887–1999. doi: 10.1242/dev.02847. [DOI] [PubMed] [Google Scholar]
- 15.Liu Y, Rao MS. Glial progenitors in the CNS and possible lineage relationships among them. Biol Cell. 2004;96(4):279–290. doi: 10.1111/j.1768-322X.2004.tb01416.x. [DOI] [PubMed] [Google Scholar]
- 16.Schmechel DE, Rakic PA. Golgi study of radial glial cells in developing monkey telencephalon: morphogenesis and transformation into astrocytes. Anat Embryol (Berl) 1979;156(2):115–152. doi: 10.1007/BF00300010. [DOI] [PubMed] [Google Scholar]
- 17.Gates MA, Thomas LB, Howard EM, Laywell ED, Sajin B, Faissner A, Götz B, Silver J, Steindler DA. Cell and molecular analysis of the developing and adult mouse subventricular zone of the cerebral hemispheres. J Comp Neurol. 1995;361(2):249–266. doi: 10.1002/cne.903610205. [DOI] [PubMed] [Google Scholar]
- 18.Cameron HA, Woolley CS, McEwen BS, Gould E. Differentiation of newly born neurons and glia in the dentate gyrus of the adult rat. Neuroscience. 1993;56(2):337–344. doi: 10.1016/0306-4522(93)90335-D. [DOI] [PubMed] [Google Scholar]
- 19.Doetsch F, Caillé I, Lim DA, García-Verdugo JM, Alvarez-Buylla A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell. 1999;97(6):703–716. doi: 10.1016/S0092-8674(00)80783-7. [DOI] [PubMed] [Google Scholar]
- 20.Seri B, García-Verdugo JM, McEwen BS, Alvarez-Buylla A. Astrocytes give rise to new neurons in the adult mammalian hippocampus. J Neurosci. 2001;21(18):7153–7160. doi: 10.1523/JNEUROSCI.21-18-07153.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Altman J. Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J Comp Neurol. 1969;137(4):433–457. doi: 10.1002/cne.901370404. [DOI] [PubMed] [Google Scholar]
- 22.Luskin MB. Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron. 1993;11(1):173–189. doi: 10.1016/0896-6273(93)90281-U. [DOI] [PubMed] [Google Scholar]
- 23.Fuller GN, Burger PC. In: Central nervous system in Histology for pathologists. Sternberg SS, editor. New York: Raven Press; 1992. pp. 145–167. [Google Scholar]
- 24.Burger PC, Scheithauer BW. Atlas of Tumor Pathology. 3 1993. Tumors of the Central Nervous System-Armed Forces Institute of Pathology. [Google Scholar]
- 25.Liu X, Bolteus AJ, Balkin DM, Henschel O, Bordey A. GFAP-expressing cells in the postnatal subventricular zone display a unique glial phenotype intermediate between radial glia and astrocytes. Glia. 2006;54(5):394–410. doi: 10.1002/glia.20392. [DOI] [PubMed] [Google Scholar]
- 26.Spassky N, Merkle FT, Flames N, Tramontin AD, García-Verdugo JM, Alvarez-Buylla A. Adult ependymal cells are postmitotic and are derived from radial glial cells during embryogenesis. J Neurosci. 2005;25(1):10–18. doi: 10.1523/JNEUROSCI.1108-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Buniatian GH, Hartmann HJ, Traub P, Wiesinger H, Albinus M, Nagel W, Shoeman R, Mecke D, Weser U. Glial fibrillary acidic protein-positive cells of the kidney are capable of raising a protective biochemical barrier similar to astrocytes: expression of metallothionein in podocytes. Anat Rec. 2002;267(4):296–306. doi: 10.1002/ar.10115. [DOI] [PubMed] [Google Scholar]
- 28.Davidoff MS, Middendorff R, Köfüncü E, Müller D, Jezek D, Holstein AF. Leydig cells of the human testis possess astrocyte and oligodendrocyte marker molecules. Acta Histochem. 2002;104(1):39–49. doi: 10.1078/0065-1281-00630. [DOI] [PubMed] [Google Scholar]
- 29.Danielyan L, Tolstonog G, Traub P, Salvetter J, Gleiter CH, Reisig D, Gebhardt R, Buniatian GH. Colocalization of glial fibrillary acidic protein, metallothionein, and MHC II in human, rat, NOD/SCID, and nude mouse skin keratinocytes and fibroblasts. J Invest Dermatol. 2007;127(3):555–563. doi: 10.1038/sj.jid.5700575. [DOI] [PubMed] [Google Scholar]
- 30.Kasantikul V, Shuangshoti S. Positivity to glial fibrillary acidic protein in bone, cartilage, and chordoma. J Surg Oncol. 1989;41(1):22–26. doi: 10.1002/jso.2930410109. [DOI] [PubMed] [Google Scholar]
- 31.Baudier J, Glasser N, Gerard D. Ions binding to S100 proteins. I. Calcium- and zinc-binding properties of bovine brain S100 alpha alpha, S100a (alpha beta), and S100b (beta beta) protein: Zn2+ regulates Ca2+ binding on S100b protein. J Biol Chem. 1986;261(18):8192–8203. [PubMed] [Google Scholar]
- 32.Schousboe A. Role of astrocytes in the maintenance and modulation of glutamatergic and GABAergic neurotransmission. Neurochem Res. 2003;28(2):347–352. doi: 10.1023/A:1022397704922. [DOI] [PubMed] [Google Scholar]
- 33.Martinez-Hernandez A, Bell KP, Norenberg MD. Glutamine synthetase: glial localization in brain. Science. 1977;195(4284):1356–1358. doi: 10.1126/science.14400. [DOI] [PubMed] [Google Scholar]
- 34.Cammer W. Glutamine synthetase in the central nervous system is not confined to astrocytes. J Neuroimmunol. 1990;26(2):173–178. doi: 10.1016/0165-5728(90)90088-5. [DOI] [PubMed] [Google Scholar]
- 35.D’Amelio F, Eng LF, Gibbs MA. Glutamine synthetase immunoreactivity is present in oligodendroglia of various regions of the central nervous system. Glia. 1990;3(5):335–341. doi: 10.1002/glia.440030504. [DOI] [PubMed] [Google Scholar]
- 36.Takumi T, Ishii T, Horio Y, Morishige K, Takahashi N, Yamada M, Yamashita T, Kiyama H, Sohmiya K, Nakanishi S. A novel ATP-dependent inward rectifier potassium channel expressed predominantly in glial cells. J Biol Chem. 1995;270(27):16339–16346. doi: 10.1074/jbc.270.27.16339. [DOI] [PubMed] [Google Scholar]
- 37.Higashi K, Fujita A, Inanobe A, Tanemoto M, Doi K, Kubo T, Kurachi Y. An inwardly rectifying K(+) channel, Kir4.1, Expressed in astrocytes surrounds synapses and blood vessels in brain. Am J Physiol Cell Physiol. 2001;281(3):C922–C931. doi: 10.1152/ajpcell.2001.281.3.C922. [DOI] [PubMed] [Google Scholar]
- 38.Nielsen S, Nagelhus EA, Amiry-Moghaddam M, Bourque C, Agre P, Ottersen OP. Specialized membrane domains for water transport in glial cells: high-resolution immunogold cytochemistry of aquaporin-4 in rat brain. J Neurosci. 1997;17(1):171–180. doi: 10.1523/JNEUROSCI.17-01-00171.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, Xing Y, Lubischer JL, Krieg PA, Krupenko SA, Thompson WJ, Barres BA. A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function. J Neurosci. 2008;28(1):264–278. doi: 10.1523/JNEUROSCI.4178-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Emsley JG, Macklis JD. Astroglial heterogeneity closely reflects the neuronal-defined anatomy of the adult murine CNS. Neuron Glia Biol. 2006;2(3):175–186. doi: 10.1017/S1740925X06000202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nishiyama A, Yang Z, Butt A. Astrocytes and NG2-glia: what’s in a name? J Anat. 2005;207(6):687–693. doi: 10.1111/j.1469-7580.2005.00489.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fiacco T, Casper K, Sweger E, Agulhon C, Taves S, Kurtzer-Minton S. Molecular approaches for studying astrocytes. In: Parpura V, Haydon PG, editors. Astrocytes in (patho)physiology of the nervous system. New York: Springer; 2009. pp. 383–405. [Google Scholar]
- 43.Bushong EA, Martone ME, Jones YZ, Ellisman MH. Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. J Neurosci. 2002;22:183–192. doi: 10.1523/JNEUROSCI.22-01-00183.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ogata K, Kosaka T. Structural and quantitative analysis of astrocytes in the mouse hippocampus. Neuroscience. 2002;113:221–233. doi: 10.1016/S0306-4522(02)00041-6. [DOI] [PubMed] [Google Scholar]
- 45.Sofroniew MV, Vinters HV. Astrocytes: biology and pathology. Acta Neuropathol. 2010;119:7–35. doi: 10.1007/s00401-009-0619-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Oberheim NA, Takano T, Han X, He W, Lin JH, Wang F, Xu Q, Wyatt JD, Pilcher W, Ojemann JG, Ransom BR, Goldman SA, Nedergaard M. Uniquely hominid features of adult human astrocytes. J Neurosci. 2009;29:3276–3287. doi: 10.1523/JNEUROSCI.4707-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Retzius G. Die neuroglia des gehirns beim menschen und bei saeugethieren. Biol Untersuchungen. 1894;6:1–28. [Google Scholar]
- 48.Colombo JA, Reisin HD. Interlaminar astroglia of the cerebral cortex: a marker of the primate brain. Brain Res. 2004;1006:126–131. doi: 10.1016/j.brainres.2004.02.003. [DOI] [PubMed] [Google Scholar]
- 49.Colombo JA, Yanez A, Puissant V, Lipina S. Long, interlaminar astroglial cell processes in the cortex of adult monkeys. J Neurosci Res. 1995;40:551–556. doi: 10.1002/jnr.490400414. [DOI] [PubMed] [Google Scholar]
- 50.Oberheim NA, Wang X, Goldman S, Nedergaard M. Astrocytic complexity distinguishes the human brain. Trends Neurosci. 2006;29:547–553. doi: 10.1016/j.tins.2006.08.004. [DOI] [PubMed] [Google Scholar]
- 51.Nishiyama A, Watanabe M, Yang Z, Bu J. Identity, distribution, and development of polydendrocytes: NG2-expressing glial cells. J Neurocytol. 2002;31:437–455. doi: 10.1023/A:1025783412651. [DOI] [PubMed] [Google Scholar]
- 52.Livet J, Weissman TA, Kang H, Draft RW, Lu J, Bennis RA, Sanes JR, Lichtman JW. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 2007;450:56–62. doi: 10.1038/nature06293. [DOI] [PubMed] [Google Scholar]
- 53.Halassa MM, Fellin T, Takano H, Dong JH, Haydon PG. Synaptic islands defined by the territory of a single astrocyte. J Neurosci. 2007;27:6473–6477. doi: 10.1523/JNEUROSCI.1419-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Oberheim NA, Tian GF, Han X, Peng W, Takano T, Ransom B, Nedergaard M. Loss of astrocytic domain organization in the epileptic brain. J Neurosci. 2008;28:3264–3276. doi: 10.1523/JNEUROSCI.4980-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Benediktsson AM, Schachtele SJ, Green SH, Dailey ME. Ballistic labeling and dynamic imaging of astrocytes in organotypic hippocampal slice cultures. J Neurosci Methods. 2005;141:41–53. doi: 10.1016/j.jneumeth.2004.05.013. [DOI] [PubMed] [Google Scholar]
- 56.Hirrlinger J, Hulsmann S, Kirchhoff F. Astroglial processes show spontaneous motility at active synaptic terminals in situ. Eur J Neurosci. 2004;20:2235–2239. doi: 10.1111/j.1460-9568.2004.03689.x. [DOI] [PubMed] [Google Scholar]
- 57.Hewett JA. Determinants of regional and local diversity within the astroglial lineage of the normal central nervous system. J Neurochem. 2009;110:1717–1736. doi: 10.1111/j.1471-4159.2009.06288.x. [DOI] [PubMed] [Google Scholar]
- 58.Silver J, Miller JH. Regeneration beyond the glial scar. Nat Rev Neurosci. 2004;5:146–156. doi: 10.1038/nrn1326. [DOI] [PubMed] [Google Scholar]
- 59.Pekny M, Nilsson M. Astrocyte activation and reactive gliosis. Glia. 2005;50:427–434. doi: 10.1002/glia.20207. [DOI] [PubMed] [Google Scholar]
- 60.Sofroniew MV. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009;32(12):638–647. doi: 10.1016/j.tins.2009.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bush TG, Puvanachandra N, Horner CH, Polito A, Ostenfeld T, Svendsen CN, Mucke L, Johnson MH, Sofroniew MV. Leukocyte infiltration, neuronal degeneration and neurite outgrowth after ablation of scar-forming, reactive astrocytes in adult transgenic mice. Neuron. 1999;23:297–308. doi: 10.1016/S0896-6273(00)80781-3. [DOI] [PubMed] [Google Scholar]
- 62.Herrmann JE, Imura T, Song B, Qi J, Ao Y, Nguyen TK, Korsak RA, Takeda K, Akira S, Sofroniew MV. STAT3 Is a critical regulator of astrogliosis and scar formation after spinal cord injury. J Neurosci. 2008;28:7231–7243. doi: 10.1523/JNEUROSCI.1709-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Brambilla R, Bracchi-Ricard V, Hu WH, Frydel B, Bramwell A, Karmally S, Green EJ, Bethea JR. Inhibition of astroglial nuclear factor kappaB reduces inflammation and improves functional recovery after spinal cord injury. J Exp Med. 2005;202:145–156. doi: 10.1084/jem.20041918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Codeluppi S, Svensson CI, Hefferan MP, Valencia F, Silldorff MD, Oshiro M, Marsala M, Pasquale EB. The rheb-mTOR pathway is upregulated in reactive astrocytes of the injured spinal cord. J Neurosci. 2009;29:1093–1104. doi: 10.1523/JNEUROSCI.4103-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Chen Y, Miles DK, Hoang T, Shi J, Hurlock E, Kernie SG, Lu QR. The basic helix-loop-helix transcription factor olig2 is critical for reactive astrocyte proliferation after cortical injury. J Neurosci. 2008;28:10983–10989. doi: 10.1523/JNEUROSCI.3545-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Eddleston M, Mucke L. Molecular profile of reactive astrocytes -implications for their role in neurological disease. Neuroscience. 1993;54:15–36. doi: 10.1016/0306-4522(93)90380-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Swanson RA, Ying W, Kauppinen TM. Astrocyte influences on ischemic neuronal death. Curr Mol Med. 2004;4:193–205. doi: 10.2174/1566524043479185. [DOI] [PubMed] [Google Scholar]
- 68.Chen Y, Vartiainen NE, Ying W, Chan PH, Koistinaho J, Swanson RA. Astrocytes protect neurons from nitric oxide toxicity by a glutathione-dependent mechanism. J Neurochem. 2001;77:1601–1610. doi: 10.1046/j.1471-4159.2001.00374.x. [DOI] [PubMed] [Google Scholar]
- 69.John GR, Lee SC, Brosnan CF. Cytokines: powerful regulators of glial cell activation. Neuroscientist. 2003;9:10–22. doi: 10.1177/1073858402239587. [DOI] [PubMed] [Google Scholar]
- 70.Zador Z, Stiver S, Wang V, Manley GT. Role of aquaporin-4 in cerebral edema and stroke. Handb Exp Pharmacol. 2009;190:159–170. doi: 10.1007/978-3-540-79885-9_7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Simard M, Nedergaard M. The neurobiology of glia in the context of water and ion homeostasis. Neuroscience. 2004;129:877–896. doi: 10.1016/j.neuroscience.2004.09.053. [DOI] [PubMed] [Google Scholar]
- 72.Maragakis NJ, Rothstein JD. Mechanisms of disease: astrocytes in neurodegenerative disease. Nat Clin Pract Neurol. 2006;2:679–689. doi: 10.1038/ncpneuro0355. [DOI] [PubMed] [Google Scholar]
- 73.Rothstein JD, Dykes-Hoberg M, Pardo CA, Bristol LA, Jin L, Kuncl RW, Kanai Y, Hediger MA, Wang Y, Schielke JP, Welty DF. Knockout of glutamate transporters reveals a major role for astroglial transport in excitotoxicity and clearance of glutamate. Neuron. 1996;16:675–686. doi: 10.1016/S0896-6273(00)80086-0. [DOI] [PubMed] [Google Scholar]
- 74.Iadecola C, Nedergaard M. Glial regulation of the cerebral microvasculature. Nat Neurosci. 2007;10:1369–1376. doi: 10.1038/nn2003. [DOI] [PubMed] [Google Scholar]
- 75.Gordon GR, Mulligan SJ, MacVicar BA. Astrocyte control of the cerebrovasculature. Glia. 2007;55:1214–1221. doi: 10.1002/glia.20543. [DOI] [PubMed] [Google Scholar]
- 76.Pellerin L, Bouzier-Sore AK, Aubert A, Serres S, Merle M, Costalat R, Magistretti PJ. Activity-dependent regulation of energy metabolism by astrocytes: an update. Glia. 2007;55:1251–1262. doi: 10.1002/glia.20528. [DOI] [PubMed] [Google Scholar]
- 77.Christopherson KS, Ullian EM, Stokes CC, Mullowney CE, Hell JW, Agah A, Lawler J, Mosher DF, Bornstein P, Barres BA. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell. 2005;120:421–433. doi: 10.1016/j.cell.2004.12.020. [DOI] [PubMed] [Google Scholar]
- 78.Stevens B, Allen NJ, Vazquez LE, Howell GR, Christopherson KS, Nouri N, Micheva KD, Mehalow AK, Huberman AD, Stafford B, Sher A, Litke AM, Lambris JD, Smith SJ, John SW, Barres BA. The classical complement cascade mediates CNS synapse elimination. Cell. 2007;131:1164–1178. doi: 10.1016/j.cell.2007.10.036. [DOI] [PubMed] [Google Scholar]
- 79.Hamby ME, Hewett JA, Hewett SJ. TGF-beta1 potentiates astrocytic nitric oxide production by expanding the population of astrocytes that express NOS-2. Glia. 2006;54:566–577. doi: 10.1002/glia.20411. [DOI] [PubMed] [Google Scholar]
- 80.Lee S, Park JY, Lee WH, Kim H, Park HC, Mori K, Suk K. Lipocalin-2 is an autocrine mediator of reactive astrocytosis. J Neurosci. 2009;29:234–249. doi: 10.1523/JNEUROSCI.5273-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gadea A, Schinelli S, Gallo V. Endothelin-1 regulates astrocyte proliferation and reactive gliosis via a JNK/c-Jun signaling pathway. J Neurosci. 2008;28:2394–2408. doi: 10.1523/JNEUROSCI.5652-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Panenka W, Jijon H, Herx LM, Armstrong JN, Feighan D, Wei T, Yong VW, Ransohoff RM, MacVicar BA. P2X7-Like receptor activation in astrocytes increases Chemokine Monocyte chemoattractant protein-1 expression via mitogen-activated protein kinase. J Neurosci. 2001;21:7135–7142. doi: 10.1523/JNEUROSCI.21-18-07135.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Farina C, Aloisi F, Meinl E. Astrocytes are active players in cerebral innate immunity. Trends Immunol. 2007;28:138–145. doi: 10.1016/j.it.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 84.Bekar LK, He W, Nedergaard M. Locus coeruleus alpha-adrenergic-mediated activation of cortical astrocytes in vivo. Cereb Cortex. 2008;18:2789–2795. doi: 10.1093/cercor/bhn040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Simpson JE, Ince PG, Lace G, Forster G, Shaw PJ, Matthews F, Savva G, Brayne C, Wharton SB. Neurobiol Aging. 2008. Astrocyte phenotype in relation to Alzheimer-type pathology in the ageing brain. [DOI] [PubMed] [Google Scholar]
- 86.Neary JT, Kang Y, Willoughby KA, Ellis EF. Activation of extracellular signal-regulated kinase by stretch-induced injury in astrocytes involves extracellular ATP and P2 purinergic receptors. J Neurosci. 2003;23:2348–2356. doi: 10.1523/JNEUROSCI.23-06-02348.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.De Keyser J, Mostert JP, Koch MW. Dysfunctional astrocytes as key players in the pathogenesis of central nervous system disorders. J Neurol Sci. 2008;267:3–16. doi: 10.1016/j.jns.2007.08.044. [DOI] [PubMed] [Google Scholar]
- 88.Gris P, Tighe A, Levin D, Sharma R, Brown A. Transcriptional regulation of scar gene expression in primary astrocytes. Glia. 2007;55:1145–1155. doi: 10.1002/glia.20537. [DOI] [PubMed] [Google Scholar]
- 89.Voskuhl RR, Peterson RS, Song B, Ao Y, Morales LB, Tiwari-Woodruff S, Sofroniew MV. J Neurosci. 2009. Reactive astrocytes form scar-like barriers to leukocytes during adaptive immune inflammation of the central nervous system. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Hamby ME, Sofronew MV. Reactive astrocytes as therapeutic targets for CNS disorders. Neurotherapeutics. 2010;7(4):494–506. doi: 10.1016/j.nurt.2010.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]