

Abstract
Conservation biologists have only finite resources, and so must prioritise some species over others. The EDGE-listing approach ranks species according to their combined evolutionary distinctiveness and degree of threat, but ignores the uncertainty surrounding both threat and evolutionary distinctiveness. We develop a new family of measures for species, which we name EDAM, that incorporates evolutionary distinctiveness, the magnitude of decline, and the accuracy with which decline can be predicted. Further, we show how the method can be extended to explore phyogenetic uncertainty. Using the vascular plants of Britain as a case study, we find that the various EDAM measures emphasise different species and parts of Britain, and that phylogenetic uncertainty can strongly affect the prioritisation scores of some species.
Introduction
Global biodiversity is declining [1, 2], forcing conservation biologists to prioritise their finite conservation budgets in order to save as many species as possible (the ‘Noah’s Ark problem’;[3]). The EDGE approach [4] prioritises species according to a combination of how evolutionary distinctive (‘ED’) and how globally endangered (‘GE’) they are. EDGE listing is a popular approach (cited over 100 times on Web of Knowledge), and has been applied to mammals [4] and amphibians [5], and related methods have been used for birds [6, 7]. Evolutionary distinctiveness is an appealing metric to many biologists [8]; beauty and utility are difficult to quantify, but the millions of years of evolutionary history a species or clade uniquely represents are not.
EDGE listing has proven a useful tool, but its components are not optimised for local-scale prioritisation. Red Listing status (the GE component) is a global ranking, as its maintainer (IUCN) acknowledges by funding the ‘National Red List’ project (http://www.nationalredlist.org/). Similarly, calculating evolutionary distinctiveness with a global phylogeny will underestimate the national distinctiveness of species with close relatives living only in other countries [9]. Such clades might be distinctive and important local components of many ecosystems, but could paradoxically be prioritised in none of them.
More fundamentally, EDGE does not attempt to distinguish between the magnitude of a threat and the extent to which we understand that threat. A species undergoing a large decline that is understood to be a transient part of a natural cycle may be of lesser concern than a species undergoing a smaller decline that we do not understand or know how to reverse. Distinguishing among the finer points of species’ threats is impractical when dealing with the thousands of species in the global Red Lists, but individual countries have fewer species and often have more detailed, comparable within-country data.
We propose a family of prioritisation strategies (‘EDAM’) that extends the EDGE system, incorporating species’ evolutionary distinctiveness, magnitude of decline, and the accuracy with which we can predict declines in the absence of successful conservation intervention. Each of these components is transformed to be on a common numerical scale, making their implicit trade-off in EDGE explicit in EDAM. Using a novel genus-level phylogeny for the majority of vascular plant species in Britain, we report the species and parts of Britain that EDAM and EDGE prioritise.
EDAM framework
EDGE consists of two components: evolutionary distinctiveness (‘ED’; the phylogenetic diversity of a clade split equally among its members [4], which is related to how much branch length is unique to each species), and how globally endangered (‘GE’) a species is according to the IUCN Red List. We propose a general family of prioritisation indices (‘EDAM’), which incorporate ED, the accuracy with which decline (or threat) can be predicted (‘A’), and the magnitude of that decline (or threat) (‘M’). There are many ways of assessing all of these components; subscripts can be used to distinguish among them where there is ambiguity, but where there are no subscripts we use precision to measure accuracy, and range change to measure magnitude. We describe the specific evolutionary, accuracy, and magnitude components that we use in this study in the next section, and Table 1 summarises each of them as well. EDAM indices, like EDGE, are the sum of their components, but under EDAM each of these components is scaled to have a mean of zero and standard deviation of one. Thus, each component contributes equally to the measure, and judgements about which components are more important can readily be made explicit by multiplying components by a weighting factor.
Table 1. Summary of measures.
Note that all measures prioritise evolutionary distinctiveness (‘ED’ above), and that all components are scaled such that their means are zero and standard deviations one unless stated. As discussed in the text, in the EDAM framework EDGE values could be called EDM RL, although EDGE components are not scaled.
| Measure | Formula | Prioritises |
|---|---|---|
| EDGE (EDM RL) | unscaled ln(ED) + Red List | Global threat |
| EDM | ln(ED) + range change | British threat |
| EDAM | ln(ED) + range change + precision of decrease | Range change and low precision of decrease |
| EDAM′ | ln(ED) + reversed range change + precision of increase | Range change and low precision of increase |
| ED(AM)max | ln(ED) + max(range change, precision) | Greatest value of threat and accuracy of prediction |
An EDAM measure need not contain all three terms. For instance, EDM RL incorporates only evolutionary distinctiveness and Red Listing status; and differs from EDGE only in that its components are scaled. However, incorporates evolutionary distinctiveness and the negation of Red Listing status (indicated by the prime)—this would prioritise species that are evolutionarily distinct and not threatened. Such species are not of conservation concern, but evolutionarily distinct species with increasing ranges could be potentially damaging invasive species (as discussed with EDAM′ in the case study below). If components are to be multiplied by scaling factors, we suggest they are represented in the superscripts of the measures; for example, EDM 2 would weight the magnitude of decline as twice as important as evolutionary distinctiveness, and vice-versa for ED 2 M. This labelling scheme has been chosen such that superscript signifies multipliers of the quantitative effect of a term, while subscript denotes modifiers that indicate a different kind of measure. There are also precautionary measures such as ED(AM)max, for which evolutionary distinctiveness (ED) is added to the greatest of the accuracy (A) and magnitude (M) measures, emphasising species that are either declining rapidly, or with range dynamics we understand poorly. Such a measure would not be possible if both accuracy and magnitude had not been transformed to be on a common scale.
Weighting conservation actions according to the confidence we place in the composition of clades in a phylogeny (e.g., bootstrap values) would be unhelpful, but it is useful to know the effect that phylogenetic uncertainty has on prioritisation. Most methods of phylogeny construction produce a set of credible, but not optimal, trees, and repeating analyses across this subset accommodates phylogenetic uncertainty. Estimates could be weighted by the likelihood of the tree across which each measure was calculated, estimating the impact of phylogenetic uncertainty.
Case study: British vascular plants
EDAM measures have three components: evolutionary distinctiveness, accuracy of decline or threat, and magnitude of decline or threat. Below we describe how each of these components was generated, and then how each EDAM measure was calculated. For comparison, EDGE scores were also calculated. Note that the EDAM results are based on decline, whereas the Red Listing data upon which the EDGE scores are based somewhat blur the distinction between decline and threat depending upon the criterion under which a species is listed [10].
Phylogeny building (evolutionary distinctiveness)
Samples from 548 species were collected from natural British populations, each representing a different plant genus. All samples were collected as part of the UK Flora DNA Bank Project, under licence from Natural England. The rbcL locus was amplified either as a single fragment, or two overlapping, PCR product(s) to obtain the whole region. Ten primers were utilised in a number of different combinations to obtain rbcL sequences from diverse groups of plant taxa. Eight of these primers were from previous publications: 1F, 1460R (5′ TCC TTT TAG TAA AAG ATT GGG CCG AG 3′), 724R, 636F [11]; 724Rm [12]; 1360R [13]; 32F, 1367R [14]. The remaining primers were developed at RBG Kew (Genetics Section): 1FA (5′ ATG TCA CCA CAA ACA GAG AC 3′) and 627F (5′ CAT TTA TGC GCT GGA GAG ACC G 3′).
Double stranded PCR amplification of rbcL was performed in an ABI thermal cycler, using pre-made 2.5mM MgCl2 PCR Mastermix (ABgene), 14μM of forward and reverse primer, 1.0μl BSA (0.4% w/v), and between 50–100ng of total DNA, in 50μl reaction volumes. Thermal cycler conditions were (1) 96°C, 1min; (2) 96°C, 1min, (3) 48°C, 30sec, (4) 72°C, 1min; cycle (2)-(4) was repeated for 28 cycles, (5) 72°C, 7min; (6) 4°C. Products were cleaned using QIAquick PCR Purification Kit (Qiagen) according to the manufacturer’s protocol. These DNA fragments were sequenced using 2.0μl BigDye™ ABI PRISM™ (Perkin-Elmer), 10–20ng of PCR DNA, and either 5′ forward or 3′ reverse primer (1.4μM) in 5.25μl reaction volumes. ABI thermal cycler conditions for dye-terminating reactions were 96°c, 1min; 66°c, 5sec; 72°c, 45sec; for 26 cycles. The sequencing products were analysed on an ABI 3100 automated DNA Sequencer (Perkin-Elmer). Both 5′ (forward) and 3′ (reverse) DNA sequences were obtained from each PCR product, and assembled and edited using Autoassembler (Version 1.4.0).
All samples sequenced as above have been released onto GenBank [15] (see S1 File for accession numbers), but only 364 of them were used for this project. An additional 312 different rbcL sequences were downloaded from GenBank using phyloGenerator [16], meaning the alignment altogether represents 91% of native genera in Britain (according to PLANTATT; [17]). We constructed a family-level constraint tree based on the APG III classification [18] included with Phylomatic [19]. After these analyses were performed, fourteen of the sequences were found to have sequencing errors that caused frame-shifts at the extreme 3′ end of the sequences. The corrected sequences were uploaded to GenBank, but the uncorrected sequences, along with a demonstration that these sequence-changes do not alter phylogenies built with them, are presented in S1 File.
We aligned the sequences using MAFFT [20, 21], and chose the phylogeny with the greatest likelihood (under a GTR-γ DNA substitution model) from two separate RAxML [22] runs. Each run partitioned the alignment into three codon positions, used the ‘GTR-PSR’ (previously called ‘GTR-CAT’) DNA substitution model, and was constrained using the constraint tree described above. The first run used 500 random starting trees (the log likelihood of the best tree was −93656.22), and the second was an integrated rapid bootstrap search with 2000 random bootstrap searches and 400 subsequent thorough maximum likelihood searches (the log likelihood of the best tree was −93669.50). Since the best-scoring tree was found in the first search, we annotated that tree with the 2000 rapid bootstrap trees from the second search, and rate smoothed it using PATHd8 [23], setting the root age to 1.
Genera in the phylogeny containing more than one species were replaced with a polytomy containing all the species listed in PLANTATT in that genus, with the polytomy placed either half-way along the branch that led to the representative of that genus in the phylogeny, or at the 80th quantile of genus age in the phylogeny, whichever was smaller. This reduces bias introduced by particularly isolated sister species; many of the gymnosperm genera are distantly related to one another, and excessively long branches within genera could have biased the results. While more complex algorithms are available to assign ages to polytomies (e.g., [24]) the quantile at which the cut is made does not affect these results (see S1 Fig), and it is practical to apply this method to large numbers of phylogenies (see below).
Evolutionary distinctiveness was calculated across the phylogeny using the ed.calc function in caper [25] with the Isaac correction for polytomies. The natural logarithm of these ED values was used to calculate EDGE, and the same (log-transformed) data were scaled to have a mean of 0 and standard deviation of 1 for use in the EDAM measures. In the original EDGE list 1 was added to the ED values before taking their logarithm to normalise the result [4]; doing so was not necessary in this case since the data were normally distributed, and in fact would have made the data extremely non-normally distributed. The entire procedure of rate smoothing, species addition, and ED calculation was additionally performed using each of the 2000 phylogenies produced during the rapid bootstrap search.
Magnitude of decline or threat
We used the Telfer et al. [26] relative change index (RCI) to measure species decline, using the same methods as in [27]. Briefly, RCI was defined for each species as the residuals from a regression of the logit-transformed proportion of occupied 10km × 10km cells in 1987–1999 (taken from [28]) against the logit-transformed proportion of occupied cells in 1930–1969 (taken from an update to [29]). RCI is not an absolute measure of decline; the observed decline is relative to the other species in the dataset, but for the purpose of prioritisation this distinction is unimportant. Range change values were reversed so that greater numbers indicate a greater decline, and scaled to have a mean of 0 and standard deviation of 1. When calculating EDGE, Red List category (taken from the 2011 update of [30]) was treated as a continuous variable on a coarse scale (Least Concern = 0, Near Threatened and Conservation Dependent = 1, Vulnerable = 2, Endangered = 3, Critically Endangered = 4; as in [4]).
Accuracy of decline
Correlative species distribution models (SDMs) predict species’ potential range expansions and contractions based on the redistribution of the species’ realised environment over time and/or space [31]; the predictive accuracy of SDMs in new environmental domains is therefore a good measure of how well species’ declines may be predicted based on given environmental variables. We modelled distribution data at the 10km square resolution for each plant species in the period 1930–1969 as a function of climate and land use using generalised boosted models (GBMs; [32]); we then quantified the predictive accuracy of models by comparing model forecasts to independent observations for the period 1987–1999. We used the same underlying data and methods as [33], but in addition accounted for historic change in land use using data from [34] for the 1930–1969 period and [35] for 1987–1999 period.
The predictive accuracy of SDMs over time is generally calculated by comparing model predictions with independent observations using widespread discrimination measures such as the AUC (e.g., [36, 37]). Here, however, we use one of three new measures of temporal validation for SDMs—Acc TV [38]—which has two main advantages over most commonly-used alternatives: it makes use of modelled probabilities of presence over time directly, without requiring the choice of arbitrary probability thresholds, and it focuses on accuracy over portions of species’ ranges that have been either observed or predicted to change, thus quantifying a model’s ability to predict decline. Briefly, Acc TV can be derived from temporal validation (TV) plots, which are extensions of presence-absence calibration plots [39] for use with data from two time periods. TV plots model observed gains and losses as a function of changes in modelled probability of presence between time periods using natural splines (see Rapacciuolo et al. [38] for detailed methods). Ideally, summing interpolated gains and losses across values of change in modelled probability of presence should result in an ideal TV curve: a line with a slope of 1, passing through the origin (0, 0). Acc TV is the weighted average distance between the model’s and the ideal TV curve, subtracted from 1. It can be calculated using the following formula:
| (1) |
where y model and y ideal are the y values of the model curve and ideal curve, respectively, at each observed site q, and x q are the proportional changes in modelled probability of presence at each site q. Species’ Acc TV estimates were scaled to have a mean of zero and a standard deviation of one.
Calculating the measures
We calculated the measures listed in Table 1 using the range change and predictive accuracy data described above, and evolutionary distinctiveness values from the best-scoring phylogenetic tree. We compared EDGE and EDM by regressing them against each other and their components; all components of EDAM were uncorrelated with each other (all ∣r∣ < 0.11). Additionally, we calculated EDM across all 2000 rapid bootstrap trees, to assess the impact of phylogenetic uncertainty on its values.
Although the various measures may rank the importance of species differently, many conservation efforts are targeted at the habitat-level and so this variation might be of little importance if prioritised species frequently co-occur. To see whether the indices prioritised different parts of Britain, we map the top-fifty ranked species according to each index of the species within each grid cell of the Preston et al. distribution data for Britain [28]. Means of prioritisation scores are difficult to interpret due to shifts in species richness across Britain; however, maps of this mean richness are available in S2 Fig.
Results
Phylogeny
All novel rbcL sequences have been uploaded to GenBank [15], and the accession numbers of all sequences used are given in S1 File. The optimal phylogeny and the bootstrap phylogenies are available in S1 File (all rate-smoothed and in Newick format), as is a OneZoom [40] file that allows the tree to be interactively explored.
Comparing EDGE and EDM RC
EDGE and EDM were strongly correlated, but many more species were tied for EDGE values (Fig 1), suggesting that EDGE was less discriminating (reflecting the coarser nature of the Red List data). Both measures were significantly correlated with their components, but EDGE was more correlated with ED than Red Listing status (Fig 2a and 2b), suggesting EDGE is driven by ED in this dataset. EDM was correlated almost equally with ED and range change (Fig 2c and 2d), confirming that it reflects its two components equally.
Fig 1. Plot of EDGE scores against EDM, showing they are strongly correlated (red regression line; F 1,1051 = 1846, r 2 = 0.64, p < 0.0001).

The standard deviations of the EDM and EDGE values’ bootstrapped estimates are shown as grey whiskers around each point; these were not incorporated in the regression quoted above. There are 553 tied EDGE scores, and no tied EDM scores; a line of tied EDGE species can be seen at the left of the plot. The greater number of tied EDGE species suggest EDGE is worse at discriminating among species.
Fig 2. EDGE and EDM plotted against their components.

In (A) and (B), EDGE is plotted against Red List category (increasing in threat level from left-to-right; r 2 = 0.20) and the logarithm of evolutionary distinctiveness (r 2 = 0.78) respectively. The larger r 2 when regressed against log(ED) suggests EDGE is more strongly related to ED than GE. (C) and (D) show EDM against change index (r 2 = 0.47) and the logarithm of evolutionary distinctiveness (r 2 = 0.56) respectively. EDM is related almost equally strongly to its components, and so is less biased than EDGE. All models above were statistically significant (p < 0.0001) and were linear regressions, with the exception of comparison of EDGE and Red Lists status, which was an ANOVA. Note that EDM is calculated with scaled values, but is not regressed against them above.
The EDAM values were stable across the bootstrap phylogenies. All sets of EDAM values calculated using the bootstrapped phylogenies, when compared with the EDAM values from the optimal phylogeny, had correlation coefficients greater than 0.92. However, Fig 3a reveals EDM estimates for some species vary considerably across the bootstraps. The species with the greatest standard deviation of bootstrapped EDM is shown in Fig 3b, where three distinct groupings of EDM values can be seen.
Fig 3. Variability of EDM values across bootstrap phylogenies.

In (A), the ranks of the best phylogeny’s EDM values are plotted against the median ranks across all the bootstrap phylogenies, with grey whiskers showing the standard deviations of those ranks. (B) plots the rank of EDM for Wahlenbergia hederacea in each bootstrap phylogeny against the log. likelihood of that phylogeny. Wahlenbergia hederacea is the species with the largest standard deviation in (A). Three distinct groupings of EDM values can be seen in (B), probably reflecting equally likely islands in treespace (see discussion).
Conservation priorities
The highest ranking twenty species for each measure are listed in Tables 2 and 3; Fig 4 plots the values of each species under each metric against one-another for comparison. EDGE and EDAM-family values for all 1060 species in the study are available in S1 File. Although EDM, EDAM, and ED(AM)max species scores are correlated with one another, EDAM′, with its focus on range expansion, highlights a different set of species (Fig 4).
Table 2. Species rankings according to EDGE and EDM.
| EDM | EDGE |
|---|---|
| Selaginella selaginoides | Lycopodiella inundata |
| Pteridium aquilinum | Pilularia globulifera |
| Galeopsis angustifolia | Osmunda regalis |
| Pilularia globulifera | Selaginella selaginoides |
| Himantoglossum hircinum | Hymenophyllum wilsonii |
| Hymenophyllum wilsonii | Daphne mezereum |
| Sinapis arvensis | Wolffia arrhiza |
| Lycopodiella inundata | Isoetes echinospora |
| Botrychium lunaria | Isoetes lacustris |
| Osmunda regalis | Zostera noltei |
| Ranunculus arvensis | Adonis annua |
| Cryptogramma crispa | Spartina maritima |
| Huperzia selago | Astragalus danicus |
| Dioscorea communis | Cuscuta epithymum |
| Hymenophyllum tunbrigense | Botrychium lunaria |
| Oxalis acetosella | Frankenia laevis |
| Scleranthus annuus | Myriophyllum verticillatum |
| Mentha pulegium | Colchicum autumnale |
| Narthecium ossifragum | Ruppia cirrhosa |
| Tofieldia pusilla | Pteridium aquilinum |
Species rankings according to EDGE and EDM. The highest ranking species is listed first.
Table 3. Species rankings according to EDAM, EDAM′, and ED(AM)max.
| EDAM | EDAM′ | ED(AM)max |
|---|---|---|
| Himantoglossum hircinum | Polypodium vulgare | Cicendia filiformis |
| Cicendia filiformis | Tripleurospermum inodorum | Radiola linoides |
| Radiola linoides | Polygonum arenastrum | Lycopodiella inundata |
| Lycopodiella inundata | Polypodium interjectum | Osmunda regalis |
| Mentha pulegium | Sedum album | Zostera marina |
| Zostera marina | Osmunda regalis | Selaginella selaginoides |
| Galeopsis angustifolia | Dryopteris expansa | Drosera intermedia |
| Spartina maritima | Tripleurospermum maritimum | Spartina maritima |
| Cuscuta epithymum | Tilia platyphyllos | Hymenophyllum tunbridgense |
| Scleranthus annuus | Isoetes echinospora | Adoxa moschatellina |
| Drosera intermedia | Polystichum setiferum | Pinguicula lusitanica |
| Pinguicula lusitanica | Papaver somniferum | Viola lactea |
| Jasione montana | Equisetum telmateia | Equisetum telmateia |
| Silene gallica | Agrostis stolonifera | Polypodium vulgare |
| Viola lactea | Polystichum aculeatum | Cuscuta epithymum |
| Hymenophyllum tunbrigense | Adoxa moschatellina | Pteridium aquilinum |
| Pteridium aquilinum | Vulpia ciliata | Galeopsis angustifolia |
| Oxalis acetosella | Cicendia filiformis | Narthecium ossifragum |
| Adonis annua | Radiola linoides | Pilularia globulifera |
| Erica tetralix | Lycopodiella inundata | Himantoglossum hircinum |
Species rankings according to EDAM, EDAM′, and ED(AM)max. The highest ranking species is listed first.
Fig 4.

Scatter plots of the EDAM measures against each another (lower panels), histograms of their distributions (along the diagonal), and correlation coefficients of the measures (upper panels). Note the poor correlation between EDAM′ and EDM.
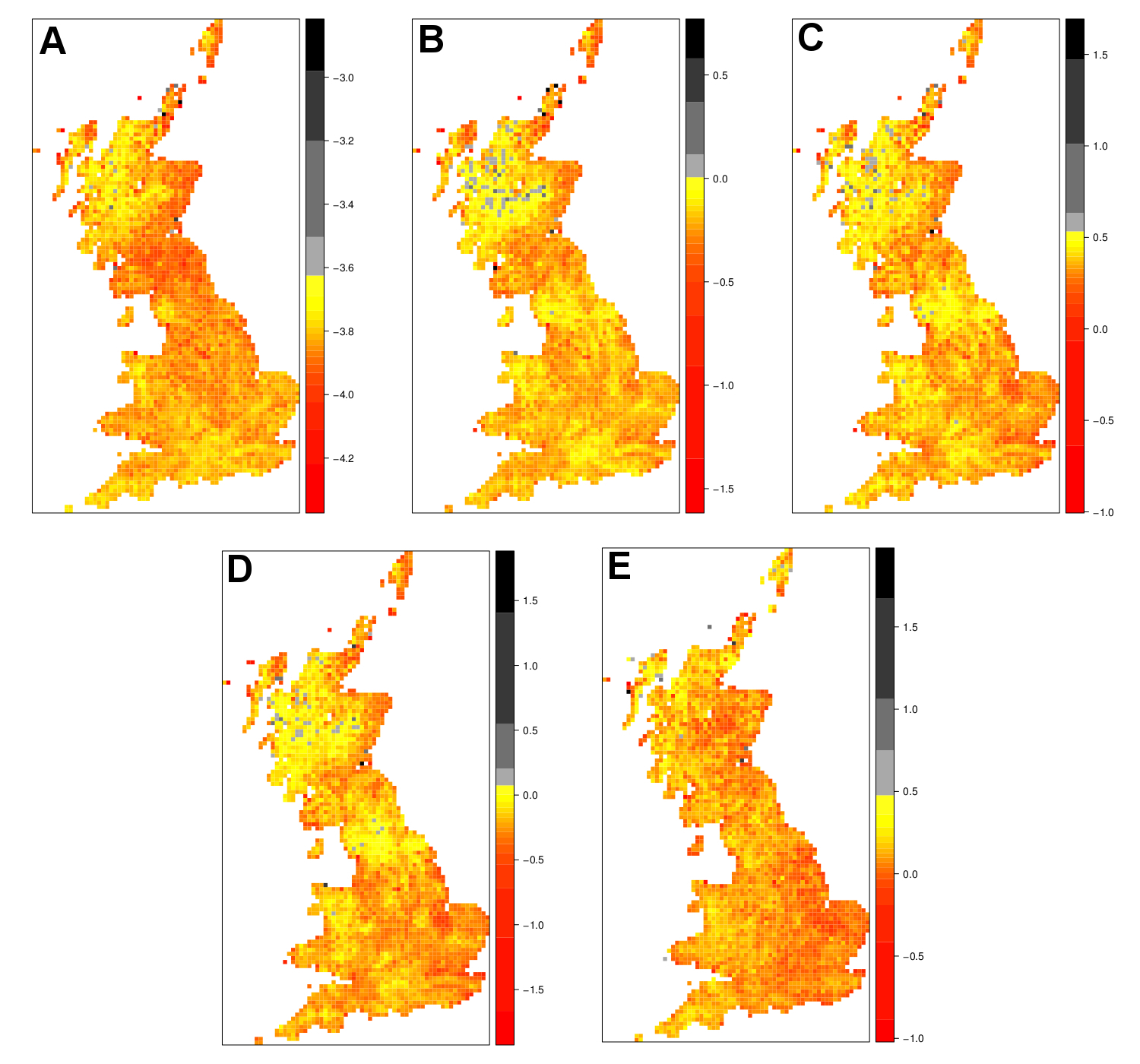
Maps of the distribution of the top-fifty species for each measure are shown in Fig 5. EDGE and EDM are correlated (r = 0.81, t 2811 = 72.12, p < 0.0001); both prioritise the Scottish Highlands, North of Wales, and patches of Northern and Southern England. In general, EDGE prioritises more diffuse areas of Britain, while more intense clusters of species are detected with EDM. EDAM and ED(AM)max highlight similar grid cells to EDM (correlations of grid cells’ values; all r > = 0.85, t 2811 > = 84.10, p < 0.0001). EDAM′ places a greater emphasis on England and Wales, prioritising large parts of the South-West and Wales. Maps of the mean scores of the species in each grid cell are shown in the S2 Fig.
Fig 5. Distribution of top-fifty ranked species across Britain.

The sub-panels show EDGE (A), EDM (B), ED(AM)max (C), EDAM (D), and EDAM′ (E). Each map has a legend to the right hand side, splitting the counts of species into twenty ‘Jenks’ quantiles (classIntervals function in the R package classInt [58]). The greatest four quantiles have been coloured differently, to emphasise the high-priority sites.
Discussion
Performance of EDAM
EDM was more discriminating than EDGE, which we suggest means EDM provides a more useful prioritisation of British plant species. If species are to be compared and prioritised, it is helpful if the scheme according to which they are compared is unambiguous as to the ranking of those species (there are no tied rankings). We also argue that, since EDM gives explicit (and, in our case, equal) weight to both evolutionary distinctiveness and decline, EDM gives a clearer prioritisation scheme. It is important to note that rankings mask the magnitude of differences among species; that one species out-ranks another does not mean it does so by much. We would advise against binning EDGE and EDAM lists (e.g., into ‘highly ranked’ and ‘low-ranked’ species, etc.) where possible since this would remove information from the data. It is better to rank species with more information and simply to be more cautious about closely-ranked species, and if necessary collect more information such as the likelihood of successful conservation intervention (see below).
The data required for the EDAM approach means it is unlikely to supplant EDGE as a global prioritisation scheme. Predictive accuracy is difficult to quantify, and species distribution models, although tractable and relatively quick to produce, typically ignore biotic interactions [41] and assume species’ distributions are at equilibrium with their environment [31]. However, other methods of assessing predictive accuracy, such as literature reviews and community-based models, can be prohibitively time-consuming and difficult to directly compare among taxa.
EDM ranks of some species vary greatly among the bootstrap phylogenies, which is concerning in species where there is no obvious relationship between EDM rank and the likelihood of the phylogeny across which it was calculated (Fig 3). In particular, the three distinct groupings in Fig 3b suggest that three islands in treespace were sampled (discussed in [42]); since each island seems equally likely, there may be no correct score for such species. Accurately prioritising species that are difficult to place in a phylogeny (rogue taxa; see [43]) without more sequence data may not be possible, potentially affecting the potential value of phylogenetic diversity (see [44]). However, rogue taxa are the exception, not the rule, and on the whole the bootstrap replicates were strongly correlated with the rankings of the best phylogeny. Rogue taxa pose no problem for a prioritisation system as long as they are identified, but testing prioritisation lists by recalculating across candidate trees (as in [5, 45]) will not detect them. Our phylogeny was built using a single DNA locus; future phylogenetic work with more data will likely revise our listing in ways we cannot readily predict or account for with randomisations.
Prioritisation
At first glance, the number of common species in the lists of prioritised species (e.g., Pteridium aquilinum, bracken; table 2) might be surprising. Such surprises reflect how evolutionarily distinct British non-angiosperm plants are relative to each other (i.e., ignoring close relatives outside the UK), but the measures also prioritise several severely declining species (e.g., Galeopsis angustifolia—red hemp-nettle). Limiting the lists to angiosperms or down-weighting evolutionary distinctiveness would alter the rankings if desired, and we consider it a strength of the EDAM (and EDGE) approach that we can make our decisions explicit in this way. More importantly, the purpose of a quantitative prioritisation exercise is not necessarily to produce a single, definitive list for conservation, but to help us consider how we prioritise nature. For example, Selaginella selaginoides is fairly uncharismatic (even for a clubmoss), yet it has the highest EDM score. This species is declining in Britain, and gives its name to an entire (declining) species group in one text [46], yet is not a UK Biodiversity Action Plan species [47] (and has not been added to the subsequent Section 41 list published under the Natural Environment and Rural Communities Act). S. selaginoides is widespread throughout mainland Europe but declining with Britain; these EDM rankings re-open the question of whether distinct, declining components of our flora should be conserved regardless of their status elsewhere.
Ours is not the first study to examine the phylogenetic pattern of threat in the UK flora (e.g., [48]), however, we are the first to prioritise species (not clades) and parts of the country. EDGE and EDM prioritise broadly similar parts of the Britain, but the intensity and resolution of prioritised areas is much greater for EDM. This greater intensity likely directly stems from the dependence of EDM on range change data and as such is unsurprising, but it shows the EDAM approach can give a more precise, and thus feasible, set of geographic priorities. The precautionary ED(AM)max measure seems intermediate between the EDM and EDAM, which is perhaps to be expected since it is essentially a compound measure, but EDAM′ is different in highlighting large parts of southern England. There is potentially a causal link between the high concentration of species with high EDM values in the Scottish Highlands (known to species with contracting southern ranges [27]) and the emphasis EDAM′ places on Southern England. As with the highlighting of S. selaginoides, we suggest this is another example of how systematic prioritisation can (re-)draw attention to potentially important conservation issues.
Given our EDAM approach allows for any arbitrary (explicit) relative weighting of its three components, it is reasonable to ask what weighting should be used in practice. We suggest that, while rankings will be sensitive to the weighting used, there is little utility in deciding empirically what the “correct” weighting is. Instead, weighting should be chosen a priori based on beliefs and wishes about the importance of species, or chosen as part of an ongoing discussion with stakeholders and the general public. Such an iterative, reflective process should reveal more about what we value about nature, and as such be of greater use than a static list generated by a distant stranger. Conservation prioritisation (triage), and the making of prioritisation lists, is controversial [49, 50]; we argue that the process of making lists forces us to confront our prior beliefs with hard data, and under an EDAM approach quantify and weight exactly what we value about biodiversity. We feel that weighting everything equally reflects more readily the naïve expectations of someone viewing a ranking, and that is why we have used it. There is no objective criterion by which a person’s subjective beliefs and values can be measured, and open dialogue is the best way to move through disagreement over beliefs. While we have made no attempt to measure the relative costs or likelihood of success of saving these species (c.f. [51]), we emphasise that such modifications to the EDAM approach are possible. An additional precautionary measure such as EDAM(CS)min, where species are additionally ranked according to the worst of their cost (C; which should be reversed as we reversed range change) and likelihood of success (S), could make a good continuation of the EDAM approach. EDAM (and EDGE; [52]) exhibit spatial pattern; spatial clumping of threatened species may hinder or help conservation, and this too could be incorporated in future prioritisation schemes.
Wider conservation issues
There is some evidence that species distantly related to an assemblage are more likely to invade [53], and more damaging when they do [54], although there are counter-examples [55]. More work is needed, but it is reasonable to highlight evolutionarily distinct species with expanding ranges as potential future problems, particularly given that distantly related species tend to be ecologically dissimilar (reviewed in [56]). Although precise ecological data on invasive species are often missing, invasives can usually be placed (perhaps coarsely) within a phylogenetic tree without much difficulty. Thus an EDAM approach could help identify potential problems, particularly in concert with information on the phylogenetic structure of protected areas. The UK has excellent data on the species composition of most protected areas, and our phylogeny based upon rbcL could be used for such analyses.
Prioritising species on the basis that their declines are poorly-understood might seem an unusual strategy for two reasons. Firstly, such species might be more difficult to save, although this could make them of greater academic interest and investigating them may uncover new conservation techniques. Secondly, it might seem better to weight the decline of a species according to our confidence in that decline, rather than treat magnitude and accuracy as distinct. However, to do so would not necessarily be precautionary (see [57]), and could lead us to prioritise species on the basis of sampling effort. Uncertainty is already enshrined in the Red List criteria [10]; for example, a species qualifies as endangered under criterion A at a lower level of decline if that decline is not understood (and continuing). Accuracy of prediction (ideally) relates to our understanding of the drivers of range change, and, if desired, we could choose to prioritise species with declines we understand well enough to reverse. The EDAM framework could be extended to explicitly trade-off the cost of saving species with their distinctiveness or magnitude of threat, along with any other species-specific data a conservation biologist can quantify.
Conservationists can rarely achieve all their goals simultaneously, and instead several components (e.g., evolutionary distinctiveness, species diversity, financial cost, likelihood of success, and ecosystem services) must be traded off against one another. By scaling the components of EDAM so that each is on the same scale, we have a starting point from which we can explore the implications of prioritising different aspects of our biota, and make our subjective decisions more explicit. The EDAM lists presented here are designed for different purposes, and it is unlikely that a single priority list will ever suffice for British plants, let alone other threatened taxa. However, we feel that the success of the EDGE program demonstrates that incorporating evolutionary distinctiveness into conservation strategies strikes a chord with the majority of biologists; the measures we propose here allow uncertainty to be explicitly incorporated as well.
Supporting Information
Genera were cut into the best-scoring phylogeny as described in the text, but at each integer quantile of genus age, and the evolutionary distinctiveness scores of all species correlated with the scores when the genera were cut at the 1st quantile. In the figure, the correlation coefficients are plotted against quantile at which the cuts were made.
(PDF)
The sub-panels show EDGE (A), EDM (B), ED(AM)max (C), EDAM (D), and EDAM′ (E). Each map has a legend to the right hand side, splitting the values into twenty ‘Jenks’ quantiles (classIntervals function in the R package classInt [58]). The greatest four quantiles have been coloured differently, to emphasise the high-priority sites. Note that the distributions of all five measures have extremely long tails (as shown by the size of the quantiles in the legends).
(JPG)
{kind=link}
- analysed_accession_numbers.doc—Accession numbers of sequences used to build phylogeny
- bootstrap_trees.zip—All bootstrap replicates of phylogeny
- metrics.csv—CSV file with species scores under all metrics
- novel_sequence_accession_numbers.doc—Accession numbers of sequences uploaded to GenBank
- OneZoom.html—Best phylogeny, viewable in OneZoom, embedded in a web page (HTML file)
- phylogeny.tre—Best dated phylogeny in Newick format
- uncorrected_sequences.fasta—Uncorrected sequences used in analysis
- uncorrect_vs_corrected_sequence_check.doc—Demonstration that corrections made to sequences do not affect the results of the manuscript
(ZIP)
Acknowledgments
We are grateful to the Botanical Society of the British Isles, and all the volunteer recorders whose efforts went into the collection of the distribution data we used to calculate range change. We would like to thank two anonymous reviewers, R. Guralnick, and T. Barraclough and J. Silvertown, who provided extremely useful feedback on this manuscript. WDP was funded by a NERC CASE PhD studentship. This article is a contribution to Imperial College’s Grand Challenges in Ecosystems and the Environment initiative.
Data Availability
All relevant data, including Genbank Accession numbers, are in the paper and its Supporting Information files.
Funding Statement
WDP was funded by a Natural Environment Research Council (NERC) Collaborative Award in Science and Engineering (CASE) PhD studentship.
References
- 1. Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: synthesis. Island Press, Washington, DC. [Google Scholar]
- 2. Butchart SHM, Walpole M, Collen B, van Strien A, Scharlemann JPW, Rosamunde EA, et al. (2010) Global biodiversity: indicators of recent declines. Science 328: 1164–1168. 10.1126/science.1187512 [DOI] [PubMed] [Google Scholar]
- 3. Weitzman ML (1998) The Noah’s Ark problem. Econometrica 66: 1279–1298. 10.2307/2999617 [DOI] [Google Scholar]
- 4. Isaac NJB, Turvey ST, Collen B, Waterman C, Baillie JEM (2007) Mammals on the EDGE: conservation priorities based on threat and phylogeny. PLoS ONE 2: e296 10.1371/journal.pone.0000296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Isaac NJB, Redding DW, Meredith HM, Safi K (2012) Phylogenetically-informed priorities for amphibian conservation. PLoS ONE 7: e43912 10.1371/journal.pone.0043912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Redding DW, Mooers AØ (2006) Incorporating evolutionary measures into conservation prioritization. Conservation Biology 20: 1670–1678. 10.1111/j.1523-1739.2006.00555.x [DOI] [PubMed] [Google Scholar]
- 7. Jetz W, Thomas GH, Joy JB, Redding DW, Hartmann K, Mooers AØ. (2014) Global distribution and conservation of evolutionary distinctness in birds. Current Biology 24: 919–930. 10.1016/j.cub.2014.03.011 [DOI] [PubMed] [Google Scholar]
- 8. Winter M, Devictor V, Schweiger O (2013) Phylogenetic diversity and nature conservation: where are we? Trends in Ecology & Evolution 28: 199–204. 10.1016/j.tree.2012.10.015 [DOI] [PubMed] [Google Scholar]
- 9. Mooers AØ, Heard SB, Chrostowski E (2005) Evolutionary heritage as a metric for conservation. In: Purvis A, Gittleman JL, Thomas B, editors, Phylogeny and Conservation, Cambridge University Press; pp. 120–138. [Google Scholar]
- 10. Mace GM, Collar NJ, Gaston KJ, Hilton-Taylor C, Akçakaya HR, Leader-Williams N, et al. (2008) Quantification of extinction risk: IUCN’s system for classifying threatened species. Conservation Biology 22: 1424–1442. 10.1111/j.1523-1739.2008.01044.x [DOI] [PubMed] [Google Scholar]
- 11. Fay MF, Swensen SM, Chase MW (1997) Taxonomic a nities of Medusagyne oppositifolia (Medusagynaceae). Kew Bulletin 52: 111–120. 10.2307/4117844 [DOI] [Google Scholar]
- 12. Fay MF, Bayer C, Alverson WS, de Bruijn AY, Chase MW (1998) Plastid rbcL sequence data indicate a close a nity between Diegodendron and Bixa . Taxon 47: 43–50. 10.2307/1224017 [DOI] [Google Scholar]
- 13. Kores P, Cameron K, Molvray M, Chase M (1997) The phylogenetic relationships of Orchidoideae and Spiranthoideae (Orchidaceae) as inferred from rbcL plastid sequences. Lindleyana 12: 1–11. [Google Scholar]
- 14. Lledó MD, Crespo MB, Cameron KM, Fay MF, Chase MW (1998) Systematics of Plumbaginaceae based upon cladistic analysis of rbcL sequence data. Systematic Botany 23: 21–29. 10.2307/2419571 [DOI] [Google Scholar]
- 15. Benson DA, Cavanaugh M, Clark K, Karsch-Mizrachi I, Lipman DJ, Ostell J, et al. (2013) Genbank. Nucleic Acids Research 41: D36–D42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Pearse WD, Purvis A (2013) phyloGenerator: an automated phylogeny generation tool for ecologists. Methods in Ecology and Evolution 4: 692–698. 10.1111/2041-210X.12055 [DOI] [Google Scholar]
- 17. Hill MO, Preston CD, Roy DB (2004) PLANTATT—attributes of British and Irish plants: status, size, life history, geography and habitats. Centre for Ecology and Hydrology. [Google Scholar]
- 18. APG III (2009) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161: 105–121. 10.1111/j.1095-8339.2009.00996.x [DOI] [Google Scholar]
- 19. Webb CO, Donoghue MJ (2005) Phylomatic: tree assembly for applied phylogenetics. Molecular Ecology Notes 5: 181–183. 10.1111/j.1471-8286.2004.00829.x [DOI] [Google Scholar]
- 20. Katoh K, Kuma Ki, Toh H, Miyata T (2005) MAFFT version 5: improvement in accuracy of multiple sequence alignment. Nucleic Acids Research 33: 511–518. 10.1093/nar/gki198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Katoh K, Toh H (2008) Recent developments in the MAFFT multiple sequence alignment program. Briefings in Bioinformatics 9: 286–298. 10.1093/bib/bbn013 [DOI] [PubMed] [Google Scholar]
- 22. Stamatakis A (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688–2690. 10.1093/bioinformatics/btl446 [DOI] [PubMed] [Google Scholar]
- 23. Britton T, Anderson CL, Jacquet D, Lundqvist S, Bremer K (2007) Estimating divergence times in large phylogenetic trees. Systematic Biology 56: 741–752. 10.1080/10635150701613783 [DOI] [PubMed] [Google Scholar]
- 24. Kuhn TS, Mooers AØ, Thomas GH (2011) A simple polytomy resolver for dated phylogenies. Methods in Ecology and Evolution 2: 427–436. 10.1111/j.2041-210X.2011.00103.x [DOI] [Google Scholar]
- 25.Orme D, Freckleton R, Thomas G, Petzoldt T, Fritz S, Isaac N, et al. (2013) caper: comparative analyses of phylogenetics and evolution in R. URL http://CRAN.R-project.org/package=caper. R package version 0.5.2.
- 26. Telfer MG, Preston CD, Rothery P (2002) A general method for measuring relative change in range size from biological atlas data. Biological Conservation 107: 99–109. 10.1016/S0006-3207(02)00050-2 [DOI] [Google Scholar]
- 27. Powney GD, Rapacciuolo G, Preston CD, Purvis A, Roy DB (2014) A phylogenetically-informed trait-based analysis of range change in the vascular plant flora of Britain. Biodiversity and Conservation 23: 171–185. 10.1007/s10531-013-0590-5 [DOI] [Google Scholar]
- 28. Preston CD, Pearman DA, Dines TD (2002) New atlas of the British and Irish flora. Oxford University Press. [Google Scholar]
- 29. Perring FH, Walters SM (1962) Atlas of the British flora. T. Nelson and Sons. [Google Scholar]
- 30.Cheffings C, Farrell L (2005) The vascular plant Red Data List for Great Britain. Technical report, Joint Nature Conservation Committee.
- 31. Araújo MB, Peterson AT (2012) Uses and misuses of bioclimatic envelope modeling. Ecology 93: 1527–1539. 10.1890/11-1930.1 [DOI] [PubMed] [Google Scholar]
- 32.Ridgeway G (2007) gbm: Generalized Boosted Regression Models. URL http://CRAN.R-project.org/package=gbm. R package version 1.6–3.
- 33. Rapacciuolo G, Roy DB, Gillings S, Fox R, Walker K, Purvis A. (2012) Climatic associations of british species distributions show good transferability in time but low predictive accuracy for range change. PLoS ONE 7: e40212 10.1371/journal.pone.0040212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Southall H, Aucott P (2009). A vision of britain through time. URL http://visionofbritain.org.uk.
- 35.Bossard M, Feranec J, Otahel J (2000) Corine land cover technical guide. Technical Report 40, European Environment Agency.
- 36. Kharouba HM, Algar AC, Kerr JT (2009) Historically calibrated predictions of butterfly species’ range shift using global change as a pseudo-experiment. Ecology 90: 2213–2222. 10.1890/08-1304.1 [DOI] [PubMed] [Google Scholar]
- 37. Dobrowski SZ, Thorne JH, Greenberg JA, Safford HD, Mynsberge AR, Thorne JH, et al. (2011) Modeling plant ranges over 75 years of climate change in California, USA: temporal transferability and species traits. Ecological Monographs 81: 241–257. 10.1890/10-1325.1 [DOI] [Google Scholar]
- 38. Rapacciuolo G, Roy DB, Gillings S, Purvis A (2014) Temporal validation plots: quantifying how well correlative species distribution models predict species’ range changes over time. Methods in Ecology and Evolution 5: 407–420. 10.1111/2041-210X.12181 [DOI] [Google Scholar]
- 39. Phillips SJ, Elith J (2010) POC plots: calibrating species distribution models with presence-only data. Ecology 91: 2476–2484. 10.1890/09-0760.1 [DOI] [PubMed] [Google Scholar]
- 40. Rosindell J, Harmon LJ (2012) OneZoom: a fractal explorer for the tree of life. PLoS Biology 10: e1001406 10.1371/journal.pbio.1001406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Kissling WD, Dormann CF, Groeneveld J, Hickler T, Kühn I, McIneryny GJ, et al. (2012) Towards novel approaches to modelling biotic interactions in multispecies assemblages at large spatial extents. Journal of Biogeography 39: 2163–2178. 10.1111/j.1365-2699.2011.02663.x [DOI] [Google Scholar]
- 42. Sanderson MJ, McMahon MM, Steel M (2011) Terraces in phylogenetic tree space. Science 333: 448–450. 10.1126/science.1206357 [DOI] [PubMed] [Google Scholar]
- 43. Aberer AJ, Krompass D, Stamatakis A (2013) Pruning rogue taxa improves phylogenetic accuracy: an efficient algorithm and webservice. Systematic Biology 1: 162–166. 10.1093/sysbio/sys078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Hartmann K, André J (2013) Should evolutionary history guide conservation? Biodiversity and Conservation 22: 449–458. 10.1007/s10531-012-0422-z [DOI] [Google Scholar]
- 45. Collen B, Turvey ST, Waterman C, Meredith HMR, Kuhn TS, Baillie JEM, et al. (2011) In vesting in evolutionary history: implementing a phylogenetic approach for mammal conservation. Philosophical Transactions of the Royal Society B: Biological Sciences 366: 2611–2622. 10.1098/rstb.2011.0109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Braithwaite ME, Ellis RW, Preston CD (2006) Change in the British flora 1987–2004. Botanical Society of the British Isles, London. [Google Scholar]
- 47.UK Biodiversity Reporting and Information Group (2007) Report on the Species and Habitat Review. Technical report, DEFRA.
- 48. Pilgrim ES, Crawley MJ, Dolphin K (2004) Patterns of rarity in the native british flora. Biological Conservation 120: 161–170. 10.1016/j.biocon.2004.02.008 [DOI] [Google Scholar]
- 49. Bottrill MC, Joseph LN, Carwardine J, Bode M, Cook C, Game ET, et al. (2008) Is conservation triage just smart decision making? Trends in Ecology & Evolution 23: 649–654. 10.1016/j.tree.2008.07.007 [DOI] [PubMed] [Google Scholar]
- 50. Possingham HP, Andelman SJ, Burgman MA, Medellın RA, Master LL, Keith DA. (2002) Limits to the use of threatened species lists. Trends in ecology & evolution 17: 503–507. 10.1016/S0169-5347(02)02614-9 [DOI] [Google Scholar]
- 51. Bennett JR, Elliott G, Mellish B, Joseph LN, Tulloch AIT, Probert WJM, et al. (2014) Balancing phylogenetic diversity and species numbers in conservation prioritization, using a case study of threatened species in new zealand. Biological Conservation 174: 47–54. 10.1016/j.biocon.2014.03.013 [DOI] [Google Scholar]
- 52. Safi K, Armour-Marshall K, Baillie JE, Isaac NJ (2013) Global patterns of evolutionary distinct and globally endangered amphibians and mammals. PloS one 8: e63582 10.1371/journal.pone.0063582 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. van Wilgen NJ, Richardson DM (2012) The roles of climate, phylogenetic relatedness, introduction effort, and reproductive traits in the establishment of non-native reptiles and amphibians. Conservation Biology 26: 267–277. 10.1111/j.1523-1739.2011.01804.x [DOI] [PubMed] [Google Scholar]
- 54. Strauss SY, Webb CO, Salamin N (2006) Exotic taxa less related to native species are more invasive. Proceedings of the National Academy of Sciences of the USA 103: 5841–5845. 10.1073/pnas.0508073103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Tingley R, Phillips BL, Shine R (2011) Establishment success of introduced amphibians increases in the presence of congeneric species. The American Naturalist 177: 382–388. 10.1086/658342 [DOI] [PubMed] [Google Scholar]
- 56. Wiens JJ, Ackerly DD, Allen AP, Anacker BL, Buckley LB, Cornell HV, et al. (2010) Niche conservatism as an emerging principle in ecology and conservation biology. Ecology Letters 13: 1310–1324. 10.1111/j.1461-0248.2010.01515.x [DOI] [PubMed] [Google Scholar]
- 57. Myers N (1993) Biodiversity and the precautionary principle. Ambio 22: 74–79. [Google Scholar]
- 58.Bivand R (2013) classInt: Choose univariate class intervals. URL http://CRAN.R-project.org/package=classInt. R package version 0.1–21.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Genera were cut into the best-scoring phylogeny as described in the text, but at each integer quantile of genus age, and the evolutionary distinctiveness scores of all species correlated with the scores when the genera were cut at the 1st quantile. In the figure, the correlation coefficients are plotted against quantile at which the cuts were made.
(PDF)
The sub-panels show EDGE (A), EDM (B), ED(AM)max (C), EDAM (D), and EDAM′ (E). Each map has a legend to the right hand side, splitting the values into twenty ‘Jenks’ quantiles (classIntervals function in the R package classInt [58]). The greatest four quantiles have been coloured differently, to emphasise the high-priority sites. Note that the distributions of all five measures have extremely long tails (as shown by the size of the quantiles in the legends).
(JPG)
- analysed_accession_numbers.doc—Accession numbers of sequences used to build phylogeny
- bootstrap_trees.zip—All bootstrap replicates of phylogeny
- metrics.csv—CSV file with species scores under all metrics
- novel_sequence_accession_numbers.doc—Accession numbers of sequences uploaded to GenBank
- OneZoom.html—Best phylogeny, viewable in OneZoom, embedded in a web page (HTML file)
- phylogeny.tre—Best dated phylogeny in Newick format
- uncorrected_sequences.fasta—Uncorrected sequences used in analysis
- uncorrect_vs_corrected_sequence_check.doc—Demonstration that corrections made to sequences do not affect the results of the manuscript
(ZIP)
Data Availability Statement
All relevant data, including Genbank Accession numbers, are in the paper and its Supporting Information files.