

Abstract
The Southern Ocean is a key region for global carbon uptake and is characterized by a strong seasonality with the annual CO2 uptake being mediated by biological carbon drawdown in summer. Here we show that the contribution of biology to CO2 uptake will become even more important until 2100. This is the case even if biological production remains unaltered and can be explained by the decreasing buffer capacity of the ocean as its carbon content increases. The same amount of biological carbon drawdown leads to a more than twice as large reduction in CO2(aq) concentration and hence to a larger CO2 gradient between ocean and atmosphere that drives the gas exchange. While the winter uptake south of 44°S changes little, the summer uptake increases largely and is responsible for the annual mean response. The combination of decreasing buffer capacity and strong seasonality of biological carbon drawdown introduces a strong and increasing seasonality in the anthropogenic carbon uptake.
Key Points
Decrease of buffer capacity leads to stronger summer CO2 uptake in the future
Biology will contribute more to future CO2 uptake in Southern Ocean
Seasonality affects anthropogenic carbon uptake strongly
Keywords: ocean CO2 sink, buffer capacity, anthropogenic carbon, feedback process, biological carbon draw down, ocean acidification
1. Introduction
The global ocean takes up about a quarter of current CO2 emissions [Le Quéré et al., 2014]. The ocean can take up more CO2 from the atmosphere than expected for an inert gas (such as oxygen and nitrogen) because CO2 reacts with seawater. An equilibrium comes into place:
| (1) |
and only 1% of the CO2 that was taken up as a gas remains in the form of CO2(aq). This property of the carbonate system is known as buffering and can be quantified using the Revelle factor, R [Revelle and Suess, 1957]:
| (2) |
The Revelle factor R is the ratio of the relative change of seawater pCO2 (or, alternatively, CO2(aq)) to the relative change of dissolved inorganic carbon (DIC, the sum of CO2(aq), HCO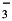 and CO
and CO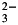 ) [Zeebe and Wolf-Gladrow, 2001; Egleston et al., 2010]. It varies between 8 in warm and 15 in cold waters [Broecker et al., 1979]. The lower the Revelle factor, the more efficient is the anthropogenic CO2 uptake [Völker et al., 2002; Sabine et al., 2004]. As the ocean continues to take up CO2, the carbonate system is pushed toward higher CO2(aq) concentrations and the Revelle factor increases [Zeebe and Wolf-Gladrow, 2001]. Consequently, the buffer capacity of the ocean and its CO2 uptake capacity decrease. This is a positive feedback limiting the increase of the ocean CO2 sink in the future [Wallace, 2001].
) [Zeebe and Wolf-Gladrow, 2001; Egleston et al., 2010]. It varies between 8 in warm and 15 in cold waters [Broecker et al., 1979]. The lower the Revelle factor, the more efficient is the anthropogenic CO2 uptake [Völker et al., 2002; Sabine et al., 2004]. As the ocean continues to take up CO2, the carbonate system is pushed toward higher CO2(aq) concentrations and the Revelle factor increases [Zeebe and Wolf-Gladrow, 2001]. Consequently, the buffer capacity of the ocean and its CO2 uptake capacity decrease. This is a positive feedback limiting the increase of the ocean CO2 sink in the future [Wallace, 2001].
The Southern Ocean turned from a source of CO2 in preindustrial times into a CO2 sink, today being the key region for CO2 uptake together with the North Atlantic. It is the strong uptake of anthropogenic carbon [Khatiwala et al., 2009] that nearly balances the outgassing of natural carbon and results in a net CO2 sink [Gruber et al., 2009]. The contemporary CO2 flux is characterized by a strong seasonality driven by deep winter mixing and photosynthesis in summer [Takahashi et al., 2009]. The Southern Ocean is a CO2 source in winter and a CO2 sink in summer [Lenton et al., 2013] highlighting the importance of biological carbon drawdown in summer [Bakker et al., 1997]. The effect of seasonal temperature variability on the solubility of CO2 would work in the opposite direction and is apparently of secondary role in the Southern Ocean [Takahashi et al., 2002].
In this study we will show that the seasonal drawdown of carbon by biological production will contribute an even larger share to total CO2 uptake as the Revelle factor increases toward the end of the century. Our analysis focuses on the Southern Ocean, a key region for global carbon uptake, but the process might be at work in all regions governed by strong seasonality.
2. Model
We use the Massachusetts Institute of Technology general circulation model (MITgcm) [Marshall et al., 1997; Massachusetts Institute of Technology General Circulation Model Group, 2014] with a state-of-the-art sea ice model [Losch et al., 2010] coupled to the Regulated Ecosystem Model with two phytoplankton classes (REcoM-2). The horizontal resolution is 2° in longitude and 0.38 to 2° in latitude, with the higher resolution in the Southern Ocean and at the equator. The thickness of the vertical layers is 10 m at the surface, increasing downward to 500 m at a depth of 3700 m. Eddies are parameterized, using the Gent and McWilliams [1990] formulation. REcoM-2 carries 21 prognostic tracers, including dynamic intracellular pools of carbon, chlorophyll, nitrogen, and silicate (for diatoms) for the two phytoplankton classes (nanophytoplankton and diatoms) [Geider et al., 1998] as well as actively advected detritus carbon, nitrogen, and silicate pools. Iron is taken up by the cells in a fixed ratio to carbon. CO2 gas exchange across the air-sea interface considers chemical enhancement [Wanninkhof, 1992]. More details on the ocean and ecosystem model setup and present-day validation is given in Hauck et al. [2013a].
The future simulation of the period 2011 to 2100 is started from a hindcast of the period 1948-2010 [Hauck et al., 2013a] that was preceded by a 47 year spin-up. The 6-hourly atmospheric forcing fields are obtained from the Model for Interdisciplinary Research on Climate (MIROC5) Representative Concentration Pathway 8.5 scenario (RCP8.5) simulation that is available from the Coupled Model Intercomparison Project phase 5 (CMIP5) archive [Watanabe et al., 2011].
The atmospheric CO2 concentration follows the RCP8.5 scenario [Meinshausen et al., 2011] in the standard model run (RCP85). In an additional simulation (CONST), atmospheric CO2 is kept constant at the preindustrial concentration of 278 ppm. Both simulations experience climate change as projected by the MIROC5 atmospheric fields.
Results will be presented in three latitudinal bands following RECCAP protocols [Lenton et al., 2013]. The region south of 58°S refers to the Antarctic Zone south of the Polar Front; a latitudinal band between 44°S and 58°S spans large parts of the subantarctic and polar frontal zones and includes the polar front itself. The zonal band between 30°S and 44°S consists of the subtropical zone, with a minor contribution from the subantarctic zone.
3. Results
Figure 1 depicts the time series of CO2 fluxes in the RCP85 and CONST simulations as well as their mean seasonal cycle for the beginning and end of the simulations. There is a general increase of CO2 uptake in RCP85 as expected from increasing atmospheric CO2 concentrations.
Figure 1.
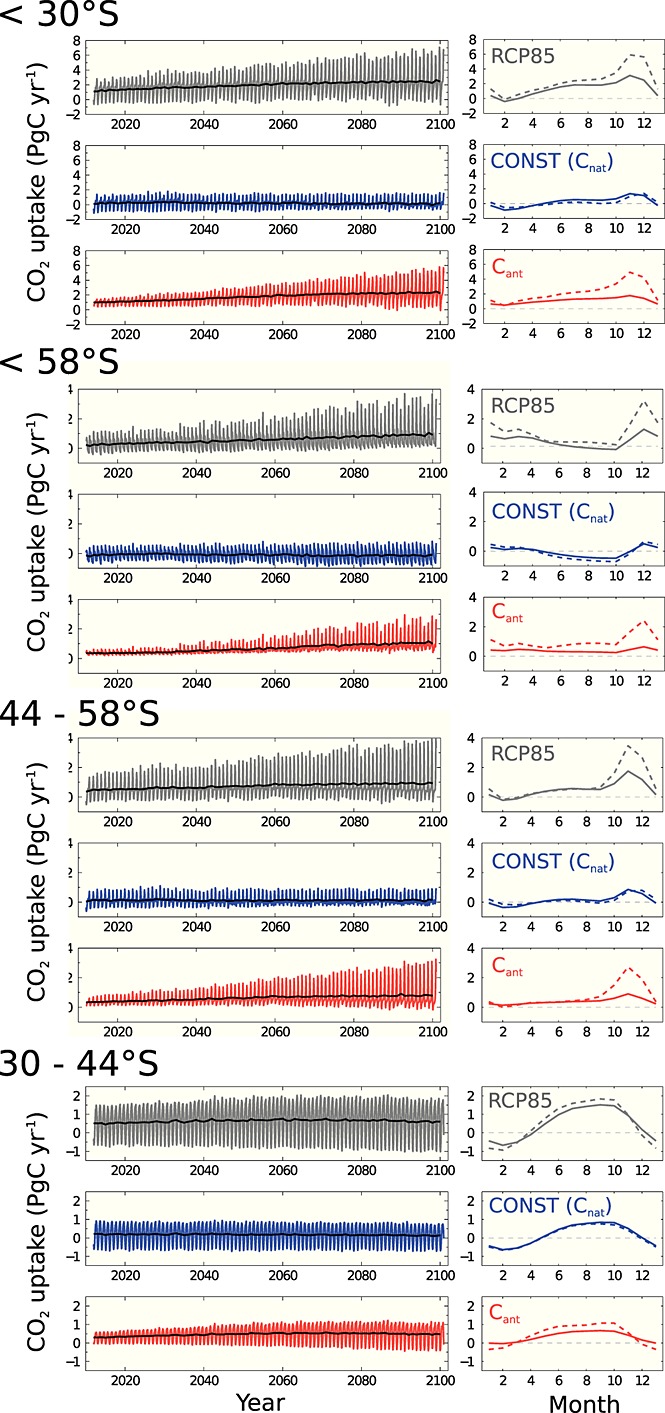
(first column) Monthly time series of CO2 uptake in the model simulations following the RCP8.5 scenario (RCP85, dark grey), in the scenario with constant atmospheric CO2 (CONST, equivalent to natural CO2 flux, blue) and the difference between the two simulations (anthropogenic CO2 flux, red) in MITgcm-REcoM-2 averaged over the full Southern Ocean (<30°S), and the subregions <58°S, 44°S–58°S, and 30°S–44°S as indicated in the figures. The annual mean is overlain in black. (second column) The mean seasonal cycle of CO2 uptake for the periods 2012–2031 (line) and 2081–2100 (dashed) in the same regions and model runs. January is shown as the first and again as the last month of the seasonal cycle.
More surprisingly, the amplitude of the seasonal cycle increases in all subregions of the Southern Ocean in RCP85. The CONST simulation does not exhibit this behavior. In the regions south of 44°S, the strong summer CO2 uptake drives the annual mean increase with winter CO2 uptake changing only little (Figure 1, second column). The absence of this effect in the CONST simulation demonstrates that the strengthening of the seasonal cycle is not a response of biology to changing climate and altered resource availability. This implies that (1) the change in the integrated Southern Ocean CO2 uptake is to a large part driven by changes in anthropogenic CO2 flux, i.e., by the increase of atmospheric CO2 itself and only to a smaller degree by circulation changes that are expected to occur in the Southern Ocean [e.g., Le Quéré et al., 2007; Hauck et al., 2013a] and that (2) future CO2 uptake has a strong seasonality that becomes more important toward the end of the century.
The peak of CO2 uptake occurs in November to January (south of 58°S) and October to December (44°S–58°S, Figure 1, second column). This coincides with the peak of carbon drawdown by biological production in these regions (Figure 2).
Figure 2.
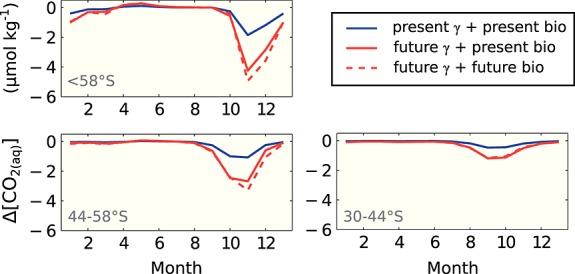
Effect of buffer factor on CO2(aq) drawdown by biology calculated as in equation (4). Blue: CO2 drawdown with present-day buffer factor and biological carbon drawdown (gross primary production minus respiration minus remineralization). Red: future buffer factor and present-day biological carbon drawdown. Red, dashed: future buffer factor and future biological carbon drawdown. Present day: mean seasonal cycle 2012 to 2031, future: 2081 to 2100. Change in buffer factor has a much larger effect on CO2(aq) drawdown (and hence CO2 uptake) than increase in biological production. January is shown as the first and again as the last month of the seasonal cycle.
We explain this emergent strengthening of seasonality in CO2 uptake by the increase of the Revelle factor over the course of the century (Figure 3) that brings the system into a new state, which is less well buffered and therefore more sensitive to seasonal DIC variations [Riebesell et al., 2009; Egleston et al., 2010]. As a consequence, the summer DIC drawdown by biology leads to a stronger decrease in CO2(aq). As it is the CO2(aq) concentration (and not DIC concentration) that controls the gas exchange with the atmosphere, this allows more CO2 to be taken up by the ocean. In other words, the same amount of biological production leads to a stronger CO2 uptake at the end of the century than at the beginning of the simulation. An increase of export production south of 44°S apparent in our model simulation reinforces this effect, yet the buffer factor is driving it.
Figure 3.
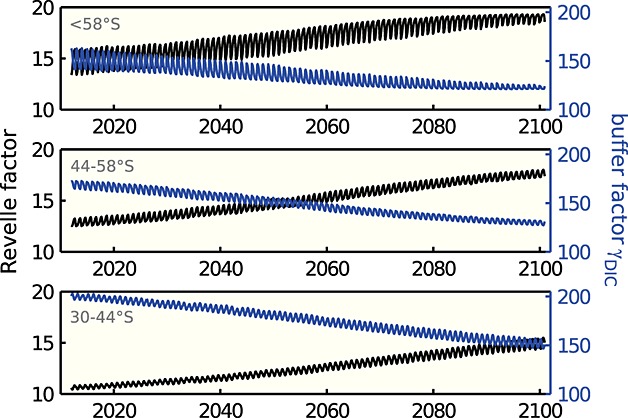
Monthly time series of Revelle factor (black, unitless) and buffer factor γDIC (blue, μmol kg−1) [Egleston et al., 2010] for the RCP8.5 scenario in the period 2012 to 2100 and the regions <58°S, 44°S–58°S, and 30°S–44°S as indicated in the figures.
This is illustrated in Figure 2 where we use the mean surface carbonate system from REcoM-2 for the periods 2012–2031 and 2081–2100 to calculate the sensitivity of aqueous CO2 concentration [CO2(aq)] to DIC perturbations using the buffer factor γDIC which is linked to the Revelle factor via
| (3) |
an can be explicitly derived from the carbonate system parameters [Egleston et al., 2010]. The decrease of the buffer capacity that is described by an increase in the Revelle factor manifests itself as a decrease in the buffer factor γDIC (Figure 3). The reduction in [CO2(aq)] due to biological carbon drawdown is then calculated as
| (4) |
where ΔDIC is the carbon drawdown by biological production in the top 100 m layer of the ocean taken from REcoM-2 (as gross primary production minus remineralization minus respiration). The total CO2(aq) drawdown by biology integrated over the year increases by a factor of 2.3, 2.5, and 2.6 due to changes in the buffer factor for the regions <58°S, 44–58°S, and 30–44°S, respectively (calculated as Δ[CO2(aq)]2081 − 2100/Δ[CO2(aq)]2012 − 2031). The contribution of altered biological production is small as indicated by similar numbers for the combined effects of production and buffer capacity changes (2.7, 2.9, and 2.5 from south to north, Figure 2).
North of 44°S, summer outgassing of CO2 becomes stronger and the amplitude of the seasonal cycle widens in both directions. The Revelle factor is still lower than in the other two regions, so that the buffer effect on CO2(aq) drawdown is smaller in absolute numbers in this region (Figure 2). As the buffer capacity of the system decreases, the system reacts stronger to DIC perturbations, but biology is only one factor among others and seasonality is not as pronounced as in the other regions. The source minus sink term for DIC becomes positive after the spring bloom, leading to (stronger) outgassing of CO2 in the less well-buffered system in austral summer (January and February).
4. Discussion
The increase of the Revelle factor with rising pCO2 acts as a strong positive feedback, reducing potential CO2 uptake in the ocean in the future [Wallace, 2001]. The other side of the coin, however, is that CO2(aq) becomes more sensitive to DIC variations as pCO2 rises, such that a strong seasonal drawdown of DIC is responsible for the increasing CO2 uptake. This is particularly relevant in systems with a strong seasonality such as the high latitudes where darkness inhibits biological production in winter.
The buffer factor γDIC reaches its theoretical minimum of 120 μmol kg−1in winter in the region <58°S at the end of the simulation. The buffer factor has a minimum when alkalinity approximately equals DIC or in other words when the system is equally distant from both pK values [see Egleston et al., 2010]. Based on this concept, we would expect that the buffer capacity would further decrease in the regions 30°S–44°S and 44°S–58°S beyond 2100 (assuming constant alkalinity, note that alkalinity is not expected to increase strongly in the Southern Ocean as carbonate sediments are sparse [Hauck et al., 2013b]). The buffer capacity south of 58°S would rise again with further CO2 uptake after the system passed through the minimum.
In the literature the concept of so-called natural and anthropogenic CO2 is used recurrently [e.g., Sarmiento et al., 1992]. The natural CO2 (Cnat) flux describes the CO2 exchange between ocean and atmosphere in preindustrial times at an atmospheric CO2 concentration of 278 μatm. The concept assumes that this background flux is today overlain by an uptake of anthropogenic CO2(Cant) which is not affected by biology and/or seasonal variability. (however, Rodgers et al. [2008] and Gorgues et al. [2010] demonstrate that there is a seasonality of Cant uptake at present). Indeed, the seasonal cycle of Cant uptake that can be explicitly simulated in the model (RCP85 − CONST) is smaller than the seasonal cycle of Cnat uptake at the beginning of the simulation (Figure 1).
In the Southern Ocean, CO2 is taken up from the atmosphere when biology draws down carbon in spring and summer. As the Revelle factor increases due to anthropogenic CO2 emissions, seasonality becomes more important and the peak CO2 uptake is evident in the anthropogenic carbon flux (Figure 1). A strong seasonal amplitude in Cant flux is established which becomes larger than the seasonal amplitude of Cnat uptake after approximately 2050. This implies that biological processes impact anthropogenic CO2 uptake, and this effect becomes more important as time and emissions progress.
We would like to note that typical methods for calculating anthropogenic carbon (based on DIC or CFC measurements) that implicitly assume that biologically induced carbon fluxes do not change Cant uptake [e.g., Wallace, 1995; Gruber et al., 1996; Goyet et al., 1999; Touratier and Goyet, 2004; Friis et al., 2005; Waugh et al., 2006] might become biased in the future. While this was observed previously [Goodkin et al., 2011], we propose that this is due to the increased importance of the biological carbon pump for CO2 uptake. The same amount of biological production and hence nutrient uptake leads to more CO2 uptake per buildup of biomass in the future. This questions the basic assumption that anthropogenic carbon can be calculated from the DIC concentration after applying a correction for biological production based on a linear relationship between nutrients and carbon. In contrast, we show that anthropogenic carbon is directly affected by nutrient variability or seasonality when taken as a proxy for biological production (Figure 1). To demonstrate this point, we calculate the Cant inventory using the extended multiple regression method (eMLR) method [Friis et al., 2005] with the parameters potential temperature, alkalinity, and nitrate from the RCP85 simulation (Figure 4, see supporting information for details of calculation). There is an increasing gap between the Cant inventory as simulated with the model (RCP85 −CONST) and that calculated using the eMLR method. We ascribe this to the impact of biology on Cant uptake that is not accounted for by the method. We hypothesize that other methods that are based on the assumption of a linear and constant relationship between either carbon and nutrients or possibly carbon and CFCs (biology does impact uptake of CO2 but not of CFCs) may experience similar problems, but leave this to be tested in future work.
Figure 4.
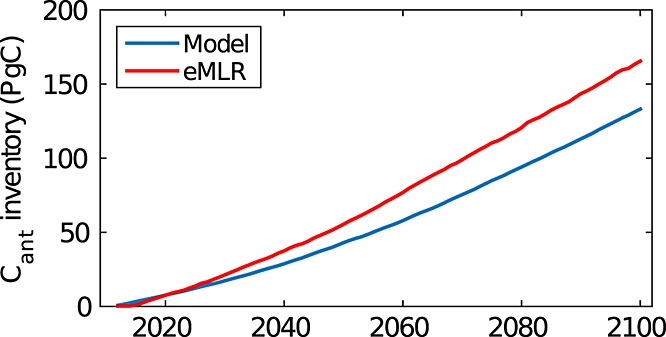
Buildup of Cant inventory (Pg C) relative to the reference year 2011 as simulated with the model (blue, RCP85 − CONST) and as calculated with the eMLR method from the RCP85 simulation using the parameters theta, alkalinity, and dissolved inorganic nitrogen (red). See supporting information for details of eMLR calculation.
Biology will contribute substantially more to CO2 uptake as the buffer capacity decreases. We demonstrate this in an example calculation using model output (Figure 2). While the timing and magnitude of the described effect is model dependent, the underlying carbonate chemistry is well understood [Zeebe and Wolf-Gladrow, 2001; Egleston et al., 2010], and this is an intrinsic feature of the carbonate system.
Acknowledgments
This research was supported through EU FP7 project CARBOCHANGE (grant agreement 264879) and through the Helmholtz PostDoc Programme. The results of model simulation RCP85 are available via the MARine Ecosystem Model Intercomparison Project (MAREMIP). In addition, the data used to produce the figures are available at http://dx.doi.org/10.1594/PANGAEA.841611. We thank two anonymous reviewers for their comments on the manuscript.
The Editor thanks two anonymous reviewers for their assistance in evaluating this paper.
Supporting Information
Text S1
References
- Bakker D, Baar HD, Bathmann U. Changes of carbon dioxide in surface waters during spring in the southern ocean. Deep Sea Res., Part II. 1997;44(1–2):91–127. doi: 10.1016/S0967-0645(96)00075-6. [Google Scholar]
- Broecker W, Takahashi T, Simpson H, Peng T-H. Fate of fossil fuel carbon dioxide and the global carbon budget. Science. 1979;206(4417):409–418. doi: 10.1126/science.206.4417.409. doi: 10.1126/science.206.4417.409. [DOI] [PubMed] [Google Scholar]
- Egleston ES, Sabine CL, Morel FMM. Revelle revisited: Buffer factors that quantify the response of ocean chemistry to changes in DIC and alkalinity. Global Biogeochem. Cycles. 2010;24:GB1002. doi: 10.1029/2008GB003407. [Google Scholar]
- Friis K, Körtzinger A, Patsch J, Wallace DWR. On the temporal increase of anthropogenic CO2 in the subpolar North Atlantic. Deep Sea Res., Part I. 2005;52(5):681–698. [Google Scholar]
- Geider RJ, MacIntyre HL, Kana TM. A dynamic regulatory model of phytoplankton acclimation to light, nutrients, and temperature. Limnol. Oceanogr. 1998;43:679–694. doi: 10.4319/lo.1998.43.4.0679. [Google Scholar]
- Gent PR, McWilliams JC. Isopycnal mixing in ocean circulation models. J. Phys. Oceanogr. 1990;20:150–155. [Google Scholar]
- Goodkin NF, Levine NM, Doney SC, Wanninkhof R. Impacts of temporal CO2 and climate trends on the detection of ocean anthropogenic CO2 accumulation. Global Biogeochem. Cycles. 2011;25:GB3023. doi: 10.1029/2010GB004009. [Google Scholar]
- Gorgues T, Aumont O, Rodgers KB. A mechanistic account of increasing seasonal variations in the rate of ocean uptake of anthropogenic carbon. Biogeosciences. 2010;7(8):2581–2589. doi: 10.5194/bg-7-2581-2010. [Google Scholar]
- Goyet C, Coatanoan C, Eischeid G, Amaoka T, Okuda K, Healy R, Tsunogai S. Spatial variation of total alkalinity in the northern Indian Ocean: A novel approach for the quantification of anthropogenic CO2 in seawater. J. Mar. Res. 1999;57:135–163. [Google Scholar]
- Gruber N, Sarmiento JL, Stocker TF. An improved method for detecting anthropogenic CO2 in the oceans. Global Biogeochem. Cycles. 1996;10(4):809–837. [Google Scholar]
- Gruber N, et al. Oceanic sources, sinks, and transport of atmospheric CO2. Global Biogeochem. Cycles. 2009;23:GB1005. doi: 10.1029/2008GB003349. [Google Scholar]
- Hauck J, Völker C, Wang T, Hoppema M, Losch M, Wolf-Gladrow DA. Seasonally different carbon flux changes in the Southern Ocean in response to the southern annular mode. Global Biogeochem. Cycles. 2013a;27:1236–1245. doi: 10.1002/2013GB004600. doi: 10.1002/2013GB004600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauck J, Arrigo KR, Hoppema M, Dijken G, Völker C, Wolf-Gladrow D. Insignificant buffering capacity of Antarctic shelf carbonates. Global Biogeochem. Cycles. 2013b;27:11–20. doi: 10.1029/2011GB004211. [Google Scholar]
- Khatiwala S, Primeau F, Hall T. Reconstruction of the history of anthropogenic CO2 concentrations in the ocean. Nature. 2009;462:346–349. doi: 10.1038/nature08526. doi: 10.1038/nature08526. [DOI] [PubMed] [Google Scholar]
- Le Quéré C, et al. Saturation of the Southern Ocean CO2 sink due to recent climate change. Science. 2007;316:1735–1738. doi: 10.1126/science.1136188. doi: 10.1126/science.1136188. [DOI] [PubMed] [Google Scholar]
- Le Quéré C, et al. Global carbon budget 2014. Earth Syst. Sci. Data Discuss. 2014;7(2):521–610. doi: 10.5194/essdd-7-521-2014. [Google Scholar]
- Lenton A, et al. Sea-air co2 fluxes in the southern ocean for the period 1990–2009. Biogeosciences. 2013;10(6):4037–4054. doi: 10.5194/bg-10-4037-2013. [Google Scholar]
- Losch M, Menemenlis D, Campin J-M, Heimbach P, Hill C. On the formulation of sea-ice models. Part 1: Effects of different solver implementations and parameterizations. Ocean Model. 2010;33:129–144. doi: 10.1016/j.ocemod.2009.12.008. [Google Scholar]
- Marshall J, Adcroft A, Hill C, Perelman L, Heisey C. A finite-volume, incompressible Navier Stokes model for studies of the ocean on parallel computers. J. Geophys. Res. 1997;102:5753–5766. doi: 10.1029/96JC02775. [Google Scholar]
- Massachusetts Institute of Technology General Circulation Model Group. MITgcm User Manual (Online Documentation) Cambridge, Mass: MIT/EAPS; 2014. [Available at http://mitgcm.org/public/r2_manual/latest/online_documents/manual.html.] [Google Scholar]
- Meinshausen M, et al. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Change. 2011;109(1–2):213–241. doi: 10.1007/s10584-011-0156-z. [Google Scholar]
- Revelle R, Suess HE. Carbon dioxide exchange between atmosphere and ocean and the question of an increase of atmospheric CO2 during the past decades. Tellus. 1957;9(1):18–27. doi: 10.1111/j.2153-3490.1957.tb01849.x. [Google Scholar]
- Riebesell U, Körtzinger A, Oschlies A. Sensitivities of marine carbon fluxes to ocean change. Proc. Natl. Acad. Sci. U.S.A. 2009;106(49):20,602–20,609. doi: 10.1073/pnas.0813291106. doi: 10.1073/pnas.0813291106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodgers KB, Sarmiento JL, Aumont O, Crevoisier C, de Boyer Montégut C, Metzl N. A wintertime uptake window for anthropogenic CO2 in the North Pacific. Global Biogeochem. Cycles. 2008;22 GB2020, doi: 10.1029/2006GB002920. [Google Scholar]
- Sabine CL, et al. The oceanic sink for anthropogenic CO2. Science. 2004;305(5682):367–371. doi: 10.1126/science.1097403. [DOI] [PubMed] [Google Scholar]
- Sarmiento JL, Orr JC, Siegenthaler U. A perturbation simulation of CO2 uptake in an ocean general circulation model. J. Geophys. Res. 1992;97(C3):3621–3645. doi: 10.1029/91JC02849. [Google Scholar]
- Takahashi T, Sutherland S, Sweeney C, Poisson A, Metzl N, Tilbrook B, Bates N, Wanninkhof R, Feely R, Sabine C. Global sea-air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects. Deep Sea Res., Part II. 2002;49:1601–1622. doi: 10.1016/S0967-0645(02)00003-6. [Google Scholar]
- Takahashi T, et al. Climatological mean and decadal change in surface ocean pCO2, and net sea-air CO2 flux over the global oceans. Deep Sea Res., Part II. 2009;56:554–577. doi: 10.1016/j.dsr2.2008.12.009. [Google Scholar]
- Touratier F, Goyet C. Definition, properties, and Atlantic Ocean distribution of the new tracer TrOCA. J. Mar. Syst. 2004;46:169–179. [Google Scholar]
- Völker C, Wallace DWR, Wolf-Gladrow DA. On the role of heat fluxes in the uptake of anthropogenic carbon in the North Atlantic. Global Biogeochem. Cycles. 2002;16(4):85–1–85–9. doi: 10.1029/2002GB001897. [Google Scholar]
- Wallace D. Storage and transport of excess CO2 in the oceans: The JGOFS/WOCE global CO2 survey. In: Siedler J, Church J, Gould J, editors. Ocean Circulation and Climate: Observing and Modelling the Global Ocean. San Diego, Calif: Academic; 2001. pp. 489–521. [Google Scholar]
- Wallace DWR. Monitoring global ocean carbon inventories. OOSDP Background Report, no. 5. 1995 Texas A and M Univ., College Station, Tex. [Google Scholar]
- Wanninkhof R. Relationship between wind speed and gas exchange over the ocean. J. Geophys. Res. 1992;97:7373–7382. doi: 10.1029/92JC00188. [Google Scholar]
- Watanabe S, et al. MIROC-ESM 2010: Model description and basic results of CMIP5-20c3m experiments. Geosci. Model Dev. 2011;4(4):845–872. doi: 10.5194/gmd-4-845-2011. [Google Scholar]
- Waugh DW, Hall TM, McNeil BI, Key R, Matear RJ. Anthropogenic CO2 in the oceans estimated using transit time distributions. Tellus B. 2006;58(5):376–389. doi: 10.1111/j.1600-0889.2006.00222.x. [Google Scholar]
- Zeebe R, Wolf-Gladrow D. CO2 in Seawater: Equilibrium, Kinetics, Isotopes, Elsevier Oceanogr. Ser. Vol. 65. Amsterdam, Netherlands: Elsevier; 2001. p. 346. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Text S1