

Abstract
Since BRCA1 associated breast cancers are frequently classified as hormone receptor negative or even triple negative, the application of endocrine therapies is rather limited in these patients. Like hormone receptors that bind to estrogen or progesterone, thyroid hormone receptors (TRs) are members of the nuclear hormone receptor superfamily. TRs might be interesting biomarkers - especially in the absence of classical hormone receptors. The current study aimed to investigate whether TRs may be specifically expressed in BRCA1 associated cancer cases and whether they are of prognostic significance in these patients as compared to sporadic breast cancer cases. This study analyzed TRα and TRβ immunopositivity in BRCA1 associated (n = 38) and sporadic breast cancer (n = 86). Further, TRs were studied in MCF7 (BRCA1 wildtype) and HCC3153 (BRCA1 mutated) cells. TRβ positivity rate was significantly higher in BRCA1 associated as compared to sporadic breast cancers (p = 0.001). The latter observation remained to be significant when cases that had been matched for clinicopathological criteria were compared (p = 0.037). Regarding BRCA1 associated breast cancer cases TRβ positivity turned out to be a positive prognostic factor for five-year (p = 0.007) and overall survival (p = 0.026) while TRα positivity predicted reduced five-year survival (p = 0.030). Activation of TRβ resulted in down-modulation of CTNNB1 while TRα inhibition reduced cell viability in HCC3153. However, only BRCA1 wildtype MCF7 cells were capable of rapidly degrading TRα1 in response to T3 stimulation. Significantly, this study identified TRβ to be up-regulated in BRCA1 associated breast cancer and revealed TRs to be associated with patients’ prognosis. TRs were also found to be expressed in triple negative BRCA1 associated breast cancer. Further studies need to be done in order to evaluate whether TRs may become interesting targets of endocrine therapeutic approaches, especially when tumors are triple-negative.
Introduction
Breast cancers diagnosed in patients carrying a BRCA1 germline mutation display distinct histo-pathological as well as molecular characteristics and have been observed to differ from sporadic cases also regarding chemotherapeutic sensitivity [1]. Unlike sporadic cases, BRCA1 associated breast cancers display a higher incidence of medullary or basal-like histology and might overexpress cell cycle stimulator genes [2–4]. The fact that BRCA1 associated breast cancers are mostly diagnosed as being negative for either classical hormone receptors (estrogen receptor (ER), progesterone receptor (PR)), for human epidermal growth factor receptor-2 (HER2) or for all of them (so called triple negative cases), is considered to be one of the most characteristic features [2,5]. As a consequence, BRCA1 associated breast cancers require a specially tailored therapeutic regimen, since the frequent lack of hormone receptors (ER/PR) or Her2 extensively narrows the application of (anti-)endocrine therapies [5].
Like classical steroid hormone receptors (ER/PR), which are routinely used as predictive markers, thyroid hormone receptors (TRs) are members of the nuclear hormone receptor superfamily acting via transcriptional cis-regulation of target genes. However, the exact role of thyroidal effector hormones and TRs in breast cancer remains still to be elucidated. Recently, expression of TRs has been identified in up to about 79% of breast cancer cases and furthermore TRs have been shown to be associated with clinicopathological parameters such as tumor size, grade, lymph node involvement and hormone receptor status [6]. In addition, TRs have been reported to regulate a plethora of genes including those being involved in mediating cell differentiation, proliferation and apoptosis [7,8] and were found to be predictive for patient prognosis in hepatocellular carcinoma [8]. Since patients’ TR statuses can be determined easily by immunohistochemistry and as TRs were demonstrated to be accessible targets [6,9], TRs might be novel alternative biomarkers, especially for hormone receptor negative or even triple negative breast cancer patients. With a high percentage of BRCA1 associated breast cancers being classified as triple-negative, the assessment of TR expression in those patients may turn out to be attractive in terms of alternative treatment options, applicable especially for breast cancer patients carrying a BRCA1 germline mutation.
So far, no report exists on immunhistochemical TR reactivity in BRCA1 associated breast cancer. Therefore, this study aimed to investigate the presence of TRs in breast cancers diagnosed in patients carrying a BRCA1 germline mutation as compared to sporadic (i.e. without any family or personal history of breast cancer) cases. Further, TR immunostaining was tested for association with clinicopathological parameters and patient survival in breast cancer samples obtained from BRCA1 carrier vs. sporadic cases. Breast cancer cell lines were used to determine whether TRs are active in the case of BRCA1 deficiency.
Patients and Methods
Patients and Specimen Characteristics
Breast cancer patients diagnosed with sporadic (n = 86) or BRCA1 associated breast cancer (n = 38) were included in this study. The majority of cases were diagnosed with a tumor of non-specific type (NST, n = 96, 77.4%), high grade (G3, n = 77, 62.6%) or staged higher than pT1 (n = 78, 62.9%). A significant number of patients also presented with either lymph node (n = 66, 57.9%) or distant metastasis (n = 70, 63.1%).
Formalin fixed paraffin embedded (FFPE) breast cancer tissue was collected from patients who had undergone surgery due to a malignant tumor of the breast, either without positive family history of breast cancer (sporadic cancer cases, n = 86) or with the diagnosis of carrying a BRCA1 germline mutation (n = 38). Breast cancer tissue was gained at surgery and underwent routine histopathological processing and examination.
Study Design
Patients were recruited at the Department of Gynecology and Obstetrics at the Ludwig-Maximilians-University of Munich, Germany between 1987 and 2009. Women only diagnosed for benign tumors of the breast or for in situ carcinoma were excluded from the study. Clinical as well as follow-up data were retrieved retrospectively from patients’ charts, from the Munich Cancer Registry or by direct contact. Overall mean survival of the cohort was 7.3 years (95% CI: 6.2–8.3 years) and mean follow up time was 6.6 years (95% CI: 5.7–7.5 years). The outcome assessed was patient five-year and overall survival. Mean age (± STDV) of the cohort was 50.0 ± 13.3 years (BRCA1 associated cases: 41.9 ± 10.8 years; sporadic breast cancer: 53.7 ± 12.8 years).
Ethical Approval
All patient data were fully anonymized and the study was performed according to the standards set in the declaration of Helsinki 1975. All tumor tissue used was left-over material that had initially been collected for histo-pathological diagnostics. All diagnostic procedures had already been fully completed when samples were retrieved for the study. The current study was approved by the Ethics Committee of the Ludwig-Maximilians-University, Munich, Germany (approval number 048-08). Authors were blinded from the clinical information during experimental analysis.
Assay Methods
Mutation Screening
Mutation screening was performed in a standardized manner at a German center for BRCA1 mutation testing (Technical University of Munich, Munich, Germany) as described by Fischer et al. [10]. In brief, PCR products comprising all coding exons of the BRCA1 gene were analyzed by high performance liquid chromatography (dHPLC) followed by sequencing of conspicuous amplicons or by direct sequencing of all BRCA1 amplicons. The NCBI (National Center for Biotechnology Information) cDNA sequence U14680.1 (BRCA1) served as a reference. Multiplex ligation-dependent probe amplification (MLPA) was used to screen for deletions or duplications in BRCA1 in case of negative sequencing results. So called variants of unknown significance (VUS) characterized as Class III were not considered as mutations.
Cell Culture
MCF7 (BRCA1 wt) and HCC3153 (BRCA1 943ins10) breast cancer cell lines were bought from the European Collection of Cell Cultures (MCF7) or were gently provided by Adi F. Gazdar (Hamon Center for Therapeutic Oncology Research and Department of Pathology, University of Texas Southwestern Medical Center, Dallas, TX) (HCC3153). While MCF7 carry a wildtype BRCA1 allele, HCC3153 show a homozygous insertion in exon eleven of BRCA1 (BRCA1 943ins10) leading to a premature stop codon and thus encoding a truncated BRCA1 protein [11]. Cells were cultured in DMEM (Biochrom, Berlin, Germany) containing stable glutamine and supplemented with 10% fetal bovine serum (FBS) without antibiotics/antimycotics. Mycoplasma testing was performed routinely.
Seeding densities were as follows: 24 well plate or 4-perm Chamberslide—7.5 x 104 cells per well, 96 well plate—3.0 x 103 cells per well (stimulation assays) and 6.0 x 103 cells per well (siRNA knockdown), Quadriperm—7.5 x 105 cells per well.
Immunostaining
Immunohistochemistry of TRα and TRβ on FFPE sections has been extensively described by our group [6,12]. In brief, rabbit polyclonal antibodies detecting TRα (Abcam, Cambridge, UK); Zytomed, Berlin, Germany) or TRβ (Zytomed)) were stained by employing commercially available kits (Vectastain Elite rabbit-IgG-Kit (VectorLabs, Burlingame, CA); ZytoChem Plus HRP Polymer System (Zytomed). Appropriate positive (struma and vaginal tissue [6]) and negative controls were included in each experiment (Fig 1). Tissue sections treated with pre-immune IgGs (supersensitive rabbit negative control, BioGenex, Fremont, CA) instead of the primary antibody served as negative controls. Immunoreactivity was quantified by applying a well-established semiquantitaive scoring system (IR-score; also known as Remmele’s score) by two independent observers by consensus. This scoring method has already been used in numerous studies [6,13–15] and quantifies immunoreactivity by multiplication of optical staining intensity (graded as 0: no, 1: weak, 2: moderate and 3: strong staining) and the percentage of positive stained cells (0: no staining, 1: ≤ 10% of the cells, 2: 11–50% of the cells, 3: 51–80% of the cells and 4: ≥ 81% of the cells). According to previously published data tissue samples that had been assigned an IRS higher than 2 were scored as positive [16,17].
Fig 1. TRα and TRβ immunostaining.
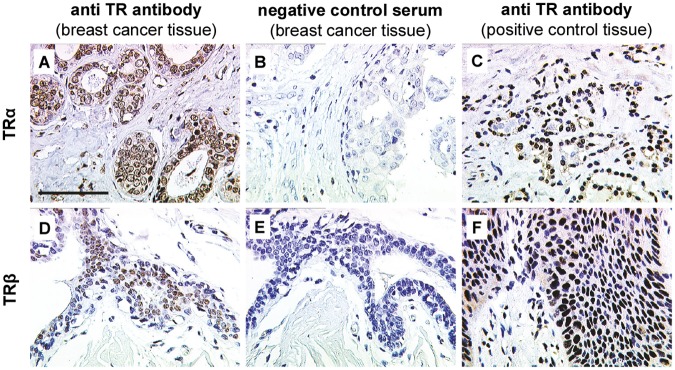
Positive TRα (A) and TRβ (D) staining was detected in breast cancer tissue. Negative (TRα (B) and TRβ (E)) and positive controls (TRα (C) and TRβ (F)) were performed to validate staining specificity. Thyroid gland and vaginal epithelium served as tissue positive controls for TRα (C) and TRβ (F), respectively. Scale bar represents 100 μm and applies to A-F. Representative photomicrographs are shown.
Double immunofluorescence of FFPE tissue sections was done as explained in [18] using TRα2 (Abcam) and TRβ (Zytomed. Berlin, Germany) as primary antibodies.
MCF7 (BRCA1 wt) and HCC3153 (BRCA1 943ins10) were grown on glass slides and fixed in 3.7% neutral buffered formaline (15 min). Following PBS washes and blocking in PBS 0.1% TritonX for one hour cells were stained by immunocytochemistry as described in [18] using TRα (diluted 1:200 in PBS) and TRβ (diluted 1:400 in PBS) antibodies.
For immunofluorescence cells were treated with T3 (10-7 M) for two hours under serum free conditions, processed as depicted above and stained for rabbit-anti-TRα1 (Abcam) or co-stained for rabbit-anti-TRα1 and for mouse-anti-ubiquitin (Enzo Life Sciences, Farmingdale, NY). Primary antibodies were diluted 1:100 and incubated overnight at 4°C. Secondary antibodies (goat-anti-rabbit-Cy2, goat-anti-mouse-Cy3; Jackson Immunolabs, West Grove, PA) were diluted 1:300 and incubated for 30 min at room temperature. Finally, samples were covered in Vectashield (VectorLabs) mounting medium containing DAPI and were imaged on a Leica confocal microscope (Leica TCS SP5 II, Leica, Wetzlar, Germany).
Western Blotting
Cells were seeded in 24 well plates and medium was changed to serum free DMEM two hours after plating. Cells were transfected using THRA-specific siRNA (si2, si3; both from Qiagen, Hilden, Germany) or scrambled control RNA (AllStars negative control, Qiagen) for 24 hours as per manufacturer’s protocol using HiPerFect (HiP) transfection reagent (Qiagen) [18] straight after changing to serum free DMEM or were stimulated with T3 (10-7 M for two hours) the next day. siRNAs targeting THRA (si2, si3, both from Qiagen, Hilden, Germany) were used. A scrambled siRNA (AllStars negative control, Qiagen) and samples treated with the transfection reagent only served as controls.
Protein samples were quantified, processed and blotted as explained elsewhere [18]. Antibodies detecting TRα1 and Histone H2B were from Abcam (Cambridge, UK) and diluted 1:500 in 2% marvel TBST. Histone H2B was used as a loading control.
Quantitative Real-time PCR
Total mRNA was isolated employing the NucleoSpin RNA II kit (Machery-Nagel, Düren, Germany). Having adjusted RNA concentrations cDNA synthesis was carried out as described elsewhere [19]. Gene expression of THRA, THRB and CTNNB1 was quantified by TaqMan real-time PCR and ACTB was used as a housekeeping gene. PCR conditions were: 20 s at 95°C and 40 cycles of 3 s (95°C) plus 30 s (60°C) employing the following primers (all from Applied Biosystems, Carlsbad, CA): THRA (Hs00268470_m1), THRB (Hs00230861_m1), CTNNB1 (Hs00170025_m1), ACTB (Hs99999903_m1). Differences in gene expression were calculated using the Rest2009 software [20] and graphs were built afterwards from Rest2009 outputs.
BrdU and WST-1 measurements
Cells were seeded in 96 well plates and medium was changed to DMEM w/o FBS two hours after plating. Cells were treated with 2-(2-(-(4-Nitrophenyl)-4-piperidinylidene)-acetyl-N-(3-(trifluoromethyl)phenyl)-1-hydrazine Carboxamide (1–850, Merck, Darmstadt, Germany) at concentrations of 10-5 M and 10-4 M. Controls treated with equal amounts of carrier solution (DMSO) served as controls. Cells were stimulated for six days before proliferation and viability were quantified by BrdU and WST-1 (both Roche, Penzberg, Germany), respectively. SiRNA transfections were performed as described before [18]. Cells were transfected in serum-free DMEM using siRNAs targeting THRA (si2, si3; both from Qiagen) while samples just treated with a scrambled siRNA (AllStars negative control, Qiagen) were included in each experiment. Cell viability was quantified by WST-1 six days after transfection.
Statistical Analysis Methods
This study has been carried out according to the REMARK (Reporting Recommendations for Tumor Marker Prognostic Studies) criteria [21].
The IBM statistic package SPSS (version 22) was used to test data for statistical significance. Fisher’s exact test and the Mann-Whitney test were used. Survival times were compared by Kaplan-Meier graphics and differences in patient overall survival times were tested for significance by using the chi-square statistics of the log rank test. Cell culture experiments were repeated three times achieving equal results and data were assumed to be statistically different in case of p < 0.05.
Statistical analyses were done in the whole sample as well as in a group of 56 patients (n (sporadic) = 28, n (BRCA1 associated) = 28) that had been matched (p = 1.000) according to tumor size, lymph node status, presence of metastasis and tumor grade.
Results
Study Cohort
Most cases investigated were diagnosed with invasive breast cancer of non-specific type (NST, n = 96, 77.4%) or of low differentiation (G3, n = 77, 62.6%). Patients carrying a BRCA1 mutation (n = 38) were diagnosed with high grade (G3, p = 0.025), pT1 (p = 0.008) and pN0 (p = 0.040) staged breast cancer significantly more often than sporadic cancer cases (n = 86). In addition, BRCA1 mutated and sporadic breast cancers differed regarding patient age (p < 0.001). With respect to hormone receptor expression, BRCA1 mutant carcinomas were significantly more frequently found to be ER negative (p = 0.001), PR negative (p = 0.001) or even triple negative (p = 0.002). Interestingly, overall survival rate of patients carrying a BRCA1 germline mutation was significantly higher than in sporadic breast cancer cases (p < 0.001). Patient characteristics and absolute numbers are listed in Table 1.
Table 1. Patient characteristics (whole sample).
| breast cancer type | BRCA1 | sporadic breast cancer | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BRCA1 | sporadic | TRα | TRβ | TRα | TRβ | ||||||||||
| p | negative | positive | p | negative | positive | p | negative | positive | p | negative | positive | p | |||
| Histology | |||||||||||||||
| non NST | 8 | 20 | ns | 4 | 4 | ns | 3 | 5 | ns | 9 | 11 | ns | 15 | 5 | ns |
| NST | 30 | 66 | 17 | 13 | 15 | 15 | 27 | 39 | 52 | 14 | |||||
| Grading | |||||||||||||||
| G1, G2 | 8 | 38 | 0.025 | 2 | 6 | ns | 5 | 3 | ns | 12 | 26 | ns | 28 | 10 | ns |
| G3 | 29 | 48 | 18 | 11 | 12 | 17 | 24 | 24 | 39 | 9 | |||||
| pT | |||||||||||||||
| pT1 | 21 | 25 | 0.008 | 12 | 9 | ns | 9 | 12 | ns | 14 | 11 | ns | 18 | 7 | ns |
| pT2-4 | 17 | 61 | 9 | 8 | 9 | 8 | 22 | 39 | 49 | 12 | |||||
| pN | |||||||||||||||
| pN0 | 20 | 28 | 0.040 | 13 | 7 | ns | 7 | 13 | ns | 13 | 15 | ns | 19 | 9 | ns |
| pN1-3 | 15 | 51 | 6 | 9 | 10 | 5 | 20 | 31 | 42 | 9 | |||||
| pM | |||||||||||||||
| pM0 | 18 | 23 | ns | 11 | 7 | ns | 7 | 11 | ns | 14 | 9 | 0.011 | 16 | 7 | ns |
| pM1 | 18 | 52 | 8 | 10 | 11 | 7 | 15 | 37 | 41 | 11 | |||||
| ER | |||||||||||||||
| negative | 27 | 27 | 0.001 | 14 | 13 | ns | 15 | 12 | ns | 12 | 15 | ns | 21 | 6 | ns |
| positive | 11 | 44 | 7 | 4 | 3 | 8 | 21 | 23 | 33 | 11 | |||||
| PR | |||||||||||||||
| negative | 27 | 25 | 0.001 | 14 | 13 | ns | 14 | 13 | ns | 13 | 12 | ns | 21 | 4 | ns |
| positive | 11 | 46 | 7 | 4 | 4 | 7 | 20 | 26 | 33 | 13 | |||||
| Her2 | |||||||||||||||
| negative | 22 | 29 | ns | 10 | 12 | ns | 10 | 12 | ns | 14 | 15 | ns | 21 | 8 | ns |
| positive | 6 | 20 | 4 | 2 | 3 | 3 | 12 | 8 | 17 | 3 | |||||
| Triple negative | |||||||||||||||
| no | 17 | 57 | 0.002 | 10 | 7 | ns | 7 | 10 | ns | 29 | 28 | ns | 43 | 14 | ns |
| yes | 12 | 7 | 5 | 7 | 7 | 5 | 3 | 4 | 6 | 1 | |||||
| Patient age | |||||||||||||||
| ≤ 42 y | 22 | 19 | < 0.001 | 12 | 10 | ns | 8 | 14 | ns | 6 | 13 | ns | 14 | 5 | ns |
| > 42 y | 16 | 66 | 9 | 7 | 10 | 6 | 30 | 36 | 52 | 14 | |||||
To compare further BRCA1 associated vs. sporadic cases, a second study panel of 56 patients that had been matched (p = 1.000) according to tumor size, lymph node status, presence of metastasis and tumor grade, was selected from the whole study cohort (Table 2). This matched group did not significantly differ regarding histologic subtype, ER, PR or Her2. However, patient age remained to be different in matched groups (p = 0.011).
Table 2. Patient characteristics (matched groups).
| BRCA1 | sporadic breast cancer | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TRα | TRβ | TRα | TRβ | |||||||||
| negative | positive | p | negative | positive | p | negative | positive | p | negative | positive | p | |
| Histology | ||||||||||||
| non NST | 2 | 3 | ns | 3 | 2 | ns | 5 | 1 | ns | 6 | 0 | ns |
| NST | 12 | 11 | 13 | 10 | 11 | 11 | 18 | 4 | ||||
| Grading | ||||||||||||
| G1, G2 | 2 | 5 | ns | 5 | 2 | ns | 3 | 4 | ns | 6 | 1 | ns |
| G3 | 12 | 9 | 11 | 10 | 13 | 8 | 18 | 3 | ||||
| pT | ||||||||||||
| pT1 | 7 | 6 | ns | 7 | 6 | ns | 9 | 4 | ns | 11 | 2 | ns |
| pT2-4 | 7 | 8 | 9 | 6 | 7 | 8 | 13 | 2 | ||||
| pN | ||||||||||||
| pN0 | 10 | 5 | ns | 6 | 9 | ns | 8 | 7 | ns | 12 | 3 | ns |
| pN1-3 | 4 | 9 | 10 | 3 | 8 | 5 | 12 | 1 | ||||
| pM | ||||||||||||
| pM0 | 7 | 4 | ns | 5 | 6 | ns | 8 | 3 | ns | 10 | 1 | ns |
| pM1 | 7 | 10 | 11 | 6 | 8 | 9 | 14 | 3 | ||||
| ER | ||||||||||||
| negative | 9 | 10 | ns | 13 | 6 | ns | 9 | 8 | ns | 14 | 3 | ns |
| positive | 5 | 4 | 3 | 6 | 7 | 4 | 10 | 1 | ||||
| PR | ||||||||||||
| negative | 10 | 10 | ns | 12 | 8 | ns | 10 | 6 | ns | 14 | 2 | ns |
| positive | 4 | 4 | 4 | 4 | 6 | 6 | 10 | 2 | ||||
| Her2 | ||||||||||||
| negative | 8 | 9 | ns | 9 | 8 | ns | 7 | 6 | ns | 11 | 2 | ns |
| positive | 2 | 2 | 3 | 1 | 7 | 2 | 9 | 0 | ||||
| Triple negative | ||||||||||||
| no | 7 | 7 | ns | 7 | 7 | ns | 12 | 6 | ns | 16 | 2 | ns |
| yes | 4 | 4 | 6 | 2 | 3 | 3 | 5 | 1 | ||||
| Patient age | ||||||||||||
| ≤ 42 y | 8 | 7 | ns | 7 | 8 | ns | 3 | 2 | ns | 4 | 1 | ns |
| > 42 y | 6 | 7 | 9 | 4 | 13 | 10 | 20 | 3 | ||||
TRs are Frequently Expressed in BRCA1 Associated Breast Cancer
TRs were found to be expressed in breast cancer tissue through a nuclear staining signal (Fig 1A and 1D). Specificity of TR staining was controlled by incubating breast cancer tissue sections with species-matched control serum instead of the primary antibody. No staining signal was visible in these control sections (Fig 1B and 1E). Tissue sections of thyroid gland struma (Fig 1C) and vaginal epithelium (Fig 1F) constantly presented a high TR immunoreactivity and were thus chosen as positive controls.
Regarding sporadic cancer cases (Fig 2A and 2D) 57 out of 86 (66.3%) stained positive for at least one of the two TRs (i.e. T0052α (50/86) and/or TRβ (19/86)), 12 out of 86 (14.0%) were scored as double positive (i.e. expressing TRα and TRβ at the same time) and 33.7% (29/86) were double negative. The majority of BRCA1 mutated tumors (30/38; 78.9%) were scored as positive for TRα and/or TRβ while only eight (8/38; 21.1%) cases expressed neither TRα nor TRβ. 17/38 (44.7%) cases stained positive for TRα (Table 3, Fig 2B and 2C), and 20/38 (52.6%) stained positive for TRβ (Table 3, Fig 2E and 2F). Finally, seven out of the 38 (18.4%) patients included were diagnosed a double positive tumor (i.e. a tumor expressing both TRα and TRβ at the same time). Regarding TRα positivity, there was no significant difference comparing sporadic (50/86; 58.1%) vs. BRCA1 associated cases (17/38; 44.7%; Fig 2A–2C). As compared to BRCA1 mutated cases (20/38; 52.6%) significantly fewer sporadic cancers (19/86; 22.1%) expressed TRβ (p = 0.001, Fig 2D–2F). With respect to TRβ this effect remained to be significant when cancers of high grade (p < 0.001) or of negative lymph node status (p = 0.039) were compared. Furthermore, also in the matched group, TRβ positivity was associated with the presence of mutant BRCA1 (p = 0.037). Again, we did not observe a significant difference in receptor expression ratios in the case of TRα comparing sporadic (12/28; 42.9%) vs. BRCA1 (14/28; 50.0%) associated cases. TRα and TRβ were not correlated among each other, neither in sporadic nor in BRCA1 associated cases. However co-expression of both receptors as determined by staining serial sections (Fig 3A) or by double-immunofluorescence (Fig 3B) was observed in some cells. Finally, in BRCA1 mutated cases there was no relation of TR positivity and circulating hormone levels (Table 4).
Fig 2. TRα and TRβ staining in breast cancer tissue.
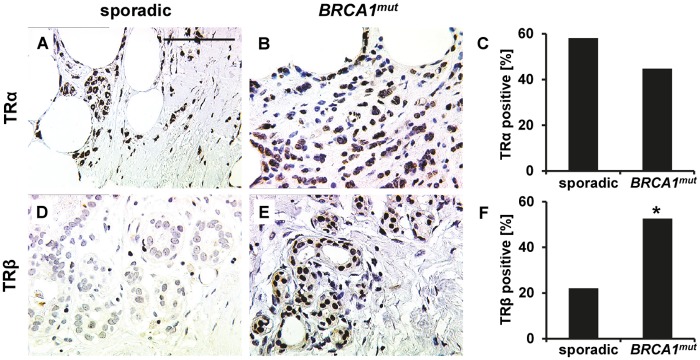
Representative photomicrographs of TRα and TRβ immunohistochemistry staining in breast cancer tissue are shown. TRα was found to be abundantly expressed though there was no significant difference regarding the number of TRα positive cases when sporadic (A) vs. BRCA1 (B) mutated cancers were compared (C). Representative images of a TRβ staining scored as negative (D) and a TRβ staining scored as positive (E) in spontaneous (D) and BRCA1 mutated (E) cancer tissue are presented, respectively. TRβ was found to be expressed more frequently in BRCA1 mutated (E) cases as compared to spontaneous breast cancers (D). The fraction of TRβ positive cases in each group is illustrated in F. Significant changes are indicated by stars (*) and scale bar (applies to A, B, D and E) represents 100 μm.
Table 3. Overall TR immunoreactivity.
| n | % | |||
|---|---|---|---|---|
| total | BRCA1 | 38 | 30.6 | |
| sporadic | 86 | 69.4 | ||
| BRCA1 | TRα | negative | 21 | 55.3 |
| positive | 17 | 44.7 | ||
| TRβ | negative | 18 | 47.4 | |
| positive | 20 | 52.6 | ||
| sporadic breast cancer | TRα | negative | 36 | 41.9 |
| positive | 50 | 58.1 | ||
| TRβ | negative | 67 | 77.9 | |
| positive | 19 | 22.1 |
Fig 3. Co-localisation of TRα and TRβ.
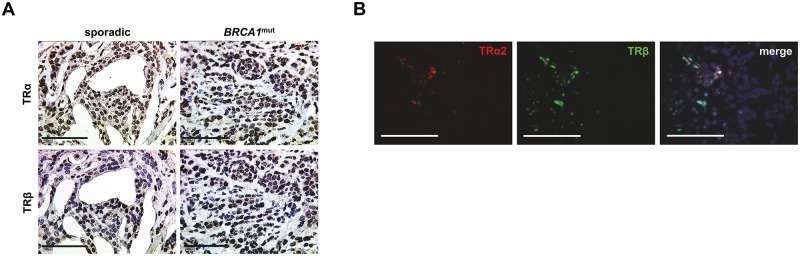
Co-localisation of TRα and TRβ was observed in some tumor cells as shown by staining serial slides (A) or by co-staining of TRs (TRα2: red signal, TRβ: green signal, DNA: blue signal) by double-immunoflourescence (B). Scale bars represent 100 μm and representative photomicrographs are shown.
Table 4. TSH, fT3 and fT4 serum levels.
| BRCA1 | sporαdic | BRCA1 | sporαdic | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BRCA1 | sporadic | p | TRα neg. | TRα pos. | p | TRα neg. | TRα pos. | p | TRβ neg. | TRβ pos. | p | TRβ neg. | TRβ pos. | p | |
| TSH | |||||||||||||||
| ref. range | 10 | 20 | 5 | 5 | 12 | 8 | 4 | 6 | 17 | 3 | |||||
| < ref. range | 2 | 4 | ns | 1 | 1 | ns | 3 | 1 | ns | 2 | 0 | ns | 2 | 2 | ns |
| > ref. range | 1 | 2 | ns | 0 | 1 | ns | 1 | 1 | ns | 0 | 1 | ns | 0 | 2 | 0.043 |
| fT3 | |||||||||||||||
| ref. range | 6 | 10 | 3 | 3 | 6 | 4 | 4 | 2 | 7 | 3 | |||||
| < ref. range | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na |
| > ref. range | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na |
| fT4 | |||||||||||||||
| ref. range | 7 | 11 | 3 | 4 | 8 | 3 | 4 | 3 | 8 | 3 | |||||
| < ref. range | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na | 0 | 0 | na |
| > ref. range | 0 | 2 | ns | 0 | 0 | na | 1 | 1 | ns | 0 | 0 | na | 1 | 1 | ns |
TSH and thyroid hormone levels in breast cancer patients as quantified at time of first diagnosis are shown. Since clinical data were retrieved retrospectively, data regarding thyroid function were not available in all the cases. ref = reference, ns = not significant, na = not applicable
Both, ER and PR, were expressed in about one-third (ER: 11/38, PR: 11/38) of BRCA1 mutant breast cancer. Information on Her2 was only available in 28 BRCA1 mutated cases, and six of these 28 patients (6/28, 21.4%) were scored as Her2 positive. Sufficient information to conclude on potential presence of triple negativity was available in 29 of 38 BRCA1 associated cancers. Twelve out of these 29 cases were finally classified as triple negative (12/29, 41.4%) while the remaining 17 cases were scored positive for at least one of the three hormone receptors (ER, PR, Her2). Interestingly, nine of these twelve (9/12, 75.0%) triple negative BRCA1 associated breast cancer cases were found to express at least one of the two TRs. TRα was detected in seven out of twelve (7/12, 58.3%) and TRβ was detected in five of twelve (5/12, 41.7%) triple negative breast cancer cases, respectively. Three triple negative cases (3/12, 25.0%) stained positive for both TRs at the same time while another three cases (3/12, 25.0%) expressed neither TRα nor TRβ.
TRα and TRβ are of Opposing Prognostic Significance in BRCA1 Related Breast Cancer
Five year survival of TR positive vs. negative cases was compared. TRα positivity was associated with significantly reduced five-year survival in BRCA1 carriers (p = 0.030) (Fig 4A), while no effect of TRα on patient survival was observed in sporadic cancer cases (Fig 4B). In contrast, BRCA1 associated cancers characterized as TRβ positive presented a significantly higher five-year survival rate as compared to TRβ-negative patients (p = 0.007), while TRβ failed to be of prognostic significance in sporadic breast cancer. Regarding BRCA1 associated cancer cases neither TRα nor TRβ presented a significant association to any of the clinicopathological variables examined (Table 1). However, to reduce the effect of possible confounders, survival analysis was also performed in selected subgroups classified as NST, high grade, pN1-3, presence of metastasis, ER negative, PR negative and patient age below 42 years. TRα and TRβ remained to be predictive for five-year survival (TRα positivity—reduced five-year survival; TRβ positivity—advanced five-year survival) in breast cancer of non-specific type (TRα: p = 0.024; TRβ: p = 0.014), in high grade cancer (TRα: p = 0.042; TRβ: p = 0.005) and in patients aged younger than 42 years (TRα: p = 0.045; TRβ: p = 0.019). Further, TR positivity was also associated with five-year survival (TRα positivity—reduced five-year survival; TRβ positivity—advanced five-year survival) in ER negative (TRα: p = 0.032; TRβ: p = 0.020), PR negative (TRα: p = 0.011; TRβ: p = 0.029) BRCA1 mutated breast cancer. Finally, TRβ positivity predicted a higher 5-year survival rate in metastasized (TRβ: p = 0.036) or lymph node positive (TRβ: p = 0.031) BRCA1 mutated cancer.
Fig 4. TR immuno-positivity predicts prognosis.
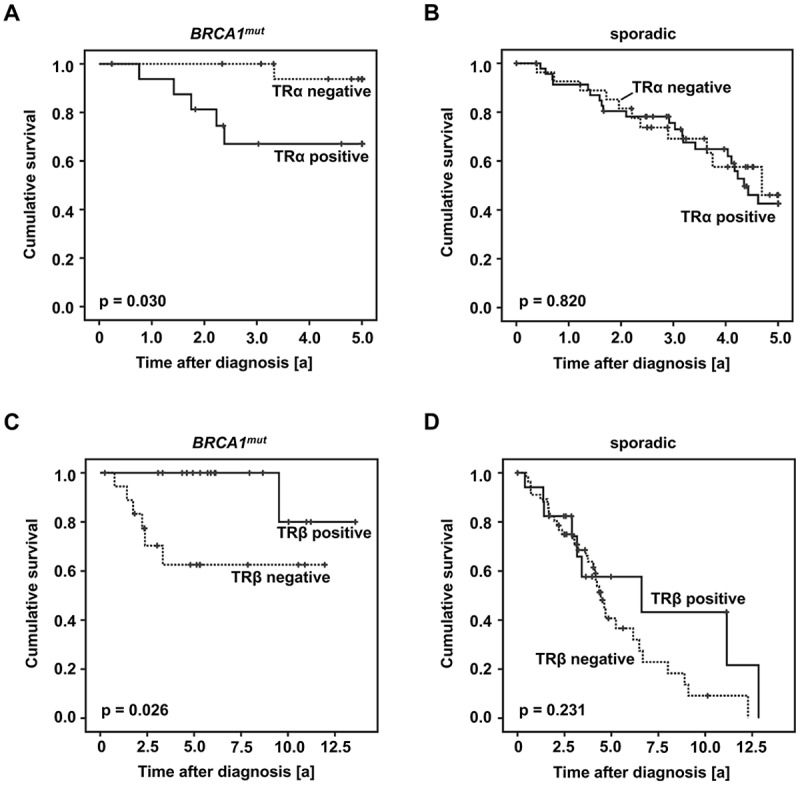
Survival of TRα positive vs. negative cases (A, B) and TRβ positive vs. negative cases (C, D) was plotted in accordance with Kaplan-Meier survival curves. TRα was a negative predictor regarding five-year survival of patients carrying a BRCA1 mutation (A). In sporadic cancer cases TRα was not associated with five-year survival (B). TRβ positivity was associated with a significantly prolonged overall survival in patients carrying a BRCA1 mutation (C). In sporadic cancer cases TRβ failed to predict prognosis (D).
More importantly, TRβ positivity predicted longer overall survival in BRCA1 mutated patients (p = 0.026) (Fig 4C). Again, no relation was found between TRβ and overall survival in sporadic breast cancer (Fig 4D). TRβ remained to be a positive prognostic factor in BRCA1 mutated cancers classified as NST (p = 0.036), high grade (p = 0.018), ER negative (p = 0.020) or PR negative (p = 0.029). However, though TRβ was also related to favorable prognosis in metastasized cancers (p = 0.036), it failed to be of prognostic significance in lymph node positive (p = 0.108) cases. The association of TRα positivity and reduced overall survival rate of BRCA1 related cases was of borderline significance (p = 0.064).
Regarding the matched group, TRβ remained to be predictive for advanced overall survival in BRCA1 associated cases (p = 0.018), while survival rates of TRβ positive vs. negative cases did not differ among sporadic breast cancers. A trend of TRα positivity being associated with reduced overall survival (p = 0.059) was observed in patients of the matched group carrying a BRCA1 germline mutation. Again TRα was not predictive for sporadic breast cancer overall survival.
TRs are active in BRCA1 mutant HCC3153
TRβ overexpression in BRCA1 mutant cases was confirmed on protein and mRNA level in HCC3153 carrying a homozygous insertion in BRCA1 as compared to BRCA1 competent MCF7 (Fig 5). In line with our observation made in sporadic vs. BRCA1 associated patients TRβ protein positivity rate was increased 8.4-fold in HCC3153 as compared to MCF7 (p = 0.009) (Fig 5A and 5C). THRB mRNA was increased in HCC3153 by a mean factor of 3.7 (S.E. range: 2.3–7.6) as compared to MCF7 (p = 0.034) (Fig 5E). Though there was no difference regarding TRα immunopositivity (Fig 5A and 5B), THRA mRNA was found to be elevated in HCC3153 by a mean factor of 9.3 (S.E. range: 5.3–20, p < 0.001) (Fig 5D).
Fig 5. TRβ is strongly expressed in the absence of functional BRCA1.

TRβ was significantly over-expressed on protein (A, C) and mRNA level (E) when BRCA1 competent MCF7 and HCC3153 carrying stop mutation in BRCA1 were compared. However, in the case of TRα this difference was significant on mRNA level only (A, B, D). Mann-Whitney U Tests (B, C) and the REST2009 algorithm for determining relative gene expression (D, E) have been applied. Significant changes are indicated by stars (*) and representative images are shown.
We further questioned whether TRs are active in the BRCA1 deficient HCC3153 cell line. TRβ activation was shown to repress CTNNB1, the gene encoding tumor promoting β-catenin [22], while TRα activation was reported to induce CTNNB1 [23]. Hence T3 stimulation was performed in HCC3153 silenced for THRA as well as in HCC3153 transfected with an off-target control siRNA. In scrambled control (scr) transfected cells treated with T3 CTNNB1 expression was induced by a mean factor of 1.3 (S.E. range: 1.1–1.9, p < 0.001) while those HCC3153 silenced for THRA appeared to repress CTNNB1 when stimulated with T3. Both THRA specific siRNAs were able to reverse the T3 effect on CTNNB1 observed in scr treated cells resulting in repression of CTNNB1 by a mean factor of si2: 0.69 (S.E. range: 0.61–0.86; p = 0.023) or si3: 0.60 (S.E. range: 0.54–0.70; p = 0.034) in those cells silenced for THRA and at the same time treated with T3 (Fig 6A).
Fig 6. TRs are active in the absence of functional BRCA1.

The TR target gene CTNNB1 encoding β-Catenin was quantified in HCC3153 (BRCA1 mut) upon T3 stimulation. While T3 induced CTNNB1 in HCC3153 expressing THRA as well as THRB, CTNNB1 expression was significantly down-regulated upon T3 stimulation in HCC3153 silenced for THRA (A). Blocking TRs significantly reduced proliferation (B) and viability (C) of HCC3153 as quantified by BrdU (B) and WST-1 (C) assay, respectively. Reduced viability was also observed in HCC3153 silenced for THRA (D). Efficiency of THRA knockdown was validated on mRNA (E) and protein (F) level. THRB mRNA was not significantly altered in cells silenced for THRA (E). Mann-Whitney U Tests (B-D, F) and the REST2009 algorithm for determining relative gene expression (A, E) have been applied. Significant changes are indicated by stars (*) representative images are shown. scr—scrambled siRNA control, HiP—HighPerFect transfection reagent only control.
TRα has been reported to act as a proliferation factor [24]. When HCC3153 were treated with the non-selective TR blocker 1–850 their proliferation (as quantified by BrdU incorporation) and viability (as quantified by WST-1 turnover) was found to be significantly reduced as compared to carrier solution (DMSO) treated samples. Median reduction of proliferation was 0.50 (p = 0.004) in samples treated with 10-5 M of 1–850 and 0.52 (p = 0.004) when cells were incubated with 1–850 at a concentration of 10-4 M (Fig 6B). Median viability was also decreased in HCC3153 stimulated with 1–850 (10-5 M: 0.64-fold, p = 0.004; 10-4 M: 0.73-fold, p = 0.004) (Fig 6C). Knocking down THRA in HCC3153 significantly lowered median cell viability (si2: 0.43-fold, p = 0.004; si3: 0.79-fold, p = 0.004) as determined by WST-1 assay (Fig 6D).
Knockdown efficiency was validated and THRA mRNA was significantly reduced in samples silenced for THRA (si2: 0.47-fold (S.E. range: 0.33–0.61), p = 0.027; si3: 0.24-fold (S.E. range is 0.17–0.39), p = 0.024) as compared to transfection reagent treated samples (HiP) (Fig 6E). THRA mRNA was also significantly repressed in THRA silenced samples (si2: 0.59-fold (S.E. range: 0.55–0.66), p = 0.033; si3: 0.31-fold (S.E. range is 0.19–0.43), p < 0.001) when compared to samples that had been transfected with the off target control (scr) (Fig 6E). THRA knockdown did not affect THRB mRNA expression (Fig 6E). Samples silenced for THRA showed significantly reduced TRα protein (Fig 6F) as compared to HiP treated (median reduction (si2): 0.27-fold, p = 0.02; median reduction (si3): 0.18-fold, p = 0.02) and as compared to scr transfected (median reduction (si2): 0.36-fold, p = 0.02; median reduction (si3): 0.24-fold, p = 0.02) samples.
BRCA1 mutant cells fail to degrade TRα1
Wildtype BRCA1 has been reported to regulate protein half-life of nuclear hormone receptors e.g. progesterone receptor via its ubiquitinilation and sumoylation activity [25]. Though TRs are regulated by ubiquitinilation as well, it is not known whether this effect is reliant on functional BRCA1. Though BRCA1 competent MCF7 cells significantly lost TRα1 positivity (median reduction: 0.20-fold, p = 0.021) in response to T3 stimulation, no such phenomenon was obvious in HCC3153 (Fig 7A). When stimulated with T3, subcellular localisation of TRα1 changed from a former predominant nuclear signal to a diffuse nuclear-cytoplasmic staining pattern. No such change in TRα1 localisation was visible in HCC3153 (Fig 7B).
Fig 7. TRα1 positivity is reduced in MCF7 upon T3 stimulation.
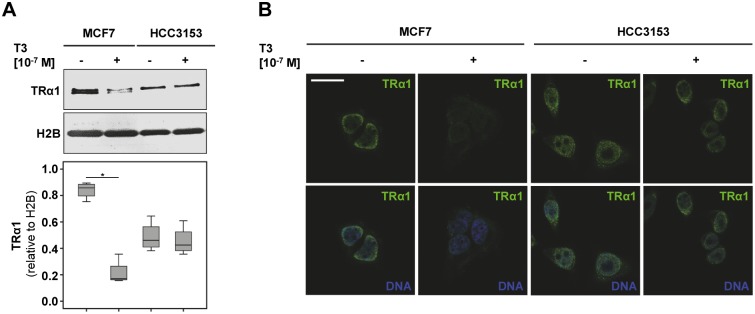
While BRCA1 competent MCF7 cells significantly lost TRα1 positivity in response to T3 stimulation, no such phenomenon was obvious in HCC3153 (A). Double immunofluorescence was performed (TRα1: green signal, DNA: blue signal). When stimulated with T3 subcellular localisation of TRα1 changed from a former predominant nuclear signal to a diffuse nuclear-cytoplasmic staining pattern (B). No such change in TRα1 localisation was visible in HCC3153 (B). Scale bar represents 25 μm. Significant changes as determined by relevant Mann-Whitney U Tests are indicated by stars (*) and representative images are shown.
Upon T3 treatment some co-localisation of TRα1 and ubiquitin was observed in BRCA1 competent MCF7 (Fig 8A–8F, magnification in D’-F’), potentially suggesting an ubiquitinilation mediated degradation process.
Fig 8. TRα1 co-localizes with ubiquitin upon T3 stimulation.

BRCA1 competent MCF7 were co-stained for TRα1 (green signal) and ubiquitin (Ubq, red signal). TRα1 was located in the nucleus (DNA: blue signal) of untreated MCF7 (A, C). The red channel was slightly enhanced in B, C thus to show the faint Ubq signal of un-stimulated MCF7 (B, C). In BRCA1 competent MCF7 co-localization of TRα1 and Ubq was observed in samples stimulated with T3 (D-F, magnified in D’-F’). Scale bar in A equals 25 μm applies to A-F, scale bar in D’ equals 12.5 μm applies to D’-F’. Representative images are shown.
Discussion
This work showed TRβ to be more frequently expressed in BRCA1 associated breast cancers as compared to sporadic breast cancer. TRα and TRβ were observed to be of opposing prognostic significance regarding five year survival in BRCA1 mutation carriers. In addition, TRβ positivity remained to be significantly associated with overall survival. Using cell lines we were able to show that TRs are active in a BRCA1 mutant genetic background.
While a direct link is still missing, thyroid hormones and breast cancer have been associated for quite a while; e. g. an elevated incidence of thyroid dysfunction has been observed in breast cancer patients [26–28]. While several studies frequently found relative hyperthyreosis in breast cancer patients [26,29–31], others report aggravating hypothyroidism during breast cancer therapy or demonstrated an increased prevalence of autoimmune thyroid disorders in breast cancer [27,32]. In line with this, a former prospective clinical trial of our group revealed higher levels of auto-immune antibodies targeting thyroid hormone receptors (TRAKs) and of thyroidal effector hormones (triiodothyronine (T3), Thyroxine (T4)) in breast cancer patients as compared to controls [26]. Thyroid hormones did not correlate with TR expression in the current analysis. However, to the best of our knowledge no data exist upon thyroid function in BRCA1 associated breast cancers so far. At least in the case of estrogen and ERs, nuclear hormone receptor expression does not seem to be related to circulating hormone levels [33]. More important, though anti-estrogen endocrine treatments are widely applied to ER positive patients, circulating estrogens are not accessed during clinical routine indicating that even if circulating estrogens alter ER expression this does not seem to be of clinical relevance.
Our current analysis on TRs indicated opposing roles of TRα and TRβ on survival of women carrying a BRCA1 germline mutation and on target gene activation in BRCA1 mutant cancer cells. Interestingly, a couple of other in vitro cell culture studies also report a tumor-promoting effect of TRα activation [34–37], while TRβ stimulation resulted in just the opposite [38–40]. For instance, quite recently TRα was found to enhance cancer cell migration and metastasis via inhibition of miR-17 [34] as well as to maintain pro-neoplastic potency by interacting with the cyclin D1/CDK/Rb/E2F cascade [35], the wnt pathway [3] or by inducing CTNNB1 gene expression [23]. Since our data demonstrate that selective THRA knockdown inhibited cell growth, a tumor promoting action of TRα is further affirmed. In contrast, TRβ exerted anti-tumor activity by inhibiting beta catenin action [39] or by interfering with AKT-mTOR-p70S6K signaling in xenografted mice [38]. In line with our data on differential regulation of CTNNB1 in BRCA1 deficient HCC3153 either expressing both TRα and TRβ or just expressing TRβ (i.e. silenced for THRA) further supports an opposing role of TRs in breast cancer [22]. Simultaneous blockade of TRα and TRβ by the non-selective TR antagonist 1–850 reduced cell growth potentially indicating that inhibition of pro-proliferative TRα may outweigh TRβ mediated effects. Interestingly, regarding both CTNNB1 expression and cell growth TRα effects seem to dominate the influence of TRβ.
Surprisingly, TRs demonstrated prognostic significance only in those patients diagnosed for a BRCA1 germline mutation but not in sporadic breast cancer cases. This is in agreement with reports where neither TRα nor TRβ were significantly associated with patient prognosis in a different, independent sample of 82 sporadic breast cancers [6]. Although experimental evidence explaining the impact of TRs in BRCA1 associated cancers is still lacking, the current study showed TRβ to be more frequently expressed in BRCA1 mutation carriers. This was confirmed by comparing TRβ expression on both mRNA and protein level in BRCA1 mutant HCC3153 vs. BRCA1 competent MCF7. Immuno-positivity of TRα was not significantly different when BRCA1 mutant and sporadic breast cancers were compared though THRA mRNA expression was induced in HCC3153. This may be explained by the fact that correlation of protein and mRNA expression of certain genes has been reported to be rather poor [41–45]. The latter phenomenon may be due to complex post-transcriptional regulatory processes, many of which have not been sufficiently understood so far. In addition the probability of a direct correlation between mRNA and protein seems to be dependent on e.g. ribosomal occupancy, protein stability, codon usage or on whether mRNA concentrations of a certain ORFs [42,43]. Since the wildtype BRCA1 protein contributes to protein degradation via its ubiquitinilation and sumoylation activity of nuclear hormone receptors [25], loss of functional BRCA1 may explain TRβ overexpression and prognostic relevance of both TRs. This hypothesis is supported by increasing evidence of ubiquitinylation and sumoylation mediated regulation of TRs upon hormone binding [46,47]. Our results on degradation of TRα1 may indicate that BRCA1 is involved in posttranscriptional regulation of TRs. However co-immunoprecipitation assays will be needed to confirm a physical interaction of TRα1 and Ubiquitin. BRCA1 mutants may be used to confirm whether reduced TRα1 degradation observed in HCC3153 may indeed be reliant on BRCA1. Since, from a clinical point of view, activation of a tumor suppressor gene is less attractive than blocking an oncogene, only posttranscriptional regulation of TRα1 was studied.
Survival analysis, cell growth and target gene activation assays performed within this study indicate that TRs not only hold prognostic significance but also are active in a BRCA1 deficient genetic background. Therefore we hypothesize that BRCA1 mutant cells may undergo a TR mediated phenomenon termed ‘oncogenic addiction’ [48]. Within this scenario, cancer cells acquire abnormalities in several onco (e.g. TRα)—and tumor suppressor (e.g. TRβ) genes. Gene products being crucial for cancer cell survival provide an Achilles heel for tumors and display interesting targets to be exploited in cancer treatment [48]. In case of BRCA1 associated breast cancers, TRs might display excellent targets for novel drugs, especially in triple-negative cases. In line with this, the current study demonstrated TRs to be expressed in 9 out of 12 triple negative cancers and to be highly sensitive to TR modulation in triple negative, BRCA1 mutant HCC3153.
Within the last decade a couple of selective as well as non-selective TR modifiers have been investigated. Dronedarone was found to inhibit TRα in vitro as well as in vivo [49] and a study in Xenopus laevis reported the TRα stimulating compound CO23 to support brain cell proliferation, while such induction of proliferation was not seen in animals treated with the TRβ selective agonists GC1 or GC24 [24].
In conclusion, our work revealed at least that TRs are active in BRCA1 associated breast cancer, that TRβ expression in BRCA1 mutant tumor samples is associated with a prolonged overall survival, and that both TRs may arise as interesting alternative targets for endocrine treatment of BRCA1 associated triple negative breast cancer.
Acknowledgments
The authors thank Christina Kuhn, Susanne Kunze, Verena Nick and Irmgard Wiest for their excellent technical assistance. Further we would like to thank Professor Jutta Engel (Munich Cancer Registry) and Verena Lanser for their help with retrieving follow-up data. This work was supported by a Deutschlandstipendium scholarship of the Ludwig-Maximilians-University of Munich Medical Faculty to SH. The funder had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by a Deutschlandstipendium scholarship of the Ludwig-Maximilians-University of Munich Medical Faculty to SH. The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Bayraktar S, Gluck S (2012) Systemic therapy options in BRCA mutation-associated breast cancer. Breast Cancer Res Treat 135: 355–366. 10.1007/s10549-012-2158-6 [DOI] [PubMed] [Google Scholar]
- 2. Da Silva L, Lakhani SR (2010) Pathology of hereditary breast cancer. Mod Pathol 23 Suppl 2: S46–51. 10.1038/modpathol.2010.37 [DOI] [PubMed] [Google Scholar]
- 3. Foulkes WD, Stefansson IM, Chappuis PO, Begin LR, Goffin JR, Wong N, et al. (2003) Germline BRCA1 mutations and a basal epithelial phenotype in breast cancer. J Natl Cancer Inst 95: 1482–1485. [DOI] [PubMed] [Google Scholar]
- 4. Honrado E, Osorio A, Palacios J, Benitez J (2006) Pathology and gene expression of hereditary breast tumors associated with BRCA1, BRCA2 and CHEK2 gene mutations. Oncogene 25: 5837–5845. [DOI] [PubMed] [Google Scholar]
- 5. Stevens KN, Vachon CM, Couch FJ (2013) Genetic susceptibility to triple-negative breast cancer. Cancer Res 73: 2025–2030. 10.1158/0008-5472.CAN-12-1699 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Ditsch N, Toth B, Himsl I, Lenhard M, Ochsenkuhn R, Friese K, et al. (2013) Thyroid hormone receptor (TR)alpha and TRbeta expression in breast cancer. Histol Histopathol 28: 227–237. [DOI] [PubMed] [Google Scholar]
- 7. Perri A, Catalano S, Bonofiglio D, Vizza D, Rovito D, Qi H, et al. (2014) T3 enhances thyroid cancer cell proliferation through TRbeta1/Oct-1-mediated cyclin D1 activation. Mol Cell Endocrinol 382: 205–217. 10.1016/j.mce.2013.10.001 [DOI] [PubMed] [Google Scholar]
- 8. Chi HC, Liao CH, Huang YH, Wu SM, Tsai CY, Liao CJ, et al. (2013) Thyroid hormone receptor inhibits hepatoma cell migration through transcriptional activation of Dickkopf 4. Biochem Biophys Res Commun 439: 60–65. 10.1016/j.bbrc.2013.08.028 [DOI] [PubMed] [Google Scholar]
- 9. Raparti G, Jain S, Ramteke K, Murthy M, Ghanghas R, Ramanand S, et al. (2013) Selective thyroid hormone receptor modulators. Indian J Endocrinol Metab 17: 211–218. 10.4103/2230-8210.109663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Fischer C, Kuchenbacker K, Engel C, Zachariae S, Rhiem K, Meindl A, et al. (2013) Evaluating the performance of the breast cancer genetic risk models BOADICEA, IBIS, BRCAPRO and Claus for predicting BRCA1/2 mutation carrier probabilities: a study based on 7352 families from the German Hereditary Breast and Ovarian Cancer Consortium. J Med Genet 50: 360–367. 10.1136/jmedgenet-2012-101415 [DOI] [PubMed] [Google Scholar]
- 11. Kao J, Salari K, Bocanegra M, Choi YL, Girard L, Gandhi J, et al. (2009) Molecular profiling of breast cancer cell lines defines relevant tumor models and provides a resource for cancer gene discovery. PLoS One 4: e6146 10.1371/journal.pone.0006146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Ditsch N, Mayr D, Lenhard M, Strauss C, Vodermaier A, Gallwas J, et al. (2012) Correlation of thyroid hormone, retinoid X, peroxisome proliferator-activated, vitamin D and oestrogen/progesterone receptors in breast carcinoma. Oncol Lett 4: 665–671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Ditsch N, Toth B, Mayr D, Lenhard M, Gallwas J, Weissenbacher T, et al. (2012) The association between vitamin D receptor expression and prolonged overall survival in breast cancer. J Histochem Cytochem 60: 121–129. 10.1369/0022155411429155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Ditsch N, Vrekoussis T, Lenhard M, Ruhl I, Gallwas J, Weissenbacher T, et al. (2012) Retinoid X receptor alpha (RXRalpha) and peroxisome proliferator-activated receptor gamma (PPARgamma) expression in breast cancer: an immunohistochemical study. In Vivo 26: 87–92. [PubMed] [Google Scholar]
- 15. Lenhard M, Lennerova T, Ditsch N, Kahlert S, Friese K, Mayr D, et al. (2011) Opposed roles of follicle-stimulating hormone and luteinizing hormone receptors in ovarian cancer survival. Histopathology 58: 990–994. 10.1111/j.1365-2559.2011.03848.x [DOI] [PubMed] [Google Scholar]
- 16. Lenhard M, Tereza L, Heublein S, Ditsch N, Himsl I, Mayr D, et al. (2012) Steroid hormone receptor expression in ovarian cancer: Progesterone receptor B as prognostic marker for patient survival. BMC Cancer 12: 553 10.1186/1471-2407-12-553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Lenhard M, Tsvilina A, Schumacher L, Kupka M, Ditsch N, Mayr D, et al. (2012) Human chorionic gonadotropin and its relation to grade, stage and patient survival in ovarian cancer. BMC Cancer 12: 2 10.1186/1471-2407-12-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Heublein S, Mayr D, Vrekoussis T, Friese K, Hofmann SS, Jeschke U, et al. (2013) The G-protein coupled estrogen receptor (GPER/GPR30) is a gonadotropin receptor dependent positive prognosticator in ovarian carcinoma patients. PLoS One 8: e71791 10.1371/journal.pone.0071791 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Heublein S, Lenhard M, Vrekoussis T, Schoepfer J, Kuhn C, Friese K, et al. (2012) The G-Protein-Coupled Estrogen Receptor (GPER) is Expressed in Normal Human Ovaries and is Upregulated in Ovarian Endometriosis and Pelvic Inflammatory Disease Involving the Ovary. Reprod Sci 19: 1197–1204. 10.1177/1933719112446085 [DOI] [PubMed] [Google Scholar]
- 20. Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29: e45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. McShane LM, Altman DG, Sauerbrei W, Taube SE, Gion M, Clark GM (2005) Reporting recommendations for tumor marker prognostic studies. J Clin Oncol 23: 9067–9072. [DOI] [PubMed] [Google Scholar]
- 22. Guigon CJ, Kim DW, Zhu X, Zhao L, Cheng SY (2010) Tumor suppressor action of liganded thyroid hormone receptor beta by direct repression of beta-catenin gene expression. Endocrinology 151: 5528–5536. 10.1210/en.2010-0475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Plateroti M, Kress E, Mori JI, Samarut J (2006) Thyroid hormone receptor alpha1 directly controls transcription of the beta-catenin gene in intestinal epithelial cells. Mol Cell Biol 26: 3204–3214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Denver RJ, Hu F, Scanlan TS, Furlow JD (2009) Thyroid hormone receptor subtype specificity for hormone-dependent neurogenesis in Xenopus laevis. Dev Biol 326: 155–168. 10.1016/j.ydbio.2008.11.005 [DOI] [PubMed] [Google Scholar]
- 25. Calvo V, Beato M (2011) BRCA1 counteracts progesterone action by ubiquitination leading to progesterone receptor degradation and epigenetic silencing of target promoters. Cancer Res 71: 3422–3431. 10.1158/0008-5472.CAN-10-3670 [DOI] [PubMed] [Google Scholar]
- 26. Ditsch N, Liebhardt S, Von Koch F, Lenhard M, Vogeser M, Spitzweg C, et al. (2010) Thyroid function in breast cancer patients. Anticancer Res 30: 1713–1717. [PubMed] [Google Scholar]
- 27. Rasmusson B, Feldt-Rasmussen U, Hegedus L, Perrild H, Bech K, Hoier-Madsen M (1987) Thyroid function in patients with breast cancer. Eur J Cancer Clin Oncol 23: 553–556. [DOI] [PubMed] [Google Scholar]
- 28. Perry M, Goldie DJ, Self M (1978) Thyroid function in patients with breast cancer. Ann R Coll Surg Engl 60: 290–293. [PMC free article] [PubMed] [Google Scholar]
- 29. Santos CT, Howe HY, Siew FK (2009) Breast cancer in a male with thyroid-associated orbitopathy and subclinical hyperthyroidism: a rare occurrence. Orbit 28: 162–165. [DOI] [PubMed] [Google Scholar]
- 30. Ferreira E, da Silva AE, Serakides R, Gomes MG, Cassali GD (2007) Ehrlich tumor as model to study artificial hyperthyroidism influence on breast cancer. Pathol Res Pract 203: 39–44. [DOI] [PubMed] [Google Scholar]
- 31. Tsuchiya A, Kimijima I, Suzuki S, Higuchi I, Watanabe I, Endo S (1981) Breast cancer concurrent with hyperthyroidism: a case report. Jpn J Surg 11: 108–110. [DOI] [PubMed] [Google Scholar]
- 32. Giani C, Fierabracci P, Bonacci R, Gigliotti A, Campani D, De Negri F, et al. (1996) Relationship between breast cancer and thyroid disease: relevance of autoimmune thyroid disorders in breast malignancy. J Clin Endocrinol Metab 81: 990–994. [DOI] [PubMed] [Google Scholar]
- 33. Honma N, Saji S, Hirose M, Horiguchi S, Kuroi K, Hayashi S, et al. (2011) Sex steroid hormones in pairs of tumor and serum from breast cancer patients and pathobiological role of androstene-3beta, 17beta-diol. Cancer Sci 102: 1848–1854. 10.1111/j.1349-7006.2011.02018.x [DOI] [PubMed] [Google Scholar]
- 34. Lin YH, Liao CJ, Huang YH, Wu MH, Chi HC, Wu SM, et al. (2013) Thyroid hormone receptor represses miR-17 expression to enhance tumor metastasis in human hepatoma cells. Oncogene 32: 4509–4518. 10.1038/onc.2013.309 [DOI] [PubMed] [Google Scholar]
- 35. Sirakov M, Skah S, Lone IN, Nadjar J, Angelov D, Plateroti M (2012) Multi-level interactions between the nuclear receptor TRalpha1 and the WNT effectors beta-catenin/Tcf4 in the intestinal epithelium. PLoS One 7: e34162 10.1371/journal.pone.0034162 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 36. Wang X, Sun Z (2010) Thyroid hormone induces artery smooth muscle cell proliferation: discovery of a new TRalpha1-Nox1 pathway. J Cell Mol Med 14: 368–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Furuya F, Shimura H, Yamashita S, Endo T, Kobayashi T (2010) Liganded thyroid hormone receptor-alpha enhances proliferation of pancreatic beta-cells. J Biol Chem 285: 24477–24486. 10.1074/jbc.M109.100222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Kim WG, Zhao L, Kim DW, Willingham MC, Cheng SY (2014) Inhibition of tumorigenesis by the thyroid hormone receptor beta in xenograft models. Thyroid 24: 260–269. 10.1089/thy.2013.0054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Kim WG, Zhu X, Kim DW, Zhang L, Kebebew E, Cheng SY (2013) Reactivation of the silenced thyroid hormone receptor beta gene expression delays thyroid tumor progression. Endocrinology 154: 25–35. 10.1210/en.2012-1728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Kim DW, Zhao L, Hanover J, Willingham M, Cheng SY (2012) Thyroid hormone receptor beta suppresses SV40-mediated tumorigenesis via novel nongenomic actions. Am J Cancer Res 2: 606–619. [PMC free article] [PubMed] [Google Scholar]
- 41. Chen G, Gharib TG, Huang CC, Taylor JM, Misek DE, Kardia SL, et al. (2002) Discordant protein and mRNA expression in lung adenocarcinomas. Mol Cell Proteomics 1: 304–313. [DOI] [PubMed] [Google Scholar]
- 42. Olivares-Hernandez R, Bordel S, Nielsen J (2011) Codon usage variability determines the correlation between proteome and transcriptome fold changes. BMC Syst Biol 5: 33 10.1186/1752-0509-5-33 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Greenbaum D, Colangelo C, Williams K, Gerstein M (2003) Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol 4: 117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Lichtinghagen R, Musholt PB, Lein M, Romer A, Rudolph B, Kristiansen G, et al. (2002) Different mRNA and protein expression of matrix metalloproteinases 2 and 9 and tissue inhibitor of metalloproteinases 1 in benign and malignant prostate tissue. Eur Urol 42: 398–406. [DOI] [PubMed] [Google Scholar]
- 45. Anderson L, Seilhamer J (1997) A comparison of selected mRNA and protein abundances in human liver. Electrophoresis 18: 533–537. [DOI] [PubMed] [Google Scholar]
- 46. Brunelle M, Fayad T, Langlois MF (2011) Degradation of thyroid hormone receptor beta 1: existence of stable and unstable forms. Thyroid 21: 311–318. 10.1089/thy.2009.0363 [DOI] [PubMed] [Google Scholar]
- 47. Dace A, Zhao L, Park KS, Furuno T, Takamura N, Nakanishi M, et al. (2000) Hormone binding induces rapid proteasome-mediated degradation of thyroid hormone receptors. Proc Natl Acad Sci U S A 97: 8985–8990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Weinstein IB (2002) Cancer. Addiction to oncogenes—the Achilles heal of cancer. Science 297: 63–64. [DOI] [PubMed] [Google Scholar]
- 49. Van Beeren HC, Jong WM, Kaptein E, Visser TJ, Bakker O, Wiersinga WM (2003) Dronerarone acts as a selective inhibitor of 3,5,3'-triiodothyronine binding to thyroid hormone receptor-alpha1: in vitro and in vivo evidence. Endocrinology 144: 552–558. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.