

Abstract
It is well established that the organ damage that complicates human diabetes is caused by prolonged hyperglycemia, but the cellular and molecular mechanisms by which high levels of glucose cause tissue damage in humans are still not fully understood. The prevalent hypothesis explaining the mechanisms that may underlie the pathogenesis of diabetes complications includes overproduction of reactive oxygen species, increased flux through the polyol pathway, overactivity of the hexosamine pathway causing intracellular formation of advanced glycation end products, and activation of protein kinase C isoforms. In addition, experimental and clinical evidence reported in past decades supports a strong link between the complement system, complement regulatory proteins, and the pathogenesis of diabetes complications. In this article, we summarize the body of evidence that supports a role for the complement system and complement regulatory proteins in the pathogenesis of diabetic vascular complications, with specific emphasis on the role of the membrane attack complex (MAC) and of CD59, an extracellular cell membrane-anchored inhibitor of MAC formation that is inactivated by nonenzymatic glycation. We discuss a pathogenic model of human diabetic complications in which a combination of CD59 inactivation by glycation and hyperglycemia-induced complement activation increases MAC deposition, activates pathways of intracellular signaling, and induces the release of proinflammatory, prothrombotic cytokines and growth factors. Combined, complement-dependent and complement-independent mechanisms induced by high glucose promote inflammation, proliferation, and thrombosis as characteristically seen in the target organs of diabetes complications.
Introduction
The MAC: Formation and Function
The MAC as a Mediator of Cellular Signaling and an Effector of Organ Pathology
Complement Regulatory Proteins
-
Clinical Evidence for a Role of Complement in the Pathogenesis of Diabetes Complications
Diabetic nephropathy
Diabetic retinopathy
Diabetic neuropathy
Diabetic cardiovascular disease
-
Glycation-Inactivation of CD59: a Molecular Link Between Complement and the Complications of Diabetes
Human studies
Animal studies
-
Functional Evidence for Glycation-Inactivation of CD59 in Individuals With Diabetes, and Presence of Glycated CD59 in Target Organs of Diabetes Complications
Functional inactivation of CD59 in individuals with diabetes
Colocalization of GCD59 and MAC in target organs of diabetic complications
Glycated CD59 as a diabetes biomarker
Complement-targeted therapeutics
Conclusions
I. Introduction
Diabetes is reaching epidemic proportions worldwide; if it continues increasing at the current rate, diabetes will affect almost 10% of the world population by the year 2035. However, an epidemic of diabetes is in fact an epidemic of its complications; diabetes is associated with: 1) accelerated macrovascular disease resulting in atherosclerotic coronary heart disease, stroke, and peripheral artery disease; and 2) microvascular disease that damages the retina, leading to blindness; the kidneys, leading to end-stage renal failure; and peripheral nerves, leading to severe forms of neuropathy, which combined with peripheral artery disease are the leading cause of nontraumatic amputations. The cost of treating complications of diabetes exceeds 10% of the total healthcare expenditure worldwide.
Large-scale prospective studies for both type 1 and type 2 diabetes, including the Diabetes Control and Complications Trial (1, 2), the UK Prospective Diabetes Study (3), and the Steno-2 Study (4), established that the complications of diabetes are caused by prolonged hyperglycemia, and that the extent of tissue damage in individuals with diabetes is influenced by genetic determinants of susceptibility and by the presence of accelerating factors such as hypertension and dyslipidemia. A hypothesis summarizing different mechanisms that may underlie the pathogenesis of diabetes complications proposes that hyperglycemia-induced overproduction of reactive oxygen species (ROS) fuels an increased flux of sugars through the polyol pathway, an increased intracellular formation of advanced glycation end products (AGEs), an increase in reactive carbonyl compounds, increased expression of the receptor for AGEs and signaling upon binding to their activating ligands, the activation of protein kinase C (PKC) isoforms, and an overactivity of the hexosamine pathway (reviewed in Refs. 5–7). However, the actual cellular and molecular mechanisms by which high levels of glucose cause tissue damage in humans are still not fully understood.
A body of clinical and experimental evidence reported in past decades supports a link between the complement system, complement regulatory proteins, and the pathogenesis of diabetes complications (8–23). Emerging evidence also indicates that the complement system is involved in several features of cardiometabolic disease, including dysregulation of adipose tissue metabolism, low-grade focal inflammation, increased expression of adhesion molecules and proinflammatory cytokines in endothelial cells contributing to endothelial dysfunction, and insulin resistance (reviewed in Ref. 24). Here we will review the biology of complement with particular emphasis on the membrane attack complex (MAC) as a potential effector of pathology seen in target organs of diabetic complications, and of CD59, an extracellular cell membrane-anchored inhibitor of MAC formation that is inactivated by nonenzymatic glycation forming glycated CD59 (GCD59), an emerging biomarker for the diagnosis and clinical management of diabetes.
The complement system is an effector of both adaptive and innate immunity. It is composed of more than 30 plasma and cell-membrane proteins that are synthesized by hepatocytes or locally in peripheral tissues (25–28) and that normally circulate as inactive precursors (proproteins). Complement proteins interact with one another in three enzymatic activation cascades known as the classical, the alternative and the mannose-binding lectin (MBL) pathways (summarized in Figure 1 and reviewed in Refs. 29–31). The three activation pathways eventually converge at the level of C3 and C5; thereafter, the three complement pathways share a common reaction sequence through the late components C6, C7, C8, and C9, leading to the generation of the MAC, the main effector of complement-mediated tissue damage.
Figure 1.

The complement activation pathways and their regulators. The Complement System: The classical pathway is initiated by immune complexes that activate C1. Activated C1 then recruits C4 and C2 to form a C3-convertase (C4b2a) that fragments C3 into C3a, an anaphylatoxin, and C3b, a molecule at the core of the complement system that binds covalently to hydroxyl groups on carbohydrates or proteins in bacterial surfaces and/or cell membranes. C3b tags invading microorganisms for opsonization and also amplifies the activation cascade acting as a focal site for further complement activation. The phylogenetically older alternative pathway is activated by low-grade cleavage of C3 in plasma; the resulting C3b binds factor B, a protein homologous to C2, to form the C3bB complex. Bound to C3b, factor B is cleaved by the activator factor D, which circulates in an active form as a serine esterase to form C3bBb, the C3-convertase of the alternative pathway. Because basal “tick over” deposition of C3b occurs in all cells exposed to complement, C3b is always available to prime the alternative pathway. However, continued activation and amplification are normally restricted because cell membranes do not bind factor B efficiently, and the inhibitory factor H prevents the association of factor B with C3b. Amplification of the alternative pathway occurs when factor H binds to foreign carbohydrates leaving factor B free to complex with C3b. Specificity for activation of the alternative pathway resides in the ability of factor H to discriminate between “self” and foreign carbohydrate determinants (30, 31, 67). The MBL pathway is initiated when the plasma MBL protein, structurally similar to C1q (165), in a complex with the MASP1 and MASP2 binds to an array of mannose groups on the surface of microorganisms. This binding activates MASPs, which then cleave C2 and C4 leading to the formation of the C3-convertase C4b2a in a manner comparable to activated C1 (30). Terminal complement pathway: The three complement activation pathways eventually converge at the level of C3 with formation of C3b and a C5 convertase that cleaves C5 into C5a, an anaphylatoxin, and C5b. This is the last enzymatic step of the activation cascades; thereafter, the complement pathways share a common sequence through the terminal components C6, C7, C8, and C9, leading to the generation of the MAC, the main effector of complement-mediated tissue damage. The terminal complement components—C6 through C9—necessary to form the MAC are constitutively present in plasma. Formation of the MAC is initiated by C5b, followed by the sequential association of C6 into C5b6, and then C7, C8, and C9. C5b6 is a stable protein that binds to C7 to form the C5b-7 complex that inserts into the lipid bilayer of the plasma membrane. Membrane-bound C5b67 exposes a binding site for C8 leading to formation of the C5b-8 complex, which functions as a docking site for C9. A single C9 first binds to C8 in the C5b-8 complex; then C9 polymerizes, forming the C5b-9 complex known as the MAC. The MAC is a circular polymer of a variable number of C9 monomers (166–170) with the capacity to insert into cell membranes (171) and form a transmembrane pore with hydrophobic domains on the outside and hydrophilic domains in the inside and an effective internal radius of 5–7 nm (172–174). Restriction of complement activity by complement regulators: In the fluid phase, the C1 inhibitor regulates the classical pathway by targeting C1, factors H and I regulate the alternative pathway, and S-protein, clusterin, and serum lipids compete with membrane lipids for reacting with nascent C5b67 (29). In addition to these fluid phase inhibitors, several membrane proteins, DAF (175), membrane cofactor (MCP) (176, 177), and complement receptors 1 and 2 (CR1, CR2) regulate the C3-convertases (178), a major amplification step in the early complement activation cascade, whereas clusterin (179), S-protein (180), and CD59 inhibit formation of the MAC (115, 116, 181–185). Massive or unrestricted activation of complement and MAC formation leads to cell lysis. In the context of reduced restriction, just basal (tick over) complement activity is sufficient to form transient MAC pores in nucleated cells leading to influx and efflux of ions and macromolecules, and activation of cellular pathways that contribute to proliferation, inflammation and thrombosis.
II. The MAC: Formation and Function
As described in Figure 1, the MAC is a transmembrane pore that allows influx of salt and water leading to colloid osmotic lysis of MAC-targeted cells. The formation of the MAC pore, however, seems to be the end stage of a process that transits through a reversible phase during which MAC pores can be formed transiently (32–34), generating significant changes in the membrane permeability and internal composition of MAC-targeted cells without compromising their viability (32, 35, 36). Figure 2 illustrates the large increase of intracellular Ca++ levels caused by transient, nonlytic MAC pores on isolated guinea-pig cardiomyocytes (36). Thus, whereas the matured MAC pore can kill nonself cells by perforating their plasma membrane, the transient MAC pore can induce nonlethal biological responses in “self” cells and mediate physiological and/or pathological response (37).
Figure 2.

Nonlytic transient MAC triggers a large increase of intracellular Ca++ concentration ([Ca++]i). The figure shows pseudocolor images of isolated adult rat ventricular muscle cell loaded with fura-2, a fluorescent calcium-indicator dye. Increasing emission intensities were associated with increasing [Ca++]i, coded in a pseudocolor spectrum from blue (low Ca++) to red (high Ca++). A, Reference, digitalized phase-contrast image. Before adding complement factors, the cell shows low-emission intensity (yellow-green) (B) that does not significantly change after the addition of C5b6 plus C7 and C8 (C). After addition of C9, the cell showed a marked emission increase (D and E), representing an increase in [Ca++]i that lasted approximately 1 minute. After an additional 3 minutes, [Ca++]i decreased to near baseline levels (F–H). [Reproduced from H.J. Berger et al. Activated complement directly modifies the performance of isolated heart muscle cells from guinea pig and rat. Am J Physiol. 1993;265:H267–H272 (36). J.A. Halperin was an author, no permission needed by The American Journal of Physiology to reproduce.]
III. The MAC as a Mediator of Cellular Signaling and an Effector of Organ Pathology
Activation products of complement such as C3b and C4b or the anaphylatoxins C3a and C5a exert their biological activities through binding to specific receptors expressed in many cell types. Upon binding to their cognate ligands, these receptors trigger signaling processes mostly linked to inflammation and immune responses (38). In contrast, there are no specific receptors for the MAC, which inserts directly into the lipid bilayer of the plasma membrane. Insertion of transient nonlethal MAC pores into cell membranes induces an array of biological responses that promote cell proliferation, inflammation, and thrombosis, as characteristically seen in all diseases in which focal complement activation is a characteristic pathological abnormality. These responses are mediated by growth factors and cytokines including basic fibroblast growth factor and IL-1 (33, 39), which are directly released through the MAC pore, and platelet-derived growth factor (39), monocyte chemotactic protein-1 (MCP-1) (40), and von Willebrand factor (41), which are released through canonical secretion pathways in response to intracellular perturbations induced by the MAC. Acting auto- and paracrinically on extracellular receptors, these leaked/secreted molecules promote: 1) proliferation of fibroblasts, endothelial and smooth muscle cells (39); 2) inflammation through the expression of proinflammatory adhesion molecules such as P-selectin, vascular cell adhesion molecule 1, and E-selectin (42, 43), and the attraction of monocytes and macrophages to the site of focal complement activation (44); and 3) thrombosis through the expression of prothrombotic tissue factor and the transient assembly of the prothrombinase complex in both the endothelial cell surface and microvesicles shed from their plasma membrane (45–47). Our group has also shown that the MAC promotes in vitro accumulation of cholesteryl esters into mouse macrophages, fostering their conversion into foam cells (48).
Reversible insertion of transient MAC into cell membranes also activates pathways of intracellular signaling (reviewed in Ref. 49), including increased: 1) Ca++ influx (36, 50) and Ca++-activated K+ efflux (35); 2) production of ROS (51, 52); 3) activation of Ca++-sensitive and Ca++-insensitive PKC (53, 54); 4) activation of heterotrimeric G proteins of the Gi/Go subfamily (55); 5) mitotic signaling through the small G protein Ras, which induces Raf-1 translocation triggering activation of the ERK pathway (56); and 6) activation of phosphatidylinositol-3 kinase (57). The MAC also activates the transcription factor nuclear factor-κB (NF-κB), which induces the production of IL-6 as well as IL-8 and MCP-1 (58, 59). In glomerular mesangial cells, insertion of the MAC results in increased production of ROS (51) and the release and/or up-regulation of: 1) basic fibroblast growth factor and platelet-derived growth factor (39), which induce mesangial cell proliferation and transcriptional up-regulation of TGF-β (60, 61); 2) MCP-1, which attracts monocytes and macrophages to the mesangium (62); and 3) transcriptional activation of collagen type IV (63).
In the context of this review, it is important to note that many mediators and cellular pathways activated by the MAC are established players in the tissue damage seen in the target organs of diabetic complications (64, 65) and are common to or intersect with complement-independent mechanisms up-regulated by hyperglycemia, such as overproduction of ROS and up-regulation of PKC and NF-κB (reviewed in Refs. 5–7).
IV. Complement Regulatory Proteins
Given the potentially devastating effects of uncontrolled complement activation, it is not surprising that an array of mechanisms have evolved to protect “self” cells from complement attack and MAC formation. On the one hand, the inherent instability of the enzymes in the activation pathways and the extremely short half-life of the activated complement components coupled to the transient nature of the reaction sequences are intrinsic properties of the complement system that keep its activation restricted and limit the effects of complement to the vicinity of the activation site. On the other hand, a multitude of complement regulatory molecules exist that restrict critical steps of the pathways (Figure 1).
In particular, CD59 is a specific inhibitor of MAC formation that restricts C9 polymerization. Ubiquitously expressed in mammalian cells, CD59 is an 18- to 20-kDa glycan phosphatidylinositol (GPI)-linked glycoprotein (66, 67). CD59 inhibits the binding of C9 to C5b-8 by competing for binding to a nascent epitope on C8, thereby affecting the association of C9 to the C5b-8 complex (68). CD59 can not only function as an inhibitor of the formation of large MACs but also allow cells to eliminate newly formed MACs by blocking the early, functional channel formation of MAC complexes (69).
Several lines of evidence indicate that CD59 is the most critical of the complement regulators in protecting human cells from MAC formation and MAC-induced phenomena: erythrocytes from a family carrying the Inab blood group phenotype totally lacked decay-accelerating factor (DAF), an inhibitor of early complement activation, without any association with hemolytic disease (70–73); in contrast, individuals carrying CD59 mutations leading to undetectable levels of CD59 on their cell membranes exhibited an early-onset hemolytic phenotype associated with vascular disease or chronic relapsing polyneuropathy (74, 75).
The GPI anchor is attached to the proteins as a post-translational modification mediated by a protein called Pig-A that is required for the synthesis of the GPI-lipid tail (76–79). Deficiency of GPI-linked proteins including CD59 in erythrocytes and platelets is phenotypically expressed as a complement-mediated hemolytic anemia, known as paroxysmal nocturnal hemoglobinuria (PNH) (80, 81). This paradigmatic disease, together with the silencing CD59 mutations described above, illustrates how the delicate balance between complement activation and restriction can be broken by just the functional deficiency of CD59, leading to a potentially harmful pathological response even in the absence of complement-activating stressors.
V. Clinical Evidence for a Role of Complement in the Pathogenesis of Diabetes Complications
Evidence for a potential role of complement in the pathogenesis of diabetes complications was initially provided by the identification of activated complement proteins in target organs of diabetes complications biopsied or excised (9, 10, 82–84) from individuals with diabetes.
A. Diabetic nephropathy
Falk et al (82–84) used antibodies against activated complement proteins including a neoantigen of the C9 portion of the MAC to perform immunoelectron and immunofluorescent microscopy on kidney tissue from normal humans and individuals with insulin-dependent diabetes. Staining of tissues with specific antibodies directed against the MAC neoantigen is the most direct method for determining whether complement activation has occurred to completion because the neoantigen is exposed when the MAC is assembled and inserted into a cell membrane. Immunofluorescent microscopy of kidney tissue from individuals with insulin-dependent diabetes and varying degrees of mesangial expansion and glomerulosclerosis demonstrated a positive association between the degree of tissue damage and the amount of MAC deposited in the mesangium. Immunoelectron microscopy of specimens from individuals with diabetic nephropathy revealed that the anti-MAC neoantigen monoclonal antibody reacted with linear and circular membranous structures within the mesangium, tubular basement membranes, blood vessel walls, and the glomerular basement membranes (82–84). Our own studies confirmed glomerular MAC deposition (colocalized with GCD59, Section VII.B) in kidney biopsies from individuals with diabetes (19). We also reported the presence of extensive MAC deposition in glomeruli and blood vessels in the biopsy of a kidney transplanted 2 years earlier and diagnosed as “recurrent diabetic nephropathy with no signs of rejection” (19).
Also, several studies have found that serum concentrations of MBL, an indicator of the lectin complement pathway, were significantly elevated in patients with type 1 diabetes (85, 86) and even more elevated in patients with vascular complications including diabetic nephropathy (13, 15, 87). Furthermore, the FinnDiane Study showed that the baseline levels of MBL were positively associated with the progression and prognosis of diabetic nephropathy in type 1 diabetes (16, 88). Transcriptome analyses of human kidneys have shown that the canonical complement-signaling pathway is differentially up-regulated in both glomeruli and tubules of individuals with diabetic nephropathy and is associated with increased glomerulosclerosis. Woroniecka et al (18) showed that increased expression of C3 mRNA and protein in glomeruli was positively correlated with diabetic kidney disease. C3 is a key protein central to all complement activation pathways.
B. Diabetic retinopathy
A combination of biochemical, histological, and genetic studies suggest that aberrant function of the complement system plays a key role in the etiology of age-related macular degeneration, a common degenerative ocular disease (22). Two independent studies have documented that complement activation occurred extensively and to completion in the choriocapillaries of eyes from donors with diabetic retinopathy, whereas similar deposits were not found in eyes from donors without retinopathy. In the study by Gerl et al (23), intense positive staining for MAC deposits (neoantigens) and C3d was noted without exception in all 50 eyes with diabetic retinopathy analyzed, but in only one of 26 eye donors without diabetic retinopathy (10, 23).
C. Diabetic neuropathy
Nerve damage through complement activation has been reported in several peripheral neuropathies (89). Interestingly, a recent study showed that a cell surface deficiency of the complement regulatory protein CD59 due to a single missense mutation of the CD59 gene was clinically expressed by a form of chronic peripheral neuropathy in children (75). The presence of activated complement proteins and MAC neoantigen in sural nerve biopsies from individuals with diabetes was first reported by Hays and collaborators (9); in the same nerve biopsies analyzed in that study, we observed colocalization of MAC and GCD59 (19).
D. Diabetic cardiovascular disease
A relatively large body of evidence supports a role of complement in cardiovascular pathology in general and atherosclerosis in particular (reviewed in Refs. 24, 44, 48, 90–97). Diabetes and prediabetes are major independent risk factors for cardiovascular disease (98, 99), and a potential role of C3, its activation products, and other components of the early complement cascade in the pathogenesis of type 2 diabetes has been suggested from associations found in several cross-sectional and longitudinal human studies, as recently reviewed in Ref. 24. Also, cleavage of C3 generates the anaphylatoxins, C3a and C5a, and further cleavage of C3a by carboxypeptidases leads to production of acylation stimulating protein (ASP/C3adesArg), a lipogenic hormone that plays a role in atherosclerosis (100–102). However, the potential role of complement activation in diabetic cardiovascular disease has not been fully explored in human studies. In a subanalysis of the DIGAMI-2 trial, which prospectively studied more than 1200 individuals with type 2 diabetes who had a myocardial infarction, high blood levels of soluble MAC at the time of hospitalization strongly and independently predicted the risk of a cardiovascular event in the subsequent 3.5 years (12). In the same subanalysis, levels of MBL-associated protease (MASP)-2 appeared to be associated with a poor cardiovascular prognosis, although this did not remain significant after multivariable adjustments.
Together, the findings summarized above suggest an important role of the complement system in the development and progression of diabetic complications. However, it is important to note that whether the reported associations of elements of the complement system with diabetes represent causative pathogenic precursors or are just a collateral consequence of diabetes and its complications is still unclear and deserves further investigation.
Complementary to the clinical evidence discussed above, in vitro and animal studies seem to suggest that the lectin pathway could be activated by increased levels of MBL induced by hyperglycemia and by a cooperative binding of MBL with fructoselysine, the product of the nonenzymtic attachment of glucose to selective α- or ϵ-amino groups in proteins (103, 104).
VI. Glycation-Inactivation of CD59: a Molecular Link Between Complement and the Complications of Diabetes
A. Human studies
There are different ways in which the “delicate balance” between complement activation and restriction can be potentially broken in diabetes. On the one hand, autoantibodies to glycated and glycol-oxidized proteins can activate the classical pathway (17, 105, 106), fructosamines reportedly activate the lectin pathway by an increased ligand-receptor binding (103), and increased AGEs on the arterial walls can serve as neo-epitopes for MBL binding (11). The presence of activated products of C3 (C3d and iC3b), C2, and C4 in target organs of diabetic complications (9, 23) is consistent with activation of complement through either or both of these two pathways. On the other hand, functional inhibition by glycation or reduced expression of complement regulatory proteins such as CD59 (8, 10) could lead to the increase of complement-mediated damage and MAC-induced biological processes likely to contribute to the pathogenesis of diabetic complications (9, 10, 82, 84, 107).
Of all the above-mentioned putative mechanisms that potentially contribute to complement-mediated tissue damage in diabetes, glycation-inactivation of CD59 is the most extensively studied and documented. Hereafter, we will summarize the experimental and clinical evidence for a role of GCD59 in the pathophysiology of diabetes complications.
Protein glycation on α- or ϵ-amino groups is a well-established mechanism of tissue damage in diabetes. Despite the many amino groups exposed on the surface of either soluble or membrane proteins, very few ϵ-amino groups are glycated to a significant extent. For example, despite the very close proximity of lysyl residues K61, K65, and K66 in β-hemoglobin, only the ϵ-amino group of K66 is preferentially glycated in vivo (108). This is because significant ϵ-amino glycation requires effective acid-base catalysis, which occurs at “glycation motives” such as histidyl/lysyl residues in which the imidazolyl moiety is located at ≈ 5A° from the ϵ-amino group of the lysyl residue or at lysyl-lysyl pairs (108–113).
Analyzing the nuclear magnetic resonance structure of human CD59 reported by Fletcher et al (114), our group predicted that the protein contains within its active site a glycation motif formed by amino acid residues K41 and H44 and that glycation could inhibit the activity of CD59 (Figure 3A). This idea was further supported by the fact that K41 is adjacent to W40, a conserved residue that is essential for CD59 function (115, 116). Exposing purified or recombinant human CD59 to glucose, we demonstrated that the anti-MAC activity of CD59 is indeed inhibited by glycation, as documented with antiglycated CD59 antibodies (Figure 3B) (8). Site-directed mutagenesis of human CD59 confirmed experimentally that: 1) human CD59 is inhibited by glycation of its K41 residue; and 2) H44 is required for glycation-inactivation of human CD59 (8) (Figure 3B).
Figure 3.
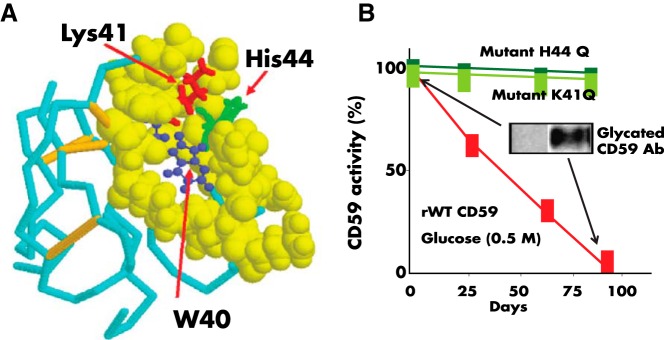
A, Nuclear magnetic resonance structure of human CD59 (from Ref. 114). The figure highlights the Lys41 (red) and His44 (green) amino acid residues that conform the glycation motif and the W40 (blue) residue, a conserved amino acid critical for CD59 activity. The His44 of human CD59 is not found in CD59 from any other species sequenced to date. B, Glycation inactivates human CD59: the activity of recombinant human CD59 was measured in a hemolytic assay using guinea-pig erythrocytes exposed to human MAC formed with purified terminal complement components, as reported in Ref. 8. Incubation of CD59 in the presence of glucose progressively inhibits CD59 activity (red line). Inhibition of activity is associated with glycation of CD59 because: 1) parallel incubation of the same preparation with the nonglycating sugar sorbitol did not affect CD59 activity (see Figure 2 in Ref. 8), and an antibody specific for the glycated form of CD59 showed the presence of glycated CD59 in the protein inactivated after incubation with glucose (inset). Site-directed mutagenesis of either K41 or H44 generated two CD59 mutants (mutant K41Q and mutant H44Q) that were active against human MAC but not inhibited by exposure to glucose.
B. Animal studies
The significance of the glycation motif in human CD59 is highlighted by the fact that the residue H44 is absent in CD59 from any animal species sequenced to date (Table 1 in Ref. 8). This is remarkable because humans are particularly prone to develop diabetic vascular complications, and it is well known that no single animal model of the disease can recapitulate the extensive vascular disease that commonly complicates human diabetes (117). This observation has led to the intriguing hypothesis that the presence of a glycation motif K41-H44 in human CD59 (not present in CD59 from other species) could contribute to the unique sensitivity of humans to develop extensive vascular disease in response to hyperglycemia. To test whether the functional inactivation of CD59 confers higher risk for diabetes-induced atherosclerosis, our group has generated molecular-engineered mice that are completely deficient in CD59 (mCD59KO mice), crossed them into the ApoE−/− background (one of the models recommended by the Diabetes Complications Consortium for studying diabetes-induced atherosclerosis) (118), and rendered them diabetic by injection of low doses of streptozotocin (an experimental model of type 1 diabetes also recommended by the Diabetes Complications Consortium) (119). The ablation of CD59 (mCD59 knockout mice in an ApoE−/− background) promoted accelerated atherosclerosis with occlusive coronary disease, vulnerable plaque, and premature death (48, 120). In contrast, either transgenic overexpression of human CD59 in the endothelium or the administration of a neutralizing anti-C5 monoclonal antibody in mice significantly attenuated the development of atherosclerosis (67). These observations in molecular-engineered mice are consistent with a body of experimental evidence supporting a role of complement in the development of atherosclerosis (90, 91, 94, 95, 120, 121).
Initial evidence demonstrated that diabetic mCd59KO/Apoe−/− mice developed a severe and accelerated form of atherosclerosis characterized by significantly larger atherosclerotic lesion areas in the aorta (>2-fold increased lesion area in en-face aorta preparations) and the aortic roots, as compared to their nondiabetic littermates. This accelerated atherosclerosis observed in diabetic CD59-deficient mice was associated with significantly increased MAC deposition (detected with antimouse C9 antibodies) and occurred at levels of hyperglycemia, lipid profiles, and body weights similar to those in diabetic CD59-expressing mice (122). Interestingly, diabetic mCD59KO mice had significantly increased MAC deposition and higher levels of serum iC3b, a byproduct of C3 cleavage during activation of complement, than nondiabetic mCD59KO mice. These two observations are consistent with the emerging evidence for activation of the complement system in diabetes (11, 17, 103, 105), as discussed above.
VII. Functional Evidence for Glycation-Inactivation of CD59 in Individuals With Diabetes, and Presence of Glycated CD59 in Target Organs of Diabetes Complications
The identification of the cellular and molecular mechanisms by which hyperglycemia causes tissue damage in humans has been hampered by: 1) the lack of animal models that recapitulate the combination and intensity of complications seen in human diabetes (117); 2) the chronic nature of the complications that can take decades to become clinically evident; and 3) ethical and practical considerations that preclude conduction of the functional studies required to assess mechanistic hypotheses in humans. Despite these limitations, two lines of human experimental evidence seem to support the potential role of glycation-inactivation of CD59 in the pathogenesis of diabetes complications.
A. Functional inactivation of CD59 in individuals with diabetes
If glycation inactivates CD59 in vivo, one would predict an increased sensitivity to MAC-mediated phenomena in cells/tissues carrying glycation-inactivated CD59. This prediction was confirmed in an ex vivo study using red blood cells (RBCs) from donors with or without diabetes, in which we measured both the activity of CD59 in the RBC membranes and the RBC sensitivity to human MAC. The results showed that RBCs from individuals with diabetes selected to have glycated hemoglobin (HbA1c > 8%) exhibited a significantly reduced activity of CD59 and were significantly more sensitive to MAC-mediated lysis than RBCs from individuals without diabetes (HbA1c < 5.5%) (19). This reduced activity of CD59 in RBCs from individuals with diabetes is consistent with inactivation of CD59 by glycation and would explain some reversible hematological abnormalities reportedly seen in individuals with poorly controlled diabetes, including increased reticulocyte count and shorter RBC life span (123–125), both suggestive of mild hemolytic anemia. Also, individuals with diabetes experience a high incidence of thrombotic episodes, their platelets are hyperactive, and their plasma contains high levels of platelet-derived microparticles (PMPs) (126). Thrombosis is also a major cause of morbidity and mortality in individuals with PNH (80, 127, 128), whose CD59-deficient platelets are exquisitely sensitive to MAC-induced activation and release of PMP (129); platelets from CD59 knockout mice are also activated spontaneously and release PMPs (130–132). These combined clinical and experimental observations indicate that inactivation of CD59 by glycation could be one factor contributing to the high incidence of thrombosis in individuals with diabetes.
B. Colocalization of GCD59 and MAC in target organs of diabetic complications
Using an antibody that specifically recognizes the glycated from of CD59 (GCD59), we were able to identify the presence of GCD59 in renal and sural nerve biopsies from individuals with diabetes. No detectable amounts of GCD59 were found in biopsies from individuals without diabetes. As expected from the functional inactivation of CD59 by glycation, serial sections of the same renal and nerve biopsies showed colocalization of GCD59 with MAC deposits (19). Increased levels of GCD59 and MAC have also been observed in other tissues extracted from individuals with diabetes, namely skin biopsies and saphenous veins removed for grafting of occluded arteries (J. A. Halperin et al, our unpublished observation).
C. Glycated CD59 as a diabetes biomarker
Although CD59 is a cell membrane-bound protein, a soluble form of CD59 that is shed from cell membranes by phospholipases is present in human blood, urine, saliva, and other bodily fluids (133–136). Ghosh et al (137) reported the development of a highly specific and sensitive ELISA assay to measure circulating levels of GCD59. In human association studies conducted to assess blood levels of GCD59 as a biomarker of diabetes using this ELISA assay (138), levels of GCD59 were 3- to 4-fold higher in individuals with type 2 diabetes. Furthermore, a positive and independent association between HbA1c and GCD59 was observed among 400 subjects (r = 0.58; P < .001); moreover, this association remained robust even in subgroup analyses of subjects with diabetes and those without diabetes, suggesting that this positive relationship was not confined to abnormal glucose handling alone. Beyond associations with HbA1c, in a cohort of subjects with no prior diagnosis of diabetes (n = 109), higher GCD59 concentrations were strongly associated with higher glucose levels after 2-hour oral glucose tolerance tests, independently of other known predictors of the 2-hour oral glucose tolerance test (β = 19.8; P = .02) (138). Lastly, the GCD59 level has also been shown to acutely parallel changes in glycemic control during therapeutic intervention with insulin. In an intervention study involving intensification of insulin therapy in persons with poorly controlled diabetes, reductions in average weekly glucose mediated by insulin treatment tightly paralleled the reductions in GCD59 levels. In contrast, other markers of glycemic control such as HbA1c and fructosamine did not change significantly in response to average weekly glucose reductions during those 2 weeks (138). Collectively, these human studies strongly implicate GCD59 as a biomarker that may be useful for the diagnosis of diabetes and in the chronic management of glycemic control in diabetic individuals.
Other complement proteins that have been reportedly used to study the pathways of complement activation and could be potentially explored as diabetes biomarkers include: C1q, C3, C4d (classical and lectin pathways), factors D and Bb (alternative pathway), 3a and C5a (general complement activation), soluble MAC complexes (terminal complement activation), and soluble forms of complement regulatory proteins DAF, MCP and CR1 (139).
D. Complement-targeted therapeutics
Progressively recognized as a major mediator of a variety of clinical disorders that include both acute and chronic diseases and affect a wide range of organs, modulation of the complement system with either small molecules or biologicals is now considered a promising strategy in drug discovery (reviewed in Refs. 140 and 141). Fueled by 1) a wealth of high-resolution structural models (142–147), functional assays (148–156), and genome-wide association studies (157), which together offer new insight into the molecular details and ligand interaction sites of complement components; 2) the approval and therapeutic success of the anti-C5 antibody, eculizumab (Alexion), for the treatment of PNH (158–160) and more recently of atypical hemolytic-uremic syndrome (161); and 3) the evidence that even small changes in the activity of individual complement proteins may add up to a significant effect over a disease progression, the field of complement-targeted drug discovery has seen a surge in approaches to therapeutically intervene at various stages of the complement cascades. Potential therapeutic target mechanisms include inhibition of C3 activation, inhibition of C3 convertase assembly, acceleration of C3 convertase decay, promotion of C3B proteolysis, inhibition of C3 and/or C5 convertase activities, inhibition of MAC assembly by CD59, and reestablishment of normal alternative complement pathway control with protective Factor H protein. Compounds directed against some of these targets that are currently in early clinical trials for other indications include: compstatin/POT-4, a peptide inhibitor that inhibits the central step of the complement cascade by preventing cleavage of C3; and ARC1905, a pegylated, aptamer-based C5 inhibitor that inhibits the cleavage of C5 into C5a and C5b. Other complement pathway-modulating compounds currently being considered for and/or under preclinical development include a humanized antifactor D antibody (TNX-234), a Fab fragment of an anti-CFB antibody (TA106), a CR2-factor H hybrid protein, a small molecule C5aR peptidomimetic (JPE-1375), and antiproperdin antibody, an inhibitor of the classical pathway (C1-INH), and a soluble form of complement receptor 1.
Based on the extensive clinical and experimental evidence supporting a role of complement in the pathogenesis of diabetes complications, it is conceivable that in the future, complement inhibition could offer an early point of intervention that could potentially retard, prevent, or even reverse progression of those complications. However, given the pleomorphic biological activities of the complement system, risk-benefit analyses of therapeutic application targeting complement to prevent or treat complications of diabetes will have to evaluate cautiously the potential systemic effects of chronic complement pathway modulation.
VIII. Conclusions
In this article we have summarized the body of evidence that supports a role for the complement system and complement regulatory proteins in the pathogenesis of diabetic vascular complications, with specific emphasis on the novel phenomenon of glycation-inactivation of CD59. Progress in this area has been remarkable considering that: 1) ethical and practical issues preclude conducting in-depth mechanistic studies in humans, especially for chronic conditions such as the complications of diabetes that take years to develop; and 2) nonclinical research is hampered due to the absence of animal models of diabetes that fully recapitulate diabetic vascular disease in the intensity and distribution seen in humans.
Similar to other tightly regulated biological systems, phenotypic endpoints derived from complement activation depend on the delicate balance between complement activators and inhibitors. In the specific case of complement and its regulators, this “delicate balance” paradigm is well exemplified by the human disease PNH, an acquired complement-mediated hemolytic anemia in which blood cells lack GPI-anchored proteins including CD59 (78, 81). Under “normal” conditions, the basal “tick-over” activation of complement is sufficient to induce only a mild degree of hemolysis. In contrast, when stressors like infections amplify complement activation, MAC attack on unprotected RBCs and platelets results in “paroxysmal” crises of hemolysis, platelet activation, and potentially lethal thrombotic episodes (128, 162, 163). The emerging clinical evidence summarized in this article indicates that comparable mechanisms could be operative in the pathogenesis of diabetes complications: on the one hand, biological responses to hyperglycemia like formation of AGEs and fructosamine adducts in proteins as well as autoantibodies against oxidized and/or glycated proteins could activate complement, whereas on the other hand, glycation-inactivation of CD59 increases the susceptibility of tissues to complement (MAC) mediated damage.
Based on the evidence summarized above, it is tempting to propose a pathogenic model of human diabetic complications in which the delicate balance between complement activation and restriction is broken by a combination of glycation-inactivation of CD59 and complement activation by autoantibodies, increased activity of the MBL pathway, and possibly other still uncovered mechanisms. This results in an increased MAC deposition as observed in target organs of diabetes complications. Increased MAC deposition activates pathways of intracellular signaling, including increased production of ROS, activation of PKC, and up-regulation of NF-κB and induces the release of proinflammatory, prothrombotic cytokines and growth factors. These complement-dependent mechanisms, together, perhaps synergistically, with the multiple complement-independent mechanisms summarized by Brownlee (5–7), promote inflammation, proliferation, and thrombosis as characteristically seen in the target organs of diabetes complications. Figure 4 summarizes these complement-dependent and complement-independent mechanisms, highlighting the points of convergence common to both mechanisms such as increased ROS and activation of PKC and transcription factor NF-κB.
Figure 4.

A model of the cellular and molecular mechanisms leading to cell proliferation, inflammation, and thrombosis characteristically seen in the target organs of diabetic complications. High levels of glucose in diabetes glycates and thereby inactivates CD59 and also increases complement activation. Combined, these effects of hyperglycemia on the complement system trigger more MAC deposition on cell membranes. The figure illustrates the different cellular signaling pathways induced by high glucose; the complement/MAC-dependent mechanisms summarized in this review are shown on the right, and the complement-independent mechanisms (reviewed in Refs. 5 and 7) are shown on the left. The center of the figure highlights the common signaling pathways reportedly triggered by either high glucose or the MAC. The evidence summarized in this review indicates that both mechanisms are operative in the pathogenesis of diabetes complications.
Future research in the area should comprise studies to elucidate: 1) the relative roles of these complement-related mechanisms in the pathogenesis of the different diabetes complications; 2) whether differences in complement activity or in expression of complement regulatory proteins such as CD59 could explain the increased risk of diabetes complications among ethnic minorities (164); and 3) how complement-induced biological phenomena and other hyperglycemia-induced cellular responses interplay in the pathogenesis of diabetes complications. Because the complement-related mechanisms described in this review are driven by hyperglycemia, it is conceivable that they will operate similarly in type 1 and type 2 diabetes, a notion to be tested in future human studies. Such research will aid in the development of novel strategies to stratify earlier individuals with diabetes who are at higher risk of complications, improve prevention of complications, and develop new complement-targeted therapeutic approaches for the complications of diabetes that represent a major cause of morbidity and mortality in adults.
Acknowledgments
This work was supported in part by National Institutes of Health Grants DK62294, DK089206, and DK101442 (all to J.A.H.) and by the National Heart, Lung, and Blood Institute of the National Institutes of Health under award no. K23HL111771 (to A.V.).
Disclosure Summary: J.A.H. and M.C. have a financial interest in Mellitus, LLC. Mellitus is developing diagnostic tools for diabetes. J.A.H. and M.C.'s interests were reviewed and are managed by Brigham and Women's Hospital and Partners HealthCare in accordance with their conflict of interest policies. The other authors have nothing to declare.
Footnotes
- AGE
- advanced glycation end product
- DAF
- decay-accelerating factor
- GPI
- glycan phosphatidylinositol
- HbA1c
- glycated hemoglobin
- MAC
- membrane attack complex
- MASP
- MBL-associated protease
- MBL
- mannose-binding lectin
- MCP-1
- monocyte chemotactic protein-1
- NF-κB
- nuclear factor-κB
- PKC
- protein kinase C
- PMP
- platelet-derived microparticle
- PNH
- paroxysmal nocturnal hemoglobinuria
- RBC
- red blood cell
- ROS
- reactive oxygen species.
References
- 1. Effect of intensive diabetes management on macrovascular events and risk factors in the Diabetes Control and Complications Trial. Am J Cardiol. 1995;75:894–903. [DOI] [PubMed] [Google Scholar]
- 2. Effect of intensive therapy on the development and progression of diabetic nephropathy in the Diabetes Control and Complications Trial. The Diabetes Control and Complications (DCCT) Research Group. Kidney Int. 1995;47:1703–1720. [DOI] [PubMed] [Google Scholar]
- 3. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33). UK Prospective Diabetes Study (UKPDS) Group. Lancet. 1998;352:837–853. [PubMed] [Google Scholar]
- 4. Pedersen O, Gaede P. Intensified multifactorial intervention and cardiovascular outcome in type 2 diabetes: the Steno-2 study. Metabolism. 2003;52:19–23. [DOI] [PubMed] [Google Scholar]
- 5. Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414:813–820. [DOI] [PubMed] [Google Scholar]
- 6. Giacco F, Brownlee M. Oxidative stress and diabetic complications. Circ Res. 2010;107:1058–1070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Brownlee M. The pathobiology of diabetic complications: a unifying mechanism. Diabetes. 2005;54:1615–1625. [DOI] [PubMed] [Google Scholar]
- 8. Acosta J, Hettinga J, Flückiger R, et al. Molecular basis for a link between complement and the vascular complications of diabetes. Proc Natl Acad Sci USA. 2000;97:5450–5455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Rosoklija GB, Dwork AJ, Younger DS, Karlikaya G, Latov N, Hays AP. Local activation of the complement system in endoneurial microvessels of diabetic neuropathy. Acta Neuropathol (Berl). 2000;99:55–62. [DOI] [PubMed] [Google Scholar]
- 10. Zhang J, Gerhardinger C, Lorenzi M. Early complement activation and decreased levels of glycosylphosphatidylinositol-anchored complement inhibitors in human and experimental diabetic retinopathy. Diabetes. 2002;51:3499–3504. [DOI] [PubMed] [Google Scholar]
- 11. Flyvbjerg A. Diabetic angiopathy, the complement system and the tumor necrosis factor superfamily. Nat Rev Endocrinol. 2010;6:94–101. [DOI] [PubMed] [Google Scholar]
- 12. Mellbin LG, Bjerre M, Thiel S, Hansen TK. Complement activation and prognosis in patients with type 2 diabetes and myocardial infarction: a report from the DIGAMI 2 trial. Diabetes Care. 2012;35:911–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Hansen TK, Tarnow L, Thiel S, et al. Association between mannose-binding lectin and vascular complications in type 1 diabetes. Diabetes. 2004;53:1570–1576. [DOI] [PubMed] [Google Scholar]
- 14. Hansen TK. Mannose-binding lectin (MBL) and vascular complications in diabetes. Horm Metab Res. 2005;37(suppl 1):95–98. [DOI] [PubMed] [Google Scholar]
- 15. Hansen TK, Forsblom C, Saraheimo M, et al. Association between mannose-binding lectin, high-sensitivity C-reactive protein and the progression of diabetic nephropathy in type 1 diabetes. Diabetologia. 2010;53:1517–1524. [DOI] [PubMed] [Google Scholar]
- 16. Hovind P, Hansen TK, Tarnow L, et al. Mannose-binding lectin as a predictor of microalbuminuria in type 1 diabetes: an inception cohort study. Diabetes. 2005;54:1523–1527. [DOI] [PubMed] [Google Scholar]
- 17. Uesugi N, Sakata N, Nangaku M, et al. Possible mechanism for medial smooth muscle cell injury in diabetic nephropathy: glycoxidation-mediated local complement activation. Am J Kidney Dis. 2004;44:224–238. [DOI] [PubMed] [Google Scholar]
- 18. Woroniecka KI, Park AS, Mohtat D, Thomas DB, Pullman JM, Susztak K. Transcriptome analysis of human diabetic kidney disease. Diabetes. 2011;60:2354–2369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Qin X, Goldfine A, Krumrei N, et al. Glycation inactivation of the complement regulatory protein CD59: a possible role in the pathogenesis of the vascular complications of human diabetes. Diabetes. 2004;53:2653–2661. [DOI] [PubMed] [Google Scholar]
- 20. Engström G, Hedblad B, Eriksson KF, Janzon L, Lindgärde F. Complement C3 is a risk factor for the development of diabetes: a population-based cohort study. Diabetes. 2005;54:570–575. [DOI] [PubMed] [Google Scholar]
- 21. Wlazlo N, van Greevenbroek MM, Ferreira I, et al. Complement factor 3 is associated with insulin resistance and with incident type 2 diabetes over a 7-year follow-up period: the CODAM study. Diabetes Care. 2014;37:1900–1909. [DOI] [PubMed] [Google Scholar]
- 22. Gehrs KM, Jackson JR, Brown EN, Allikmets R, Hageman GS. Complement, age-related macular degeneration and a vision of the future. Arch Ophthalmol. 2010;128:349–358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Gerl VB, Bohl J, Pitz S, Stoffelns B, Pfeiffer N, Bhakdi S. Extensive deposits of complement C3d and C5b-9 in the choriocapillaris of eyes of patients with diabetic retinopathy. Invest Ophthalmol Vis Sci. 2002;43:1104–1108. [PubMed] [Google Scholar]
- 24. Hertle E, Stehouwer CD, van Greevenbroek MM. The complement system in human cardiometabolic disease. Mol Immunol. 2014;61:135–148. [DOI] [PubMed] [Google Scholar]
- 25. Morgan BP, Gasque P. Extrahepatic complement biosynthesis: where, when and why? Clin Exp Immunol. 1997;107:1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Laufer J, Katz Y, Passwell JH. Extrahepatic synthesis of complement proteins in inflammation. Mol Immunol. 2001;38:221–229. [DOI] [PubMed] [Google Scholar]
- 27. Andoh A, Fujiyama Y, Sakumoto H, et al. Detection of complement C3 and factor B gene expression in normal colorectal mucosa, adenomas and carcinomas. Clin Exp Immunol. 1998;111:477–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Passwell JH, Schreiner GF, Wetsel RA, Colten HR. Complement gene expression in hepatic and extrahepatic tissues of NZB and NZB x W (F1) mouse strains. Immunology. 1990;71:290–294. [PMC free article] [PubMed] [Google Scholar]
- 29. Morgan BP, Harris CL. Complement Regulatory Proteins. London, UK: Academic Press; 1999. [Google Scholar]
- 30. Walport MJ. Complement. First of two parts. N Engl J Med. 2001;344:1058–1066. [DOI] [PubMed] [Google Scholar]
- 31. Walport MJ. Complement. Second of two parts. N Engl J Med. 2001;344:1140–1144. [DOI] [PubMed] [Google Scholar]
- 32. Halperin JA, Taratuska A, Rynkiewicz M, Nicholson-Weller A. Transient changes in erythrocyte membrane permeability are induced by sublytic amounts of the complement membrane attack complex (C5b-9). Blood. 1993;81:200–205. [PubMed] [Google Scholar]
- 33. Acosta JA, Benzaquen LR, Goldstein DJ, Tosteson MT, Halperin JA. The transient pore formed by homologous terminal complement complexes functions as a bidirectional route for the transport of autocrine and paracrine signals across human cell membranes. Mol Med. 1996;2:755–765. [PMC free article] [PubMed] [Google Scholar]
- 34. Halperin JA, Nicholson-Weller A, Brugnara C, Tosteson DC. Complement induces a transient increase in membrane permeability in unlysed erythrocytes. J Clin Invest. 1988;82:594–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Halperin JA, Brugnara C, Nicholson-Weller A. Ca2+-activated K+ efflux limits complement-mediated lysis of human erythrocytes. J Clin Invest. 1989;83:1466–1471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Berger HJ, Taratuska A, Smith TW, Halperin JA. Activated complement directly modifies the performance of isolated heart muscle cells from guinea pig and rat. Am J Physiol. 1993;265:H267–H272. [DOI] [PubMed] [Google Scholar]
- 37. Nicholson-Weller A, Halperin JA. Membrane signaling by complement C5b-9, the membrane attack complex. Immunol Res. 1993;12:244–257. [DOI] [PubMed] [Google Scholar]
- 38. Carroll MC. The role of complement and complement receptors in induction and regulation of immunity. Annu Rev Immunol. 1998;16:545–568. [DOI] [PubMed] [Google Scholar]
- 39. Benzaquen LR, Nicholson-Weller A, Halperin JA. Terminal complement proteins C5b-9 release basic fibroblast growth factor and platelet-derived growth factor from endothelial cells. J Exp Med. 1994;179:985–992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Torzewski J, Oldroyd R, Lachmann P, Fitzsimmons C, Proudfoot D, Bowyer D. Complement-induced release of monocyte chemotactic protein-1 from human smooth muscle cells. A possible initiating event in atherosclerotic lesion formation. Arterioscler Thromb Vasc Biol. 1996;16:673–677. [DOI] [PubMed] [Google Scholar]
- 41. Hattori R, Hamilton KK, McEver RP, Sims PJ. Complement proteins C5b-9 induce secretion of high molecular weight multimers of endothelial von Willebrand factor and translocation of granule membrane protein GMP-140 to the cell surface. J Biol Chem. 1989;264:9053–9060. [PubMed] [Google Scholar]
- 42. Marks RM, Todd RF, 3rd, Ward PA. Rapid induction of neutrophil-endothelial adhesion by endothelial complement fixation. Nature. 1989;339:314–317. [DOI] [PubMed] [Google Scholar]
- 43. Libby P. Inflammation in atherosclerosis. Nature. 2002;420:868–874. [DOI] [PubMed] [Google Scholar]
- 44. Kostner KM. Activation of the complement system: a crucial link between inflammation and atherosclerosis? Eur J Clin Invest. 2004;34:800–802. [DOI] [PubMed] [Google Scholar]
- 45. Hamilton KK, Hattori R, Esmon CT, Sims PJ. Complement proteins C5b-9 induce vesiculation of the endothelial plasma membrane and expose catalytic surface for assembly of the prothrombinase enzyme complex. J Biol Chem. 1990;265:3809–3814. [PubMed] [Google Scholar]
- 46. Christiansen VJ, Sims PJ, Hamilton KK. Complement C5b-9 increases plasminogen binding and activation on human endothelial cells. Arterioscler Thromb Vasc Biol. 1997;17:164–171. [DOI] [PubMed] [Google Scholar]
- 47. Saadi S, Holzknecht RA, Patte CP, Stern DM, Platt JL. Complement-mediated regulation of tissue factor activity in endothelium. J Exp Med. 1995;182:1807–1814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wu G, Hu W, Shahsafaei A, et al. Complement regulator CD59 protects against atherosclerosis by restricting the formation of complement membrane attack complex. Circ Res. 2009;104:550–558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Tegla CA, Cudrici C, Patel S, et al. Membrane attack by complement: the assembly and biology of terminal complement complexes. Immunol Res. 2011;51:45–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Morgan BP, Campbell AK. The recovery of human polymorphonuclear leucocytes from sublytic complement attack is mediated by changes in intracellular free calcium. Biochem J. 1985;231:205–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Adler S, Baker PJ, Johnson RJ, Ochi RF, Pritzl P, Couser WG. Complement membrane attack complex stimulates production of reactive oxygen metabolites by cultured rat mesangial cells. J Clin Invest. 1986;77:762–767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Bellosillo B, Villamor N, López-Guillermo A, et al. Complement-mediated cell death induced by rituximab in B-cell lymphoproliferative disorders is mediated in vitro by a caspase-independent mechanism involving the generation of reactive oxygen species. Blood. 2001;98:2771–2777. [DOI] [PubMed] [Google Scholar]
- 53. Cybulsky AV, Bonventre JV, Quigg RJ, Lieberthal W, Salant DJ. Cytosolic calcium and protein kinase C reduce complement-mediated glomerular epithelial injury. Kidney Int. 1990;38:803–811. [DOI] [PubMed] [Google Scholar]
- 54. Carney DF, Lang TJ, Shin ML. Multiple signal messengers generated by terminal complement complexes and their role in terminal complement complex elimination. J Immunol. 1990;145:623–629. [PubMed] [Google Scholar]
- 55. Niculescu F, Rus H, Shin ML. Receptor-independent activation of guanine nucleotide-binding regulatory proteins by terminal complement complexes. J Biol Chem. 1994;269:4417–4423. [PubMed] [Google Scholar]
- 56. Niculescu F, Rus H, van Biesen T, Shin ML. Activation of Ras and mitogen-activated protein kinase pathway by terminal complement complexes is G protein dependent. J Immunol. 1997;158:4405–4412. [PubMed] [Google Scholar]
- 57. Niculescu F, Rus H. Mechanisms of signal transduction activated by sublytic assembly of terminal complement complexes on nucleated cells. Immunol Res. 2001;24:191–199. [DOI] [PubMed] [Google Scholar]
- 58. Kilgore KS, Schmid E, Shanley TP, et al. Sublytic concentrations of the membrane attack complex of complement induce endothelial interleukin-8 and monocyte chemoattractant protein-1 through nuclear factor-κ B activation. Am J Pathol. 1997;150:2019–2031. [PMC free article] [PubMed] [Google Scholar]
- 59. Viedt C, Hänsch GM, Brandes RP, Kübler W, Kreuzer J. The terminal complement complex C5b-9 stimulates interleukin-6 production in human smooth muscle cells through activation of transcription factors NF-κ B and AP-1. FASEB J. 2000;14:2370–2372. [DOI] [PubMed] [Google Scholar]
- 60. Torbohm I, Schönermark M, Wingen AM, Berger B, Rother K, Hänsch GM. C5b-8 and C5b-9 modulate the collagen release of human glomerular epithelial cells. Kidney Int. 1990;37:1098–1104. [DOI] [PubMed] [Google Scholar]
- 61. Lovett DH, Haensch GM, Goppelt M, Resch K, Gemsa D. Activation of glomerular mesangial cells by the terminal membrane attack complex of complement. J Immunol. 1987;138:2473–2480. [PubMed] [Google Scholar]
- 62. Stahl RA, Thaiss F, Disser M, Helmchen U, Hora K, Schlöndorff D. Increased expression of monocyte chemoattractant protein-1 in anti-thymocyte antibody-induced glomerulonephritis. Kidney Int. 1993;44:1036–1047. [DOI] [PubMed] [Google Scholar]
- 63. Wagner C, Braunger M, Beer M, Rother K, Hänsch GM. Induction of matrix protein synthesis in human glomerular mesangial cells by the terminal complement complex. Exp Nephrol. 1994;2:51–56. [PubMed] [Google Scholar]
- 64. Kotajima N, Kimura T, Kanda T, et al. Type IV collagen as an early marker for diabetic nephropathy in non-insulin-dependent diabetes mellitus. J Diabetes Complications. 2000;14:13–17. [DOI] [PubMed] [Google Scholar]
- 65. Adler SG, Feld S, Striker L, et al. Glomerular type IV collagen in patients with diabetic nephropathy with and without additional glomerular disease. Kidney Int. 2000;57:2084–2092. [DOI] [PubMed] [Google Scholar]
- 66. Kooyman DL, Byrne GW, McClellan S, et al. In vivo transfer of GPI-linked complement restriction factors from erythrocytes to the endothelium. Science. 1995;269:89–92. [DOI] [PubMed] [Google Scholar]
- 67. Morgan BP. Complement regulatory molecules: application to therapy and transplantation. Immunol Today. 1995;16:257–259. [DOI] [PubMed] [Google Scholar]
- 68. Ninomiya H, Sims PJ. The human complement regulatory protein CD59 binds to the α-chain of C8 and to the “b”domain of C9. J Biol Chem. 1992;267:13675–13680. [PubMed] [Google Scholar]
- 69. Kimberley FC, Sivasankar B, Paul Morgan B. Alternative roles for CD59. Mol Immunol. 2007;44:73–81. [DOI] [PubMed] [Google Scholar]
- 70. Lin RC, Herman J, Henry L, Daniels GL. A family showing inheritance of the Inab phenotype. Transfusion. 1988;28:427–429. [DOI] [PubMed] [Google Scholar]
- 71. Telen MJ, Green AM. The Inab phenotype: characterization of the membrane protein and complement regulatory defect. Blood. 1989;74:437–441. [PubMed] [Google Scholar]
- 72. Reid ME, Mallinson G, Sim RB, et al. Biochemical studies on red blood cells from a patient with the Inab phenotype (decay-accelerating factor deficiency). Blood. 1991;78:3291–3297. [PubMed] [Google Scholar]
- 73. Sun X, Funk CD, Deng C, Sahu A, Lambris JD, Song WC. Role of decay-accelerating factor in regulating complement activation on the erythrocyte surface as revealed by gene targeting. Proc Natl Acad Sci USA. 1999;96:628–633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Yamashina M, Ueda E, Kinoshita T, et al. Inherited complete deficiency of 20-kilodalton homologous restriction factor (CD59) as a cause of paroxysmal nocturnal hemoglobinuria. N Engl J Med. 1990;323:1184–1189. [DOI] [PubMed] [Google Scholar]
- 75. Nevo Y, Ben-Zeev B, Tabib A, et al. CD59 deficiency is associated with chronic hemolysis and childhood relapsing immune-mediated polyneuropathy. Blood. 2013;121:129–135. [DOI] [PubMed] [Google Scholar]
- 76. Luzzatto L, Bessler M. The dual pathogenesis of paroxysmal nocturnal hemoglobinuria. Curr Opin Hematol. 1996;3:101–110. [DOI] [PubMed] [Google Scholar]
- 77. Miyata T, Takeda J, Iida Y, et al. The cloning of PIG-A, a component in the early step of GPI-anchor biosynthesis. Science. 1993;259:1318–1320. [DOI] [PubMed] [Google Scholar]
- 78. Takeda J, Miyata T, Kawagoe K, et al. Deficiency of the GPI anchor caused by a somatic mutation of the PIG-A gene in paroxysmal nocturnal hemoglobinuria. Cell. 1993;73:703–711. [DOI] [PubMed] [Google Scholar]
- 79. Miyata T, Yamada N, Iida Y, et al. Abnormalities of PIG-A transcripts in granulocytes from patients with paroxysmal nocturnal hemoglobinuria. N Engl J Med. 1994;330:249–255. [DOI] [PubMed] [Google Scholar]
- 80. Halperin JA, Nicholson-Weller A. Paroxysmal nocturnal hemoglobinuria. A complement-mediated disease. Complement Inflamm. 1989;6:65–72. [DOI] [PubMed] [Google Scholar]
- 81. Parker CJ. Molecular basis of paroxysmal nocturnal hemoglobinuria. Stem Cells. 1996;14:396–411. [DOI] [PubMed] [Google Scholar]
- 82. Falk RJ, Dalmasso AP, Kim Y, et al. Neoantigen of the polymerized ninth component of complement. Characterization of a monoclonal antibody and immunohistochemical localization in renal disease. J Clin Invest. 1983;72:560–573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Falk RJ, Scheinman JI, Mauer SM, Michael AF. Polyantigenic expansion of basement membrane constituents in diabetic nephropathy. Diabetes. 1983;32(suppl 2):34–39. [DOI] [PubMed] [Google Scholar]
- 84. Falk RJ, Sisson SP, Dalmasso AP, Kim Y, Michael AF, Vernier RL. Ultrastructural localization of the membrane attack complex of complement in human renal tissues. Am J Kidney Dis. 1987;9:121–128. [DOI] [PubMed] [Google Scholar]
- 85. Bouwman LH, Eerligh P, Terpstra OT, et al. Elevated levels of mannose-binding lectin at clinical manifestation of type 1 diabetes in juveniles. Diabetes. 2005;54:3002–3006. [DOI] [PubMed] [Google Scholar]
- 86. Hansen TK, Thiel S, Knudsen ST, et al. Elevated levels of mannan-binding lectin in patients with type 1 diabetes. J Clin Endocrinol Metab. 2003;88:4857–4861. [DOI] [PubMed] [Google Scholar]
- 87. Saraheimo M, Forsblom C, Hansen TK, et al. Increased levels of mannan-binding lectin in type 1 diabetic patients with incipient and overt nephropathy. Diabetologia. 2005;48:198–202. [DOI] [PubMed] [Google Scholar]
- 88. Kaunisto MA, Sjölind L, Sallinen R, et al. Elevated MBL concentrations are not an indication of association between the MBL2 gene and type 1 diabetes or diabetic nephropathy. Diabetes. 2009;58:1710–1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Kaida K, Kusunoki S. Antibodies to gangliosides and ganglioside complexes in Guillain-Barré syndrome and Fisher syndrome: mini-review. J Neuroimmunol. 2010;223:5–12. [DOI] [PubMed] [Google Scholar]
- 90. Haskard DO, Boyle JJ, Mason JC. The role of complement in atherosclerosis. Curr Opin Lipidol. 2008;19:478–482. [DOI] [PubMed] [Google Scholar]
- 91. Niculescu F, Rus H. Complement activation and atherosclerosis. Mol Immunol. 1999;36:949–955. [DOI] [PubMed] [Google Scholar]
- 92. Lappegård KT, Garred P, Jonasson L, et al. A vital role for complement in heart disease. Mol Immunol. 2014;61:126–134. [DOI] [PubMed] [Google Scholar]
- 93. Niculescu F, Rus HG, Vlaicu R. Activation of the human terminal complement pathway in atherosclerosis. Clin Immunol Immunopathol. 1987;45:147–155. [DOI] [PubMed] [Google Scholar]
- 94. Pang AS, Katz A, Minta JO. C3 deposition in cholesterol-induced atherosclerosis in rabbits: a possible etiologic role for complement in atherogenesis. J Immunol. 1979;123:1117–1122. [PubMed] [Google Scholar]
- 95. Schmiedt W, Kinscherf R, Deigner HP, et al. Complement C6 deficiency protects against diet-induced atherosclerosis in rabbits. Arterioscler Thromb Vasc Biol. 1998;18:1790–1795. [DOI] [PubMed] [Google Scholar]
- 96. Seifert PS, Hugo F, Hansson GK, Bhakdi S. Prelesional complement activation in experimental atherosclerosis. Terminal C5b-9 complement deposition coincides with cholesterol accumulation in the aortic intima of hypercholesterolemic rabbits. Lab Invest. 1989;60:747–754. [PubMed] [Google Scholar]
- 97. Seifert PS, Kazatchkine MD. The complement system in atherosclerosis. Atherosclerosis. 1988;73:91–104. [DOI] [PubMed] [Google Scholar]
- 98. Tominaga M, Eguchi H, Manaka H, Igarashi K, Kato T, Sekikawa A. Impaired glucose tolerance is a risk factor for cardiovascular disease, but not impaired fasting glucose. The Funagata Diabetes Study. Diabetes Care. 1999;22:920–924. [DOI] [PubMed] [Google Scholar]
- 99. Jarrett RJ, McCartney P, Keen H. The Bedford survey: ten year mortality rates in newly diagnosed diabetics, borderline diabetics and normoglycaemic controls and risk indices for coronary heart disease in borderline diabetics. Diabetologia. 1982;22:79–84. [DOI] [PubMed] [Google Scholar]
- 100. Onat A, Can G, Rezvani R, Cianflone K. Complement C3 and cleavage products in cardiometabolic risk. Clin Chim Acta. 2011;412:1171–1179. [DOI] [PubMed] [Google Scholar]
- 101. Hertle E, van Greevenbroek MM, Stehouwer CD. Complement C3: an emerging risk factor in cardiometabolic disease. Diabetologia. 2012;55:881–884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Cianflone K, Xia Z, Chen LY. Critical review of acylation-stimulating protein physiology in humans and rodents. Biochim Biophys Acta. 2003;1609:127–143. [DOI] [PubMed] [Google Scholar]
- 103. Fortpied J, Vertommen D, Van Schaftingen E. Binding of mannose-binding lectin to fructosamines: a potential link between hyperglycaemia and complement activation in diabetes. Diabetes Metab Res Rev. 2010;26:254–260. [DOI] [PubMed] [Google Scholar]
- 104. Ostergaard JA, Bjerre M, Dagnaes-Hansen F, Hansen TK, Thiel S, Flyvbjerg A. Diabetes-induced changes in mannan-binding lectin levels and complement activation in a mouse model of type 1 diabetes. Scand J Immunol. 2013;77:187–194. [DOI] [PubMed] [Google Scholar]
- 105. Orchard TJ, Virella G, Forrest KY, Evans RW, Becker DJ, Lopes-Virella MF. Antibodies to oxidized LDL predict coronary artery disease in type 1 diabetes: a nested case-control study from the Pittsburgh Epidemiology of Diabetes Complications Study. Diabetes. 1999;48:1454–1458. [DOI] [PubMed] [Google Scholar]
- 106. Witztum JL, Steinbrecher UP, Fisher M, Kesaniemi A. Nonenzymatic glucosylation of homologous low density lipoprotein and albumin renders them immunogenic in the guinea pig. Proc Natl Acad Sci USA. 1983;80:2757–2761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Weiss JS, Sang DN, Albert DM. Immunofluorescent characteristics of the diabetic cornea. Cornea. 1990;9:131–138. [PubMed] [Google Scholar]
- 108. Shapiro R, McManus MJ, Zalut C, Bunn HF. Sites of nonenzymatic glycosylation of human hemoglobin A. J Biol Chem. 1980;255:3120–3127. [PubMed] [Google Scholar]
- 109. Iberg N, Flückiger R. Nonenzymatic glycosylation of albumin in vivo. Identification of multiple glycosylated sites. J Biol Chem. 1986;261:13542–13545. [PubMed] [Google Scholar]
- 110. Shilton BH, Walton DJ. Sites of glycation of human and horse liver alcohol dehydrogenase in vivo. J Biol Chem. 1991;266:5587–5592. [PubMed] [Google Scholar]
- 111. Shilton BH, Campbell RL, Walton DJ. Site specificity of glycation of horse liver alcohol dehydrogenase in vitro. Eur J Biochem. 1993;215:567–572. [DOI] [PubMed] [Google Scholar]
- 112. Calvo C, Ulloa N, Campos M, Verdugo C, Ayrault-Jarrier M. The preferential site of non-enzymatic glycation of human apolipoprotein A-I in vivo. Clin Chim Acta. 1993;217:193–198. [DOI] [PubMed] [Google Scholar]
- 113. Flückiger R, Strang CJ. Structural requirements for protein glycation. Protein Sci. 1995;4:699S. [Google Scholar]
- 114. Fletcher CM, Harrison RA, Lachmann PJ, Neuhaus D. Structure of a soluble, glycosylated form of the human complement regulatory protein CD59. Structure. 1994;2:185–199. [DOI] [PubMed] [Google Scholar]
- 115. Bodian DL, Davis SJ, Morgan BP, Rushmere NK. Mutational analysis of the active site and antibody epitopes of the complement-inhibitory glycoprotein, CD59. J Exp Med. 1997;185:507–516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Yu J, Abagyan R, Dong S, Gilbert A, Nussenzweig V, Tomlinson S. Mapping the active site of CD59. J Exp Med. 1997;185:745–753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Vischer UM. Hyperglycemia and the pathogenesis of atherosclerosis: lessons from murine models. Eur J Endocrinol. 1999;140:1–3. [DOI] [PubMed] [Google Scholar]
- 118. Hsueh W, Abel ED, Breslow JL, et al. Recipes for creating animal models of diabetic cardiovascular disease. Circ Res. 2007;100:1415–1427. [DOI] [PubMed] [Google Scholar]
- 119. Brosius FC, 3rd, Alpers CE, Bottinger EP, et al. Mouse models of diabetic nephropathy. J Am Soc Nephrol. 2009;20:2503–2512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. An G, Miwa T, Song WL, et al. CD59 but not DAF deficiency accelerates atherosclerosis in female ApoE knockout mice. Mol Immunol. 2009;46:1702–1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Buono C, Come CE, Witztum JL, et al. Influence of C3 deficiency on atherosclerosis. Circulation. 2002;105:3025–3031. [DOI] [PubMed] [Google Scholar]
- 122. Liu F, Ge X, Sahoo R, Qin X, Halperin J. The deficiency of CD59 accelerates the development of diabetic atherosclerosis in mice. Mol Immunol. 2014;61:223. [Google Scholar]
- 123. Peterson CM, Jones RL, Koenig RJ, Melvin ET, Lehrman ML. Reversible hematologic sequelae of diabetes mellitus. Ann Intern Med. 1977;86:425–429. [DOI] [PubMed] [Google Scholar]
- 124. Bern MM, Busick EJ. Disorders of the blood and diabetes. In: Marble A, Krall L, Bradley R, Chistlieb A, Soeldner J, eds. Joslin's Diabetes Mellitus. Philadelphia, PA: Lea & Febiger; 1985:748. [Google Scholar]
- 125. Giugliano D. Glycosylated haemoglobin and reticulocyte count in diabetes. Diabetologia. 1982;22:223. [DOI] [PubMed] [Google Scholar]
- 126. Carr ME. Diabetes mellitus: a hypercoagulable state. J Diabetes Complications. 2001;15:44–54. [DOI] [PubMed] [Google Scholar]
- 127. Gardner FH, Robin ED, Travis DM, Julian DG, Crump CH. Some pathophysiologic aspects of paroxysmal nocturnal hemoglobinuria. J Clin Invest. 1958;37:895. [Google Scholar]
- 128. Hill A, Kelly RJ, Hillmen P. Thrombosis in paroxysmal nocturnal hemoglobinuria. Blood. 2013;121:4985–4996; quiz 5105. [DOI] [PubMed] [Google Scholar]
- 129. Wiedmer T, Hall SE, Ortel TL, Kane WH, Rosse WF, Sims PJ. Complement-induced vesiculation and exposure of membrane prothrombinase sites in platelets of paroxysmal nocturnal hemoglobinuria. Blood. 1993;82:1192–1196. [PubMed] [Google Scholar]
- 130. Qin X, Hu W, Song W, et al. Balancing role of nitric oxide in complement-mediated activation of platelets from mCd59a and mCd59b double-knockout mice. Am J Hematol. 2009;84:221–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Qin X, Hu W, Song W, et al. Generation and phenotyping of mCd59a and mCd59b double-knockout mice. Am J Hematol. 2009;84:65–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Qin X, Krumrei N, Grubissich L, et al. Deficiency of the mouse complement regulatory protein mCd59b results in spontaneous hemolytic anemia with platelet activation and progressive male infertility. Immunity. 2003;18:217–227. [DOI] [PubMed] [Google Scholar]
- 133. Meri S, Lehto T, Sutton CW, Tyynelä J, Baumann M. Structural composition and functional characterization of soluble CD59: heterogeneity of the oligosaccharide and glycophosphoinositol (GPI) anchor revealed by laser-desorption mass spectrometric analysis. Biochem J. 1996;316:923–935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Lehto T, Meri S. Interactions of soluble CD59 with the terminal complement complexes. CD59 and C9 compete for a nascent epitope on C8. J Immunol. 1993;151:4941–4949. [PubMed] [Google Scholar]
- 135. Lehto T, Honkanen E, Teppo AM, Meri S. Urinary excretion of protectin (CD59), complement SC5b-9 and cytokines in membranous glomerulonephritis. Kidney Int. 1995;47:1403–1411. [DOI] [PubMed] [Google Scholar]
- 136. Väkevä A, Lehto T, Takala A, Meri S. Detection of a soluble form of the complement membrane attack complex inhibitor CD59 in plasma after acute myocardial infarction. Scand J Immunol. 2000;52:411–414. [DOI] [PubMed] [Google Scholar]
- 137. Ghosh P, Sahoo R, Vaidya A, et al. A specific and sensitive assay for blood levels of glycated CD59: a novel biomarker for diabetes. Am J Hematol. 2013;88:670–676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Ghosh P, Vaidya A, Sahoo R, et al. Glycation of the complement regulatory protein CD59 is a novel biomarker for glucose handling in humans. J Clin Endocrinol Metab. 2014;99:E999–E1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Cataland SR, Holers VM, Geyer S, Yang S, Wu HM. Biomarkers of terminal complement activation confirm the diagnosis of aHUS and differentiate aHUS from TTP. Blood. 2014;123:3733–3738. [DOI] [PubMed] [Google Scholar]
- 140. Ricklin D, Lambris JD. Complement-targeted therapeutics. Nat Biotechnol. 2007;25:1265–1275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Ricklin D, Lambris JD. Progress and trends in complement therapeutics. Adv Exp Med Biol. 2013;735:1–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Forneris F, Ricklin D, Wu J, et al. Structures of C3b in complex with factors B and D give insight into complement convertase formation. Science. 2010;330:1816–1820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Janssen BJ, Christodoulidou A, McCarthy A, Lambris JD, Gros P. Structure of C3b reveals conformational changes that underlie complement activity. Nature. 2006;444:213–216. [DOI] [PubMed] [Google Scholar]
- 144. Janssen BJ, Huizinga EG, Raaijmakers HC, et al. Structures of complement component C3 provide insights into the function and evolution of immunity. Nature. 2005;437:505–511. [DOI] [PubMed] [Google Scholar]
- 145. Torreira E, Tortajada A, Montes T, Rodríguez de Córdoba S, Llorca O. 3D structure of the C3bB complex provides insights into the activation and regulation of the complement alternative pathway convertase. Proc Natl Acad Sci USA. 2009;106:882–887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Wiesmann C, Katschke KJ, Yin J, et al. Structure of C3b in complex with CRIg gives insights into regulation of complement activation. Nature. 2006;444:217–220. [DOI] [PubMed] [Google Scholar]
- 147. Wu J, Wu YQ, Ricklin D, Janssen BJ, Lambris JD, Gros P. Structure of complement fragment C3b-factor H and implications for host protection by complement regulators. Nat Immunol. 2009;10:728–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Rooijakkers SH, Wu J, Ruyken M, et al. Structural and functional implications of the alternative complement pathway C3 convertase stabilized by a staphylococcal inhibitor. Nat Immunol. 2009;10:721–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Garlatti V, Chouquet A, Lunardi T, et al. Cutting edge: C1q binds deoxyribose and heparan sulfate through neighboring sites of its recognition domain. J Immunol. 2010;185:808–812. [DOI] [PubMed] [Google Scholar]
- 150. Laursen NS, Gordon N, Hermans S, et al. Structural basis for inhibition of complement C5 by the SSL7 protein from Staphylococcus aureus. Proc Natl Acad Sci USA. 2010;107:3681–3686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Shaw CD, Storek MJ, Young KA, et al. Delineation of the complement receptor type 2-C3d complex by site-directed mutagenesis and molecular docking. J Mol Biol. 2010;404:697–710. [DOI] [PubMed] [Google Scholar]
- 152. Gingras AR, Girija UV, Keeble AH, et al. Structural basis of mannan-binding lectin recognition by its associated serine protease MASP-1: implications for complement activation. Structure. 2011;19:1635–1643. [DOI] [PubMed] [Google Scholar]
- 153. Kajander T, Lehtinen MJ, Hyvärinen S, et al. Dual interaction of factor H with C3d and glycosaminoglycans in host-nonhost discrimination by complement. Proc Natl Acad Sci USA. 2011;108:2897–2902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Morgan HP, Schmidt CQ, Guariento M, et al. Structural basis for engagement by complement factor H of C3b on a self surface. Nat Struct Mol Biol. 2011;18:463–470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. van den Elsen JM, Isenman DE. A crystal structure of the complex between human complement receptor 2 and its ligand C3d. Science. 2011;332:608–611. [DOI] [PubMed] [Google Scholar]
- 156. Hadders MA, Bubeck D, Roversi P, et al. Assembly and regulation of the membrane attack complex based on structures of C5b6 and sC5b9. Cell Reports. 2012;1:200–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Degn SE, Jensenius JC, Thiel S. Disease-causing mutations in genes of the complement system. Am J Hum Genet. 2011;88:689–705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Hillmen P, Young NS, Schubert J, et al. The complement inhibitor eculizumab in paroxysmal nocturnal hemoglobinuria. N Engl J Med. 2006;355:1233–1243. [DOI] [PubMed] [Google Scholar]
- 159. Hillmen P, Muus P, Dührsen U, et al. Effect of the complement inhibitor eculizumab on thromboembolism in patients with paroxysmal nocturnal hemoglobinuria. Blood. 2007;110:4123–4128. [DOI] [PubMed] [Google Scholar]
- 160. Rother RP, Rollins SA, Mojcik CF, Brodsky RA, Bell L. Discovery and development of the complement inhibitor eculizumab for the treatment of paroxysmal nocturnal hemoglobinuria. Nat Biotechnol. 2007;25:1256–1264. [DOI] [PubMed] [Google Scholar]
- 161. Legendre CM, Licht C, Muus P, et al. Terminal complement inhibitor eculizumab in atypical hemolytic-uremic syndrome. N Engl J Med. 2013;368:2169–2181. [DOI] [PubMed] [Google Scholar]
- 162. Hillmen P, Richards SJ. Implications of recent insights into the pathophysiology of paroxysmal nocturnal haemoglobinuria. Br J Haematol. 2000;108:470–479. [DOI] [PubMed] [Google Scholar]
- 163. Risitano AM. Paroxysmal nocturnal hemoglobinuria and other complement-mediated hematological disorders. Immunobiology. 2012;217:1080–1087. [DOI] [PubMed] [Google Scholar]
- 164. Lanting LC, Joung IM, Mackenbach JP, Lamberts SW, Bootsma AH. Ethnic differences in mortality, end-stage complications, and quality of care among diabetic patients: a review. Diabetes Care. 2005;28:2280–2288. [DOI] [PubMed] [Google Scholar]
- 165. Matsushita M, Endo Y, Nonaka M, Fujita T. Complement-related serine proteases in tunicates and vertebrates. Curr Opin Immunol. 1998;10:29–35. [DOI] [PubMed] [Google Scholar]
- 166. Podack ER. Molecular composition of the tubular structure of the membrane attack complex of complement. J Biol Chem. 1984;259:8641–8647. [PubMed] [Google Scholar]
- 167. Podack ER, Müller-Eberhard HJ, Horst H, Hoppe W. Membrane attach complex of complement (MAC): three-dimensional analysis of MAC-phospholipid vesicle recombinants. J Immunol. 1982;128:2353–2357. [PubMed] [Google Scholar]
- 168. Podack ER, Tschopp J. Polymerization of the ninth component of complement (C9): formation of poly(C9) with a tubular ultrastructure resembling the membrane attack complex of complement. Proc Natl Acad Sci USA. 1982;79:574–578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Podack ER, Tschopp J. Membrane attack by complement. Mol Immunol. 1984;21:589–603. [DOI] [PubMed] [Google Scholar]
- 170. Tschopp J, Engel A, Podack ER. Molecular weight of poly(C9). 12 to 18 C9 molecules form the transmembrane channel of complement. J Biol Chem. 1984;259:1922–1928. [PubMed] [Google Scholar]
- 171. Laine RO, Esser AF. Detection of refolding conformers of complement protein C9 during insertion into membranes. Nature. 1989;341:63–65. [DOI] [PubMed] [Google Scholar]
- 172. Mayer MM. Complement. Historical perspectives and some current issues. Complement. 1984;1:2–26. [PubMed] [Google Scholar]
- 173. Ramm LE, Whitlow MB, Mayer MM. Transmembrane channel formation by complement: functional analysis of the number of C5b6, C7, C8, and C9 molecules required for a single channel. Proc Natl Acad Sci USA. 1982;79:4751–4755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174. Ramm LE, Whitlow MB, Mayer MM. The relationship between channel size and the number of C9 molecules in the C5b-9 complex. J Immunol. 1985;134:2594–2599. [PubMed] [Google Scholar]
- 175. Nicholson-Weller A, March JP, Rosen CE, Spicer DB, Austen KF. Surface membrane expression by human blood leukocytes and platelets of decay-accelerating factor, a regulatory protein of the complement system. Blood. 1985;65:1237–1244. [PubMed] [Google Scholar]
- 176. Brooimans RA. Relative roles of decay-accelerating factor, membrane cofactor protein, and CD59 in the protection of human endothelial cells against complement mediated lysis. Eur J Immunol. 1992;22:3135–3140. [DOI] [PubMed] [Google Scholar]
- 177. Cervoni F, Oglesby TJ, Adams EM, et al. Identification and characterization of membrane cofactor protein of human spermatozoa. J Immunol. 1992;148:1431–1437. [PubMed] [Google Scholar]
- 178. Beynon HL, Davies KA, Haskard DO, Walport MJ. Erythrocyte complement receptor type 1 and interactions between immune complexes, neutrophils, and endothelium. J Immunol. 1994;153:3160–3167. [PubMed] [Google Scholar]
- 179. McDonald JF, Nelsestuen GL. Potent inhibition of terminal complement assembly by clusterin: characterization of its impact on C9 polymerization. Biochemistry. 1997;36:7464–7473. [DOI] [PubMed] [Google Scholar]
- 180. Podack ER, Müller-Eberhard HJ. Binding of desoxycholate, phosphatidylcholine vesicles, lipoprotein and of the S-protein to complexes of terminal complement components. J Immunol. 1978;121:1025–1030. [PubMed] [Google Scholar]
- 181. Davies A, Simmons DL, Hale G, et al. CD59, an LY-6-like protein expressed in human lymphoid cells, regulates the action of the complement membrane attack complex on homologous cells. J Exp Med. 1989;170:637–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182. Huang Y, Qiao F, Abagyan R, Hazard S, Tomlinson S. Defining the CD59–C9 binding interaction. J Biol Chem. 2006;281:27398–27404. [DOI] [PubMed] [Google Scholar]
- 183. Hüsler T, Lockert DH, Kaufman KM, Sodetz JM, Sims PJ. Chimeras of human complement C9 reveal the site recognized by complement regulatory protein CD59. J Biol Chem. 1995;270:3483–3486. [DOI] [PubMed] [Google Scholar]
- 184. Meri S, Morgan BP, Wing M, et al. Human protectin (CD59), an 18–20-kD homologous complement restriction factor, does not restrict perforin-mediated lysis. J Exp Med. 1990;172:367–370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185. Petranka JG, Fleenor DE, Sykes K, Kaufman RE, Rosse WF. Structure of the CD59-encoding gene: further evidence of a relationship to murine lymphocyte antigen Ly-6 protein. Proc Natl Acad Sci USA. 1992;89:7876–7879. [DOI] [PMC free article] [PubMed] [Google Scholar]