

Abstract
Dioscoreaceae, a kind of yam plant, has been recommended for treatment of chronic inflammatory conditions. However, the mechanisms are poorly defined. Methyl protodioscin (MPD) is one of the main bioactive components in Dioscoreaceae. Here, we aim to determine the mechanisms by which MPD ameliorates intestinal inflammation. Surgical intestinal specimens were collected from inflammatory bowel diseases (IBD) patients to perform organ culture. Experimental colitis was induced in mice by dextran sulfate sodium (DSS) or Citrobacter rodentium, and was then treated with MPD. NF-κB activation, expression of mucosal pro-inflammatory cytokines, disease severity, and epithelial proliferation/apoptosis were determined. Mouse crypts and Caco-2 monolayers were cultured to observe the effect of MPD upon intestinal epithelial differentiation and barrier function. We found that MPD increased the percentage of survival from high-dose DSS-(4%) treated mice, and accelerated mucosal healing and epithelial proliferation in low-dose DSS-(2.5%) treated mice characterized by marked reduction in NF-κB activation, pro-inflammatory cytokines expression and bacterial translocation. Consistently, MPD protected colonic mucosa from C. rodentium-induced colonic inflammation and bacterial colonization. In vitro studies showed that MPD significantly increased crypt formation and restored intestinal barrier dysfunction induced by pro-inflammatory cytokines. In conclusion, MPD ameliorates the intestinal mucosal inflammation by modulating the intestinal immunity to enhance intestinal barrier differentiation. MPD could be an alternative for treating chronic intestinal inflammatory diseases.
Keywords: Colitis, inflammatory bowel diseases (IBD), intestinal barrier function, intestinal stem cells, methyl protodioscin (MPD), traditional chinese herbal (TCH)
Introduction
The inflammatory bowel diseases (IBD), including Crohn's Disease (CD) and ulcerative colitis (UC), are believed to be caused by a complex interaction between genetic susceptibility, environmental triggers and immune-mediated tissue injury leading to chronic relapsing intestinal inflammation (Bouma and Strober 2003). Pediatric IBD patients often experience growth velocity impairment, malnutrition, pubertal delay, and bone demineralization (Grossman and Baldassano 2008). Therefore, pediatric IBD patients require special consideration, and medical treatment must be optimized to promote clinical improvement and reverse growth failure with minimal toxicity. However, corticosteroids (Prednisone or Methylprednisolone) may cause profound growth retardation in children. Immunomodulators, including azathioprine (AZA) and 6-mercaptopurine (6-MP), have been determined the adverse effects including hepatotoxicity, myelosuppression, pancreatitis, and allergic reactions (Andres and Friedman 1999). Current biologic therapies like infliximab have been used to treat moderate to severe disease in patients who do not respond to 5-ASA, antibiotics, steroids, or immunomodulators; however, approximately 30% of patients do not respond to infliximab (Hanauer 2007). Therefore, exploring safer and more efficacious approaches is a long-term strategy for pediatric IBD treatment.
Traditional Chinese herbal (TCH) remedies are rapidly gaining attention in the West as sources of new drugs, dietary supplements and functional foods. TCH also has a very long history and rich experience in effectively treating some intestinal symptoms of IBD, for instance, diarrhea and intestinal spasm, etc (Hilsden et al. 1998; Langmead and Rampton 2006). Dioscoreaceae, a kind of yam plants, is recommended as a treatment for rheumatic conditions, biliary colic, irritable bowel syndrome, diverticulitis and intestinal inflammation, or spasm in TCH (Ulbricht et al. 2003). However, the primary bioactive elements are elusive and dosage is extremely variable. Methyl protodioscin (MPD) is a member of the furostan saponin family, which broadly exists in Dioscoreaceae plant. MPD was extracted and purified from the rhizome of Dioscorea collettii var. hypoglauca (Dioscoreaceae) (Hu et al. 1997). Importantly, MPD can be also synthesized (Cheng et al. 2003). MPD was tested for in vitro cytotoxicity against 60 human cancer cell lines in the anticancer drug screen by National Cancer Institute (NCI) and showed potent activity against most cell lines from leukemia and solid tumors (Hu and Yao 2003). It has been shown that MPD induces a dramatic decline of cytoplasmic Ca2+ concentration, which could lead to G2/M arrest and apoptosis of K562 cells (human leukemia cell line) through the efflux of Ca2+ from cell and Ca2+ uptake into mitochondria (Liu et al. 2005) Most importantly, a preclinical pharmacodynamic study showed that high dosage (80 mg/kg i.v.) had no serious adverse effects (He et al. 2006). A recent study found that it also had antiosteoporotic activity without any side effects (Yin et al. 2004). MPD has a clear pharmacokinetic (PK)/pharmacodynamic (PD) profile and a defined chemical structure, which is very rare in TCH since often combinatorial substances are thought to be effective (He et al. 2006; Cao et al. 2008).
Immune responses in the intestine are tightly regulated to maintain a delicate intercellular balance that promotes host defense to suppress chronic inflammation (Maloy and Powrie 2011). Disruption of this balance can lead to chronic intestinal inflammation characteristic of IBD (Salzman et al. 2010). Amongst these factors, intestinal epithelial (IEC) innate immunity including intestinal barrier, antimicrobial peptides (AMP), and innate immune receptors, is critical for host defense against invading microorganisms and for the subsequent generation of an adaptive immune response, thus playing a critical role in the pathogenesis of IBD (Medzhitov 2007; Turner 2009). The transcription nuclear factor-κB (NF-κB), as well as its regulation of cytokine production, exaggerates chronic intestinal inflammation in both CD and UC (Schreiber et al. 1998; Fichtner-Feigl et al. 2005). In turn, inflammation in IBD and other conditions can be attenuated through an inhibition of NF-κB activity (Fichtner-Feigl et al. 2005). NF-κB activation in macrophages and T cells leads to increased production of cytokines in the intestinal mucosa (Tak and Firestein 2001). Furthermore, increased TNFα, IFNγ, and interleukin 1 β (IL-1β) production causes loss of IEC integrity and increased tight junction (TJ) permeability, leading to microbial translocation and chronic intestinal inflammation (Han et al. 2004; Al-Sadi et al. 2008; Boivin et al. 2009). Utilizing human organ culture, experimental colitis, and human and mouse cell culture, we here for the first time determine the unique anti-inflammatory mechanism for the steroidal herbal compound, MPD, in ameliorating chronic mucosal inflammation and infectious colitis. Furthermore, our findings could lead to a further study with IBD patients to examine the effects of MPD upon immune modulation.
Materials and Methods
Materials
All chemicals and antibodies were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise noted. Antibodies specific for Cleaved Caspase-3, and p65 (phospho-ser276) were from Cell Signaling Technology (Danvers, MA). Antibodies specific for p65, IκBα, and β and Tubulin were from Santa Cruz Biotechnology (Santa Cruz, CA). Antibody specific for Ki67 is from DAKO (Carpinteria, CA). CD3, CD4, and AnnexinV were from eBioscience (San Diego, CA). For crypt culture, Matrigel was purchased from BD Bioscience (San Jose, CA), and EGF, Noggin, and R-spondin were from R&D (Minneapolis, MN), advanced DMEM/F12 media were from Life Technologies (Danvers, MA).
MPD preparation
MPD was isolated and purified by the Xinsheng Yao as published methodologically (Cheng et al. 2003; Cao et al. 2008, 2010). Briefly, the root slice of Dioscorea nipponica Makino was extracted with 60% ethanol. The concentrated extract was then subjected to macroreticular resin column to elute with 30%, 55%, and 95% alcohol stepwise. The isolated bioactive fraction was purified with methanol to obtain MPD of a purity of 98.7% as determined by mass spectroscopy, which was kindly supplied and analyzed by the Institute of Traditional Chinese Medicine (TCM) and Natural Products, Jinan University.
Animal resources and maintenance
The animal study protocol was approved by the CHRF Institutional Animal Care and Use Committee (IACUC 2013-0051 1E03030, Han). All C57BL/6 mice used in these studies have been maintained for ten generations in specific pathogen-free (SPF) conditions in the Children's Hospital Research Foundation (CHRF) Animal Care Facility. Two-month-old mice with mixed-gender were used to study MPD treatment of colitis.
Patient-based studies
Surgical colonic specimens from colon surgical resection were obtained from children with UC after informed consent. The diagnosis of UC was made by the patient's primary gastroenterologist on the basis of established clinical, radiological, and histological criteria. The de-identified IRB protocol was approved by the Cincinnati Children's Hospital Medical Center Institutional Review Board and the Cincinnati Children's Hospital Medical Center General Clinical Research Center Scientific Advisory Committee.
Animal model of colitis and MPD administration
Intestinal inflammation was induced by providing C57BL/6 mice with 2.5% or 4% DSS water (M.W. 36,000–50,000; MP Biomedicals, Santa Ana, CA, USA) for either 7 days for acute studies or 7 days followed by 5 days of water for healing phase studies. In therapeutic groups, we intraperitoneally (i.p.) gave mice MPD once daily on day 3 together with DSS for either 5 days or 10 days, respectively to observe the effect of MPD on DSS-induced acute injury or mucosal healing phase (Fig.1A). Animal weights and stool scores (0, normal; 1, soft; 2, diarrhea/minimal anorectal bleeding; 3, diarrhea/severe anorectal bleeding) were recorded daily. Mice were sacrificed; the colon was removed. For each colon fragment, longitudinal cuts of tissue in which the three colon sections had clearly visible intestinal lumen were used in histological scoring. Scoring parameters included quantitation of the area of middle and distal colon involved, edema, erosion/ulceration of the epithelial monolayer, crypt loss/damage, and infiltration of immune cells into the mucosa. Severity was graded on a scale that, for the area involved (erosion/ulceration and crypt loss), was defined as 0 (normal), 1 (0–10%), 2 (10–25%), 3 (25–50%), and 4 (>50%). Edema and immune cell infiltration were scored as: 0, absent; 1, weak; 2, moderate; and 3, severe. Total disease score was expressed as the mean of all combined scores per genotype (Cooper et al. 1993; Hogan et al. 2006).
Figure 1.
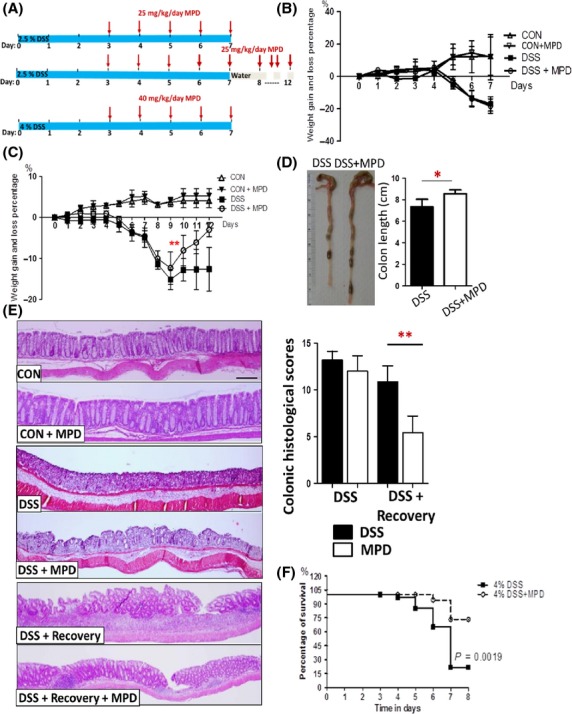
MPD treatment mitigates intestinal inflammation to increase mouse survival from lethal colitis. (A) Diagram of MPD treatment protocol upon DSS-induced colitis. (B) Intestinal inflammation was induced by 2.5% DSS in mice for acute injury study, mice were then treated with vehicle (sterile saline) or MPD (25 mg/kg) once daily for 5 days starting on day 3. Weight gain and loss were recorded. (C) Intestinal inflammation was induced by 2.5% DSS for 7 days followed by 5 days of water for healing studies. MPD was given to mice at 25 mg/kg daily on day 3 together with DSS for 10 days. Curve of weight gain and loss percentage was showed. Colon lengths were measured after mice were sacrificed on day 12 (D). Disease severity of DSS-induced colitic injury was scored (E). Results are expressed as mean ± SEM (n = 7−8 per group). Data were analyzed using ANOVA, **P < 0.01 versus controls. (F) C57BL/6 mice were orally administrated with 4% DSS in drinking water for 7 days; simultaneously, these mice were i.p. treated with MPD (40 mg/kg) once daily for 5 days. Kaplan–Meier curve was used to estimate the percent survival of DSS-induced colitis with and without MPD intervention, n = 8 each group.
Citrobacter rodentium infection of mice and MPD treatment
Groups of C57BL/6 mice with free access to food and water were infected by oral gavage with approximately 1.5 × 109 CFU of freshly prepared C. rodentium resuspended in 100 μL sterile PBS. Groups of mice were sacrificed at 10 days after infection and MPD treatment (25 mg/kg) started on day 3 for 7 days. Tissues including jejunum and ileum from additional naïve (noninfected) age- and gender-matched mice were obtained. Disease severity was scored as colonic mucosal inflammation, IEC hyperplasia, and inflammatory cell infiltration separately, and combined total scores as described as colonic mucosal inflammation (Mann et al. 2013).
Measurement of bacterial translocation
Bacterial translocation to mesenteric lymph node (MLN) was determined as described previously (Han et al. 2004).
Real-time quantitative PCR
Total RNA was isolated from frozen tissue using RNeasy Mini Kit (QIAGEN, Valencia, CA) according to the manufacturer's protocol. Using specific gene primers (IL-17, TNFα, IL12p35 & 40, and IFNγ, IL-23, β-defensin1, β-defensin3, β-defensin10, RegIIIγ, and GAPDH), PCR reactions were performed with Brilliant II SYBR Green QPCR mix (Stratagene, La Jolla, CA) in the Mx3000p thermocycler (Stratagene). A relative amount for each gene examined was obtained from a standard curve generated by plotting the cycle threshold value against the concentration of a serially diluted RNA sample expressing the gene of interest. This amount was normalized to the level of GAPDH RNA. Primer sequences (Chong et al. 2006; Denning et al. 2007; Villarino et al. 2008; Peyrin-Biroulet et al. 2010; Aguilera et al. 2011; Vaishnava et al. 2011) are listed in the Table1.
Table 1.
Primer sequences of real time PCR
| Primers for qPCR | Sequences |
|---|---|
| TNF-α | 5′-AATGGCCTCCCTCTCATCAGTT-3′ |
| 5′-CCACTTGGTGGTTTGCTACGA-3′ | |
| IFN-γ | 5′-GGCTGTCCCTGAAAGAAAGC-3′ |
| 5′-GAGCGAGTTATTTGTCATTCGG-3′ | |
| IL-23 | 5′-CATGGGGCTATCAGGGAGTA-3′ |
| 5′-GACCCACAAGGACTCAAGGA-3′ | |
| IL-17 | 5′-TTTAACTCCCTTGGCGCAAAA-3′ |
| 5′- CTTTCCCTCCGCATTGACAC-3′ | |
| IL-12p35 | 5′-GGAGAAGCAGACCCTTACAGAGTG-3′ |
| 5′-CTCAGATAGCCCATCACCCTG-3′ | |
| IL-12p40 | 5′-AAAGGCTGGGTATCGGTGG-3′ |
| 5′-ACTGGCTGTGCTGGAACTCC-3′ | |
| RegIIIγ | 5′-TTCCTGTCCTCCATGATCAAA-3′ |
| 5′-CATCCACCTCTGTTGGGTTC-3′ | |
| Defensin-1 | 5′-CCAGATGGAGCCAGGTGTTG-3′ |
| 5′-AGCTGGAGCGGAGACAGAATCC-3′ | |
| Defensin-3 | 5′-GCATTGGCAACACTCGTCAGA-3′ |
| 5′-CGGGATCTTGGTCTTCTCTA-3′ | |
| Defensin-10 | 5′-TTGTCCTGGTAATAGCAGGTTTATGA-3′ |
| 5′-CGGAGATTCTCTGGGTGACAGT-3′ | |
| GAPDH | 5′-GGTGGGTGGTCCAAGGTTTC-3′ |
| 5′-TGGTTTGACAATGAATACGGCTAC-3′ |
Western Blot (WB) and immunofluorescence (IF)
Total cellular protein (TP), cytosolic protein (CE), and nuclear protein (NE) extracts were respectively prepared from colonic mucosa using cold RIPA buffer and the NE-PER kit per the manufacturers' recommendations (Pierce, Rockford, IL). The nuclear abundance of p65 was detected in NE, SH-PTP1 was used as internal control for loading NE. IκBα, and β were determined in CE, β-tubulin was used as internal control for loading CE. Band intensities were quantified as mean area density using ImageQuant (Molecular Dynamics, Sunnyvale, CA). p65 (phosphoserine 276) was examined in paraffin-embedded intestinal sections using VECTASTAIN Elite ABC system (Vector lab, Burlingame, CA). BrdU staining followed manufacturer's instructions (BrdU In-Situ Detection Kit, BD Pharmingen, San Jose, CA) (Han et al. 2010). BrdU, Cleaved caspase-3, Ki67, and p65-positive cells were counted by a semiquantitative method and expressed as average positive cells per crypt. Images were captured using a Zeiss microscope and Axioviewer image analysis software (Carl Zeiss Corp, Germany) (Han et al. 2005, 2009).
Isolation of lamina propria mononuclear cells (LPMC) and flow cytometry (FACS)
Fresh LPMC were dissociated with EDTA and Collagen IV from human surgical specimens or mouse intestines (Munitz et al. 2010). LPMCs were stained for surface markers CD3, CD4, CD11b. The fluorescent signals were measured by FACS (LSR II, BD Biosciences, San Jose, CA, USA). CD3+ or CD11b+/7-AAD-/AnnexinV was used as a marker for apoptotic cells (BD Biosciences). Data were analyzed using FlowJo software (Tree Star, OR) (Han et al. 2009).
Human colonic biopsy organ culture
Colon biopsy samples were captured from noninflamed and inflamed areas in the collected surgical specimens. The pieces of tissue were washed with cold PBS and trimmed into explants. Explants were cultured in serum-free CMRL-1066 tissue culture medium (Gibco, Grandland, NY) on collagen I-coated plates (BioCoat; Becton Dickinson, Bedford, MA) at 37°C in a 95% oxygen/5% carbon dioxide atmosphere for 24 h. The morphological appearance of the explants was preserved under these conditions as assessed by light microscopy. After 24 h in culture, explants were treated with MPD (100 ng/mL) for another 12 h, and nuclear protein was prepared for NF-κB p65 immunoblotting (Han et al. 2005).
Enteroid culture and differentiation
Intestinal crypts were isolated from mice, and then dissociated with Chelation Buffer (1 mmol/L EDTA, 5 mmol/L EGTA, 0.5 mmol/L DTT, 43.3 mmol/L Sucrose, and 54.9 mmol/L Sorbitol). The crypts were filtered and re-suspended in Matrigel with 50 ng/mL EGF, 100 ng/mL Noggin, and 500 ng/mL R-spondin. Intestinal intact crypts were cultured and in vitro differentiated from day 1 to day 14 as published (Sato et al. 2009; Spence et al. 2011). Media was replaced once every 4 days. Different doses of MPD (0, 1, 10, or 100 ng/mL) were used to observe enteroid crypt formation. The crypt budding process was imaged and recorded with an inverted microscope (Olympus TH4-100, Tokyo, Japan).
IEC monolayer permeability assay
For subsequent permeability assays (Han et al. 2003; Wang et al. 2005), we used 17-day postconfluent Caco-2 monolayers. The medium bathing the apical surface of the monolayers was replaced with 200 μL of DMEM complete medium containing FITC-Dextran (FD4, 4 kDa, Sigma, St. Louis, MO, USA) at 25 mg/mL. The medium bathing the basolateral side of the monolayers was replaced with 500 μL of DMEM complete medium alone or DMEM supplemented with or without IFNγ (10 ng/mL) induction for 12 h and then stimulation with TNFα (10 ng/mL) respectively for 8 h followed by MPD treatment (100 ng/mL). Fluorescence in basolateral bathing medium was measured using a fluorescence spectrophotometer (Biotek Instruments, Winooski, VT). The permeability of the monolayer was expressed as a clearance (C; nL·cm−2·min−1). Transepithelial electrical resistance (TEER) was measured by E-VOM instrument (World Precision Instruments, Sarasota, FL). Results were expressed as Ohm·cm2.
Statistical analysis
Results are presented as the mean ± SEM. Data were analyzed using analysis of variance (ANOVA), 2-tailed Student's t-test, and the Mann–Whitney test as appropriate (Prism, GraphPad, San Diego, CA). Kaplan–Meier curve was used to estimate the percent survival. P values ≤0.05 were considered significant.
Results
MPD treatment mitigates intestinal inflammation, leading to increased mouse survival from lethal colitis
DSS administration results in intestinal inflammation and the formation of colitic lesions in mice, which resembles human UC pathology (Okayasu et al. 1990). To investigate the potential applications of MPD for human mucosal inflammation, intestinal inflammation in C57BL/6 mice was induced by exposing them to 2.5% DSS challenge for either 7 days for acute studies or 7 days followed by 5 days of water for healing studies. We first determined the therapeutic effects of MPD upon acute intestinal injury or mucosal recovery. We i.p. gave mice MPD (25 mg kg−1 day−1) on the day 3 of DSS administration for 7 or 10 days, respectively, to observe the effect of MPD upon DSS-induced acute injury or mucosal healing (Fig.1A). We found that 5-day MPD treatment did not ameliorate disease severity at a dose of 25 mg/kg daily (Fig.1B and E). In contrast, 10-day MPD treatment significantly improved the disease severity compared to the DSS colitis group, characterized by increased percentage of weight gain (Fig.1C), increased colon length (Fig.1D) and reduced histological scores (Fig.1E). Consistently, MDP can maintain the therapeutic effects to promote mucosal healing (Fig.1E). These data indicate that MPD treatment reduces mucosal inflammation to promote mucosal healing. We thus focus on MPD preventive and therapeutic effects upon intestinal inflammation. In order to provoke clinical signs of colitis, we induced colitis with a high dose of DSS (4%) in C57BL/6 mice. There was 75% mortality after 7 day-DSS administration (Fig.1F). In parallel, we treated another group with 40 mg/kg/day MPD for 5 consecutive days (Fig.1A and E), a dose which has shown the least toxicity to rat (Cao et al. 2010). We found 37.5% mortality, indicating that MPD treatment can prevent the high-dose DSS-induced mortality (P < 0.01) (Fig.1F). Taken together, MPD treatment can ameliorate mucosal inflammation to increase mouse survival from severe colitis.
MPD treatment reduces NF-κB activation in the inflamed intestinal mucosa to diminish the secretion of proinflammatory cytokines
The inflammation in human IBD and murine models of IBD is likely to depend, at least in part, on the activation and nuclear translocation of NF-κB family members (Neurath et al. 1998; Schreiber et al. 1998). To determine whether MPD can inhibit the activation of NF-κB to ameliorate mucosal inflammation, we collected surgical intestinal specimen from patients with pediatric UC. Some biopsies were captured from relatively normal mucosa in colon, others from areas of inflamed mucosa defined by the gross appearance. An organ culture model was employed to test whether MPD incubation can attenuate the upregulated NF-κB activation. We found that NF-κB activation was highly upregulated in the inflamed areas, compared to the relatively normal ileal and colonic mucosa (Fig.2A). MPD at 200 ng/mL significantly reduced RelA/p65 nuclear abundance in inflamed colonic mucosal biopsies after 24 h incubation (Fig.2A), indicating that MPD treatment reduced NF-κB activation.
Figure 2.
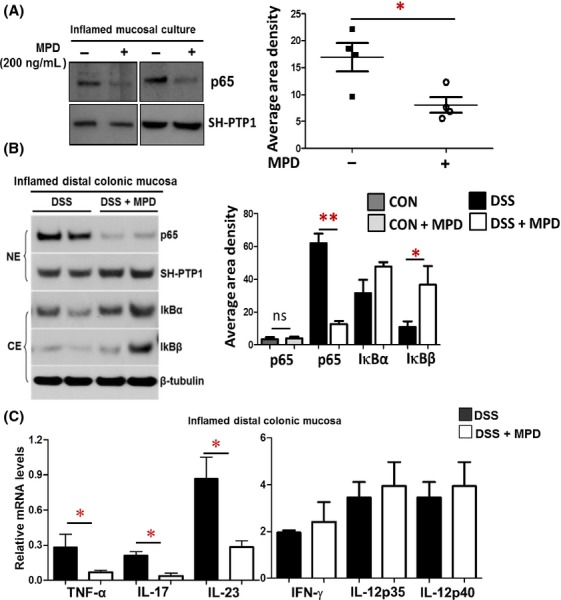
MPD treatment reduces NF-κB activation and proinflammatory cytokine production in the inflamed intestinal mucosa. (A) Surgical intestinal specimens were collected from four different patients with pediatric UC. Biopsies were captured from relatively normal colonic mucosa as control group, others from areas of inflamed mucosa as inflamed group. These biopsies were used for organ culture in the presence or absence of MPD incubation. Nuclear proteins were extracted from cultured explants after 24-h MPD treatment, n = 4. (B) Inflamed colonic mucosa was isolated from DSS-induced colitis with or without MPD treatment, and nuclear or cytosolic proteins were extracted. p65 nuclear abundance and IκBα & β cytosolic abundance were determined, n = 5, NE: nuclear protein. CE: cytosolic protein. Results are expressed as mean ± SEM, **P < 0.01 versus controls, ns, not significant. (C) RNA was extracted from inflamed colonic mucosa, levels of TNF-α, IL-17, IL-23, IFN-γ, IL-12p35 and IL-12p40 were determined with real-time PCR. Results are expressed as mean ± SEM (n = 5−6), *P < 0.05 versus controls.
In comparison with ex vivo patient-based studies, we separated colonic mucosa from DSS colitic mice with or without 10-day MPD treatment. The nuclear and cytosolic proteins were prepared as published previously (Gilbert et al. 2012). Immunoblotting analysis showed that MPD treatment significantly reduced NF-κB activation in colonic mucosa (Fig.2B). IκBa or β were demonstrated to deactivate NF-κB (Zandi et al. 1997). We consistently found that MPD significantly increased the cytosolic IκBα and β expression (Fig.2B). Therefore, MPD treatment mitigates the disease severity of colitis possibly through reducing NF-κB activation in colonic mucosa. It was known that various proinflammatory cytokines activate NF-κB in both transient and persistent phases (Schmidt et al. 2003). Persistent activated NF-κB upregulates the secretion of proinflammatory cytokines in intestinal mucosa that perpetuate the progressive mucosal inflammation to reduce mucosal healing in IBD (Schreiber et al. 1998; Fichtner-Feigl et al. 2005). Thus, we next measured pro-inflammatory cytokines (TNFα, IL-17, and IL-23) in colonic mucosa using quantitative real-time PCR (qPCR) that are particularly relevant with colonic mucosal inflammation and NF-κB activation (Xavier and Podolsky 2007). We found that 10-day MPD treatment significantly reduced TNFα, IL-17, and IL-23 production in colonic mucosa (Fig.3C), suggesting that MPD could reduce mucosal proinflammatory cytokine production to reduce persistent mucosal inflammation.
Figure 3.
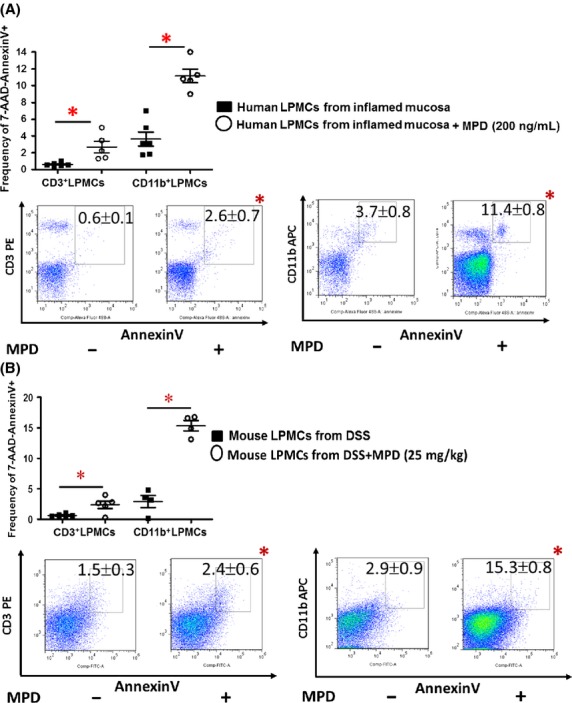
MPD treatment induces apoptosis of inflammatory laminar propria mononuclear cells (LPMC). (A) LPMCs were isolated from the collected surgical specimen from UC patients and cultured for 12 h in the presence or absence of MPD Incubation (200 ng/mL). Frequency of apoptosis in the CD3+ or CD11b+ LPMCs were determined with 7-AAD and Fluorochrome-labeled AnnexinV as percentage of 7-AAD-AnnexinV+. Representative scatter graphs are shown. (B) LMPCs were isolated from colonic tissues of colitic mice, apoptosis was determined as percentage of 7-AAD-AnnexinV+. Results are expressed as mean ± SEM (n = 5 or 6), *P < 0.05 versus controls. Representative scatter graphs are shown.
MPD treatment induces inflammatory cell apoptosis
The unrelenting mucosal inflammation of IBD is characterized by a persistently elevated number of activated lamina propria mononuclear cells (LPMC) including activated T cells and macrophages in the mucosa (Fiocchi 1998). These LPMCs secrete proinflammatory cytokines during mucosal inflammation to activate NF-κB, contributing to the pathogenesis of IBD (Strober et al. 2007). Accordingly, we tested whether MPD can induce the apoptosis of activated mucosal LPMCs, and further reduce mucosal NF-κB activation using patient materials. We isolated LPMCs in the collected surgical specimen from UC patients. We found that after 12 h-incubation, MPD induced a significant apoptosis of CD3+ and CD11b+ lamina propria cells at 200 ng/mL (Fig.3A). In comparison, we isolated LMPCs from DSS-treated colitic mice with or without MPD treatment to examine the effects of MPD upon LPMC apoptosis. Consistently, our in vivo experiments exhibited that MPD treatment at 25 mg/kg led to significantly increased apoptosis of CD11b+ LPMCs (Fig.3B), suggesting that MPD treatment could induce inflammatory cell apoptosis to inhibit mucosal inflammation. Taken together, MPD treatment could reduce mucosal proinflammatory cytokine production and NF-κB activation, leading to reduction in inflammatory cell survival that further mitigate colonic mucosal inflammation.
MPD administration promotes AMP expression and increased IEC innate function against bacterial translocation
We next tested if MPD treatment can increase IEC function to promote mucosal inflammation healing. We induced intestinal inflammation by exposing C57BL/6 mice to 2.5% DSS for 7 days followed by 5 days of water recovery. Starting on day 3, MPD treatment at 25 mg/kg was i.p. given to mice for 10 days. We found that MPD significantly increased β-defensin1 & 10, and RegIIIγ gene expression (Fig.4A), the number of goblet cells (Fig.4B), and IEC proliferation (Fig.4C and D) while reducing IEC apoptosis (Fig.4E). To confirm our observation, we examined the effect of MPD administration on bacterial translocation to MLN. We found that 10 day MPD treatment at 25 mg/kg significantly reduced the number of bacterial colonies in MLN (Fig.4F), suggesting that MPD might enhance intestinal barrier function. Together, these data suggest that MPD could promote IEC barrier function to directly protect intestinal epithelia from bacteria-induced mucosal inflammation, accelerating mucosal healing.
Figure 4.
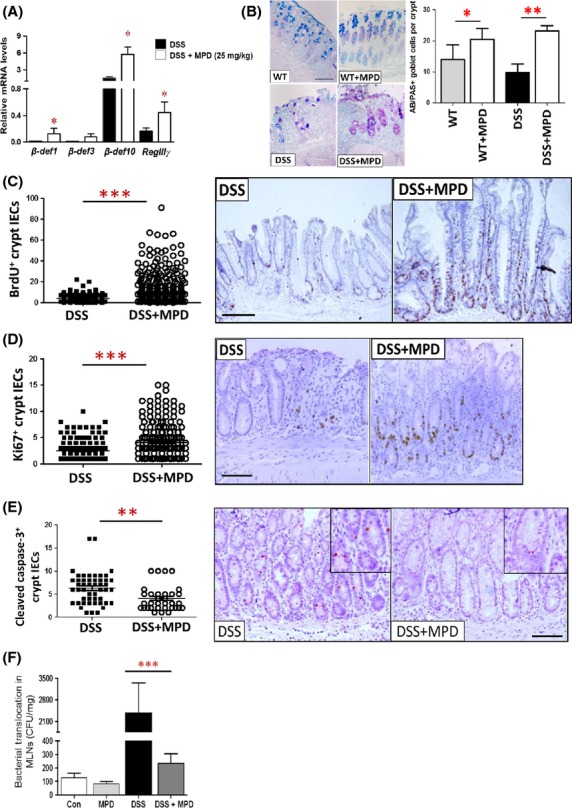
MPD administration promotes antimicrobial peptide expression and increases intestinal epithelial proliferation to enhance intestinal barrier function. Intestinal inflammation was induced by 2.5% DSS for 7 days followed by 5 days of water recovery. MPD was given to mice at 25 mg/kg daily on day 3 together with DSS for 10 days. (A) Colonic mucosal RNA was then extracted; the levels of AMP mRNA were quantitated by qPCR. *P < 0.01. (B) Goblet cells were stained with AB/PAS staining, and numbers of goblet cells were counted and expressed as AB/PAS+ cells per crypts. (C–E) IEC proliferation or apoptosis was determined with BrdU labeling (C) and Ki67 immunohistochemistry (D), or Cleaved Caspase-3 immunohistochemistry (E). Numbers of proliferative IEC or apoptotic IEC was counted in the well-orientated crypts. Results are expressed as mean ± SEM (n = 5 − 6). *P < 0.05, **P < 0.01 versus controls. (F) Intestinal inflammation was induced by 2.5% DSS for 7 days followed by 5 days of water. MPD was given to mice at 25 mg/kg daily on day 3 together with DSS for 10 days. Bacterial translocation to MLN was measured and expressed as colony-forming unit per mg lymphatic tissue. Results are expressed as mean ± SEM, n = 6 per group. ***P < 0.001 versus DSS colitis group, bar = 200 um.
MPD treatment promotes crypt formation and increases the IEC antimicrobial gene expression
Microbiota are known to shape the innate immune development in part by inducing IEC differentiation (Hooper et al. 2012). In turn, enhancing IEC differentiation into functional lineages could strengthen gut innate immunity to prevent from or protect against mucosal inflammation (Biteau et al. 2011). These enteroids provide a primary culture model used for studying IEC differentiation and regeneration (Sato et al. 2011; Liu et al. 2012). Therefore, to recapitulate our in vivo observation, we isolated mouse intact crypts and established crypt-derived enteroids. These enteroids were then incubated with MPD for 7 days to observe the effects of MPD upon crypt formation and differentiation with increasing doses (1, 10, 100 ng/mL). We found that MPD increased crypt expansion at 10 ng/mL (Fig.5A and B). However, higher dose of MPD (100 ng/mL) led to inhibited crypt budding (Fig.5A), suggesting that MPD could have a dose-dependent effect on increasing crypt formation. RNA was then extracted from cultured enteroides with or without MPD treatment; the levels of antimicrobial gene mRNA were quantitated by qPCR. We found that MPD elevated RegIIIγ mRNA expression (Fig.5C), suggesting that low dose of MPD could promote IEC differentiation into functional enterocytes. Together, MPD may increase the IEC functional differentiation to produce more antimicrobial peptide, interfering with microbial translocation.
Figure 5.
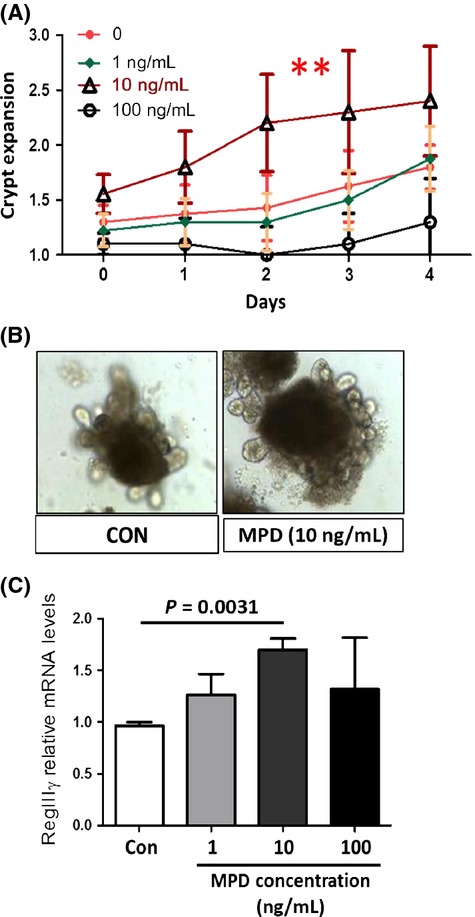
MPD incubation promotes intestinal crypt formation to up regulate the antimicrobial gene expression. Mouse small bowel intact crypts were isolated. Crypt-derived enteroids were cultured for 7 days, and then treated with MPD (1, 10, and 100 ng/mL) on day 2 for 5 days. (A and B) Number of crypt buds was counted daily in enteroids (n = 10). The results were expressed as a graph of crypt expansion showing the number of crypt buds versus time (A). One-way ANOVA is used to test for variance of two groups, *P < 0.05. Representative images were shown (B). (C) RNA was extracted and the levels of RegIIIγ mRNA were quantified by qPCR. Results are expressed as mean ± SEM, n = 10 buds each group, **P < 0.01 versus controls.
MPD treatment protects IEC barrier from proinflammatory cytokine-induced injury
Besides disturbed IEC lineage differentiation and antimicrobial activity in the immune-mediated colitis, the compromised TJ barrier in the differentiated IEC monolayers also plays a causal role in initiating colitic disease through broadly activating mucosal adaptive immunity (Blair et al. 2006; Su et al. 2009). To further pursue the mechanism of MPD action, we established a Caco-2 cell monolayer model, which is usually employed to test differentiated enterocyte maturation and barrier function (Pereira et al. 2013). We firstly induced Caco-2 monolayer barrier dysfunction by treating the monolayer with IFN-γ for 16 h followed by 8-h TNFα incubation (Gilbert et al. 2012). We then treated the Caco-2 monolayer with 100 ng/mL MPD for 12, 18, 24, and 48 h (Fig.6A). We found that MPD at 100 ng/mL significantly reduced the cytokine-induced hyperpermeability and restored the decrease in transepithelial resistance (TEER) caused by proinflammatory cytokines in this highly differentiated enterocyte model (Fig.6B). Thus, it is possible that MPD regenerates IEC barrier function to prevent luminal bacteria from crossing the IEC barrier to activate mucosal immune system. Taken together, MPD could also promote regeneration of the intestinal barrier to prevent luminal bacteria from penetrating, further ameliorating the disease severity of colitis.
Figure 6.
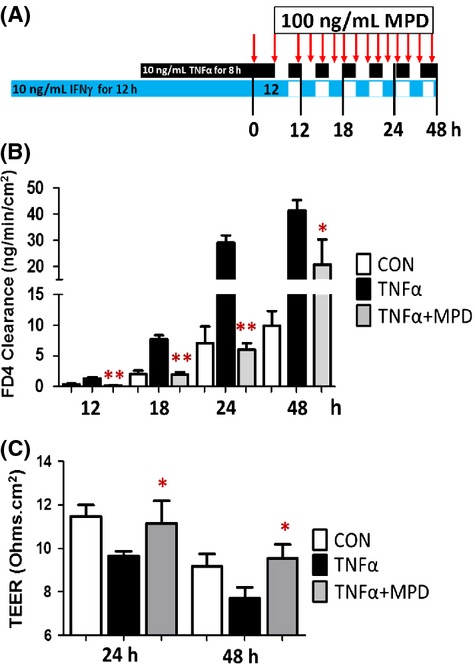
MPD treatment promotes intestinal barrier function. (A) 17 day postconfluent Caco-2 IEC monolayers, grown on Transwell filters, were challenged by IFNγ and TNFα followed by MPD (100 ng/mL) treatment. (B and C) Para-cellular permeability in monolayers was assessed as FD4 clearance and TEER. Results are expressed as mean ± SEM, n = 6 per group. *P < 0.05, **P < 0.01 versus TNFα group.
MPD treatment reduces disease severity of pathogen-induced colitis and bacterial translocation
Citrobacter rodentium (C. rodentium) is a murine-specific bacterial pathogen that causes attaching and effacing (A/E) lesion and distal colitis characterized by transmural inflammation, inflammatory cell infiltration and IEC hyperplasia (Mundy et al. 2005). We used this model to confirm the protective effects of MPD upon pathogen-induced mucosal inflammation. Mice were inoculated with C. rodentium (1 × 109) for 10 days; these mice were then administered MPD (25 mg/kg) on day 3 for 7 days. We found that 7-day MPD administration significantly restored weight loss (Fig.7A), increased colon length (Fig.7B and C), reduced disease severity of C. rodentium-induced colitis (Fig.7D and E), and reduces bacterial translocation in the MLN (Fig.7F). These data indicate that MPD can protect colonic mucosa from bacteria-induced colonic inflammation.
Figure 7.
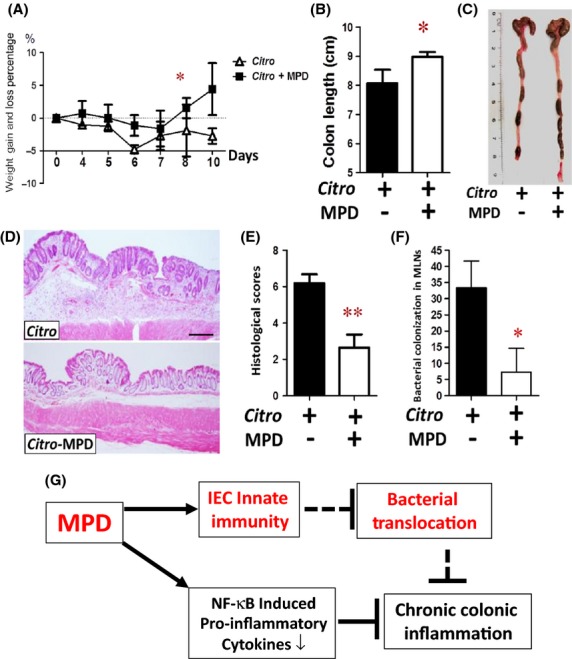
MPD treatment reduced the disease severity of C. rodentium-induced colitis. Eight-week-old C57BL/6 mice were orally inoculated with C. rodentium. Percentage of weight gain or loss was determined at the indicated time points (A), colon length (B) and number of stool pellets (C) were measured. (D) HE staining shows that MPD treatment reduced mucosal hyperplasia, diminished focal ulceration and transmural inflammation in the C. rodentium-treated C57BL/6 mice. (E) Combined colonic histological scores were determined in the MPD-treated colitic mice. (F) C. rodentium colonization in the MLNs was determined after 7-day MPD treatment. Results are expressed as colony-forming unit (CFU). Results are expressed as the mean ± SEM, n = 6, *P < 0.05, **P < 0.01 versus controls without C. rodentium infection. (G) Overview of MPD action mechanisms. On the one hand, MPD stimulates epithelial innate immunity; on the other hand, MPD inhibits the exaggerated mucosal adaptive immunity.
Discussion
Dioscoreaceae, a family of natural steroids with over 600 species (Ulbricht et al. 2003), has been recommended for the treatment of intestinal inflammation, rheumatic conditions, cancer, and gynecological symptoms in TCM over 2000 years (Komesaroff et al. 2001; Hu and Yao 2003; Ulbricht et al. 2003). MPD is a steroidal saponin that was isolated and refined from Dioscoreaceae (He et al. 2006). Recent investigations indicated that MPD improved experimental myocardial infarction (Ning et al. 2010). More importantly, MPD has a relatively high safety margin in toxicity, a clear PD/PK profile (Cao et al. 2008), and an identified chemical structure for further experimental studies (He et al. 2006). In our study, MPD administration protected against intestinal inflammation. Importantly, we found that in addition to inhibiting NF-κB activation, reducing proinflammatory cytokine expression, and inducing inflammatory cell apoptosis, MPD administration increased IEC proliferation, upregulated epithelial antimicrobial gene expression, and restored the impaired epithelial barrier function. These exciting data suggest a biphasic model of MPD action in which low doses of MPD can enhance epithelial innate immunity to promote mucosal healing; in contrast, high doses of MPD can control the exaggerated adaptive immune reaction, further keep inflammation conditions under control (Fig.7G).
IBD is due to a dysregulated mucosal innate immune response to the enteric flora, leading to a chronically activated adaptive response (Bouma and Strober 2003). Current effective biologic treatments targeting adaptive immune activation, inevitably lead to systemic immunosuppression and antimicrobial resistance (Andres and Friedman 1999). Natural products, including medicinal herbs, have provided approximately half of the drugs developed for clinical use over the past 20 years and they have low toxicity even if given in high quantity. As a complementary approach in the prevention of intestinal injury, there are no any reports that Dioscoreaceae causes systemic immunosuppression. Thus, we tested the effects of different doses of MPD upon experimental colitis. We found that a low dose of MPD (25 mg/mL) failed to prevent DSS-induced acute inflammation. In contrast, the preventive and therapeutic treatment with the same dose of MPD can protect mouse pathogen or chemical-induced colitis. Surprisingly, a high dose of MPD reduced colitis-induced mortality. These data suggest that it requires a continuous therapy to maintain the treatment efficacy of MPD in line with TCM, which adds Dioscoreaceae to food of affected patients with chronic inflammation. In this manuscript, we also focus on the potential mechanisms of preventive and therapeutic effects of MPD upon colitis.
Mucosal NF-κB activation leads to the increased production of cytokines in the intestinal mucosa that induce TJ barrier dysfunction and IEC apoptosis (Han et al. 2004; Al-Sadi et al. 2008; Boivin et al. 2009). Thus, we first studied whether MPD administration can attenuate intestinal mucosal inflammation through suppressing the aberrant activation of NF-κB in the inflamed intestinal mucosa. We combined ex vivo organ culture with mouse models where we found in both systems that MPD incubation can suppress activated NF-κB in the colonic samples from IBD patients. Our mouse experiment displayed that the preventive and therapeutic treatment of MPD reduced NF-κB activation, whereas it upregulated IκBα and β sub-units, which are the cytosolic inhibitors of NF-κB activation (Scott et al. 1993). Consistently, we found that MPD administration can significantly decrease levels of TNF-α, IL-17, and IL-23 cytokines. These data suggest that MPD can control inflammatory progress by directly reducing proinflammatory cytokine-induced NF-κB activation.
Well-established epithelial innate immune functions, such as antimicrobial peptides, epithelial barrier, and bacterial sensing receptors, temporally and spatially regulate the intestine homeostatic response to enteric microbiota and protect against pathogen injury (Vaishnava et al. 2011; Peterson and Artis 2014). Therefore, the therapeutic interventions for preserving intestinal innate immune homeostasis could avoid the negative side effects of exaggerated adaptive immune-induced tissue injury. We found that in contrast to blocking inflammatory pathways, MPD preventive and therapeutic treatment stimulated the secretion of anti-microbial peptides, β-defensin1, β-defensin10, and RegIIIγ, goblet cell differentiation, and epithelial proliferation. These enhanced innate immune functions protected intestinal epithelia from apoptosis, further promoting mucosal healing. Consequently, MPD treatment reduced bacterial translocation to MLNs. These effects of MPD therapy suggest that MPD can enhance the IEC innate function in addition to direct suppressive effects upon inflammatory pathway.
We further tested the potential mechanisms that MPD enhanced the IEC function. Crypt-derived enteroids were utilized to quantify intestinal stem cell proliferative capacity; they are also used as a model for drug discovery (Yin et al. 2014). Interestingly, we found that 10 ng/mL MPD can stimulate crypt budding that significantly increased RegIIIγ expression, indicating that MPD treatment promotes intestinal stem cell proliferation. However, MPD at 100 ng/mL significantly inhibited crypt formation. These data suggest the biphasic effects of MPD upon intestinal stem cell proliferation. Caco-2 monolayers are often used to evaluate the IEC barrier function and regeneration in response to inflammatory factors or infectious pathogens (Pollok et al. 2001; Gilbert et al. 2012). Using Caco-2 monolayers, we determined that MPD incubation can rescue inflammatory cytokine-induced monolayer disruption. Citrobacter rodentium, a murine model pathogen that shares important functional features with Enteropathogenic Escherichia coli (EPEC), colonizes mice in colon and cecum and causes barrier disruption and mucosal inflammation (Mundy et al. 2005; Flynn and Buret 2008). Taken together, MPD might reduce C. rodentium inflammation and bacterial colonization by enhancing IEC differentiation and barrier function.
Taken together, our study for the first time tests the therapeutic effects of MPD, a natural steroid extracted and refined from Dioscoreaceae upon IEC differentiation, gut infection and inflammation, thus providing unique anti-infectious and anti-inflammatory mechanisms for steroid-like herbal compounds. Our studies could lead to a new chemical entity for clinical trial with IBD patients.
Acknowledgments
We thank James Heubi and Mitchell Cohen at Cincinnati Children's Hospital Medical Center for generously supporting this project. This work was completed in part in NIH-funded digestive health center (D. H. C.). This work was supported by NIAID R21 (AI103388) (X. H.), CDMRP (PR121412) (X. H.), and NIH Clinical and Translational Research Award KL2 RR026315 (J. H. and X. H.). Cincinnati Children's Hospital Research Foundation Digestive Health Center (PHS Grant P30 DK078392). Thank Elizabeth A. Mann and Eleana Harmel-Laws for helping analyzing data.
Glossary
- AMP
antimicrobial peptides
- C. rodentium
Citrobacter rodentium
- CD
Crohn's disease
- DSS
dextran sulfate sodium
- IBD
inflammatory bowel diseases
- MPD
methyl protodioscin
- PK/PD
pharmacokinetic/pharmacodynamic
- TCH
traditional Chinese herbal
- TJ
tight junction
- UC
ulcerative colitis
Author Contributions
Participated in research design: XH and LD.
Conducted experiments: XH, SG, LD, KS, NS, RZ, MI.
Performed data analysis: XH, SG, LD, NS, JV, KS, RZ, RM, ME.
Contributed reagents/materials/analysis tools: RZ, LD, YS, NS, and CH.
Wrote or contributed to the writing of the manuscript: XH and LD.
Disclosure
None declared.
References
- Aguilera C, Nakagawa K, Sancho R, Chakraborty A, Hendrich B, Behrens A. c-Jun N-terminal phosphorylation antagonises recruitment of the Mbd3/NuRD repressor complex. Nature. 2011;469:231–235. doi: 10.1038/nature09607. [DOI] [PubMed] [Google Scholar]
- Al-Sadi R, Ye D, Dokladny K, Ma TY. Mechanism of IL-1beta-induced increase in intestinal epithelial tight junction permeability. J Immunol. 2008;180:5653–5661. doi: 10.4049/jimmunol.180.8.5653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andres PG, Friedman LS. Epidemiology and the natural course of inflammatory bowel disease. Gastroenterol Clin North Am. 1999;28:255–281. doi: 10.1016/s0889-8553(05)70056-x. , vii. [DOI] [PubMed] [Google Scholar]
- Biteau B, Hochmuth CE, Jasper H. Maintaining tissue homeostasis: dynamic control of somatic stem cell activity. Cell Stem Cell. 2011;9:402–411. doi: 10.1016/j.stem.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair SA, Kane SV, Clayburgh DR, Turner JR. Epithelial myosin light chain kinase expression and activity are upregulated in inflammatory bowel disease. Lab Invest. 2006;86:191–201. doi: 10.1038/labinvest.3700373. [DOI] [PubMed] [Google Scholar]
- Boivin MA, Roy PK, Bradley A, Kennedy JC, Rihani T, Ma TY. Mechanism of interferon-gamma-induced increase in T84 intestinal epithelial tight junction. J Interferon Cytokine Res. 2009;29:45–54. doi: 10.1089/jir.2008.0128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouma G, Strober W. The immunological and genetic basis of inflammatory bowel disease. Nat Rev Immunol. 2003;3:521–533. doi: 10.1038/nri1132. [DOI] [PubMed] [Google Scholar]
- Cao X, Yao Z, Chen H, Dai Y, Sun P, Ye W, et al. Development and validation of a liquid chromatography/tandem mass spectrometry assay for the quantification of methyl protodioscin in rat plasma: application to a pharmacokinetic study. Biomed Chromatogr. 2008;22:408–413. doi: 10.1002/bmc.948. [DOI] [PubMed] [Google Scholar]
- Cao X, Yao Z, Shao M, Chen H, Ye W, Yao X. Pharmacokinetics of methyl protodioscin in rats. Pharmazie. 2010;65:359–362. [PubMed] [Google Scholar]
- Cheng MS, Wang QL, Tian Q, Song HY, Liu YX, Li Q, et al. Total synthesis of methyl protodioscin: a potent agent with antitumor activity. J Org Chem. 2003;68:3658–3662. doi: 10.1021/jo020683w. [DOI] [PubMed] [Google Scholar]
- Chong KT, Xiang L, Wang X, Jun EL, Xi LF, Schweinfurth JM. High level expression of human epithelial beta-defensins (hBD-1, 2 and 3) in papillomavirus induced lesions. Virol J. 2006;3:75. doi: 10.1186/1743-422X-3-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper HS, Murthy SN, Shah RS, Sedergran DJ. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Labor Invest. 1993;69:238–249. [PubMed] [Google Scholar]
- Denning TL, Wang YC, Patel SR, Williams IR, Pulendran B. Lamina propria macrophages and dendritic cells differentially induce regulatory and interleukin 17-producing T cell responses. Nat Immunol. 2007;8:1086–1094. doi: 10.1038/ni1511. [DOI] [PubMed] [Google Scholar]
- Fichtner-Feigl S, Fuss IJ, Preiss JC, Strober W, Kitani A. Treatment of murine Th1- and Th2-mediated inflammatory bowel disease with NF-kappa B decoy oligonucleotides. J Clin Investig. 2005;115:3057–3071. doi: 10.1172/JCI24792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiocchi C. Inflammatory bowel disease: etiology and pathogenesis. Gastroenterology. 1998;115:182–205. doi: 10.1016/s0016-5085(98)70381-6. [DOI] [PubMed] [Google Scholar]
- Flynn AN, Buret AG. Tight junctional disruption and apoptosis in an in vitro model of Citrobacter rodentium infection. Microb Pathog. 2008;45:98–104. doi: 10.1016/j.micpath.2007.12.004. [DOI] [PubMed] [Google Scholar]
- Gilbert S, Zhang R, Denson L, Moriggl R, Steinbrecher K, Shroyer N, et al. Enterocyte STAT5 promotes mucosal wound healing via suppression of myosin light chain kinase-mediated loss of barrier function and inflammation. EMBO Mol Med. 2012;4:109–124. doi: 10.1002/emmm.201100192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grossman AB, Baldassano RN. Specific considerations in the treatment of pediatric inflammatory bowel disease. Expert Rev Gastroenterol Hepatol. 2008;2:105–124. doi: 10.1586/17474124.2.1.105. [DOI] [PubMed] [Google Scholar]
- Han X, Uchiyama T, Sappington PL, Yaguchi A, Yang R, Fink MP, et al. NAD+ ameliorates inflammation-induced epithelial barrier dysfunction in cultured enterocytes and mouse ileal mucosa. J Pharmacol Exp Ther. 2003;307:443–449. doi: 10.1124/jpet.103.056556. [DOI] [PubMed] [Google Scholar]
- Han X, Fink MP, Yang R, Delude RL. Increased iNOS activity is essential for intestinal epithelial tight junction dysfunction in endotoxemic mice. Shock. 2004;21:261–270. doi: 10.1097/01.shk.0000112346.38599.10. [DOI] [PubMed] [Google Scholar]
- Han X, Sosnowska D, Bonkowski EL, Denson LA. Growth hormone inhibits signal transducer and activator of transcription 3 activation and reduces disease activity in murine colitis. Gastroenterology. 2005;129:185–203. doi: 10.1053/j.gastro.2005.05.018. [DOI] [PubMed] [Google Scholar]
- Han X, Ren X, Jurickova I, Groschwitz K, Pasternak BA, Xu H, et al. Regulation of intestinal barrier function by signal transducer and activator of transcription 5b. Gut. 2009;58:49–58. doi: 10.1136/gut.2007.145094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han X, Gilbert S, Groschwitz K, Hogan S, Jurickova I, Trapnell B, et al. Loss of GM-CSF signalling in non-haematopoietic cells increases NSAID ileal injury. Gut. 2010;59:1066–1078. doi: 10.1136/gut.2009.203893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanauer SB. Risks and benefits of combining immunosuppressives and biological agents in inflammatory bowel disease: is the synergy worth the risk? Gut. 2007;56:1181–1183. doi: 10.1136/gut.2006.115980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He X, Qiao A, Wang X, Liu B, Jiang M, Su L, et al. Structural identification of methyl protodioscin metabolites in rats' urine and their antiproliferative activities against human tumor cell lines. Steroids. 2006;71:828–833. doi: 10.1016/j.steroids.2006.05.013. [DOI] [PubMed] [Google Scholar]
- Hilsden RJ, Scott CM, Verhoef MJ. Complementary medicine use by patients with inflammatory bowel disease. Am J Gastroenterol. 1998;93:697–701. doi: 10.1111/j.1572-0241.1998.208_a.x. [DOI] [PubMed] [Google Scholar]
- Hogan SP, Seidu L, Blanchard C, Groschwitz K, Mishra A, Karow ML, et al. Resistin-like molecule beta regulates innate colonic function: barrier integrity and inflammation susceptibility. J Allergy Clin Immunol. 2006;118:257–268. doi: 10.1016/j.jaci.2006.04.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science. 2012;336:1268–1273. doi: 10.1126/science.1223490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu K, Yao X. The cytotoxicity of methyl protodioscin against human cancer cell lines in vitro. Cancer Invest. 2003;21:389–393. doi: 10.1081/cnv-120018230. [DOI] [PubMed] [Google Scholar]
- Hu K, Dong A, Yao X, Kobayashi H, Iwasaki S. Antineoplastic agents. II. Four furostanol glycosides from rhizomes of Dioscorea collettii var. hypoglauca. Planta Med. 1997;63:161–165. doi: 10.1055/s-2006-957636. [DOI] [PubMed] [Google Scholar]
- Komesaroff PA, Black CV, Cable V, Sudhir K. Effects of wild yam extract on menopausal symptoms, lipids and sex hormones in healthy menopausal women. Climacteric. 2001;4:144–150. [PubMed] [Google Scholar]
- Langmead L, Rampton DS. Review article: complementary and alternative therapies for inflammatory bowel disease. Aliment Pharmacol Ther. 2006;23:341–349. doi: 10.1111/j.1365-2036.2006.02761.x. [DOI] [PubMed] [Google Scholar]
- Liu MJ, Yue PY, Wang Z, Wong RN. Methyl protodioscin induces G2/M arrest and apoptosis in K562 cells with the hyperpolarization of mitochondria. Cancer Lett. 2005;224:229–241. doi: 10.1016/j.canlet.2004.11.051. [DOI] [PubMed] [Google Scholar]
- Liu J, Walker NM, Cook MT, Ootani A, Clarke LL. Functional Cftr in crypt epithelium of organotypic enteroid cultures from murine small intestine. Am J Physiol Cell Physiol. 2012;302:C1492–C1503. doi: 10.1152/ajpcell.00392.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maloy KJ, Powrie F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature. 2011;474:298–306. doi: 10.1038/nature10208. [DOI] [PubMed] [Google Scholar]
- Mann EA, Harmel-Laws E, Cohen MB, Steinbrecher KA. Guanylate cyclase C limits systemic dissemination of a murine enteric pathogen. BMC Gastroenterol. 2013;13:135. doi: 10.1186/1471-230X-13-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medzhitov R. Recognition of microorganisms and activation of the immune response. Nature. 2007;449:819–826. doi: 10.1038/nature06246. [DOI] [PubMed] [Google Scholar]
- Mundy R, MacDonald TT, Dougan G, Frankel G, Wiles S. Citrobacter rodentium of mice and man. Cell Microbiol. 2005;7:1697–1706. doi: 10.1111/j.1462-5822.2005.00625.x. [DOI] [PubMed] [Google Scholar]
- Munitz A, Cole ET, Beichler A, Groschwitz K, Ahrens R, Steinbrecher K, et al. Paired immunoglobulin-like receptor B (PIR-B) negatively regulates macrophage activation in experimental colitis. Gastroenterology. 2010;139:530–541. doi: 10.1053/j.gastro.2010.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neurath MF, Fuss I, Schurmann G, Pettersson S, Arnold K, Muller-Lobeck H, et al. Cytokine gene transcription by NF-kappa B family members in patients with inflammatory bowel disease. Ann N Y Acad Sci. 1998;859:149–159. doi: 10.1111/j.1749-6632.1998.tb11119.x. [DOI] [PubMed] [Google Scholar]
- Ning Z, Li Y, Zhang R. [Effects of methyl protodioscin on [Ca2+]i and ATPase activity in cardiomyocytes and analysis of mechanisms] China journal of Chinese materia medica. 2010;35:80–83. [PubMed] [Google Scholar]
- Okayasu I, Hatakeyama S, Yamada M, Ohkusa T, Inagaki Y, Nakaya R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology. 1990;98:694–702. doi: 10.1016/0016-5085(90)90290-h. [DOI] [PubMed] [Google Scholar]
- Pereira B, Sousa S, Barros R, Carreto L, Oliveira P, Oliveira C, et al. CDX2 regulation by the RNA-binding protein MEX3A: impact on intestinal differentiation and stemness. Nucleic Acids Res. 2013;41:3986–3999. doi: 10.1093/nar/gkt087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peterson LW, Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol. 2014;14:141–153. doi: 10.1038/nri3608. [DOI] [PubMed] [Google Scholar]
- Peyrin-Biroulet L, Beisner J, Wang G, Nuding S, Oommen ST, Kelly D, et al. Peroxisome proliferator-activated receptor gamma activation is required for maintenance of innate antimicrobial immunity in the colon. Proc Natl Acad Sci USA. 2010;107:8772–8777. doi: 10.1073/pnas.0905745107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollok RC, Farthing MJ, Bajaj-Elliott M, Sanderson IR, McDonald V. Interferon gamma induces enterocyte resistance against infection by the intracellular pathogen Cryptosporidium parvum. Gastroenterology. 2001;120:99–107. doi: 10.1053/gast.2001.20907. [DOI] [PubMed] [Google Scholar]
- Salzman NH, Hung K, Haribhai D, Chu H, Karlsson-Sjoberg J, Amir E, et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nat Immunol. 2010;11:76–83. doi: 10.1038/ni.1825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato T, Vries RG, Snippert HJ, van de Wetering M, Barker N, Stange DE, et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 2009;459:262–265. doi: 10.1038/nature07935. [DOI] [PubMed] [Google Scholar]
- Sato T, Stange DE, Ferrante M, Vries RG, vanEs JH, vanden Brink S, et al. Long-term Expansion of Epithelial Organoids From Human Colon, Adenoma, Adenocarcinoma, and Barrett's Epithelium. Gastroenterology. 2011;141:1762–1772. doi: 10.1053/j.gastro.2011.07.050. [DOI] [PubMed] [Google Scholar]
- Schmidt C, Peng B, Li Z, Sclabas GM, Fujioka S, Niu J, et al. Mechanisms of proinflammatory cytokine-induced biphasic NF-kappaB activation. Mol Cell. 2003;12:1287–1300. doi: 10.1016/s1097-2765(03)00390-3. [DOI] [PubMed] [Google Scholar]
- Schreiber S, Nikolaus S, Hampe J. Activation of nuclear factor kappa B inflammatory bowel disease. Gut. 1998;42:477–484. doi: 10.1136/gut.42.4.477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott ML, Fujita T, Liou HC, Nolan GP, Baltimore D. The p65 subunit of NF-kappa B regulates I kappa B by two distinct mechanisms. Genes Dev. 1993;7:1266–1276. doi: 10.1101/gad.7.7a.1266. [DOI] [PubMed] [Google Scholar]
- Spence JR, Mayhew CN, Rankin SA, Kuhar MF, Vallance JE, Tolle K, et al. Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature. 2011;470:105–109. doi: 10.1038/nature09691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strober W, Fuss I, Mannon P. The fundamental basis of inflammatory bowel disease. J Clin Investig. 2007;117:514–521. doi: 10.1172/JCI30587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su L, Shen L, Clayburgh DR, Nalle SC, Sullivan EA, Meddings JB, et al. Targeted epithelial tight junction dysfunction causes immune activation and contributes to development of experimental colitis. Gastroenterology. 2009;136:551–563. doi: 10.1053/j.gastro.2008.10.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tak PP, Firestein GS. NF-kappaB: a key role in inflammatory diseases. J Clin Investig. 2001;107:7–11. doi: 10.1172/JCI11830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner JR. Intestinal mucosal barrier function in health and disease. Nat Rev Immunol. 2009;9:799–809. doi: 10.1038/nri2653. [DOI] [PubMed] [Google Scholar]
- Ulbricht C, Basch E, Ulbricht C, Sollars D, Hammerness P, Hashmi S. Wild yam (Dioscoreaceae) J Herb Pharmacother. 2003;3:77–91. [PubMed] [Google Scholar]
- Vaishnava S, Yamamoto M, Severson KM, Ruhn KA, Yu X, Koren O, et al. The antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the intestine. Science. 2011;334:255–258. doi: 10.1126/science.1209791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villarino AV, Artis D, Bezbradica JS, Miller O, Saris CJ, Joyce S, et al. IL-27R deficiency delays the onset of colitis and protects from helminth-induced pathology in a model of chronic IBD. Int Immunol. 2008;20:739–752. doi: 10.1093/intimm/dxn032. [DOI] [PubMed] [Google Scholar]
- Wang F, Graham WV, Wang Y, Witkowski ED, Schwarz BT, Turner JR. Interferon-gamma and tumor necrosis factor-alpha synergize to induce intestinal epithelial barrier dysfunction by up-regulating myosin light chain kinase expression. Am J Pathol. 2005;166:409–419. doi: 10.1016/s0002-9440(10)62264-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflammatory bowel disease. Nature. 2007;448:427–434. doi: 10.1038/nature06005. [DOI] [PubMed] [Google Scholar]
- Yin J, Tezuka Y, Kouda K, Le Tran Q, Miyahara T, Chen Y, et al. In vivo antiosteoporotic activity of a fraction of Dioscorea spongiosa and its constituent, 22-O-methylprotodioscin. Planta Med. 2004;70:220–226. doi: 10.1055/s-2004-815538. [DOI] [PubMed] [Google Scholar]
- Yin X, Farin HF, van Es JH, Clevers H, Langer R, Karp JM. Niche-independent high-purity cultures of Lgr5+ intestinal stem cells and their progeny. Nat Methods. 2014;11:106–112. doi: 10.1038/nmeth.2737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zandi E, Rothwarf DM, Delhase M, Hayakawa M, Karin M. The IkappaB kinase complex (IKK) contains two kinase subunits, IKKalpha and IKKbeta, necessary for IkappaB phosphorylation and NF-kappaB activation. Cell. 1997;91:243–252. doi: 10.1016/s0092-8674(00)80406-7. [DOI] [PubMed] [Google Scholar]