

Abstract
Phototoxicity consists in the capability of certain innocuous molecules to become toxic when subjected to suitable illumination. In order to discover new photoactive drugs or characterize phototoxic pollutants, it would be advantageous to use simple biological tests of phototoxicy. In this work, we present a pilot screening of 37 dyes to test for phototoxic effects in the roundworm Caenorhabditis elegans. Populations of this nematode were treated with different dyes, and subsequently exposed to 30 min of white light. Behavioral outcomes were quantified by recording the global motility using an infrared tracking device (WMicrotracker). Of the tested compounds, 17 dyes were classified as photoactive, being phloxine B, primuline, eosin Y, acridine orange and rose Bengal the most phototoxic. To assess photoactivity after uptake, compounds were retested after washing them out of the medium before light irradiation. Dye uptake into the worms was also analyzed by staining or fluorescence. All the positive drugs were incorporated by animals and produced phototoxic effects after washing. We also tested the stress response being triggered by the treatments through reporter strains. Endoplasmic reticulum stress response (hsp-4::GFP strain) was activated by 22% of phototoxic dyes, and mitochondrial stress response (hsp-6::GFP strain) was induced by 16% of phototoxic dyes. These results point to a phototoxic perturbation of the protein functionality and an oxidative stress similar to that reported in cell cultures. Our work shows for the first time the feasibility of C. elegans for running phototoxic screenings and underscores its application on photoactive drugs and environmental pollutants assessment.
Introduction
The phototoxic properties of many compounds were demonstrated long ago [1], consisting in the capability of certain molecules to be activated when subjected to suitable illumination. When this effect requires molecular oxygen it has been named photodynamic action, and allowed the development of a new cancer treatment called “photodynamic therapy” (“PDT”) [2]. In a PDT treatment patients are treated with the drug and, once absorbed, the tumor area is subjected to light irradiation in order to activate drug phototoxicity and destroy cancer cells. The molecular mechanism of photoactivation is based on the generation of reactive oxygen species (ROS, and mainly singlet oxygen, 1O2), which damage cellular structures and induce cell death, either by apoptosis or necrosis [3–5]. Specific photoactivable drugs have been discovered and are currently used in human healthcare [6], and several common dyes have shown photodynamic activity. Some well characterized molecular structures with photoactive properties are thiazine [7–9], xanthene [10], acridine [11, 12], and triarylmethane [13] derivatives.
At the moment, most of the work on photoactivity of dyes has been concerned with biomedical applications [14], particularly PDT of cancer [4, 15, 16]. However, this particular effect spreads to other fields of interest such as photosterilization of water and blood products [9, 12], and environmental pollution [17–20]. It is worth to note that dyes used in industrial activities often present mutagenic, carcinogenic, and genotoxic photoactivity [15, 21, 22], which can result in a relevant risk for both human and environmental health.
In addition, xenobiotics such as herbicides, pesticides, aromatic hydrocarbons, cosmetics and personal care products are also photoactive environmental pollutants that, when illuminated, exert adverse effects on the quality of river and lake water, soil sediments and living organisms [23–29].
Biological models, from microorganisms to higher organisms, have been thoroughly employed to evaluate phototoxic effects of chemicals in order to explore the potential of new drugs for PDT or to prevent environmental damage from xenobiotics. Several bioassays for the detection of photodynamic effects are currently on use, examples being Paramecium [30], Candida [31], Allium [32], Drosophila [22], Daphnia, sea urchin [10], amphibian embryos [33], cell cultures [34], erythrocyte hemolysis [35], and intradermal tests [36].
In order to assess more precisely the phototoxicity of compounds such as drugs or pollutants, the use of a small translucent whole organism seems to be advantageous for simple, rapid and cheap bioassays. One of the simplest animal models is the nematode Caenorhabditis elegans, which has a fully sequenced genome, rudimentary organs similar to those found in mammals, and highly conserved cellular signaling pathways [37]. Also, the ease to culture, growth and reproduction of this organism has helped to establish it as a very suitable biological model for a variety of assessments. The main attraction of C. elegans for use in pharmacological and toxicological screening consists in the capability to be cultured in microplates (96 or 384 well plates), with small amounts of liquid medium, where animals can be incubated with experimental compounds [38].
In a whole animal approach, treatment with toxic doses will cause death, whereas subtoxic effects will affect animal behavior. Based on this premise, we employed here a motility tracking system, previously developed in our laboratory [39], to characterize the effects of thirty seven dyes, some already known to have phototoxic properties. As far as we know, this is the first time that the C. elegans model has been used for screening of this kind of compound.
Materials and Methods
Animal culture and strains
For the phototoxicity, uptake assays, and for qRT-PCR experiments, synchronized populations of Caenorhabditis elegans SS104 strain (glp-4 [bn2], a temperature sensitive sterile mutant) were cultured at non permissive temperature (25°C) in Nematode Growth Medium as described previously [40]. On the 1st day of adult stage, animals were washed twice in saline solution (modified K-medium: 52 mM NaCl and 32 mM KCl [41] + 0.01% Triton X-100) and transferred to flat bottomed 96-well microplates (Greiner Bio-one Cat. 655101). An average of 50 animals was used per well with a final volume of 100 μl.
For the stress response assays the reporter lines used were zcIs4 [hsp-4::GFP], zcIs9 [hsp-60::GFP], zcIs13 [hsp-6::GFP] (SJ4005, SJ4100 and SJ4058 strains, respectively). For worm cultures the same protocol was followed but at 20°C. All strains were obtained from the Caenorhabditis Genetic Center, University of Minnesota, USA.
Behavioral phototoxicity measurement
One hour after pipetting, basal motility of worms within the wells was assessed using an infrared tracking device (WMicrotracker, Designplus SRL, Argentina) [39]. This basal activity was recorded to normalize any subsequent activity variations to that initial activity, eliminating differences between wells due to population size. After basal measurement, dyes were added to the culture medium at final concentrations of 0.1, 1, 10 or 100 μM and incubated for 1 h before light irradiation (where applied). Dye photoactivation was carried out exposing the microplates for 30 min under a fluorescent white light source (2700°K white fluorescent lamp R7s 20W, Sica, Argentina) at 10 mW/cm2 of intensity. An additional water IR-filter (3-cm wide) was used to avoid heating, as previously reported [20].
Locomotor activity was tracked continuously in darkness during the incubation with the dye and for 4 h after dye activation. At least 4 replicate wells were used for each experiment, and the reported concentration was repeated independently three times, unless otherwise mentioned in the text.
Chemical treatment
The dyes employed in this study, as well as their characteristics and known properties, are shown in Table 1. After a preliminary determination, compounds were used at 100 μM concentration (unless otherwise indicated).
Table 1. List of dyes used in the present work, with reference to the chemical group, Colour Index number (C.I.), electric charge (Z), source, and previously reported photoactivity (if any, and only for well known cases).
| Dye (abbreviation) | Chemical group | C.I. | Z | Source | Photo Activity | Reference a |
|---|---|---|---|---|---|---|
| 1. Acridine orange (AO) | Acridine | 46005 | + | Merck | + | Zdolsek et al. 1990, Herkovits et al. 2007, Alvarez el al. 2011 |
| 2. Alizarin red S (ARS) | Anthraquinone | 58005 | − | Fluka | − | Seliger & McElroy 1965, Redmond & Gamlin 1999 |
| 3. Auramine O (AuO) | Diphenylmethane | 41000 | + | Analema | + | Stockert et al. 1990 |
| 4. Berberine (Ber) | Isoquinoline | 75169 | + | Sigma | + | Molero et al. 1985, Inbaraj et al. 2001 |
| 5. Bismarck brown Y (BBY) | Bis-azo | 21000 | 2+ | R.A. Lamb | − | Barbosa & Peters 1971 |
| 6. Carmine (Car) | Anthraquinone | 75470 | 0 | Gurr | nd | |
| 7. Chrysoidine (Chr) | Mono-azo | 11270 | + | G.T. Gurr | nd | |
| 8. Congo red (CR) | Bis-azo | 22120 | 2− | Merck | nd | |
| 9. Curcumin (Cur) | β-Diketone | 75300 | 0 | Merck | + | Bruzell et al. 2005 |
| 10. Eosin Y (EY) | Hydroxyxanthene | 45380 | 2− | Sigma | + | Seliger & McElroy 1965, Knox & Dodge 1985, Deerinck et al. 1994 |
| 11. Fast green FCF (FG) | Triphenylmethane | 42053 | 2− | Sigma | nd | |
| 12. Fluorescein (F) | Hydroxyxanthene | 45350 | 2− | Sigma | − | Gandin et al. 1983, Devanathan et al. 1990 |
| 13. Hematoxylin (H) | Neoflavone | 75290 | − | Panreac | nd | |
| 14. Indigocarmine (IC) | Indigoid | 73015 | 2− | Serva | − | Herkovits et al. 2007 |
| 15. Janus green B (JGB) | Mono-azo-azine | 11050 | + | Grübler | nd | |
| 16. Luxol fast blue MBSN b | Phthalocyanine | 74180 | 2− | Merck | − d | Redmond & Gamlin 1999 |
| 17. Mercurochrome (Mer) | Hydroxyxanthene | − | 2− | Merck | + | Redmond & Gamlin 1999 |
| 18. Methylene blue (MB) | Thiazine | 52015 | + | Sigma | + | Stockert & Herkovits 2003, Smijs et al. 2004, Herkovits et al. 2007 |
| 19. Morin (Mor) | Flavonol | 75660 | 0 | Merck | nd | |
| 20. Naphtol Blue Black | Bis-azo | 20470 | 2− | Serva | nd | |
| 21. Neutral red (NR) | Azine | 50040 | + | Panreac | + | Seliger & McElroy 1965, Barbosa & Peters 1971, Gutter et al. 1977 |
| 22. NiPcS4 c | Phthalocyanine | − | 4− | Aldrich | − d | |
| 23. Nuclear fast red (NFR) | Anthraquinone | 60760 | − | Merck | + | Kuramoto & Kitao 1981, Robertson et al. 2009 |
| 24. Phloxine B (PhB) | Hydroxyxanthene | 45410 | 2− | Panreac | + | Rasooly & Weisz 2002, Herkovits et al. 2007 |
| 25. Primuline (Pri) | Benzothiazole | 49000 | − | Fluka | nd | |
| 26. Pyronine Y (PY) | Aminoxanthene | 45005 | + | G.T. Gurr | − | Herkovits et al. 2007 |
| 27. Rhodamine B (RhB) | Aminoxanthene | 45170 | + | Merck | + | Ngen et al. 2009 |
| 28. Riboflavine (Rib) | Alloxazine | − | 0 | Merck | + | Naseem et al. 1988, Redmond & Gamlin 1999 |
| 29. Rose bengal (RB) | Hydroxyxanthene | 45440 | 2− | Sigma | + | Banks et al. 1985, Marthy et al. 1990, Herkovits et al. 2007 |
| 30. Ruphen e | Ru-diimide | − | 2+ | Aldrich | − | Dobrucki 2001 |
| 31. Safranine O (SO) | Azine | 50240 | + | Fluka | + | Li et al. 2006 |
| 32. Tartrazine (Tar) | Mono-azo | 19140 | 3− | Fluka | nd | |
| 33. Thioflavin S (TS) | Benzothiazole | 49010 | − | Bayer | nd | |
| 34. Thioflavin T (TT) | Benzothiazole | 49005 | + | G.T. Gurr | + | Seliger & McElroy 1965, Villanueva et al. 1993 |
| 35. Thionine (T) | Thiazine | 52000 | + | Merck | nd | |
| 36. Toluidine Blue O (TB) | Thiazine | 52040 | + | Merck | + | Stockert et al. 1996, Herkovits et al. 2007, Blázquez-Castro et al. 2009 |
| 37. Trypan Blue (TryB) | Bis-azo | 23850 | 4- | Fluka | − | Barbosa & Peters 1971, Herkovits et al. 2007 |
(a) The reference list for reported photoactivity is not exhaustive and only reflects a few but relevant citations on the best known photoactive dyes. Reference details are shown as supplementary information (S1 Text). (b) CuPcS2:cooper phthalocyanine disulfonic acid, di-o-tolylguanidine salt.(c) Nickel phthalocyanine tetrasulfonic acid, tetrasodium salt. (d) In these compounds, the presence of paramagnetic metals (Cu, Ni) abolishes the photodynamic activity. (e) Ruthenium(II)-tris(phenanthroline) dichloride. nd: non determined
Statistical analysis
In order to quantify the toxic properties of the compounds in worms, a parameter called Vitality Rate (VR) was calculated as the ratio between dye treated worms and contol animals in the same assay. Based on previous experience on tracking C. elegans, where 10% variations in locomotor activity are common over continous 30 min periods of activity tracking (S1 Table), an activity drop below 0.8 of the control population level (significantly different from 1, p < 0.05 ONE sample t-test) was set to be considered a positive toxic effect.
Dye uptake
Worms were exposed to dyes as described previously, with the difference that the culture medium with the chemical was removed from the medium after the 1 h incubation, and worms were washed inside microplates 4 times with saline solution (modified K-medium). After this treatment, the dye accumulated inside the animal body was photoactivated for 30 min, and locomotor activity was recorded for 4 hours as described previously. In parallel, 3 replicate wells were observed using bright-field and fluorescence microscopy under blue or green excitation light with the purpose of visually assessing dye uptake. The accumulation of dyes was arbitrarily quantified by the intensity of fluorescence or staining using a 4 color scale.
Drug response pathway determination
The previously indicated GFP reporter strains were cultured in NGM plates until adulthood, and treated as described above for the behavioral assessment experiments. GFP expression was determined by visual inspection and imaged using a fluorescence stereoscope (OLYMPUS model: SZX-ILLK100) 4 h after dye photoactivation. In addition, RNA samples were prepared from a population of adult worms (SS104 strain) treated with 1 μM rose Bengal, our positive control dye, and hsp-4 plus hsp-6 genes were quantified by qRT-PCR using the protocol reported previously by Buzzi et al [42]. The ama-1 gene was employed as a constitutively expressed internal control. Primers used for real time PCR are as follows: for hsp-4, hsp-4F 5’GCAGATGATCAAGCCCAAAAAG3’and hsp4R 5’GCGATTTGAGTTTTCATCTGATAGG3’; for hsp-6, hsp-6F 5’GGACAAACCAAAGGGACATG3’ and hsp-6R 5’AACGAATGCTCCAACCTGAG3’; for ama-1, ama-1F 5’CCTACGATGTATCGAGGCAAA3’ and ama-1R 5’CCTCCCTCCGGTGTAATAATG3’. All experiments were replicated 3 times.
Results
Phototoxicity assessment in adult Caenorhabditis elegans
To perform phototoxicity experiments we developed a stepped protocol, where all animals are subjected to the compounds in the dark and then either exposed to the light (Fig 1A) or kept in darkness as control (Fig 1B). Their motility was recorded with the infrared tracking method for 1 h before exposure to light and 4 h after in order to record the behavioral response. Firstly, two well characterized dyes were tested, Luxol fast blue (not phototoxic) and rose Bengal (a photoactive molecule). Neither caused an effect without light irradiation (Fig 1B).
Fig 1. Phototoxic compound activation affects C.elegans behavior.
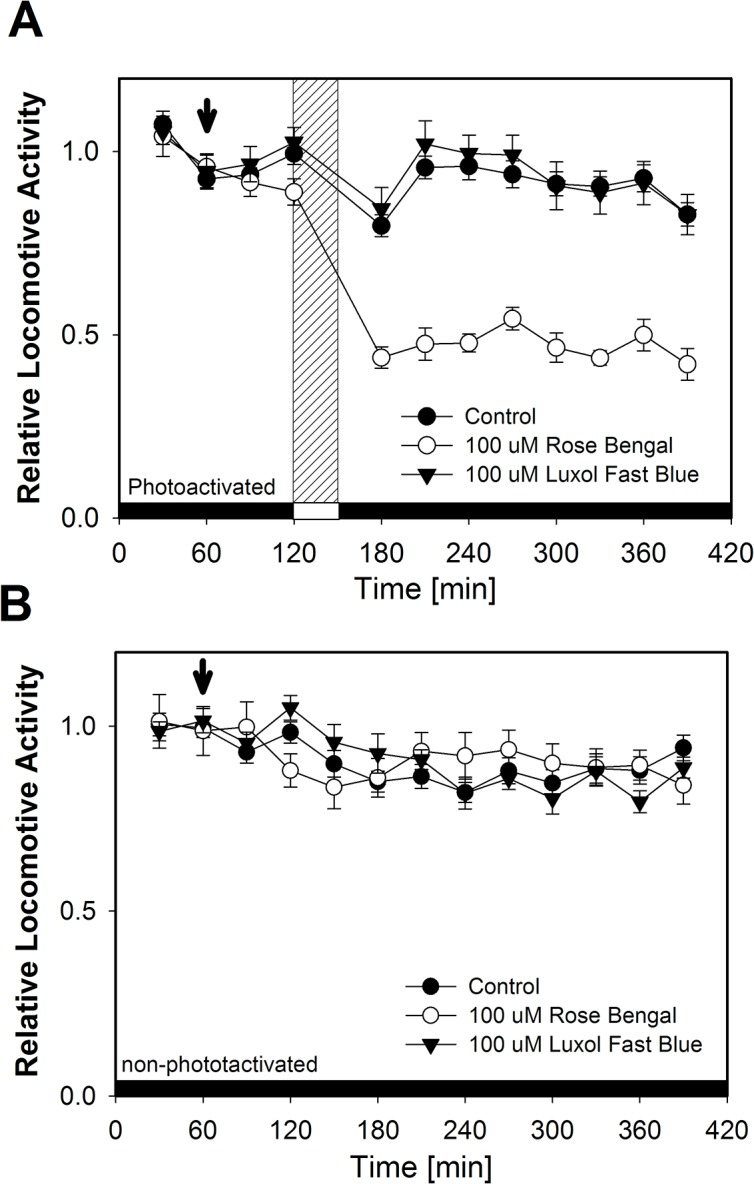
Worms where incubated in buffer (control), non-phototoxic dye (Luxol Fast Blue) and phototoxic dye (Rose Bengal). Locomotor activity was measured continously using an infrared tracking device (WMicrotracker) with exposure to photoactivating light pulse [30 min white light, 10 mW/cm2] (A) or without light exposure (B). Relative Locomotor Activity is reported as the acumulated activity of 30 min time frames over the average activity before adition of compounds.
In contrast, when white light irradiation (10 mW/cm2, 30 min) is applied (Fig 1A), a significant reduction in locomotor activity is observed in the positive control (rose Bengal), and it remains low even after 4 h. Unexpectedly, an immediate response is also observed in control animals and with a negative control (Luxol fast blue) just after light irradiation, possibly as a transient response to light. Since this short term response is magnified in animals treated with phototoxic molecules, we decided to divide biological effects in “immediate phototoxicity” and “late phototoxicity”.
We then selected a set of 37 dyes to test phototoxic effects in C. elegans cultures. Of these, 16 have been already reported to present effects in other organisms (Table 1). Of the assessed list of 37 compounds, 16 presented immediate phototoxic effects and 8 late phototoxic effects (Table 2, Fig 2A and 2B). In order of decreasing phototoxicity, the top 5 dyes were phloxine B > primuline > eosin Y > acridine orange > rose Bengal. Many of these dyes have been previously reported as phototoxic, consistent with our results (Table 1).
Table 2. Results of the screening for phototoxicity.
| Immediate Phototoxicity (30 min after light pulse) | Late Phototoxicity (240 min after light pulse) | ||||||
|---|---|---|---|---|---|---|---|
| # | Name | Vitality w/Light Pulse | Vitality non Light Pulse | Classification | Vitality w/Light Pulse | Vitality non Light Pulse | Classification |
| 24 | Phloxine B | 0.11 +/- 0.03 | 1.08 +/- 0.09 | Phototoxic | 0.26 +/- 0.06 | 0.85 +/- 0.08 | Phototoxic |
| 25 | Primuline | 0.11 +/- 0.05 | 0.86 +/- 0.07 | Phototoxic | 0.19 +/- 0.04 | 0.87 +/- 0.07 | Phototoxic |
| 10 | Eosin Y | 0.25 +/- 0.04 | 1.05 +/- 0.1 | Phototoxic | 0.63 +/- 0.07 | 1.1 +/- 0.13 | Phototoxic |
| 1 | Acridine orange | 0.42 +/- 0.05 | 1.17 +/- 0.08 | Phototoxic | 0.48 +/- 0.06 | 0.86 +/- 0.08 | Phototoxic |
| 29 | Rose bengal | 0.46 +/- 0.06 | 1.01 +/- 0.07 | Phototoxic | 0.4 +/- 0.04 | 0.91 +/- 0.05 | Phototoxic |
| 34 | Thioflavin T | 0.51 +/- 0.02 | 0.92 +/- 0.03 | Phototoxic | 0.63 +/- 0.03 | 0.81 +/- 0.05 | Phototoxic |
| 31 | Safranine T | 0.76 +/- 0.04 | 0.94 +/- 0.03 | Phototoxic | 0.64 +/- 0.03 | 0.9 +/- 0.04 | Phototoxic |
| 23 | Nuclear fast red | 0.34 +/- 0.03 | 1.07 +/- 0.04 | Phototoxic | 0.78 +/- 0.03 | 0.92 +/- 0.06 | Phototoxic |
| 7 | Chrysoidine | 0.42 +/- 0.05 | 0.63 +/- 0.06 | Phot + Tox | 0.55 +/- 0.03 | 0.5 +/- 0.05 | Toxic |
| 33 | Thioflavin S | 0.49 +/- 0.04 | 0.91 +/- 0.04 | Phototoxic | 0.81 +/- 0.07 | 0.88 +/- 0.05 | NonToxic |
| 17 | Mercurochrome | 0.56 +/- 0.04 | 1.16 +/- 0.06 | Phototoxic | 0.86 +/- 0.06 | 0.82 +/- 0.05 | NonToxic |
| 21 | Neutral red | 0.6 +/- 0.05 | 1 +/- 0.05 | Phototoxic | 0.85 +/- 0.04 | 0.93 +/- 0.04 | NonToxic |
| 37 | Trypan blue | 0.64 +/- 0.05 | 1.06 +/- 0.05 | Phototoxic | 0.91 +/- 0.06 | 0.94 +/- 0.09 | NonToxic |
| 36 | Toluidine blue O | 0.69 +/- 0.04 | 1.09 +/- 0.05 | Phototoxic | 0.86 +/- 0.04 | 0.95 +/- 0.05 | NonToxic |
| 3 | Auramine O | 0.7 +/- 0.05 | 0.9 +/- 0.07 | Phototoxic | 0.89 +/- 0.06 | 0.96 +/- 0.06 | NonToxic |
| 28 | Riboflavine | 0.76 +/- 0.06 | 0.97 +/- 0.05 | Phototoxic | 0.93 +/- 0.07 | 0.84 +/- 0.06 | NonToxic |
| 27 | Rhodamine B | 0.8 +/- 0.08 | 1 +/- 0.05 | Phototoxic | 0.98 +/- 0.05 | 0.97 +/- 0.04 | NonToxic |
| 19 | Morin | 0.73 +/- 0.05 | 0.84 +/- 0.08 | Toxic | 0.85 +/- 0.06 | 0.9 +/- 0.09 | NonToxic |
| 9 | Curcumin | 0.82 +/- 0.09 | 1.12 +/- 0.09 | NonToxic | 0.9 +/- 0.07 | 1.18 +/- 0.12 | NonToxic |
| 2 | Alizarin red S | 0.82 +/- 0.07 | 0.93 +/- 0.08 | NonToxic | 0.85 +/- 0.08 | 0.93 +/- 0.07 | NonToxic |
| 4 | Berberine | 0.82 +/- 0.09 | 0.9 +/- 0.05 | NonToxic | 0.94 +/- 0.09 | 0.92 +/- 0.09 | NonToxic |
| 5 | Bismarck brown Y | 0.87 +/- 0.08 | 0.89 +/- 0.06 | NonToxic | 0.81 +/- 0.07 | 0.85 +/- 0.06 | NonToxic |
| 6 | Carmine | 0.87 +/- 0.07 | 1.1 +/- 0.06 | NonToxic | 0.99 +/- 0.09 | 0.95 +/- 0.09 | NonToxic |
| 13 | Hematoxylin | 0.89 +/- 0.05 | 0.97 +/- 0.05 | NonToxic | 1.08 +/- 0.05 | 1.06 +/- 0.06 | NonToxic |
| 32 | Tartrazine | 0.91 +/- 0.04 | 0.93 +/- 0.06 | NonToxic | 0.99 +/- 0.05 | 0.94 +/- 0.05 | NonToxic |
| 22 | NiPcS4 | 0.92 +/- 0.07 | 0.94 +/- 0.05 | NonToxic | 0.89 +/- 0.04 | 0.89 +/- 0.06 | NonToxic |
| 26 | Pyronine Y | 0.92 +/- 0.05 | 0.93 +/- 0.03 | NonToxic | 0.94 +/- 0.04 | 0.88 +/- 0.04 | NonToxic |
| 12 | Fluorescein | 0.93 +/- 0.06 | 0.99 +/- 0.09 | NonToxic | 0.96 +/- 0.07 | 0.96 +/- 0.07 | NonToxic |
| 35 | Thionine | 1.01 +/- 0.11 | 1.01 +/- 0.08 | NonToxic | 0.85 +/- 0.07 | 0.9 +/- 0.07 | NonToxic |
| 11 | Fast green FCF | 1.01 +/- 0.07 | 0.99 +/- 0.07 | NonToxic | 0.95 +/- 0.06 | 1.02 +/- 0.09 | NonToxic |
| 20 | Naphthol Blue Black | 1.03 +/- 0.09 | 0.9 +/- 0.06 | NonToxic | 0.89 +/- 0.09 | 0.93 +/- 0.08 | NonToxic |
| 16 | Luxol fast blue | 1.03 +/- 0.07 | 1.03 +/- 0.06 | NonToxic | 0.99 +/- 0.06 | 0.98 +/- 0.05 | NonToxic |
| 18 | Methylene blue | 1.04 +/- 0.06 | 0.94 +/- 0.04 | NonToxic | 0.83 +/- 0.04 | 0.93 +/- 0.05 | NonToxic |
| 30 | Ruphen | 1.04 +/- 0.08 | 1.02 +/- 0.07 | NonToxic | 1 +/- 0.06 | 0.99 +/- 0.08 | NonToxic |
| 14 | Indigocarmine | 1.04 +/- 0.05 | 1.04 +/- 0.06 | NonToxic | 0.97 +/- 0.04 | 1.03 +/- 0.1 | NonToxic |
| 8 | Congo red | 1.12 +/- 0.09 | 1.15 +/- 0.1 | NonToxic | 1.04 +/- 0.09 | 1.06 +/- 0.11 | NonToxic |
| 15 | Janus green B | 1.12 +/- 0.07 | 0.99 +/- 0.05 | NonToxic | 0.73 +/- 0.08 | 0.71 +/- 0.07 | Toxic |
All dyes were tested at least in 2 independent experiments (with 4 internal replicates each) using 100 μM concentration, with the exception of curcumin which was assayed at 10 μM for solubility reasons. Vitality is reported as the rate of locomotor activity of dye treated worms over buffer treated worms under the same light conditions (Vitality Rate).
Fig 2. Classification of dye phototoxicity according to behavioral changes.

Vitality Rate (VR) index were calculated as [locomotor activity of dye treated worms] / [locomotor activity of control animals]. When observed in a two axis plot (VRlight vs. VRno-light) toxic effects could be classified as: Toxic (VRno-light < 0.8), Phototoxic (VRlight < 0.8) and Toxic and Phototoxic (VRlight < VRno-light < 0.8, *p < 0.05). A) Behavioral changes observed 30min after light pulse (180min from start) were classed as “immediate effects”, where 43% of compounds showed phototoxic effects. B) Changes observed 240 min after light pulse (390 min from start) were classed as “late phototoxicity effects”, where 21% of compounds showed phototoxic effects. N = 3 ± SEM.
In order to discard any synthetic effects between SS104 and a particular chemical, we test top 12 dyes in wild type (N2) young adult animals. As shown in S2 Table, although some sensitivity differences are observed, 100% of compounds manifest similar phototoxic behavior response.
Phototoxic molecules are acting directly from within the animal body
Since we measured phototoxicity without washing the compound from the medium, we asked whether the toxic effects were mainly caused by dye molecules in the culture medium or by those absorbed inside the animal. In order to clarify this point we replicated the experimental setup for 15 positive phototoxic molecules, washing the medium previous to light irradiation. Interestingly, phototoxic activity of all retested compounds remained after washout (Fig 3). Moreover, an increase in the phototoxic effect is observed in most compounds after washout. We hypothesize this increment could be attributed to higher light exposure of the worm, since the dye in the medium can be absorbing some light.
Fig 3. Phototoxic effects remain after drug washing out.

The VR of light exposed worms is shown at 180 min (30 min after light pulse) for 15 phototoxic dyes, washing them out of the medium immediately before the light pulse (gray bars) or following the original protocol (black bars). N = 3 ± SEM.
Also, in order to confirm that the compounds did in fact enter the animals, we carried out observations under the stereoscope to assess the staining or fluorescence due to dye uptake. Almost all phototoxic compounds displayed positive staining or fluorescence in living animals, demonstrating the permeability of this animal model to the dyes (Fig 4).
Fig 4. Dye uptake is observed in treated worms.

Worm staining or fluorescence (under green or blue light excitation) was visually quantified using a 3 value scale. A) Representative images of worms treated with Rose Bengal (I) under bright-field and green-light excitation, and (II) treatment with thionine observed under bright-field and blue-light excitation. B) Table of dye uptake quantification is shown for 15 phototoxic dyes.
Stress response genes are turned on in response to some phototoxic molecules in worms
Since it has been reported that phototoxic compounds produce oxidative stress and mitochondrial damage, we studied whether or not the compound were mainly active in any particular cellular compartment. For this purpose two mitochondrial-specific stress response proteins (hsp-6::GFP and hsp-60::GFP) and one endoplasmic reticulum stress response protein (hsp-4::GFP) were used. Of the 16 phototoxic compounds tested (acridine orange was rejected due to its intrinsic green fluorescence), 10 were shown to stress one or both compartments while the other 6 compounds did not seem to be active in perturbing either the mitochondria or the endoplasmic reticulum (Fig 5B).
Fig 5. Stress response machinery is activated in C. elegans under phototoxic treatment.

Two mitochondrial-specific Stress Response (SR) lines (hsp-6::GFP and hsp-60::GFP) and one endoplasmic reticulum SR line (hsp-4::GFP) were subjected to the phototoxicity protocol, and GFP-gene expression was observed 4 hours after dye photoactivation. A) Representative scale determination of each reporter line. B) Observed gene expression for 15 phototoxic dyes tested at 10 μM, 25 μM and 50 μM. N = 3. C) Photo-activated Rose Bengal stress response measured by qPCR. Hsp-4 was assayed as a marker of endoplasmic reticulum stress and hsp-6 was assayed as a marker of mitochondrial stress. RNA polymerase subunit gene (ama-1) was employed as internal reference gene. N = 3 ± SEM.
Finally, to give more support to the idea that our finding on the GFP reporter lines reflected what was happening in the phototoxicity screening, a qRT-PCR was performed on 1 μM rose Bengal exposed glp-4 worms (Fig 5C). At this low concentration, hsp-4 expression was increased 3.8 fold with respect to control (n = 3, p<0.05) while hsp-6 expression remained unaffected. No transcription was found under dark dye treatment (no photoactivation). These results confirm the response pattern observed in the reporter lines.
Correlation between our screening results and previously reported phototoxicity of compounds
A set intersection between our worm experiments and published data is shown in Fig 6. When percentage of false detections is analyzed we found a rate of 8.1% of false negatives (dyes reported as phototoxic, but not detected in our experiments) and 2.7% of false positives (dyes reported as non phototoxic but detected in our system) compared to literature reports. These differences could be attributed to variability in the uptake, efficiency for ROS production, experimental protocol, as well as specific sensitivity and threshold to oxidative stress for distinct type of cells and organisms.
Fig 6. Comparison of worm experiments with previously published data.

Results of the current study were compared to reported data for all 37 assayed dyes. Two comparisons are shown: Dyes reported as phototoxic vs. dyes determined as phototoxic in this C. elegans screening (A) and dyes reported as non-phototoxic vs. dyes determined as non-phototoxic in this C. elegans screening (B).
Discussion
Simple animal models are becoming useful to perform in vivo drug discovery experiments and testing for biological activity. In this work we propose the application of the nematode C. elegans as a reliable model for assessing phototoxicity of dyes as prototypical compounds and the utility of a simple behavioural measurement (such as global locomotor activity) as a direct readout for toxic and sub-toxic effects.
Common photoactivity experiments using cell cultures are based on the incubation with compounds, followed by washing, light exposure and measurement of viability by staining or colorimetric methods. In this work we start the experiments without dye washing. Since similar results were observed after washing of dyes, we conclude that this faster approach can be used in a rapid screening of molecules, previous or complementary to cell culture measurements. At the biological level, photoactive molecules appear to be permeating the animal cuticle or digestive tract, acting from inside the animal body (as shown in uptake experiments, Figs 4 and 5) and resulting in a reduced motility response.
It is worth to note that we were able to measure activation of the stress response machinery in at least 50% of positive compounds (Fig 5). Damage in intracellular compartments associated with protein misfolding and mitochondrial electron transport disruption are well-known mechanisms of phototoxicity action in cell cultures [3]. Our results suggest a similar mechanism of damage in worms. It might be useful to evaluate a larger set of stress-reporter strains and, even more, the amount of ROS driven by dye photoexcitation in C. elegans in order to gain a deeper understanding of the molecular mechanisms of action. These issues are the objectives of ongoing research.
In order to compare the photodynamic action of each molecule, we can define a Phototoxic Index (PI) by analyzing the phototoxicity and intrinsic toxicity of the compound. The occurrence and degree of photosensitization could be expressed as the ratio: PI = effects with light / effect without light. Thus, the five dyes showing high immediate phototoxicity would present the following values: PI(phloxine B) = 9.81; PI(primuline) = 5.45; PI(eosin Y) = 4.2, PI(acridine orange) = 2.78, and PI(rose Bengal) = 2.19. It should be noted that the higher the PI is, the stronger the phototoxic effect.
Finally, it is important to mention that this animal model and simple protocols for biotoxicity-detection are useful not only for pharmacological testing but also in ecotoxicology assays. Regarding environmental pollution, around 10,000 types of dyes and pigments are produced annually worldwide and extensively used in textile, leather, plastic and printing industries, laboratory work, and as food, pharmaceutical and cosmetic additives. About 10–15% of the total dyes used in dyeing processes are released in wastewater [17]. Therefore, contamination by dyes [19] and the resulting phototoxicity represent a significant risk for human health and wildlife preservation. In addition, other pharmacological agents showing undesired photoactivity must be taken into account (e.g., antiinflammatory, anxiolytic, antirheumatic, antibacterial, and antiparasitic drugs), which have also revealed to be phototoxic [21, 24, 26, 29, 36, 43, 44]. As a logical consequence, it is increasingly necessary to evaluate the phototoxicity of possible drugs or xenobiotics to induce or prevent, respectively, biological effects through the design and development of simple, precise and cheap bioassays for oxidative stress-dependent phototoxicity studies. The present results using an automated tracking device for assessing population motility of a whole organism such as C. elegans show that this bioassay is very suitable for easy, rapid and accurate evaluation of the phototoxic potential of photosensitizing drugs and environmental pollutants.
Supporting Information
In order to estimate the average activity and deviation over a 4 h recording period, 8 biological replicates, independently repeated, are shown in the table. Each test was normalized to the first half hour of recording. Four wells replicates, and 50 worms per well, were used for each experiment.
(DOC)
Immediate and late phototoxic response is shown in the table, and compared with SS104 strain results.
(DOC)
(DOCX)
Acknowledgments
This work was supported by grants PICTStartup and PAEBT2009-19 from Ministry of Science and Technology Argentina, and CTQ2013-48767-C3-3R from the Ministerio de Economía y Competitividad, Spain. The strains used in this work were provided by the CGC Center, which is supported by the NIH, and the C. elegans knockout consortium.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by grants PICTStartup and PAEBT2009-19 from Ministry of Science and Technology Argentina, and CTQ2013-48767-C3-3R from the Ministerio de Economía y Competitividad, Spain. The strains used in this work were provided by the CGC Center, which is supported by the NIH, and the C. elegans knockout consortium. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Seliger HH, McElroy WD (1965) Light: physical and biological action Acad Press: New York, London: pp 324–332. [Google Scholar]
- 2. Robertson CA, Hawkins D, Abrahamse H (2009) Photodynamic therapy (PDT): A short review on cellular mechanisms and cancer research applications for PDT. J Photochem Photobiol B: Biol 96: 1–8. [DOI] [PubMed] [Google Scholar]
- 3. Villanueva A, Vidania R, Stockert JC, Cañete M, Juarranz A (2003) Photodynamic effects on cultured tumor cells: cytoskeleton alterations and cell death mechanisms In: Nalwa HS, ed. Handbook of Photochemistry and Photobiology v4 California: American Scientific Publishers; pp 79–117. [Google Scholar]
- 4. Stockert JC, Juarranz A, Villanueva A, Nonell S, Horobin RW, Soltermann AT, et al. (2004) Photodynamic therapy: selective uptake of photosensitizing drugs into tumor cells. Curr Top Pharmacol 8: 185–217. [Google Scholar]
- 5. Cañete M, Stockert JC, Villanueva A (2009) Preclinical photodynamic therapy research in Spain. 3. Localization of photosensitizers and mechanisms of cell death in vitro. J Porphyrins Phthalocyanines 13: 544–551. [Google Scholar]
- 6. Huang Z (2005) A review of progress in clinical Photodynamic Therapy. Technol Cancer Res Treat 4: 283–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Stockert JC, Juarranz A, Villanueva A, Cañete M (1996) Photodynamic damage to HeLa cell microtubules induced by thiazine dyes. Cancer Chemother Pharmacol 39: 167–169. [DOI] [PubMed] [Google Scholar]
- 8. Mellish KJ, Cox RD, Vernon DI, Griffiths J, Brown SB (2002) In vitro photodynamic activity of a series of methylene blue analogues. Photochem Photobiol 75: 392–397. [DOI] [PubMed] [Google Scholar]
- 9. Wagner SJ, Skripchenko A (2003) Investigation of photosensitizing dyes for pathogen reduction in red cell suspensions. Biotech Histochem 78: 171–177. [DOI] [PubMed] [Google Scholar]
- 10. Marthy HJ, Murasecco-Suardi P, Oliveros E, Braun AM (1990) Primary effects of singlet oxygen sensitizers on eggs and embryos of sea urchin. J Photochem Photobiol B: Biol 7: 303–315. [DOI] [PubMed] [Google Scholar]
- 11. Zdolsek JM, Olsson GM, Brunck UT (1990) Photooxidative damage to lysosomes of cultured macrophages by acridine orange. Photochem Photobiol 51: 67–76. [DOI] [PubMed] [Google Scholar]
- 12. Kussovski VK, Hristov AE, Radoucheva TS (2001) Proflavine-mediated inactivation of Salmonella dublin exposed to visible sunlight in natural fresh water. Microbios 105: 119–125. [PubMed] [Google Scholar]
- 13. Kandela IK, Bartlett JA, Indig GL (2002) Effect of molecular structure on the selective phototoxicity of triarylmethane dyes toward tumor cells. Photochem Photobiol Sci 1: 309–314. [DOI] [PubMed] [Google Scholar]
- 14. Wainwright M (2003) The use of dyes in modern biomedicine. Biotech Histochem 78: 147–155. [DOI] [PubMed] [Google Scholar]
- 15. Horobin RW, Kiernan JA (2002) Conn’s biological stains. A handbook of dyes, stains and fluorochromes for use in biology and medicine 10th Ed. Bios Scientific Publishers, Oxford. [Google Scholar]
- 16. Sugden JK (2004) Photochemistry of dyes and fluorochromes used in biology and medicine: some physicochemical background and current applications. Biotech Histochem 79: 71–90. [DOI] [PubMed] [Google Scholar]
- 17. Vaidya AA, Datye KV (1982) Environmental pollution during chemical processing of synthetic fibres. Colourage 14: 3–10. [Google Scholar]
- 18. Anliker R (1986) Organic colourants. Interpretation of mammalian, geno-, and eco-toxicity data in terms of potential risks In: Richardson M, ed. Toxic hazard assessment of chemicals. London: Royal Society of Chemistry; pp 166–187. [Google Scholar]
- 19. Robinson T, McMullan G, Marchant R, Nigam P (2001) Remediation of dyes in textile effluent: a critical review on current treatment technologies with a proposed alternative. Bioresour Technol 77: 247–255. [DOI] [PubMed] [Google Scholar]
- 20. Herkovits J, Pérez-Coll CS, Stockert JC, Blázquez A (2007) The screening of photodynamic toxicity of dyes by means of a bioassay using amphibian embryos. Res J Chem Environ 11: 86–91. [Google Scholar]
- 21. Gocke E (2001) Photochemical mutagenesis: examples and toxicological relevance. J Environ Pathol Toxicol Oncol 20: 285–292. [PubMed] [Google Scholar]
- 22. Smijs TGM, Nivard MJM, Schuitmaker HJ (2004) Development of a test system for mutagenicity of photosensitizers using Drosophila melanogaster . Photochem Photobiol 79: 332–338. [DOI] [PubMed] [Google Scholar]
- 23. Gupta I, Tripathy BC (2000) Oxidative stress in cucumber (Cucumis sativus L) seedlings treated with acifluorfen. Indian J Bichem Biophys 37: 498–505. [PubMed] [Google Scholar]
- 24. Inbaraj JJ, Kukielczak BM, Bilski P, Sandvik SL, Chignell CF (2001) Photochemistry and photocytotoxicity of alkaloids from Goldenseal (Hydrastis canadensis L.) 1. Berberine. Chem Res Toxicol 14: 1529–1534. [DOI] [PubMed] [Google Scholar]
- 25. Singer H, Müller S, Tixier C, Pillonel L (2002) Triclosan: occurrence and fate of a widely used biocide in the aquatic environment: field measurements in wastewater treatment plants, surface waters and lake sediments. Environ Sci Technol 36: 4998–5004. [DOI] [PubMed] [Google Scholar]
- 26. Kolpin DW, Furlong ET, Meyer MT, Thurman EM, Saugg SD, Barber LB, et al. (2002) Pharmaceuticals, hormones, and other organic wastewater contaminants in U.S. streams, 1999–2000: a national reconnaissance. Environ Sci Technol 36: 1202–1211. [DOI] [PubMed] [Google Scholar]
- 27. Scrano L, Bufo SA, D’Auria M, Meallier P, Behechti A, Shramm KW (2002) Photochemistry and photoinduced toxicity of acifluorfen, a diphenyl-ether herbicide. J Environ Qual 31: 268–274. [DOI] [PubMed] [Google Scholar]
- 28. Song S, Song QJ, Chen Z (2007) Online phototransformation—flow injection chemiluminescence determination of triclosan. Anal Bioanal Chem 387: 2917–2922. [DOI] [PubMed] [Google Scholar]
- 29. Robertson PKJ, Black KD, Adams M, Willis K, Buchan F, Orr H, et al. (2009) A new generation of biocides for control of crustacean in fish farms. J Photochem Photobiol B: Biol 95: 58–63. doi: 10.1016/j.jphotobiol.2008.12.009 [DOI] [PubMed] [Google Scholar]
- 30. Joshi PC, Misra RB (1986) Evaluation of chemically-induced phototoxicity to aquatic organism using Paramecium as a model. Biochem Biophys Res Commun 139: 79–84. [DOI] [PubMed] [Google Scholar]
- 31. Knudsen EA (1985) The Candida phototoxicity test. The sensitivity of different strains and species of Candida, standardization attempts and analysis of the dose-response curves for 5- and 8-methoxypsoralen. Photodermatology 2: 80–85. [PubMed] [Google Scholar]
- 32. Molero ML, Hazen MJ, Stockert JC (1985) Photodynamic effect of berberine sulfate on the growth rate of Allium cepa roots. J Plant Physiol 120: 91–94. [Google Scholar]
- 33. Stockert JC, Herkovits J (2003) Photodynamic toxicity and its prevention by antioxidative agents in Bufo arenarum embryos. Toxicology 192: 211–218. [DOI] [PubMed] [Google Scholar]
- 34. Merlin J, Azzi S, Lignon D, Ramacci C, Zeghari N, Guillemin F (1992) MTT assays allow quick and reliable measurements of the response of human tumour cells to photodynamic therapy. Eur J Cancer 28: 1452–1458. [DOI] [PubMed] [Google Scholar]
- 35. Hetherington AM, Johnson BE (1984) Photohemolysis. Photodermatology 1: 255–260. [PubMed] [Google Scholar]
- 36. Wagai N, Tawara K (1991) Quinolone antibacterial agent-induced cutaneous phototoxicity: ear swelling reactions in Balb/c mice. Toxicol Lett 58: 215–223. [DOI] [PubMed] [Google Scholar]
- 37. Riddle DL, Blumenthal T, Meyer BJ, Preiss JR (1997) C. elegans II Cold Spring Harbor Laboratory Press, New York. [PubMed] [Google Scholar]
- 38. Artal-Sanz M, de Jong L, Tavernarakis N (2006) Caenorhabditis elegans: A versatile platform for drug discovery. Biotechnol J 1: 1405–1418. [DOI] [PubMed] [Google Scholar]
- 39. Simonetta SH, Golombek DA (2007) An automated tracking system for Caenorhabditis elegans locomotor behaviour and circadian studies application. J Neurosci Meth 161: 273–280. [DOI] [PubMed] [Google Scholar]
- 40. Brenner S (1974) The genetics of Caenorhabditis elegans . Genetics 77: 71–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Williams PL, Dusenbery DB (1990) Aquatic toxicology testing using the nematode Caenorhabditis elegans . Environ Toxicol Chem 9:1285–1290. [Google Scholar]
- 42. Buzzi LI, Simonetta SH, Parodi AJ, Castro OA (2011) The two Caenorhabditis elegans UDP-glucose:glycoprotein glucosyltransferase homologues have distinct biological functions. PLoS ONE 6(11): e27025 doi: 10.1371/journal.pone.0027025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Griebrock JJ (1979) Photomutagenesis by chlorinated phenothiazine tranquilizers. Proc Natl Acad Sci USA 76: 469–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Lewis DF, Ioannides C, Parke DV (1990) A retrospective study of the molecular toxicology of benoxaprofen. Toxicology 65: 33–47. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
In order to estimate the average activity and deviation over a 4 h recording period, 8 biological replicates, independently repeated, are shown in the table. Each test was normalized to the first half hour of recording. Four wells replicates, and 50 worms per well, were used for each experiment.
(DOC)
Immediate and late phototoxic response is shown in the table, and compared with SS104 strain results.
(DOC)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.