

ABSTRACT
Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified on the basis of human clinical data. The mechanisms underlying its beneficial effects are still unknown. Gnotobiotic mice harboring F. prausnitzii (A2-165) and Escherichia coli (K-12 JM105) were subjected to 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced acute colitis. The inflammatory colitis scores and a gas chromatography-time of flight (GC/TOF) mass spectrometry-based metabolomic profile were monitored in blood, ileum, cecum, colon, and feces in gnotobiotic mice. The potential anti-inflammatory metabolites were tested in vitro. We obtained stable E. coli and F. prausnitzii-diassociated mice in which E. coli primed the gastrointestinal tract (GIT), allowing a durable and stable establishment of F. prausnitzii. The disease activity index, histological scores, myeloperoxidase (MPO) activity, and serum cytokine levels were significantly lower in the presence of F. prausnitzii after TNBS challenge. The protective effect of F. prausnitzii against colitis was correlated to its implantation level and was linked to overrepresented metabolites along the GIT and in serum. Among 983 metabolites in GIT samples and serum, 279 were assigned to known chemical reactions. Some of them, belonging to the ammonia (α-ketoglutarate), osmoprotective (raffinose), and phenolic (including anti-inflammatory shikimic and salicylic acids) pathways, were associated with a protective effect of F. prausnitzii, and the functional link was established in vitro for salicylic acid. We show for the first time that F. prausnitzii is a highly active commensal bacterium involved in reduction of colitis through in vivo modulation of metabolites along the GIT and in the peripheral blood.
IMPORTANCE
Inflammatory bowel diseases (IBD) are characterized by low proportions of F. prausnitzii in the gut microbiome. This commensal bacterium exhibits anti-inflammatory effects through still unknown mechanisms. Stable monoassociated rodents are actually not a reproducible model to decipher F. prausnitzii protective effects. We propose a new gnotobiotic rodent model providing mechanistic clues. In this model, F. prausnitzii exhibits protective effects against an acute colitis and a protective metabolic profile is linked to its presence along the digestive tract. We identified a molecule, salicylic acid, directly involved in the protective effect of F. prausnitzii. Targeting its metabolic pathways could be an attractive therapeutic strategy in IBD.
INTRODUCTION
The human colonic microbiota is a highly complex microbial ecosystem, and several hundred different species of bacteria are known to colonize the gut (1). The homeostasis established between the host and the resident bacteria plays important functional roles, such as protection against pathogenic organisms, maturation and modulation of the immune system, intestinal maturation, production of short-chain fatty acids (SCFA), mucosal physiology, and production of vitamins, such as vitamin K and biotin (2). Alterations in this homeostasis, named dysbiosis, have a profound impact on human health, as demonstrated in several gastrointestinal diseases, including inflammatory bowel diseases (IBD), irritable bowel syndrome (IBS), and colorectal cancer (CRC) (3–7). For instance, in genetically predisposed subjects, bacterial dysbiosis, including a decrease of Firmicutes, could result in disordered innate immune reactions and lead to IBD (8). Interestingly, in IBD patients that exhibited endoscopic recurrence, the proportion of the most prevalent species from the Clostridium leptum group, Faecalibacterium prausnitzii, was lower than in patients without endoscopic recurrence (7). Since then, it has been observed that F. prausnitzii decreases with the onset of inflammatory status; the relative abundance of F. prausnitzii might be a sensor of intestinal health in adults suffering from Crohn’s disease (CD), one of the two major IBD (3). This observation strengthens the paradigm of a mucosal protective role of F. prausnitzii through its anti-inflammatory potential.
The mechanisms leading to the beneficial protection of F. prausnitzii are still poorly understood. F. prausnitzii is predominant in the intestine of many other mammals; such ubiquity suggests that F. prausnitzii plays an important physiological role (3, 9). In humans, F. prausnitzii accounts for more than 3.5% of the total bacterial population (10) and belongs to the functional metagenomic core (11). Because it is one of the most abundant butyrate-producing bacterial species, its beneficial effect had been first attributed to butyrate, well known for its pleiotropic and beneficial effects in the gastrointestinal tract (GIT) (12, 13). However, we showed that F. prausnitzii-mediated butyrate production is not the only host health benefit metabolite linked to this species (7). In fact, in acute or chronic colitis models, the immunomodulation potential of F. prausnitzii involves uncharacterized secreted metabolites that directly impact the host immune response (7, 14). Moreover, we have recently proposed that F. prausnitzii participates in host epithelial homeostasis (15). The establishment of F. prausnitzii along the GIT may result from a combination of environmental factors, such as other commensal bacterial species, redox mediators, oxygen concentration, and mucus layer but also bile salt concentrations, as well as pH (16, 17). In early infancy, the abundance of F. prausnitzii is very low and increases after the establishment of primocolonizing bacteria (18). The complexity of conditions required for F. prausnitzii colonization might explain why the mechanisms accounting for its protection potential in vivo are still poorly understood. Thus, we set up a new model of diassociated mice housing F. prausnitzii and Escherichia coli (as a primocolonizing bacterial companion) to characterize F. prausnitzii’s protective role and metabolic profile during a chemically induced acute colitis.
RESULTS
Establishment of a stable E. coli/F. prausnitzii-diassociated mouse model.
We first tried to obtain F. prausnitzii-monocolonized mice by intragastric inoculation of 2 × 108 ± 1 × 108 CFU into germfree (GF) mice. After 24 h, no F. prausnitzii was detected in the feces, indicating that this species was not able to stably colonize the GIT of GF mice after a single oral inoculation. We then tried to establish F. prausnitzii in GF mice by both intragastric and intrarectal inoculations, and as shown in Fig. 1A, we succeeded since the amount of F. prausnitzii A2-165 ranged from 1.5 to 5 × 108 CFU/g of mouse feces. However, this colonization was obtained with a poor yield, with only 5 mice successfully colonized from 18 inoculated. Interestingly, F. prausnitzii exhibited an unusual morphology in the cecum of these 5 F. prausnitzii monoassociated animals, with cell sizes ranging from 6 to 25 µm (Fig. 1B), which is very rarely seen in in vitro cultures.
FIG 1 .

Setup of the E. coli/F. prausnitzii-diassociated mouse gnotobiotic model. (A and B) Inoculation of F. prausnitzii without E. coli. (A) Enumeration in feces of F. prausnitzii after gavage in germfree mice (n = 18 inoculations). (B) Scanning electron microscopy images from cecum of F. prausnitzii-infected mice. (C, D, and E) Inoculation of F. prausnitzii with E. coli. (C) Establishment of E. coli (open circles) and F. prausnitzii (solid squares) in the GIT of mice (n = 29) was monitored weekly by enumeration in the feces; arrows represent the various inoculations with F. prausnitzii before successful colonization. (D) Scanning electron microscopy images of E. coli (thick arrow) and F. prausnitzii (thin arrow) in the cecum of E. coli/F. prausnitzii-diassociated mice. (E) Representation along the GIT of E. coli (white bar) and F. prausnitzii (black bars) in CFU/g of content in diassociated mice.
We thus decided to use E. coli as a bacterial companion in order to get stable F. prausnitzii-colonized mice (hereafter named the E. coli/F. prausnitzii-diassociated mouse model). E. coli strain JM105 rapidly and stably colonized mice gut during at least 4 weeks after oral inoculation (see Fig. S1 in the supplemental material). By “priming” the GIT of mice with E. coli for at least 6 days, a durable and stable implantation of F. prausnitzii was systematically obtained in all mice (n = 29) (Fig. 1C). The population of F. prausnitzii was 10-fold lower (3.4 × 108 to 2 × 109 CFU/g of feces) than that of E. coli (2 × 109 to 1.2 × 1010 CFU/g of feces). Because of their different morphologies, the two bacterial species E. coli and F. prausnitzii could easily be distinguished in the cecal content (Fig. 1D). In addition F. prausnitzii, as a strict anaerobe species, was only mainly present from the cecum to the rectum, whereas E. coli was gradually abundant from the stomach to the colon (Fig. 1E).
Impact of F. prausnitzii on the metabolite profile during inflammation in the gnotobiotic mouse model.
We compared 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced colitis in either E. coli-monoassociated or E. coli/F. prausnitzii-diassociated mice in order to establish a metabolite profile under conditions allowing F. prausnitzii to display its anti-inflammatory properties (Fig. 2A). To decipher the impact of F. prausnitzii, after TNBS induction of colitis, metabolite profiling was performed with ileum, cecum, and colon contents, feces, and serum samples using gas chromatography-time of flight (GC/TOF) mass spectrometry (see Text S1 in the supplemental material) (Fig. 2B). Using a system that combines mass spectrometric and biological metadata, 983 metabolic features were validated. The relational database system BinBase was used for automated metabolites annotation (19, 20), with a criterion of presence in 50% or more of the biological replicates (see Text S1 and Table S1 in the supplemental material). Using a stringent 3-fold-intensity criterion, the number of metabolites distinguishing E. coli-monoassociated and E. coli/F. prausnitzii-diassociated animals greatly varied, depending on the GIT segment (Fig. 2B; see Table S1). In the upper part of the GIT, 86 and 142 metabolites were overrepresented in the ileum and cecum, respectively, in the E. coli-monoassociated mice. Conversely, 187 and 482 metabolites were more abundant in the colon and feces from E. coli/F. prausnitzii-diassociated mice, suggesting a metabolic imprinting correlated to the presence of F. prausnitzii (Fig. 1E). A total of 279 metabolic signals were structurally identified and assigned to a compound involved in known chemical reactions (as shown by KEGG number in Table S1). Interestingly, the metabolites identified in the colon mapped onto distinct functional categories (amino acids, energy metabolism, SCFA, lipids, phenolic acid, and sugars), indicating a broad metabolic signature of F. prausnitzii (Table 1). Compared to previous metabolomic data (21), our E. coli/F. prausnitzii-diassociated model displayed many classes of molecules present in conventional animals but absent in axenic animals as energy metabolites, lipids and fatty acids, amino acids, and sugars. Moreover, three metabolites are common: palmitic acid, 3,4-dihydroxyhydrocinnamic acid, and glucuronic acid.
FIG 2 .
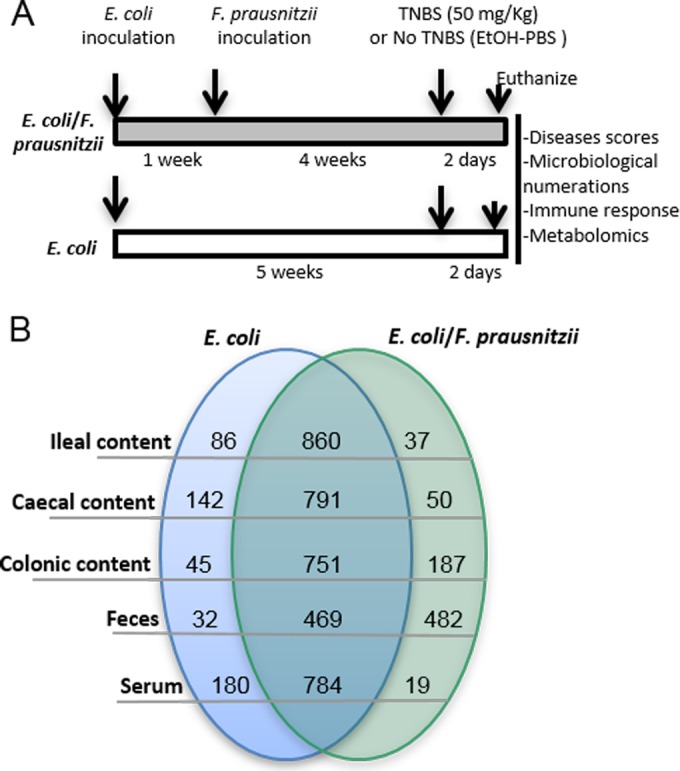
Experimental design of TNBS-induced colitis and metabolomic profiling. (A) Protocol of TNBS induction of colitis in E. coli-monoassociated and E. coli/F. prausnitzii-diassociated BALB/c mice. Different readouts are indicated in the box. (B) Numbers and comparison, using a stringent 3-fold-intensity criterion, of metabolites in the GIT segments and in the serum of E. coli-monoassociated and E. coli/F. prausnitzii-diassociated mice after a TNBS-induced colitis, identified by mass spectrometric and biological metadata.
TABLE 1 .
Known metabolites statistically overrepresented in the colon of E. coli/F. prausnitzii-diassociated mice compared to E. coli-monoassociated mice after TNBS-induced colitis
| Class | Metabolite | KEGG/PubChem ID no. | Dixenic/monoxenic intensity ratio | SEM | P value |
|---|---|---|---|---|---|
| Amino acids | Spermidine minor | C00315 | 7.675 | 0.69 | 0.009 |
| N-Acetylaspartic acid | SID 136743417 | 5.472 | 0.29 | 0.000 | |
| Quinic acid | C00296 | 3.682 | 0.30 | 0.019 | |
| O-Acetylserine | C00979 | 2.821 | 0.12 | 0.0005 | |
| Isothreonic acid | SID 103172351 | 2.220 | 0.17 | 0.047 | |
| Threonic acid | C01620 | 2.120 | 0.11 | 0.012 | |
| Homocystine | C01817 | 0.490 | 0.06 | 0.031 | |
| 2-Ketoisocaproic acid | C00233 | 0.402 | 0.05 | 0.003 | |
| Energy metabolites | Phosphoric acid | C00009 | 5.334 | 0.39 | 0.003 |
| Glucose-6-phosphate | C00092 | 2.354 | 0.11 | 0.005 | |
| Fructose-6-phosphate | C00085 | 2.0364 | 0.10 | 0.015 | |
| Glycerol | C00116 | 2.0206 | 0.07 | 0.003 | |
| β-Sitosterol | C01753 | 0.0449 | 0.13 | 0.020 | |
| Lipids and fatty acids | 4-Hydroxybutyric acid | C00989 | 3.277 | 0.16 | 0.000 |
| Palmitic acid | C00249 | 2.765 | 0.17 | 0.014 | |
| Caprylic acid | SID 11528290 | 2.637 | 0.12 | 0.001 | |
| Ethanolamine | C00189 | 4.854 | 0.22 | 0.000 | |
| Phytol | C01389 | 2.4277 | 0.15 | 0.015 | |
| Squalene | C00751 | 2.004 | 0.13 | 0.046 | |
| Phenolic acids | Idonic acid | SID 135170473 | 3.779 | 0.40 | 0.049 |
| trans-Sinapinic acid | C00482 | 2.618 | 0.14 | 0.003 | |
| Syringic acid | C10833 | 2.180 | 0.11 | 0.008 | |
| Malonic acid | C04025 | 2.131 | 0.14 | 0.037 | |
| 3-Phenyllactic acid | C05607 | 2.080 | 0.10 | 0.011 | |
| 3,4-Dihydroxyhydrocinnamic acid | SID 111677818 | 0.269 | 0.06 | 0.001 | |
| Pyrimidine | Pseudouridine | C02067 | 2.556 | 0.17 | 0.019 |
| Orotic acid | C00295 | 0.279 | 0.09 | 0.011 | |
| Sugars | Fructose | C00095 | 5.023 | 0.29 | 0.001 |
| Mannose | C00159 | 2.896 | 0.23 | 0.025 | |
| Glyceric acid | C00258 | 2.644 | 0.10 | 0.000 | |
| N-Acetyl-d-hexosamine | C03136 | 2.591 | 0.19 | 0.036 | |
| 1,5-Anhydroglucitol | C07326 | 2.484 | 0.18 | 0.027 | |
| β-Gentiobiose | C08240 | 2.423 | 0.11 | 0.003 | |
| Arabinose | C00259 | 2.356 | 0.11 | 0.005 | |
| Tagatose | SID 163300922 | 2.240 | 0.16 | 0.037 | |
| Ribonic acid | C01685 | 2.187 | 0.11 | 0.013 | |
| 2-Deoxyerythritol | SID 108236 | 2.002 | 0.11 | 0.023 | |
| Glucuronic acid | C00191 | 0.321 | 0.02 | 0.000 | |
| Glycero-guloheptose | SID 117585658 | 0.085 | 0.07 | 0.001 | |
| Raffinose | C00492 | 0.024 | 0.15 | 0.027 | |
| Inulotriose | SID 4849 | 0.004 | 0.03 | 0.000 | |
| Other | Indole-3-lactate | C02043 | 5.427 | 0.39 | 0.003 |
| Shikimic acid | C00493 | 2.220 | 0.13 | 0.017 | |
| Putrescine | C00134 | 0.487 | 0.05 | 0.015 |
Protective effect of F. prausnitzii on TNBS-induced colitis is linked to specific metabolic signatures in the colon.
In the E. coli/F. prausnitzii-diassociated mice, all biological markers indicated a decrease of the inflammation level in the presence of F. prausnitzii. After colitis induction, weight loss tended to decrease in the presence of F. prausnitzii (−3.2% ± 1.9% compared to −5.2% ± 1.5%). The disease activity index (DAI), Wallace macroscopic scores, and Ameho microscopic scores (architectural derangements, goblet cell depletion, edema/ulceration, and degree of inflammatory cell infiltrates) were significantly (P < 0.001) decreased in the colons of E. coli/F. prausnitzii-diassociated mice (Fig. 3A, B, and C). A severe disruption of the epithelial architecture was observed in E. coli-infected, TNBS-treated mice in contrast to E. coli/F. prausnitzii-diassociated, TNBS-treated mice (Fig. 3D). E. coli-infected TNBS-treated mouse colon epithelium was characterized by the absence of crypts and massive immune cell infiltrations. Such tissue erosions were less marked in the presence of F. prausnitzii. All of these observations indicated a protective effect of F. prausnitzii in our TNBS-induced-colitis model. Moreover, the F. prausnitzii population level was inversely correlated to the disease activity markers (weight loss, DAI, and Wallace and Ameho scores), suggesting that its anti-inflammatory effects could be dose dependent (see Fig. S2 in the supplemental material).
FIG 3 .
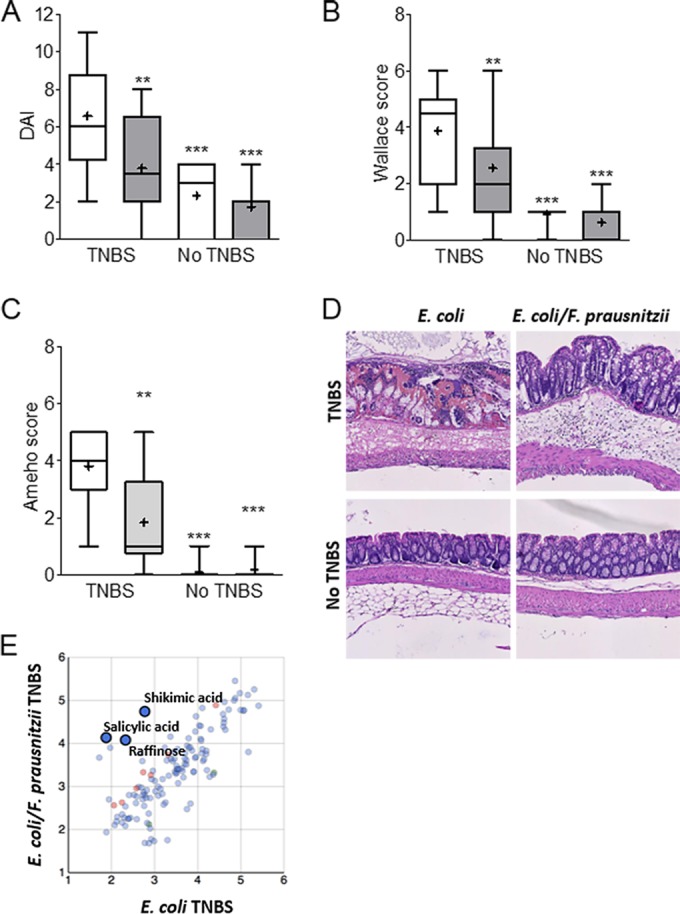
Analysis of colonic responses to a TNBS-induced colitis. (A) Disease activity index (DAI), (B) Wallace macroscopic score, (C) Ameho histologic score, (D) representative hematoxylin-eosin-safran staining of colonic tissue sections from E. coli-monoassociated (white bars) (n = 16 and 12) and E. coli/F. prausnitzii-diassociated (gray bars) (n = 18 and 11) mice with or without colitis, respectively. (E) Metabolomic analysis of E. coli-monoassociated (n = 8) compared to E. coli/F. prausnitzii-diassociated (n = 9) mouse colon content after TNBS treatment. The values are expressed as box-and-whisker plots with interquartile range, with the top portion of the box representing the 75th percentile and the bottom portion representing the 25th percentile. The horizontal bar within the box represents the median, with the mean shown as “+.” Asterisks indicate significant differences: **, P ≤ 0.01; ***, P ≤ 0.001. Metabolomics results are expressed as log intensities of metabolites with KEGG ID (t test, red and green, P ≤ 0.05).
Regarding metabolic profiles, we obtained strong evidence of the magnitude of the effect of an E. coli/F. prausnitzii microbial context in the TNBS-treated animals. The rarefaction curve showed that 80% of the known metabolites were more represented in the colon content of dixenic mice (see Fig. S3 in the supplemental material). We searched for metabolites segregating the monoxenic and dixenic animals after induction of colitis (t test on the mean). For instance, in the colon, butyrate (4-hydroxybutyric acid) (P = 0.0006), shikimic acid (P = 0.017) related to the salicylic acid pathway (P = 0.2055), and indole-3-lactate (P = 0.0035) were overrepresented in the E. coli/F. prausnitzii-diassociated mice (Table 1 and Fig. 3E). The metabolomic profiles sharply differed according the gut segment (Fig. 2B). For instance, raffinose was more discriminatory in the ileum (P = 0.0091) (data not shown) than in the colon (P = 0.1703) and was negatively related to F. prausnitzii abundance (Fig. 1E).
F. prausnitzii is a metabolically active species in vivo.
The beneficial mitigation effects, observed locally in the colon in the presence of F. prausnitzii, were confirmed in the serum, where systemic inflammation markers were reduced. First, the degree of infiltration by polymorphonuclear neutrophils (estimated by tissue myeloperoxidase [MPO] activity) was significantly decreased during TNBS-induced colitis in the presence of F. prausnitzii (P < 0.001) (Fig. 4A). In the presence of F. prausnitzii after TNBS induction of colitis, the serum cytokine response was globally similar to the one measured without TNBS (see Table S2 in the supplemental material). Interestingly, in the serum, the levels of proinflammatory cytokines, such as interleukin-1α (IL-1α), IL-2, and IL-5, were statistically decreased to a level corresponding to a noninflammatory status (Fig. 4B).
FIG 4 .

Analysis of serum immune response after TNBS-induced colitis. (A) MPO activity from E. coli-monoassociated (white bars) (n = 16 and 12) and E. coli/F. prausnitzii-diassociated (gray bars) (n = 18 and 11) mice with or without colitis, respectively. (B) Blood cytokine levels (IL-1α, IL-2, and IL-5) from E. coli-monoassociated (n = 9 and 8) and E. coli/F. prausnitzii-diassociated (n = 10 and 8) mice with or without colitis, respectively. (C) Venn diagram depicting common metabolites between gut contents (cecum, colon, ileum, and feces) and serum (serum, colon, ileum, and feces). (D) Serum metabolites from E. coli-monoassociated (n = 8) compared to E. coli/F. prausnitzii-diassociated (n = 10) mice after TNBS treatment. Values are expressed as the mean ± SEM. Asterisks indicate significant differences: *, P ≤ 0.05; **, P ≤ 0.01. Metabolomic data are expressed as log intensities of metabolites with KEGG ID (t test, red and green, P ≤ 0.05).
The high number of metabolites detected in the serum samples was noteworthy (Fig. 2B; see Table S1 in the supplemental material). Nineteen metabolites were overrepresented in the serum of E. coli/F. prausnitzii-diassociated mice. Seven were only found in the serum, and 11 were also found in the colon, where F. prausnitzii was most prevalent. Four metabolites were present in ileum, cecum, colon, and feces, but they are so far unknown (BinBase no. ID436364, -269147, -200610, and -310063) (Fig. 4C). The serum metabolome shared 5 entities with both colon and feces (uric acid, elaidic acid, lyxose, and unknown compounds BinBase ID231544 and ID459916). As observed in the colon content, the presence of F. prausnitzii is correlated to high levels of salicylic acid (P = 0.367) and shikimic acid (P = 0.4328) (Fig. 4D). Strikingly, α-ketoglutaric (AKG) acid was 10-fold more abundant in the serum of E. coli/F. prausnitzii-diassociated mice (P = 0.028), together with the linked citric acid and uric acid pathway (P = 0.085 and P = 0.0733, respectively). AKG acid is important in ammonia recycling and is reported to be depleted during gastrointestinal dysbiosis (22). Its involvement in cell proliferation and differentiation through the activity of the AKG-acid-dependent enzymes has recently been reviewed (23). The concomitant analysis of salicylic, shikimic, and AKG acids as standard in the GC mix validated their identification.
The four metabolites specifically associated with the presence of F. prausnitzii possess anti-inflammatory capacities in vitro.
We tested the immunomodulatory properties of four candidate molecules in a human colon adenocarcinoma cell line model (HT-29) by using F. prausnitzii supernatant, butyrate, and the 5-aminosalicylic acid (5-ASA) as controls. As shown in Fig. 5, salicylic acid (10 µM) blocked IL-8 production, as did butyrate (10 mM), F. prausnitzii supernatant, and 5-ASA (20 mM). Shikimic acid, raffinose, and AKG acid (100 µM) did not block IL-8 production, and a mixture of the different molecules did not show any additional effect (data not shown).
FIG 5 .
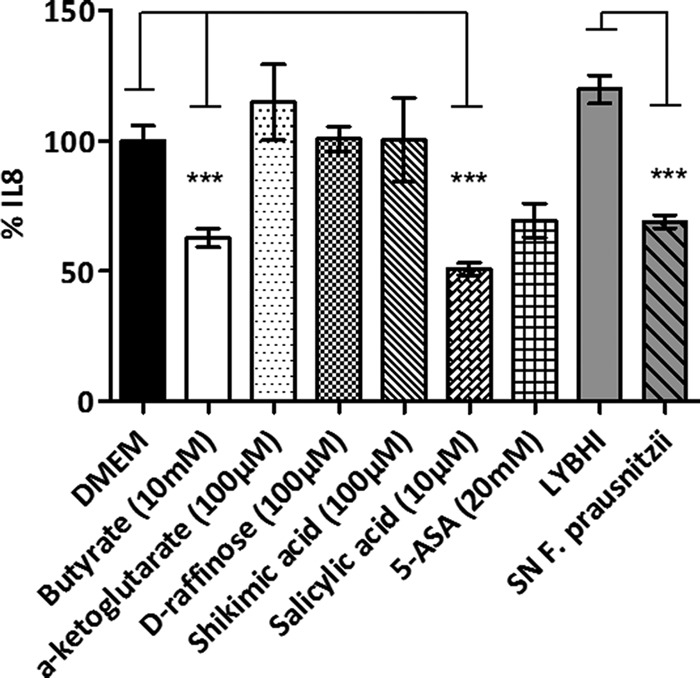
Immunomodulation capacities of F. prausnitzii supernatant and identified molecules in vitro. IL-8 production in TNF-α-stimulated HT-29 cells is shown. Experiments were done in triplicate. Results are expressed as picograms of IL-8 per milligram of protein and have been normalized with the IL-8 produced after the coincubation with PBS or LYBHI medium. SN, F. prausnitzii supernatant (20%). ***, P ≤ 0.001.
DISCUSSION
In this study, we show for the first time that F. prausnitzii is a highly active commensal bacterium involved in the reduction of colitis through in vivo modulation of metabolites along the GIT and in the peripheral blood. This novel approach clearly confirms the anti-inflammatory potential of F. prausnitzii and provides new clues about its ecological lifestyle and mechanisms in intestinal physiopathology.
We showed that efficient F. prausnitzii implantation requires the GIT preparation by E. coli, a primocolonizer of the sterile human newborn gut. Such an implantation kinetic is concordant with previous reports (15, 24) and with the late presence of F. prausnitzii in the human gut 7 months after birth (18). Thus, the time needed for F. prausnitzii colonization could correspond to the decrease of the redox potential (Eh), leading to more reduced conditions in the lower GIT (15, 25). However, other ecological factors should not be excluded, such as physiological modification of nutrient sources by the other species leading to the onset of an efficient trophic chain. Interestingly, sugars and energy metabolisms were deeply modified in the presence of F. prausnitzii (Table 1), suggesting a modification of their availability. While E. coli abundance gradually increased from stomach to colon, F. prausnitzii localization seemed to depend upon anaerobic conditions only found in the lower gut. These observations confirm that gradients in the physiochemical conditions along the GIT influence the biogeography of the colonic mucosal microbiome (26). Thus, our gnotobiotic rodents can be considered a relevant model to provide ecological information about GIT colonization kinetics by extremely oxygen-sensitive species. Moreover, we showed that the colonization pattern of F. prausnitzii in the GIT reflects its metabolic imprinting. F. prausnitzii, abundant in the lower GIT (cecum and colon) (Fig. 1E), had a gradually metabolic impact along the GIT (Fig. 2B). Even if F. prausnitzii was absent in the ileum, metabolic changes were observed in the ileum and at a distance in the serum (Fig. 4C and D). By using ileal content, we counted the F. prausnitzii population, and we cannot exclude that bacteria were present under the detection threshold or in a viable-but-not-culturable state. We detected 86 metabolites specific for E. coli in the ileum and 37 that are linked to the presence of E. coli and F. prausnitzii (Fig. 2B). Twenty-eight are unknown, and 9 could be identified (see Table S1 in the supplemental material). Among them are sugars, an antioxidant (trans-sinapinic acid), and neurotransmitter precursors (5-amino valeric acid lactone, a homologue of γ-aminobutyric acid [GABA] [27], and N-acetyl aspartic acid [28]). In the literature, an impact of F. prausnitzii on ileal physiology and particularly permeability has been already reported (29). These results, and similar findings reported by Velagalpuladi et al. (21), suggest that a direct or indirect effect of F. prausnitzii can be observed at a systemic level and not only where its population level is high.
During dysbiosis, the site-specific colonization and the dominance of F. prausnitzii in the colon are affected, which might contribute to the etiology of diseases of the large intestine (24). We have previously shown that F. prausnitzii presents anti-inflammatory properties in acute and chronic TNBS-induced-colitis models in conventional mice (7, 30), and this result was also observed in simplified gnotobiotic model. Moreover, our model allowed us to compare the population levels of E. coli and F. prausnitzii in feces to assess the impact of TNBS induction of colitis on bacterial populations (see Fig. S2 in the supplemental material). A TNBS challenge decreased the F. prausnitzii population, and the remaining level of F. prausnitzii clustered the mice into two groups (threshold of 108 F. prausnitzii cells/g of feces). Inflammation markers were decreased when the F. prausnitzii population was high. Because the level of colitis mitigation was directly correlated to its GIT abundance, our results confirmed that F. prausnitzii might be both a biomarker and a major factor of human health in adults (3). A review suggested that an increase in oxygen tension in the gut, related to the oxidative burst due to inflammatory reaction itself, could be the cause of the dysbiosis in IBD patients (25). Our results allowed us to hypothesize that the decreased F. prausnitzii level observed in IBD patients is a consequence of inflammation but also participates in the inflammation process, leading to the disease’s vicious circle (31).
In our simplified microbiota, we identified metabolites that can be produced either by the host or by the bacteria. Moreover, most of those metabolites are not assigned to any functions. This reflects the paucity of knowledge about reciprocal functional adaptation between host and microbes and the high percentage of microbial genes lacking annotation in metagenomic studies (1, 32). Li et al. had identified F. prausnitzii as an active bacterial species in humans, modifying butyrate-derived metabolites in urine (11). Using our metabolomic approach, we confirmed that the amount of 4-hydroxybutyric acid was increased in feces, colon, and cecum, where F. prausnitzii was established (see Table S1 in the supplemental material). Butyrate plays a major role in gut homeostasis, particularly through its anti-inflammatory activities in the colon mucosa (33, 34). This impact was confirmed in vitro using concentrations similar to those found in F. prausnitzii culture supernatants (10 mM) (Fig. 5). However, its role remains controversial since its effects seem to be dose and time dependent but also depend on the cellular model used (14, 35). In our study, butyrate specifically produced by F. prausnitzii (see Fig. S4A in the supplemental material) was not directly linked to health parameters (see Fig. S4B).
Based on our metabolomic approach, we identified metabolites specifically associated with the presence of F. prausnitzii. These findings support the concept that beneficial effects from commensals are linked to their large metabolic impact on host physiology. In fact, salicylic acid, at a concentration close to the one detected in the colon, decreased IL-8 production in vitro (Fig. 5). Interestingly, 40% of Fusobacterium prausnitzii (former nomenclature for F. prausnitzii) strains were able to ferment salicin, an alcoholic β-glucoside that can be converted into salicylic acid (36). In the pharmaceutical industry, salicylic acid is the precursor of 5-ASA, a drug used in the treatment of IBD (37). We showed that salicylic acid was as effective as 20 mM 5-ASA in vitro (38). Furthermore, we identified shikimic acid, a precursor for the biosynthesis aromatic compounds, including salicylic acid through the achorismate synthase pathway (39). Shikimic acid, upstream in the pathway of 5-ASA production, did not have the same anti-inflammatory effect in vitro. However, in vivo, it is demonstrated that 3,4-oxo-eisopropylideneshikimic acid (ISA), a derivative of shikimic acid, protects against TNBS-induced colonic damage in rats (40). We also identified raffinose, an oligosaccharide only fermented by the gut microbiota since monogastric animals do not express pancreatic α-galactosidase (41). Its uptake pathways are affected in celiac disease, whereas only transcellular uptake is affected in CD patients. Mucosal permeability has been found to be an important factor for raffinose permeation (42). Velagapudi et al. detected this metabolite in much higher abundance in GF than in conventional animals (21). F. prausnitzii impacts the host inflammation pattern but could also play a key role in maintaining gut permeability (29, 43), intestinal epithelial homeostasis (15), and energy balance, particularly through sugar pathway (44) (Table 1).
In contrast to the metabolites overrepresented in the presence of F. prausnitzii, underrepresented ones could be involved in anti-inflammatory process (Table 1). For instance, a significant decrease in homocystine was observed in the presence of F. prausnitzii. This amino acid results from the oxidation of homocysteine, which has been reported to be proinflammatory (45). Also, a decrease in putrescine together with an increase in spermidine in the dixenic mice could inhibit the secretion of inflammatory cytokines (46). Our approach provided a lot of hypotheses about the role of F. prausnitzii in intestinal physiopathology for future investigations.
The serum is a biofluid providing an excellent opportunity to assess the extent of interplay between bacterial metabolic and systemic human pathways (47). F. prausnitzii modulated the metabolome of the host blood by decreasing IL-1α, IL-2, and IL-5 concentrations. For instance, α-ketoglutaric (AKG) acid was statistically detected in the serum of mice colonized with F. prausnitzii and E. coli (Fig. 4). This organic acid, important for the proper metabolism of all essential amino acids and the transfer of cellular energy in the citric acid cycle, is a glutamic acid precursor involved in protein synthesis and the regulation of blood glucose levels (48). AKG did not show any significant effect on IL-8 production by HT-29 in our in vitro model (Fig. 5). This may mean that our conditions are not appropriate or that, as reported by Cairns and Mak (23), AKG acid may need a specific transporter/biochemical transformation before being active. Individuals with high protein intake, bacterial infections, or gastrointestinal dysbiosis may benefit from supplemental AKG acid to help metabolic health. Personalized health care investigations could be routinely set up to target inflammation biomarkers or check F. prausnitzii mitigation through salicylic, shikimic, and AKG acid levels in feces or serum.
To conclude, F. prausnitzii is a highly metabolic bacterium that impacts the host via different pathways to preserve its homeostasis. We have identified metabolites, particularly salicylic acid, involved in its anti-inflammatory capacities but also highlighted the complex and dynamic cross talk with the host under inflammation conditions.
MATERIALS AND METHODS
Bacterial culture.
F. prausnitzii A2-165 (DSM 17677), isolated from human fecal stool (49), was grown at 37°C in YBHI medium (brain-heart infusion medium supplemented with 0.5% yeast extract [Difco]) supplemented with cellobiose (1 mg/ml [Sigma]), maltose (1 mg/ml [Sigma]), and cysteine (0.5 mg/ml [Sigma]) in an anaerobic chamber filled with 85% N2, 10% CO2, and 5% H2. Escherichia coli JM105 was grown on LB medium (Difco) at 37°C.
Animals and experimental design.
Germfree 7- to 8-week-old BALB/c mice (male) were obtained from the germfree rodent breeding facilities of Anaxem-Micalis (INRA, Jouy-en-Josas, France). Animals were kept in flexible-film isolators (Getinge-La Calhène, Vendôme, France) in standard cages (5 mice/cage) with sterile wood shavings as animal bedding. Mice were given free access to autoclaved tap water and to a standard diet, R03-40 (Scientific Animal food and Engineering, Augy, France), sterilized by gamma irradiation at 45 kGy (IBA Mediris, Fleurus, Belgium). Isolators were maintained under controlled conditions of light (0730 to 1930 h), temperature (20 to 22°C), and humidity (45 to 55%). All procedures were carried out according to European Community Rules of Animal Care and with authorization 12-092 from the French Veterinary Services.
Inoculation of germfree mice with F. prausnitzii.
To obtain F. prausnitzii-monoassociated mice, germfree animals receive both intragastric and intrarectal inoculations of a 100× concentrated culture of F. prausnitzii A2-165 (109 CFU/inoculation). For intragastric gavages, mice were first treated with 0.1 ml of sodium bicarbonate (0.2 M) and then received 0.5 ml of the bacterial suspension 10 min later. For intrarectal inoculation, mice were first anesthetized by an intraperitoneal injection of a volume (1% of the body weight) of 10% ketamine hydrochloride (Imalgen 500 [Merial]) and 6% xylazine (Rompun [Bayer]) in 0.9% NaCl and then treated with 0.2 ml of the bacterial suspension.
To obtain dixenic E. coli/F. prausnitzii-diassociated mice, germfree mice were orally inoculated with a fresh culture of E. coli JM105 (108 to 109 CFU /ml). Twenty-four hours later, when E. coli was established in the GIT, 109 CFU of F. prausnitzii at 0.2 ml was administered to a donor mouse by intragastric gavage as described above. As soon as F. prausnitzii was established in the donor mouse, its feces were homogenized in water inside the isolator and immediately administered by intragastric gavage to other mice pretreated with sodium bicarbonate.
Viable bacteria in the inoculum were enumerated before and after gavage by plating of appropriate dilutions on supplemented YBHI agar in the anaerobic chamber. To enumerate the A2-165 strain in feces, fresh samples were introduced into the anaerobic chamber, serially 10-fold diluted, and plated on supplemented YBHI agar. In order to selectively enumerate A2-165 in the feces of dixenic mice by eliminating E. coli cells, fecal samples were treated with sodium azide (3%) for 5 min at 37°C and then plated on sodium azide (0.02%) YBHI agar.
TNBS-induced colitis.
Mice were weighed before TNBS administration and anesthetized by an intraperitoneal injection as described above. TNBS (Sigma-Aldrich) was dissolved in 0.9% NaCl–ethanol (50:50 vol/vol). Fifty microliters of TNBS (50 mg/kg body weight) was administered intrarectally (using a 3.5 French catheter [Solomon Scientific]) to mice maintained for 30 s in a vertical position. Control mice received 50 µl of 0.9% NaCl–ethanol solution intrarectally. Inflammation was monitored 48 h after TNBS administration.
Scanning electron microscopy.
Scanning electron microscopy analyses were performed at the MIMA2 platform (INRA, Massy, France) from 0.2 g of cecal samples as previously described (50).
Tissue preparation for histological analysis.
Flushed colons were opened longitudinally, cut into 2-cm sections, and rolled according to the Swiss roll procedure (51). The samples were fixed in 4% paraformaldehyde (4 h, room temperature), dehydrated, and embedded in paraffin according to standard histological protocols. Four-micrometer sections of distal colon were mounted on SuperFrost Plus slides. To assess TNBS-induced colitis, the sections were stained with hematoxylin-eosin and examined blindly according to the Ameho criteria (52).
Inflammation score assessment of TNBS-induced colitis.
The colon was removed without mesenteric fat and mesentery, carefully opened, and cleaned with phosphate-buffered saline (PBS). Colonic damage and inflammation were assessed blindly according to the Wallace score (53).
MPO activity.
MPO activity was measured using the method of Bradley et al. (54), modified as follows. Tissue samples were homogenized (50 mg/ml) in ice-cold 50-mM potassium phosphate buffer (pH 6) containing 5% hexadecyl trimethyl ammonium bromide (Sigma-Aldrich) and hydrogen peroxide. MPO is expressed in units per milligram of wet tissue.
Cytokine analysis.
Colonic samples were homogenized in 400 µl of Tris-HCl buffer containing protease inhibitors (Sigma-Aldrich) in a tissue lyser. Samples were centrifuged for 20 min, and the supernatant was frozen at −80°C until further analysis. Before sacrifice, blood samples were obtained from the retro-orbital venous plexus and centrifuged, and sera were stored at −80°C. Cytokine levels were determined by flow cytometry using Cytometric bead array analysis (mouse inflammation kit) (BD, NJ).
Metabolite extraction and analysis.
Fifty microliters of serum and feces, intestinal content, cecal content, and colonic content were lyophilized and subjected to 10 s of bead beating. Metabolite extraction was performed as previously described (20) with 700 µl of a precooled methanol-isopropanol-water (3:3:2) mixture in an ultrasonic water bath (5 min). After centrifugation, 650 µl of the supernatant was collected (5 min, 15,000 rpm) and dried before derivatization. Metabolite analysis was performed according to Fiehn et al. (55).
In vitro experiments with the HT-29 cell line.
The protocols used in this study were adapted from Kechaou et al. (56). The human colon carcinoma cell line HT-29 was cultured and stimulated with recombinant human tumor necrosis factor alpha (TNF-α) (5 ng/ml; PeproTech, NJ) as described previously. Cell incubations were done in the presence or not of different metabolites tested at the indicated concentration: butyric acid sodium salt, reference 303410, Sigma-Aldrich; shikimic acid, reference S5375, Sigma-Aldrich; salicylic acid, reference 84210, Sigma-Aldrich; d-(+)-raffinose pentahydrate, reference R0514, Sigma-Aldrich; α-ketoglutaric acid disodium salt dihydrate, reference 75892, Sigma-Aldrich; 5-aminosalicylic acid, reference 18858, Sigma-Aldrich. All samples were analyzed in triplicate. After coincubation, cell supernatants were collected and mixed with an antiprotease cocktail according to the manufacturer’s instructions (Complete EDTA-free protease inhibitor; Roche Applied Bioscience), and frozen at −80°C until further analysis of interleukin-8 (IL-8) concentrations by an enzyme-linked immunosorbent assay (ELISA) (BioLegend, San Diego, CA, and Mabtech, Sweden, respectively).
Statistical analysis.
Data are reported as means ± standard errors of the means (SEM) and for figures using box-and-whisker plots with the interquartile range, where the top portion of the box represents the 75th percentile and the lower portion represents the 25th percentile. The horizontal bar within the box represents the median, with the mean shown as “+.” The upper horizontal bar outside the box represents the 95th percentile, and the lower horizontal bar outside the box represents the 15th percentile. Statistical analysis was performed using one-way analysis of variance followed by Student-Newman-Keuls multiple comparison post hoc analysis, and with a P value of <0.05 considered significant. For metabolomic analysis, the resulting data sets were imported into the MatLab software for univariate and multivariate statistical analyses. Univariate statistics evaluated the number and percentage of features that are either unique or vary significantly between E. coli-monoassociated and E. coli/F. prausnitzii-diassociated animals. Venn diagrams were constructed as described in reference 57.
SUPPLEMENTAL MATERIAL
Supporting information. Shown are details of the methods used, including the type of segments and number of samples analyzed by GC/TOF mass spectrometry. Download
Model of E. coli monoxenic mice. Establishment of E. coli in the GIT of mice (n = 15) was monitored weekly by enumeration of bacterial counts in the feces; arrows represent the inoculation with E. coli. Download
Correlation between F. prausnitzii population level and colitis scores in BALB/c mice. (A) E. coli and F. prausnitzii levels in feces of diassociated mice before and after TNBS induction of colitis. (B) Weight variations, (C) disease activity index (DAI), and (D) Wallace macroscopic and Ameho histologic scores according to F. prausnitzii level in diassociated mouse feces (hatched bars, <108 F. prausnitzii cells/g of feces; black bars, >108 F. prausnitzii cells/g of feces) and E. coli-monoassociated mice (white bars) with TNBS-induced colitis. The values are expressed as the mean ± SEM. Asterisks indicate significant differences: *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. Download
Distribution curve of known metabolites identified in the colon content of TNBS-treated E. coli/F. prausnitzii-diassociated and E. coli-monoassociated mice. Download
SCFA production in cecal content of gnotobiotic mice according to colitis induction and F. prausnitzii levels. (A) Relative proportions of acetate and butyrate in the cecum of monoassociated (n = 15 and 12) and diassociated (n = 16 and 11) mice with or without TNBS-induced colitis, respectively. (B) Relative proportions of butyrate in the cecum of diassociated (n = 16 and 11) mice with or without TNBS-induced colitis considering the F. prausnitzii level in feces. The values are expressed as the mean ± SEM. Asterisks (*) indicate significant differences: *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. Download
Identification of metabolites increased in monoxenic (E. coli) versus dixenic (E. coli/F. prausnitzii) mice with TNBS-induced colitis
Colonic and blood cytokine levels in monoassociated and diassociated mice with or without TNBS-induced colitis
ACKNOWLEDGMENTS
This study was a part of the FPARIS collaborative project selected and supported by the Vitagora Competitive Cluster and funded by the French FUI (Fond Unique Interministériel; FUI no. F1010012D), the FEDER (Fonds Européen de Développement Régional; Bourgogne no. 34606), the Burgundy Region, the Conseil Général 21, and the Grand Dijon. This work was also supported by Merck Médication Familiale (Dijon, France) and Biovitis (Saint Étienne de Chomeil, France). R.M. and S.M. received a salary from the same grants.
We thank Sébastien Blugeon for technical assistance and Florian Chain and Olivier Berteau for fruitful discussions and critical reading of the manuscript.
Footnotes
Citation Miquel S, Leclerc M, Martin R, Chain F, Lenoir M, Raguideau S, Hudault S, Bridonneau C, Northen T, Bowen B, Bermúdez-Humarán LG, Sokol H, Thomas M, Langella P. 2015. Identification of metabolic signatures linked to anti-inflammatory effects of Faecalibacterium prausnitzii. mBio 6(2):e00300-15. doi:10.1128/mBio.00300-15.
REFERENCES
- 1.Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, Mende DR, Li J, Xu J, Li S, Li D, Cao J, Wang B, Liang H, Zheng H, Xie Y, Tap J, Lepage P, Bertalan M, Batto JM, Hansen T, Le Paslier D, Linneberg A, Nielsen HB, Pelletier E, Renault P, Sicheritz-Ponten T, Turner K, Zhu H, Yu C, Li S, Jian M, Zhou Y, Li Y, Zhang X, Li S, Qin N, Yang H, Wang J, Brunak S, Dore J, Guarner F, Kristiansen K, Pedersen O, Parkhill J, Weissenbach J, Bork P, Ehrlich SD, Wang J. 2010. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464:59–65. doi: 10.1038/nature08821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Tomas J, Wrzosek L, Bouznad N, Bouet S, Mayeur C, Noordine ML, Honvo-Houeto E, Langella P, Thomas M, Cherbuy C. 2013. Primocolonization is associated with colonic epithelial maturation during conventionalization. FASEB J 27:645–655. doi: 10.1096/fj.12-216861. [DOI] [PubMed] [Google Scholar]
- 3.Miquel S, Martín R, Rossi O, Bermúdez-Humarán LG, Chatel JM, Sokol H, Thomas M, Wells JM, Langella P. 2013. Faecalibacterium prausnitzii and human intestinal health. Curr Opin Microbiol 16:255–261. doi: 10.1016/j.mib.2013.06.003. [DOI] [PubMed] [Google Scholar]
- 4.Rajilić-Stojanović M, Biagi E, Heilig HG, Kajander K, Kekkonen RA, Tims S, de Vos WM. 2011. Global and deep molecular analysis of microbiota signatures in fecal samples from patients with irritable bowel syndrome. Gastroenterology 141:1792–1801. doi: 10.1053/j.gastro.2011.07.043. [DOI] [PubMed] [Google Scholar]
- 5.Sobhani I, Tap J, Roudot-Thoraval F, Roperch JP, Letulle S, Langella P, Corthier G, Tran Van Nhieu J, Furet JP. 2011. Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS One 6:e16393. doi: 10.1371/journal.pone.0016393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Swidsinski A, Loening-Baucke V, Vaneechoutte M, Doerffel Y. 2008. Active Crohn’s disease and ulcerative colitis can be specifically diagnosed and monitored based on the biostructure of the fecal flora. Inflamm Bowel Dis 14:147–161. doi: 10.1002/ibd.20330. [DOI] [PubMed] [Google Scholar]
- 7.Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermúdez-Humarán LG, Gratadoux JJ, Blugeon S, Bridonneau C, Furet JP, Corthier G, Grangette C, Vasquez N, Pochart P, Trugnan G, Thomas G, Blottière HM, Doré J, Marteau P, Seksik P, Langella P. 2008. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci U S A 105:16731–16736. doi: 10.1073/pnas.0804812105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sokol H, Seksik P, Furet JP, Firmesse O, Nion-Larmurier I, Beaugerie L, Cosnes J, Corthier G, Marteau P, Doré J. 2009. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflamm Bowel Dis 15:1183–1189. doi: 10.1002/ibd.20903. [DOI] [PubMed] [Google Scholar]
- 9.Miquel S, Martin R, Bridonneau C, Robert V, Sokol H, Bermúdez-Humarán L, Thomas M, Langella P. 2014. Ecology and metabolism of the beneficial intestinal commensal bacterium Faecalibacterium prausnitzii. Gut Microbes 5:146–151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hold GL, Schwiertz A, Aminov RI, Blaut M, Flint HJ. 2003. Oligonucleotide probes that detect quantitatively significant groups of butyrate-producing bacteria in human feces. Appl Environ Microbiol 69:4320–4324. doi: 10.1128/AEM.69.7.4320-4324.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li M, Wang B, Zhang M, Rantalainen M, Wang S, Zhou H, Zhang Y, Shen J, Pang X, Zhang M, Wei H, Chen Y, Lu H, Zuo J, Su M, Qiu Y, Jia W, Xiao C, Smith LM, Yang S, Holmes E, Tang H, Zhao G, Nicholson JK, Li L, Zhao L. 2008. Symbiotic gut microbes modulate human metabolic phenotypes. Proc Natl Acad Sci U S A 105:2117–2122. doi: 10.1073/pnas.0712038105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Louis P, Flint HJ. 2009. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol Lett 294:1–8. doi: 10.1111/j.1574-6968.2009.01514.x. [DOI] [PubMed] [Google Scholar]
- 13.Macfarlane GT, Macfarlane S. 2011. Fermentation in the human large intestine: its physiologic consequences and the potential contribution of prebiotics. J Clin Gastroenterol 45(Suppl):S120–S127. doi: 10.1097/MCG.0b013e31822fecfe. [DOI] [PubMed] [Google Scholar]
- 14.Martín R, Miquel S, Ulmer J, Kechaou N, Langella P, Bermúdez-Humarán LG. 2013. Role of commensal and probiotic bacteria in human health: a focus on inflammatory bowel disease. Microb Cell Fact 12:71. doi: 10.1186/1475-2859-12-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wrzosek L, Miquel S, Noordine ML, Bouet S, Joncquel Chevalier-Curt M, Robert V, Philippe C, Bridonneau C, Cherbuy C, Robbe-Masselot C, Langella P, Thomas M. 2013. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol 11:61. doi: 10.1186/1741-7007-11-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Duncan SH, Louis P, Thomson JM, Flint HJ. 2009. The role of pH in determining the species composition of the human colonic microbiota. Environ Microbiol 11:2112–2122. doi: 10.1111/j.1462-2920.2009.01931.x. [DOI] [PubMed] [Google Scholar]
- 17.Lopez-Siles M, Khan TM, Duncan SH, Harmsen HJ, Garcia-Gil LJ, Flint HJ. 2012. Cultured representatives of two major phylogroups of human colonic Faecalibacterium prausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth. Appl Environ Microbiol 78:420–428. doi: 10.1128/AEM.06858-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hopkins MJ, Macfarlane GT, Furrie E, Fite A, Macfarlane S. 2005. Characterisation of intestinal bacteria in infant stools using real-time PCR and Northern hybridisation analyses. FEMS Microbiol Ecol 54:77–85. doi: 10.1016/j.femsec.2005.03.001. [DOI] [PubMed] [Google Scholar]
- 19.Skogerson K, Wohlgemuth G, Barupal DK, Fiehn O. 2011. The volatile compound BinBase mass spectral database. BMC Bioinformatics 12:321. doi: 10.1186/1471-2105-12-321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lee do Y, Bowen BP, Nguyen DH, Parsa S, Huang Y, Mao JH, Northen TR. 2012. Low-dose ionizing radiation-induced blood plasma metabolic response in a diverse genetic mouse population. Radiat Res 178:551–555. doi: 10.1667/RR2990.1. [DOI] [PubMed] [Google Scholar]
- 21.Velagapudi VR, Hezaveh R, Reigstad CS, Gopalacharyulu P, Yetukuri L, Islam S, Felin J, Perkins R, Borén J, Oresic M, Bäckhed F. 2010. The gut microbiota modulates host energy and lipid metabolism in mice. J Lipid Res 51:1101–1112. doi: 10.1194/jlr.M002774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hamer HM, De Preter V, Windey K, Verbeke K. 2012. Functional analysis of colonic bacterial metabolism: relevant to health? Am J Physiol Gastrointest Liver Physiol 302:G1–G9. doi: 10.1152/ajpgi.00048.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cairns RA, Mak TW. 2013. Oncogenic isocitrate dehydrogenase mutations: mechanisms, models, and clinical opportunities. Cancer Discov 3:730–741. doi: 10.1158/2159-8290.CD-13-0083. [DOI] [PubMed] [Google Scholar]
- 24.Ahmed S, Macfarlane GT, Fite A, McBain AJ, Gilbert P, Macfarlane S. 2007. Mucosa-associated bacterial diversity in relation to human terminal ileum and colonic biopsy samples. Appl Environ Microbiol 73:7435–7442. doi: 10.1128/AEM.01143-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rigottier-Gois L. 2013. Dysbiosis in inflammatory bowel diseases: the oxygen hypothesis. ISME J 7:1256–1261. doi: 10.1038/ismej.2013.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Aguirre de Cárcer D, Cuív PO, Wang T, Kang S, Worthley D, Whitehall V, Gordon I, McSweeney C, Leggett B, Morrison M. 2011. Numerical ecology validates a biogeographical distribution and gender-based effect on mucosa-associated bacteria along the human colon. ISME J 5:801–809. doi: 10.1038/ismej.2010.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Callery PS, Geelhaar LA. 1984. Biosynthesis of 5-aminopentanoic acid and 2-piperidone from cadaverine and 1-piperideine in mouse. J Neurochem 43:1631–1634. doi: 10.1111/j.1471-4159.1984.tb06088.x. [DOI] [PubMed] [Google Scholar]
- 28.Yan HD, Ishihara K, Serikawa T, Sasa M. 2003. Activation by N-acetyl-l-aspartate of acutely dissociated hippocampal neurons in rats via metabotropic glutamate receptors. Epilepsia 44:1153–1159. doi: 10.1046/j.1528-1157.2003.49402.x. [DOI] [PubMed] [Google Scholar]
- 29.Carlsson AH, Yakymenko O, Olivier I, Håkansson F, Postma E, Keita AV, Söderholm JD. 2013. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice DSS colitis. Scand J Gastroenterol 48:1136–1144. doi: 10.3109/00365521.2013.828773. [DOI] [PubMed] [Google Scholar]
- 30.Martín R, Chain F, Miquel S, Lu J, Gratadoux JJ, Sokol H, Verdu EF, Bercik P, Bermúdez-Humarán LG, Langella P. 2014. The commensal bacterium Faecalibacterium prausnitzii is protective in DNBS-induced chronic moderate and severe colitis models. Inflamm Bowel Dis 20:417–430. doi: 10.1097/01.MIB.0000440815.76627.64. [DOI] [PubMed] [Google Scholar]
- 31.Sonnenberg A, Collins JF. 2006. Vicious circles in inflammatory bowel disease. Inflamm Bowel Dis 12:944–949. doi: 10.1097/01.mib.0000231577.19301.95. [DOI] [PubMed] [Google Scholar]
- 32.Human Microbiome Project Consortium 2012. Structure, function and diversity of the healthy human microbiome. Nature 486:207–214. doi: 10.1038/nature11234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Plöger S, Stumpff F, Penner GB, Schulzke JD, Gäbel G, Martens H, Shen Z, Günzel D, Aschenbach JR. 2012. Microbial butyrate and its role for barrier function in the gastrointestinal tract. Ann N Y Acad Sci 1258:52–59. doi: 10.1111/j.1749-6632.2012.06553.x. [DOI] [PubMed] [Google Scholar]
- 34.Fusunyan RD, Quinn JJ, Fujimoto M, MacDermott RP, Sanderson IR. 1999. Butyrate switches the pattern of chemokine secretion by intestinal epithelial cells through histone acetylation. Mol Med 5:631–640. [PMC free article] [PubMed] [Google Scholar]
- 35.Böcker U, Nebe T, Herweck F, Holt L, Panja A, Jobin C, Rossol S, Sartor RB, Singer MV. 2003. Butyrate modulates intestinal epithelial cell-mediated neutrophil migration. Clin Exp Immunol 131:53–60. doi: 10.1046/j.1365-2249.2003.02056.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Moore WE, Moore LV, Cato EP, Wilkins TD, Kornegay ET. 1987. Effect of high-fiber and high-oil diets on the fecal flora of swine. Appl Environ Microbiol 53:1638–1644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Messori A, Brignola C, Trallori G, Rampazzo R, Bardazzi G, Belloli C, d’Albasio G, De Simone G, Martini N. 1994. Effectiveness of 5-aminosalicylic acid for maintaining remission in patients with Crohn’s disease: a meta-analysis. Am J Gastroenterol 89:692–698. [PubMed] [Google Scholar]
- 38.Lee JS, Park SY, Thapa D, Choi MK, Chung IM, Park YJ, Yong CS, Choi HG, Kim JA. 2010. Grifola frondosa water extract alleviates intestinal inflammation by suppressing TNF-alpha production and its signaling. Exp Mol Med 42:143–154. doi: 10.3858/emm.2010.42.2.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bochkov DV, Sysolyatin SV, Kalashnikov AI, Surmacheva IA. 2012. Shikimic acid: review of its analytical, isolation, and purification techniques from plant and microbial sources. J Chem Biol 5:5–17. doi: 10.1007/s12154-011-0064-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Xing J, You C, Dong K, Sun J, You H, Dong Y, Sun J. 2013. Ameliorative effects of 3,4-oxo-isopropylidene-shikimic acid on experimental colitis and their mechanisms in rats. Int Immunopharmacol 15:524–531. doi: 10.1016/j.intimp.2013.02.008. [DOI] [PubMed] [Google Scholar]
- 41.LeBlanc JG, Silvestroni A, Connes C, Juillard V, de Giori GS, Piard JC, Sesma F. 2004. Reduction of non-digestible oligosaccharides in soymilk: application of engineered lactic acid bacteria that produce alpha-galactosidase. Genet Mol Res 3:432–440. [PubMed] [Google Scholar]
- 42.Dawson DJ, Lobley RW, Burrows PC, Notman JA, Mahon M, Holmes R. 1988. Changes in jejunal permeability and passive permeation of sugars in intestinal biopsies in coeliac disease and Crohn’s disease. Clin Sci (Lond) 74:427–431. [DOI] [PubMed] [Google Scholar]
- 43.Laval L, Martin R, Natividad J, Chain F, Miquel S, de Maredsous CD, Capronnier S, Sokol H, Verdu E, van Hylckama Vlieg J, Bermúdez-Humarán L, Smokvina T, Langella P. 2015. Lactobacillus rhamnosus CNCM I-3690 and the commensal bacterium Faecalibacterium prausnitzii A2-165 exhibit similar protective effects to induced barrier hyper-permeability in mice. Gut Microbes 6:1–9. doi: 10.4161/19490976.2014.990784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Heinken A, Khan MT, Paglia G, Rodionov DA, Harmsen HJ, Thiele I. 2014. Functional metabolic map of Faecalibacterium prausnitzii, a beneficial human gut microbe. J Bacteriol 196:3289–3302. doi: 10.1128/JB.01780-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Perła-Kaján J, Twardowski T, Jakubowski H. 2007. Mechanisms of homocysteine toxicity in humans. Amino Acids 32:561–572. doi: 10.1007/s00726-006-0432-9. [DOI] [PubMed] [Google Scholar]
- 46.Pérez-Cano FJ, Franch A, Castellote C, Castell M. 2003. Immunomodulatory action of spermine and spermidine on NR8383 macrophage line in various culture conditions. Cell Immunol 226:86–94. doi: 10.1016/j.cellimm.2003.09.009. [DOI] [PubMed] [Google Scholar]
- 47.Wikoff WR, Anfora AT, Liu J, Schultz PG, Lesley SA, Peters EC, Siuzdak G. 2009. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc Natl Acad Sci U S A 106:3698–3703. doi: 10.1073/pnas.0812874106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Waddell TG, Henderson BS, Morris RT, Lewis CM, Zimmermann AG. 1987. Chemical evolution of the citric acid cycle: sunlight photolysis of alpha-ketoglutaric acid. Orig Life Evol Biosph 17:149–153. doi: 10.1007/BF01808242. [DOI] [PubMed] [Google Scholar]
- 49.Duncan SH, Hold GL, Harmsen HJ, Stewart CS, Flint HJ. 2002. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int J Syst Evol Microbiol 52:2141–2146. doi: 10.1099/ijs.0.02241-0. [DOI] [PubMed] [Google Scholar]
- 50.Joly F, Mayeur C, Bruneau A, Noordine ML, Meylheuc T, Langella P, Messing B, Duée PH, Cherbuy C, Thomas M. 2010. Drastic changes in fecal and mucosa-associated microbiota in adult patients with short bowel syndrome. Biochimie 92:753–761. doi: 10.1016/j.biochi.2010.02.015. [DOI] [PubMed] [Google Scholar]
- 51.Rochat T, Bermúdez-Humarán L, Gratadoux JJ, Fourage C, Hoebler C, Corthier G, Langella P. 2007. Anti-inflammatory effects of Lactobacillus casei BL23 producing or not a manganese-dependent catalase on DSS-induced colitis in mice. Microb Cell Fact 6:22. doi: 10.1186/1475-2859-6-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ameho CK, Adjei AA, Harrison EK, Takeshita K, Morioka T, Arakaki Y, Ito E, Suzuki I, Kulkarni AD, Kawajiri A, Yamamoto S. 1997. Prophylactic effect of dietary glutamine supplementation on interleukin 8 and tumour necrosis factor alpha production in trinitrobenzene sulphonic acid induced colitis. Gut 41:487–493. doi: 10.1136/gut.41.4.487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wallace JL, MacNaughton WK, Morris GP, Beck PL. 1989. Inhibition of leukotriene synthesis markedly accelerates healing in a rat model of inflammatory bowel disease. Gastroenterology 96:29–36. [DOI] [PubMed] [Google Scholar]
- 54.Bradley PP, Priebat DA, Christensen RD, Rothstein G. 1982. Measurement of cutaneous inflammation: estimation of neutrophil content with an enzyme marker. J Invest Dermatol 78:206–209. doi: 10.1111/1523-1747.ep12506462. [DOI] [PubMed] [Google Scholar]
- 55.Fiehn O, Kopka J, Dörmann P, Altmann T, Trethewey RN, Willmitzer L. 2000. Metabolite profiling for plant functional genomics. Nat Biotechnol 18:1157–1161. doi: 10.1038/81137. [DOI] [PubMed] [Google Scholar]
- 56.Kechaou N, Chain F, Gratadoux JJ, Blugeon S, Bertho N, Chevalier C, Le Goffic R, Courau S, Molimard P, Chatel JM, Langella P, Bermúdez-Humarán LG. 2013. Identification of one novel candidate probiotic Lactobacillus plantarum strain active against influenza virus infection in mice by a large-scale screening. Appl Environ Microbiol 79:1491–1499. doi: 10.1128/AEM.03075-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Oliveros JC. 2007. An interactive tool for comparing lists with Venn diagrams. http://bioinfogp.cnb.csic.es/tools/venny/index.html.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information. Shown are details of the methods used, including the type of segments and number of samples analyzed by GC/TOF mass spectrometry. Download
Model of E. coli monoxenic mice. Establishment of E. coli in the GIT of mice (n = 15) was monitored weekly by enumeration of bacterial counts in the feces; arrows represent the inoculation with E. coli. Download
Correlation between F. prausnitzii population level and colitis scores in BALB/c mice. (A) E. coli and F. prausnitzii levels in feces of diassociated mice before and after TNBS induction of colitis. (B) Weight variations, (C) disease activity index (DAI), and (D) Wallace macroscopic and Ameho histologic scores according to F. prausnitzii level in diassociated mouse feces (hatched bars, <108 F. prausnitzii cells/g of feces; black bars, >108 F. prausnitzii cells/g of feces) and E. coli-monoassociated mice (white bars) with TNBS-induced colitis. The values are expressed as the mean ± SEM. Asterisks indicate significant differences: *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. Download
Distribution curve of known metabolites identified in the colon content of TNBS-treated E. coli/F. prausnitzii-diassociated and E. coli-monoassociated mice. Download
SCFA production in cecal content of gnotobiotic mice according to colitis induction and F. prausnitzii levels. (A) Relative proportions of acetate and butyrate in the cecum of monoassociated (n = 15 and 12) and diassociated (n = 16 and 11) mice with or without TNBS-induced colitis, respectively. (B) Relative proportions of butyrate in the cecum of diassociated (n = 16 and 11) mice with or without TNBS-induced colitis considering the F. prausnitzii level in feces. The values are expressed as the mean ± SEM. Asterisks (*) indicate significant differences: *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. Download
Identification of metabolites increased in monoxenic (E. coli) versus dixenic (E. coli/F. prausnitzii) mice with TNBS-induced colitis
Colonic and blood cytokine levels in monoassociated and diassociated mice with or without TNBS-induced colitis