

Abstract
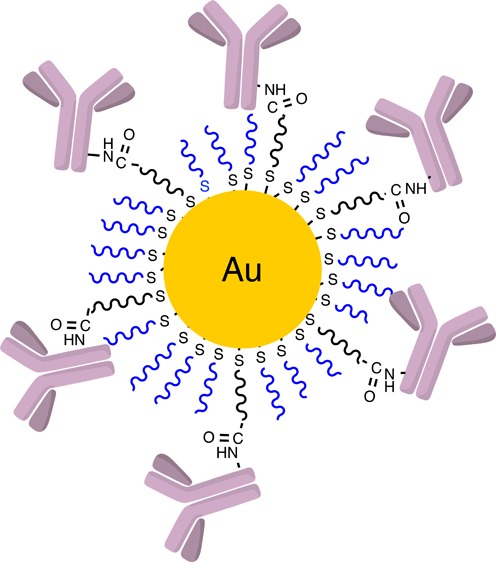
Antibody-dependent cellular cytotoxicity (ADCC) is a cytolytic mechanism that can elicit in vivo antitumor effects and can play a significant role in the efficacy of antibody treatments for cancer. Here, we prepared cetuximab, panitumumab, and rituximab containing gold nanoparticles and investigated their ability to produce an ADCC effect in vivo. Cetuximab treatment of EGFR-expressing H1975 tumor xenografts showed significant tumor regression due to the ADCC activity of the antibody in vivo, while the control antibody, panitumumab, did not. However, all three antibody containing nanoparticles are not able to suppress tumor growth in the same in vivo mouse model. The antibody containing nanoparticles localized in the tumors and did not suppress the immune function of the animals, so the lack of tumor growth suppression of the cetuximab containing nanoparticle suggests that immobilizing antibodies onto a nanoparticle significantly decreases the ability of the antibody to promote an ADCC response.
Introduction
A number of targeted nanoparticles have now been investigated in human clinical trials.1,2 At this time, there is no clinical example of a full antibody targeted nanoparticle. Since immunotherapies are finding increased importance in cancer, the use of a full antibody targeted nanoparticle could be interesting. This type of therapeutic could potentially elicit immunotherapeutic functions such as antibody-dependent cellular cytotoxicity (ADCC) and complement dependent cytotoxicity (CDC) in addition to targeting the nanoparticles to cancer cell surface receptors and blocking cell signaling from those receptors. While antibody fragments can elicit the latter two functions, they do not stimulate immunotherapeutic pathways.
Numerous preclinical studies utilize full antibody targeted nanoparticles. However, only one investigation has specifically explored the possibility of stimulating an ADCC response.3 Rituximab is an IgG1 antibody that binds to the CD20 receptor, and rituximab containing lipid nanoparticles were investigated both in vitro and in vivo for their ability to elicit ADCC. Rituximab nanoparticles exhibited ADCC cell lysis in vitro, but the observed in vivo therapeutic efficacy of the antibody–lipid conjugates could not be ascribed to ADCC function.3
Natural killer (NK) cell based immunotherapies have shown considerable potential for cancer therapy in the clinic.4,5 ADCC is an immune mechanism dependent on the activity of CD56dim CD16+ NK cells. Transgenic mouse models deficient in the CD16 receptor, also known as the activating Fcγ (FcγRIIIa/CD16) receptor, are unable to inhibit tumor growth in the presence of IgG1 antibodies that primarily work by inducing an ADCC response.6 Various types of peripheral blood mononuclear cells (PBMCs) have been studied for their antitumor ADCC activities in vitro, and NK cells have been found to induce the most potent ADCC response.7
Cetuximab and panitumumab are two antibodies that specifically target the epidermal growth factor receptor I (EGFRI) and possess similar EGFR binding affinities.8,9 In contrast to cetuximab, panitumumab is not able to elicit an ADCC response.10 Here, we address the question as to whether full antibodies that are displayed on the surface of nanoparticles can elicit an ADCC response in vivo. In order to observe antitumor effects that would be specific to an ADCC response, we selected a lung cancer cell line (H1975) that does not show any in vitro antiproliferative effects upon treatment with either cetuximab or panitumumab. Thus, any antitumor behavior observed in vivo can be ascribed to an ADCC function (positive for cetuximab and negative for panitumumab). Since gold nanoparticles will not have antitumor effects, antibody containing gold nanoparticles were prepared using cetuximab, panitumumab, and rituximab (negative control) and investigated in vivo with xenografts of the EGFR-expressing H1975 lung cancer cell line in athymic nude mice. While cetuximab alone reveals significant ADCC dependent antitumor behavior, the lack of antitumor function with the cetuximab containing gold nanoparticles shows that the ADCC function from antibody containing nanoparticles maybe be difficult to achieve in vivo.
Results and Discussion
Assembly of Antibody Containing Gold Nanoparticle
The assembly of the antibody containing gold nanoparticles was accomplished as follows. Conjugates of polyethylene glycol (PEG) and cetuximab, panitumumab, and rituximab were prepared by antibody reaction with NHS–PEG–OPSS (reacts with amine groups of antibodies to yield antibody–PEG conjugates through amide bond formation (Scheme 1)). High-pressure liquid-phase chromatography (HPLC) purified antibody–PEG conjugates were analyzed by MALDI-TOF-MS and confirmed to be mono-PEGylated. 50 nm gold nanoparticles (AuNPs) were then functionalized with the mono-PEGylated antibody–PEG conjugates and mPEG-SH (Scheme 1) and were analyzed for their average hydrodynamic diameter and surface charge (Table 1). The quantitative number of antibodies per nanoparticle was obtained using two different methods. The results from the two were consistent with each other (Supporting Information, Table S1), and the mean values obtained from the two methods are presented in Table 1. PEGylated AuNPs containing approximately 15–20 antibodies per particle have negative ζ potential values and are stable in deionized water and saline solutions.
Scheme 1. Assembly of Antibody Containing Gold Nanoparticles.

(A) Antibodies first reacted with the NHS–PEG–OPSS and then purified. (B) In a second step, the antibody conjugates were then combined with mPEG-SH and assembled onto the surface of the gold nanoparticles.
Table 1. Properties of Antibody Containing AuNPs.
| sample | size (nm) by DLSa | size (nm) by NTAb | ζ potential (mV)c | no. of antibodies on surface |
|---|---|---|---|---|
| cetuximab–AuNPs | 79.5 ± 4 | 72.6 ± 0.4 | –18.7 ± 1 | 19 ± 6 |
| panitumumab–AuNPs | 72.8 ± 1.9 | 74.8 ± 1.6 | –19.2 ± 2.1 | 20 ± 5 |
| rituximab–AuNPs | 82.5 ± 5.5 | 73.2 ± 1.3 | –20.5 ± 1.7 | 17 ± 3 |
| mPEG–AuNPs | 64.1 ± 9.5 | 72.3 ± 0.9 | –19.4 ± 4.3 | 0 |
DLS (Dynamic Light Scattering), measured in phosphate buffered saline (PBS).
NTA (Nanoparticle Tracking Analysis), measured in deionized water.
Measured in 1 mM KCl solution in deionized water.
In Vitro Cytotoxicity of Antibodies and Antibody-Functionalized Gold Nanoparticles
The H1975 cell line harbors a double mutation in the kinase domain of EGFR that makes these cells insensitive to treatments with tyrosine kinase inhibitors (TKIs). Also, EGFR specific monoclonal antibodies do not produce antiproliferative effects with the H1975 cells in vitro.11 The in vivo antitumor effects observed with cetuximab are thought to be associated with the ADCC activity of the antibody. Here, the antiproliferative effects of the antibodies cetuximab, panitumumab, and rituximab and the corresponding antibody containing AuNPs were investigated with the EGFR-TKI resistant H1975 lung cancer cell line via a cell viability assay. The H1975 cells remained viable in the presence of all antibodies and antibody containing AuNPs (Supporting Information, Figure S1) 72 h post-treatment. The results are consistent with previous reports that show no in vitro cell death upon exposure of H1975 lung cancer cells to cetuximab or panitumumab10 and reveal that multiple antibodies on the surface of the AuNPs also do not produce any antiproliferative effects in vitro.
Antitumor Effects of Antibodies and Antibody Containing AuNPs
To investigate the role of ADCC in vivo, mice bearing established H1975 xenograft tumors were dosed twice a week for 2 weeks, and tumor growth was measured over a period of 2 months. Cetuximab treated mice showed significant tumor regression, while panitumumab treated mice did not (Figure 1). Thus, cetuximab treatments inhibit in vivo H1975 tumor growth, and these results are consistent with literature reports.
Figure 1.

Effects of antibody treatment on H1975 xenograft tumors in nude mice. Mice were treated with cetuximab (circles), panitumumab (squares), or saline (triangles) as a negative control. Arrows indicate the days treatment occurred.
Antibody containing AuNPs were investigated for their antitumor activities in nude mice bearing H1975 tumor xenografts in order to compare to the results of treatments with the antibodies alone. The data illustrated in Figure 2 show that neither the cetuximab nor panitumumab containing AuNPs produced an antitumor effect in vivo (compared to saline, rituximab containing AuNPs, and AuNPs with no antibody (mPEG AuNP)). These results show that attaching the antibody to the surface of the AuNPs abolished cetuximab’s ADCC activity in vivo.
Figure 2.

H1975 xenograft tumor growth in mice treated with antibody containing AuNPs. Nonantibody containing AuNPs, rituximab containing AuNPs, and saline were used as negative controls. Arrows indicate the days treatment occurred.
Lack of ADCC with Cetuximab Containing AuNPs Is Not Due to Loss of Tumor Accumulation or Presence of Immune Cells
The strong tumor regression observed in cetuximab treated mice (Figure 1) is attributed to the ADCC activity of cetuximab. As expected, no antitumor effects are observed with panitumumab, which does not elicit an ADCC effect. This difference in the antitumor effect between the two antibodies was completely abolished when they were attached to the AuNPs (Figure 2). Unfortunately, we were not able to construct an in vitro model system for testing the ADCC activity of cetuximab or panitumumab. However, we were able to do so for trastuzumab using the BT474M1 cell line and immortalized NK cells (Supporting Information, Materials and Methods). These model data show that trastuzumab and trastuzumab AuNPs give an in vitro ADCC response (Supporting Information, Figure S2). Since we used the same conjugation chemistry with cetuximab and panitumumab as with trastuzumab, we assume that at least some fraction of the antibodies displayed on the surfaces of the nanoparticles are able to bind to their cancer cell surface receptors and stimulate an ADCC response in vitro. Thus, in order to better understand the factors that cause the in vivo loss in antitumor effects for the cetuximab that is contained on the AuNPs, we must first prove that the AuNPs are in fact reaching the tumors and that these tumors possess immune cells.
AuNPs in tissue can be visualized by silver staining and imaging by light microscopy. Images of silver stained tissues harvested from mice treated with antibody containing AuNPs (Supporting Information, Figure S3) show the presence of AuNPs in all treated tumor tissues, as well as in liver, spleen, and small amounts in kidney (nontumor organs were imaged to confirm that the AuNPs were reaching all organs expected with NPs). No staining was evident for tissues from mice treated with saline (negative control). Thus, the AuNPs are localizing to the tumors in these mice. The presence of immune cells within the tumor tissue was investigated by staining for CD45 (pan-leukocyte) and CD11b (NK) immune cell markers. In comparison to saline treated tumors, no significant difference in the presence of immune cells in tumor sections was found for antibody or antibody containing AuNP treated mice (Supporting Information, Figures S4 and S5).
Lack of ADCC with Cetuximab Containing AuNPs Is Due to Loss of ADCC Function
The in vivo antitumor data shown here reveal that an antibody that can elicit an ADCC effect when used alone loses this function when it is conjugated onto a nanoparticle. Cetuximab injected at 9.3 mg/kg showed significant tumor regression due to an ADCC effect, while cetuximab containing AuNPs dosed at 0.44 mg of antibody/kg did not show any antitumor effects. It should be noted that cetuximab maintains antitumor activities in nude mice over the range of 0.4–40 mg/kg.12
The mice dosed with 0.44 mg/kg of cetuximab containing AuNPs (0.299 nmoles of gold/gram of body weight) showed significant accumulation of AuNPs in dermis, spleen, liver, and kidney, as revealed by sharp changes in skin color and silver staining of tissues from treated mice. Similar deposition of targeted and nontargeted PEGylated AuNPs in mouse dermis and lymph nodes was previously observed when studied at a much lower dose of AuNPs per mouse.13 Most importantly, the AuNPs also trafficked to the tumors (Supporting Information, Figure S3). Thus, the lack of antitumor effects with the cetuximab containing AuNPs is not because the NPs did not reach the tumor.
It has been argued that introducing NPs into animals can modify their immune response via secretion of anti- or proinflammatory cytokines, which in turn has an impact on immune cell surveillance in tumor tissues.14−16 Here, the presence of immune cells in tumor tissues was studied using antibodies against CD45 (for all leukocytes) and CD11b (for mouse NK cells). Immune cell surveillance in the tumors used here was not significantly different regardless of treatment (Supporting Information, Figures S4 and S5). CD45 and CD11b labeled immune cells were abundant in H1975 tumor xenografts, indicating that the inability of the cetuximab containing AuNPs to show an antitumor effect in vivo was not due to a suppression of the animal’s immune response.
The results presented here lead to the conclusion that covalent immobilization of an antibody on a nanoparticle surface can eliminate the ADCC function of the antibody. Here, we conjugated the antibody to PEG via reactions with amines on the antibody. Because of the presence of several amines on the antibody, the conjugation of the PEG component will occur at various sites on the antibody. The antibody–PEG conjugates were purified by HPLC to obtain a mono-PEGylated fraction for attachment to the nanoparticle surface. However, the orientation and distribution of antibodies on the nanoparticle surface, such as whether the Fc region is accessible to immune cell binding may play an important role in eliminating the ADCC effects in vivo. Results from a model in vitro system suggest that at least a fraction of the antibodies that are contained on the surfaces of the gold nanoparticles can bind to the appropriate cancer cell surface receptor and stimulate an in vitro ADCC response. Thus, at this time, the mechanistic origin of the in vivo loss in ADCC with nanoparticle containing antibodies remains unknown. The ability to perform site-specific conjugation to the antibody like what is presently done with antibody–drug conjugates17 may help resolve some of these issues.
Acknowledgments
We thank Kristin Anderson for performing MTS cell viability assays, Mona Shahgholi for MALDI-TOF-MS training, and Andres Collazo (Beckman Imaging Center) for confocal microscopy trainning. This work was supported by the National Cancer Institute Grant CA 151819 and the National Science and Engineering Research Council of Canada (NSERC) (fellowship for M.A.).
Supporting Information Available
Materials and instrumentation, experimental procedures, and analytical data. This material is available free of charge via the Internet at http://pubs.acs.org.
Author Contributions
† M.A. and D.W.P. contributed equally to this work.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- Davis M. E.; Chen Z. G.; Shin D. M. (2008) Nanoparticle therapeutics: an emerging treatment modality for cancer. Nat. Rev. Drug Discovery 7, 771–782. [DOI] [PubMed] [Google Scholar]
- Bertrand N.; Wu J.; Xu X.; Kamaly N.; Farokhzad O. C. (2014) Cancer nanotechnology: The impact of passive and active targeting in the ear of modern cancer biology. Adv. Drug Delivery Rev. 66, 2–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popov J.; Kapanen A. I.; Turner C.; Ng R.; Tucker C.; Chiu G.; Klasa R.; Bally M. B.; Chikh G. (2011) Multivalent rituximab lipid nanoparticles as improved lymphoma therapies: indirect mechanisms of action and in vivo activity. Nanomedicine (London, U. K.) 6, 1575–1591. [DOI] [PubMed] [Google Scholar]
- Roberti M. P.; Mordoh J.; Levy E. M. (2012) Biological role of NK cells and immuno-therapeutic approaches in breast cancer. Front. Immunol. 3, 375. 10.3389/fimmu.2012.00375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng M.; Chen Y.; Xiao W.; Sun R.; Tian Z. (2013) NK cell-based immunotherapy for malignant diseases. Cell. Mol. Immunol. 10, 230–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clynes R. A.; Towers T. L.; Presta L. G.; Ravetch J. V. (2000) Inhibitory Fc receptors modulate in vivo cytoxicity against tumor targets. Nat. Med. 6, 443–446. [DOI] [PubMed] [Google Scholar]
- Seidel U. J. E.; Schlegel P.; Lang P. (2013) Natural killer cell mediated antibody-dependent cellular cytotoxicity in tumor immunotherapy with therapeutic antibodies. Front. Immunol. 4, 76. 10.3389/fimmu.2013.00076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voigt M.; Braig F.; Gothel M.; Schulte A.; Lamszus K.; Bokemeyer C.; Binder M. (2012) Functional dissection of the epidermal growth factor receptor epitopes targeted by panitumumab and cetuximab. Neoplasia 14, 1023–1031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel D.; Guo X.; Ng S.; Melchior M.; Balderes P.; Burtrum D.; Persaud K.; Luna X.; Ludwig D. L.; Kang X. (2010) IgG isotype, glycosylation, and EGFR expression determine the induction of antibody-dependent cellular cytotoxicity in vitro by cetuximab. Hum. Antibodies 19, 89–99. [DOI] [PubMed] [Google Scholar]
- Lopez-Albaitero A.; Ferris R. L. (2007) Immune activation by epidermal growth factor receptor-specific monoclonal antibody therapy for head and neck cancer. Arch. Otolaryngol. Head Neck Surg. 133, 1277–1281. [DOI] [PubMed] [Google Scholar]
- Lu Y.; Liang K.; Li X.; Fan Z. (2007) Responses of cancer cells with wild-type or tyrosine kinase domain-mutated epidermal growth factor receptor (EGFR) to EGFR-targeted therapy are linked to downregulation of hypoxia-inducible factor-1a. Mol. Cancer 6, 63–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steiner P.; Joynes C.; Bassi R.; Wang S.; Tonra J. R.; Hadari Y. R.; Hicklin D. J. (2007) Tumor growth inhibition with cetuximab and chemotherapy in non-small cell lung cancer xenografts expressing wild-type and mutated epidermal growth factor receptor. Clin. Cancer Res. 13, 1540–1551. [DOI] [PubMed] [Google Scholar]
- Sykes E. A.; Dai Q.; Tsoi K. M.; Hwang D. M.; Chan W. C. W. (2012) Nanoparticle exposure in animals can be visualized in the skin and analyzed via skin biopsy. Nat. Commun. 13, 3796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai C.-Y.; Lu S.-L.; Hu C.-W.; Yeh C.-S.; Lee G.-B.; Lei H.-Y. (2012) Size-dependent attenuation of TLR9 signaling by gold nanoparticles in macrophages. J. Immunol. 188, 68–76. [DOI] [PubMed] [Google Scholar]
- Dobrovolskaia M. A.; McNeil S. E. (2007) Immunological properties of engineered nanomaterials. Nat. Nanotechnol. 2, 469–478. [DOI] [PubMed] [Google Scholar]
- Lin A. Y.; Almeida J. P. M.; Bear A.; Liu N.; Luo L.; Foster A. E.; Drezek R. A. (2013) Gold nanoparticle delivery of modified CpG stimulates macrophages and inhibits tumor growth for enhanced immunotherapy. PLoS One 8, e63550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agarwal P.; Bertozzi C. R. (2015) Site-specific antibody-drug conjugates: the nexus of bioorthogonal chemistry, protein engineering, and drug development. Bioconjugate Chem. 26, 176–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.