

Abstract
Kisspeptin is a neuropeptide that signals via a Gαq-coupled receptor, GPR54, in gonadotropin-releasing hormone (GnRH) neurons and is essential for pubertal maturation and fertility. Kisspeptin depolarizes and excites GnRH neurons primarily through the activation of canonical transient receptor potential (TRPC) channels and inhibition of K+ channels. The gonadal steroid 17β-estradiol (E2) up-regulates not only kisspeptin (Kiss1) mRNA, but also increases the excitability of the rostral forebrain Kiss1 neurons. In addition, a primary postsynaptic action of E2 on GnRH neurons is to up-regulate the expression of channel transcripts that orchestrate the downstream signaling of kisspeptin in GnRH neurons. These include not only TRPC4 channels, but also low voltage-activated T-type calcium channels and high voltage-activated L-, N- and R-type calcium channel transcripts. Moreover, E2 has direct membrane-initiated actions to alter the excitability of GnRH neurons by enhancing ATP-sensitive potassium (KATP) channel activity, which is critical for maintaining GnRH neurons in a hyperpolarized state for recruitment of T-type calcium channels that are important for burst firing. Therefore, E2 modulates the excitability of GnRH neurons as well as Kiss1 neurons by altering the expression and/or function of ion channels; and kisspeptin provides critical excitatory input to GnRH neurons to facilitate burst firing activity and peptide release.
Introduction
Gonadotropin-releasing hormone (GnRH) neurosecretion and the control of the ovulatory cycle in females is dependent on estrogen feedback, primarily 17β-estradiol (E2) secreted from the ovaries which reaches the pituitary and the brain via the circulation. E2 treatment in ovariectomized (OVX) females initially inhibits GnRH and LH secretion by the process known as negative feedback, followed by E2-induced super-secretion (positive feedback) in a species-specific manner 12-42h later [1-3]. With the development of mouse and rat models in which GnRH neurons express enhanced green fluorescent protein (GnRH-EGFP), it has been possible to systematically study these neurons in order to evaluate the GnRH neuronal excitability and activity with the ultimate goal of understanding the neuronal activity underlying the different secretory patterns utilized by these cells [4-6]. In addition, kisspeptin neurons, located in the anteroventral and more caudal periventricular preoptic area (AVPV/PeN), express the E2 receptor alpha (ERα), and E2 stimulates Kiss1 mRNA expression in this brain region [7]. Moreover, kisspeptin is one of the most potent excitatory neurotransmitters of GnRH neurons [8-11].
Based on decades of intracellular sharp electrode and whole-cell recordings in a number of parvocellular hypothalamic neurons, we have modeled currents crucial for rhythmic burst firing and have continued to explore the role of these currents in GnRH and more recently in Kiss1 neurons [12-14]. These currents include the low-threshold T-type calcium current (IT) the hyperpolarization-activated, cyclic nucleotide-gated current (Ih), a calcium-dependent, after-hyperpolarization potassium current (IAHP) as well as a persistent, sodium current (INaP), all of which will be further discussed in this review (Figure 1). All of these currents have been studied extensively in thalamic relay neurons, in which the T-current is responsible for low threshold calcium spikes and the h-current serves as the “pacemaker” to control the rate of rhythmic oscillations in these neurons [15-17]. Recently, several models have been developed that include an ensemble of channels that appear to be critical for burst firing in parvocellular neurosecretory neurons [12,18-20]. Variations of these models have been described in a number of reviews and primary publications [21-30] and therefore will not be extensively covered in this mini-review. Rather, this review will focus on the E2 modulation of both GnRH neurons and the presynaptic kisspeptin neurons, as well as discuss the role of kisspeptin as a unique excitatory neurotransmitter of GnRH neurons.
Figure 1. Schematic diagram illustrating the Kiss1-GnRH connection and signaling pathways responsible for kisspeptin-induced depolarization and burst firing of GnRH neurons.
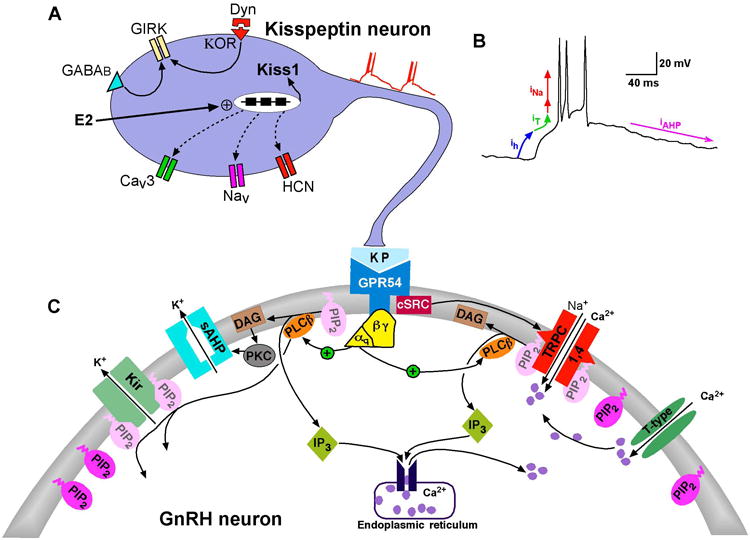
A. Kisspeptin neurons express the conductances (h-, T- and NaP) and corresponding channels (HCN, CaV3 and NaV) that generate burst firing necessary for maximum release of kisspeptin from the nerve terminal. E2 treatment increases the mRNA expression of Kiss1. In addition, the critical currents (Ih and IT.) are increased by circulating E2. The respective ion channel transcripts (HCN and CaV3, as well as NaV) are also expressed in Kiss1 neurons (dashed lines). E2 regulation of the mRNA expression of these channels is under investigation (manuscript in preparation). Kiss1 neurons also express GABAB and κ-opioid receptors, which couple to GIRK channels. B. Whole-cell current-clamp recording of spontaneous rebound burst firing recorded from an AVPV/PeN Kiss1 neuron. Arrows indicate the approximate range where the h-current (Ih, blue) and T-type calcium current (IT, green) and persistent Na current (INa, first red arrow) are active to depolarize the cell to threshold for generating an ensemble of fast Na+ spikes (INa, second red arrow) and AHP currents (IAHP, magenta) to repolarize the membrane to a hyperpolarized state to reinitiate the burst firing. The INa is composed of an initial INaP followed by an INaT (transient Na current). C. Kisspeptin binds to the Gq-coupled GPR54 receptor to activate phospholipase Cβ (PLCβ), which catabolizes PIP2, potentiates TRPC channel activity and inhibits the Kir channel activity. PKC, activated by the PIP2 hydrolysis product diacylglycerol (DAG), inhibits the activity of a calcium-activated slow afterhyperpolarization (sAHP) current. The non-receptor tyrosine (cSRC) kinase, which is activated by kisspeptin/GRP54 signaling, potentiates the activity of TRPC4 channels. Calcium entering the cell via T-type channels facilitates the activation of TRPC channels.
E2 modulation of GnRH neuronal activity through channel expression
As in thalamic neurons, IT, Ih, IAHP, and INaP are important for burst firing in GnRH as well as AVPV/PeN Kiss1 neurons [14,15,17,22,25,31-34]. We and others have continued to explore the modulation of these currents by E2 in GnRH and Kiss1 neurons in order to elucidate the conductances underlying burst firing activity.
Three subunits of the T-type calcium channel (Cav 3.1, 3.2, 3.3) have been cloned, with the specific gating properties of the channel being dependent on its subunit composition [35,36]. Thus, the kinetic properties of channels composed of Cav3.1 and 3.2 subunits are different from those of Cav3.3 subunits [37]. GnRH neurons express all three of the T-type calcium channel subunits, and the IT in GnRH neurons shows different types of inactivation kinetics, with the slow Cav3.3 channel being the most prominent of the three [32,38]. Of importance is that IT is present in the majority of GnRH neurons, and the membrane potential at which half of the T-type calcium channels are available for activation (i.e., are de-inactivated) is -80 mV [32]. As found in other neuronal systems, the role of IT is to initiate transient membrane depolarizations, which recruit high voltage-activated (HVA) calcium channels to increase calcium influx [39,40]. This is of functional significance since HVA calcium currents are prevalent in GnRH neurons [41], but they can only be activated from a more depolarized state, whereas low voltage-activated T-type currents are recruited from a more hyperpolarized state (i.e., they provide the signal for setting off burst firing) [32,38,41-44]. Importantly, the T-type CaV3.3 channel subunit expression and current density are significantly enhanced with a high LH surge-inducing E2 treatment, facilitating the excitation of GnRH neurons [32]. Therefore, the E2-dependent increase in T-type calcium channels may serve to augment the excitability of GnRH neurons in preparation for the GnRH (and LH) surge. In addition, the majority of HVA channel subtypes are expressed in GnRH neurons, and the mRNA levels of the L-type HVA calcium channel CaV1.3, the N-type HVA calcium channel CaV2.2, and the R-type HVA calcium channel CaV2.3 are also increased by LH surge-inducing E2 treatment [45]. Similarly, L- and N-type currents are increased by E2 during the afternoon as compared to morning in an E2-implant model that induces daily LH surges [41,46]. Collectively, these findings suggest a prominent role for calcium channels in the E2-induced excitation of GnRH neurons. GnRH neurons also express a hyperpolarization-activated, cyclic nucleotide-gated cation current (Ih), aka the pacemaker current, which also contributes to rhythmic firing [18,33,47,48]. Two Ih channel mRNAs, HCN1 and HCN2, are highly expressed in mouse GnRH neurons, and the mRNA expression of HCN1 is up-regulated by E2 [45]. Although E2 regulation of the h-current has not been evaluated in female GnRH neurons, Ih is up-regulated by E2 in Kiss1 neurons, an indication that this current is E2-sensitive [14,34].
Critical for hyperpolarizing the membrane to de-inactivate and recruit IT and activate Ih are the inwardly rectifying K+ (Kir) channels. It is well known that Kir channels are vital for maintaining excitable cells in a hyperpolarized resting state closer to the Nernst equilibrium potential for potassium (EK+), but once cells are depolarized the channels allow for the quick transition to long depolarizing responses because of their inwardly rectifying properties [49]. With greater membrane hyperpolarization of GnRH neurons, there is an increase in the number of Na+ spikes generated during the rebound excitation (Zhang et al; unpublished observations). Indeed, KATP and G protein-coupled inwardly rectifying potassium (GIRK) channels appear to be the critical Kir channels in GnRH neurons for maintaining the membrane in a relatively more negative resting state. Blocking KATP channels significantly depolarizes GnRH neurons, which is indicative of tonic KATP channel activity that is significantly augmented with E2 treatment [48]. Also, GABA, opioids, neuropeptide Y and perhaps melanin-concentrating hormone inputs via their respective GABAB, μ-opioid, Y5 and MCH1 receptors activate GIRK channels to hyperpolarize GnRH neurons (Lagrange et al 1995, Wu etal 2009, Zhang et al 2009, Xu et al 2008) [47,50-52]. This combination of hyperpolarizing influences would allow for the recruitment of multiple excitatory conductances that are critical for generating burst firing in CNS neurons [15,18].
Neuronal excitability is also determined by the afterhyperpolarization (AHP) that follows an action potential, and the three kinds of calcium-activated potassium channels that mediate AHPs have been identified in GnRH neurons [25,53-56]. This includes the fast AHP, which is mediated by the large-conductance Ca2+-activated K+ (BK) channels, the medium AHP (mAHP), which is mediated by small conductance, calcium-activated K+ (SK) channels, and the slow AHP (sAHP), which is mediated by a yet unidentified potassium channel. The BK channels appear to be involved in the repolarization of action potentials in GnRH neurons [57]. Inhibition of SK channels by apamin directly attenuates the mAHP as well as augments the ADP in GnRH neurons, and, thus, these channels exert a significant influence on the firing properties of these neurons [58,59]. Of the SK channel subtypes, only SK3 mRNA exhibits significant expression in GnRH neurons, and surge-inducing E2 treatments decrease the expression and function of this channel in ovariectomized females [45,59]. The channel responsible for the slow AHP current has not been identified. This current, however, slows or limits the firing frequency (called spike frequency adaptation) and is a major player controlling neuronal excitability [60]. A reduced AHP current allows a faster trajectory to burst firing of GnRH neurons [25,56,59]. Thus, the up-regulation of low voltage-activated (CaV3.3) and high voltage-activated calcium (CaV1.3, 2.2, 2.3) and hyperpolarization-activated cation (HCN1) channels and the down-regulation of SK3 channels by E2 play a key role in increasing GnRH neuronal excitability.
Rapid 17β-estradiol modulation of GnRH neuronal activity
The relatively fast (∼15 min) inhibition of GnRH and LH secretion by E2 is congruent with its initiation of a membrane signaling cascade. In fact years ago, it was found that guinea pig GnRH neurons are rapidly hyperpolarized by E2 via activation of an inwardly-rectifying (G protein-coupled) K+ (GIRK) conductance in the presence of tetrodotoxin (TTX), which blocks fast Na+ channel activity and isolates GnRH neurons from action potential- driven synaptic inputs [47,61,62]. In mice, physiological concentrations (picomolar) of E2 rapidly augment KATP (also of the inwardly rectifying family) channel activity to hyperpolarize GnRH neurons [63]. E2 signals via a protein kinase C (PKC)- protein kinase A (PKA) pathway, and hence a selective Gαq-mER ligand called STX is also able to mimic the effects of E2 in GnRH as in other hypothalamic neurons [63-66]. Both the effects of E2 and STX are abrogated by the estrogen receptor antagonist ICI 182,780 with a Ki of 0.5 nM in GnRH neurons [63,67], which is similar to the Ki for antagonism of ERα [68]. Interestingly, the membrane-initiated signaling by E2 in GnRH neurons appears to persist given that female mice treated with an E2 implant for 4-7 days similarly exhibit augmentation of the KATP current, which attenuates GnRH neuronal firing [48]. This would imply that membrane-initiated signaling by E2 is involved in negative feedback regulation of GnRH neurons.
In addition, nanomolar concentrations of E2 enhance action potential firing by modulating intrinsic afterhyperpolarizing and afterdepolarizing potentials via a PKA-dependent mechanism involving ERβ [59]. Picomolar concentrations of E2 also inhibit action potential firing via presynaptic ERα-dependent mechanisms [59]. E2 rapidly potentiates high-voltage-activated (HVA) Ca2+ currents (L- and R-type Ca2+ channels) via ERβ and a G protein-coupled receptor of 30 KD molecular weight (GPR30), suggesting that Ca2+ signaling is also a target for E2 membrane actions in GnRH neurons [41]. However, in slices prepared from GnRH-Pericam transgenic mice, E2 initiates calcium transients within 15 min in a subgroup of GnRH neurons [69]. In this transgenic mouse model the E2 effects are mediated indirectly via input from GABA neurons that express ERα. The role of ERβ to mediate rapid effects of E2 on GnRH neurons has also been shown following E2 treatment in vivo [70]. E2 rapidly (15 min) phosphorylates cAMP response element binding protein (pCREB) in GnRH neurons, an effect that is lost in ERβ-deleted animals, supporting a direct role for ERβ in mediating rapid E2 signaling in GnRH neurons [70]. However, how pCREB affects gene expression and cell excitability is not known.
In primate and mouse embryonic nasal explants (the source of immature GnRH neurons) E2 modulates Ca2+ oscillations in GnRH neurons [71-74], which synchronize with a periodicity of approximately 60 minutes in primates, a rhythm that is similar to the pulsatile GnRH release [71,73-75]. Furthermore, nanomolar concentrations of a membrane-impermeant E2 (E2-dendrimer, EDC) and the Gq-mER ligand STX alter the patterns of Ca2+ oscillations in primate GnRH neurons [75,76]. As found previously with E2, STX also elicits an increase in the frequency and synchronization of Ca2+ oscillations in rhesus macaque GnRH neurons [75,77]. The STX-induced modulation of Ca2+ oscillations (and GnRH release) from macaque nasal explants is not altered by GPR30 small interfering RNA transfection, suggesting that GPR30 is not involved. Importantly, the effects of STX are blocked by the ER antagonist ICI182,780 and by the PLC inhibitor U73122 [75]. This would suggest that STX (E2) is activating Gq-mER (as defined above) in rhesus macaque GnRH neurons that induces calcium oscillations via the IP3 receptor (Figure 1) and/or a PKC signaling pathway [78]. Therefore, although the specific role of membrane-initiated E2 signaling for GnRH physiology is uncertain, it could potentially play a critical role in sculpting GnRH burst firing activity.
Kisspeptin-GnRH neuronal circuitry
Kisspeptin-54 is the endogenous ligand of G protein-coupled receptor 54 (GPR54, aka Kiss1R) [79]. GPR54 is highly expressed in GnRH neurons [45], and mutations in GPR54 cause autosomal recessive idiopathic hypogonadism in humans and deletion of GPR54 or Kiss1 in mice results in defective sexual development and reproductive failure [80,81]. The Kiss1 gene encodes a 145 amino acid protein, which is proteolytically processed to Kisspeptin-54 and several other smaller peptide fragments, and centrally administered kisspeptins robustly stimulate GnRH and gonadotropin secretion in both prepubertal and adult animals [82,83]. Neurons in the AVPV/PeN areas express kisspeptin, GABA and opioid peptides all of which are important for regulation of GnRH neurosecretion [84-89]. The AVPV/PeN expresses high levels of ERα and also ERβ, and the actions of the gonadal steroids on kisspeptin neurons are mediated, in part, via nuclear-initiated signaling (genomic) mechanisms [90-92]. Also, Kiss1 mRNA expression is greatly increased in the AVPV/PeN following E2 treatment [7]. These findings combined with previous observations that lesions or anti-estrogen implants of the AVPV/PeN in rodents abrogate the positive feedback effects of E2 [93-96], have led to the hypothesis that E2 acts on AVPV/PeN Kiss1 neurons to induce positive feedback on GnRH and LH secretion. Notably, an E2 treatment that induces a LH surge increases both the h -current and T-type calcium current in the AVPV/PeN Kiss1 neurons [14,34].
Kisspeptin excites GnRH neurons primarily through activation of TRPC channels and to a lesser extent through inhibition of inwardly rectifying K+ channels (Figure 1) [8-11,78,97]. In addition, kisspeptin induces a transient elevation of intracellular calcium in GnRH neurons, which is thought to be due to release from intracellular calcium stores and has been hypothesized to play an important role in the kisspeptin-mediated depolarization [9]. However, the activation of TRPC channels in GnRH neurons by kisspeptin is not affected by buffering intracellular calcium levels by the calcium chelators EGTA or BAPTA or by depleting intracellular calcium stores [11,78,98]. Therefore, release of calcium from intracellular stores does not appear to play a critical role in the kisspeptin-mediated activation of TRPC channels, but may be involved in Ca2+/calmodulin-dependent inhibition of high voltage-activated Ca2+ channels [99]. The kisspeptin-activated TRPC current is attenuated by the general calcium channel blocker Cd2+ and by the low voltage-activated calcium channel blocker Ni2+, but not by the high voltage-activated calcium channel blocker amlodipine [11,98]. This would indicate that low voltage-activated (T-type) calcium channels may be involved in facilitating TRPC channel opening. However, reducing extracellular calcium to nominally calcium free has no effect on the kisspeptin-activated TRPC current, an indication that very little calcium is needed to enable the opening of TRPC channels in GnRH neurons. This is consistent with the small, but persistent, T-type calcium channel activity (window current) around -65 mV in GnRH neurons [32]. Therefore, with a sustained depolarization that exceeds that of classical neurotransmitters (e.g., glutamate), kisspeptin excites GnRH neurons primarily through the opening of a TRPC channel that is independent of intracellular calcium store release, but appears to be dependent on transient calcium influx through T-type calcium channels (Figure 1). This ensures a fast and yet sustained depolarization of GnRH neurons.
Although GnRH neurons express all of the TRPC channel subunits that are found in the brain (i.e., TRPC1,3,4,5,6,7), quanitative PCR analysis shows that TRPC4 is the main TRPC channel subtype in GnRH neurons and is expressed at levels 4-fold higher than TRPC1 and TRPC5 [11,45]. In fact, TRPC4 mRNA is increased in high E2-treated mice [45]. Phosphatidylinositol-4,5-biphosphate (PIP2) is an important regulator of TRPC channels, and depletion of PIP2 is required for kisspeptin-induced TRPC channel activation in GnRH neurons [98]. In addition to PIP2 depletion, kisspeptin activation of TRPC channels is also dependent on the non-receptor tyrosine (cSrc) kinase activation (Figure 1), since both global tyrosine kinase inhibitors such as genistein and the specific cSrc kinase inhibitor PP2 attenuate (inhibit) kisspeptin currents in GnRH neurons [98]. cSrc kinase directly regulates TPRC4 channel activity through tyrosine phosphorylation, which causes rapid insertion of TRPC4 channels into the plasma membrane [100]. Therefore, cSrc appears to be a key signaling molecule in the kisspeptin-mediated activation of TRPC channels in GnRH neurons.
PKC is an important second messenger stimulated by Gq-coupled receptors. However, PKC appears not to be involved in TRPC channel activation in cell lines expressing cloned TRPC channels [101]. Similarly in GnRH neurons, PKC activation or inhibition has only minor or no effect on the kisspeptin-induced inward TRPC current [11,98].
Since the initial studies showing that kisspeptin has prolonged effects on GnRH neuronal activity [8-11], the question has been why is there very little spike frequency adaptation (slowing of action potential firing due to activation of K+ channels) during kisspeptin-induced sustained firing? Recently it was shown that kisspeptin reduces spike frequency adaptation and prolongs firing via the inhibition of a calcium-activated slow afterhyperpolarization current (IsAHP) via PKC [56]. GnRH neurons express two distinct IsAHP, an apamin-sensitive [25,56] and a kisspeptin-sensitive IsAHP [56]. Essentially, kisspeptin inhibits 50% of the IsAHP, and apamin inhibits the other 50% of the current [56]. Therefore, in addition to increasing the firing rate through an overt depolarization, kisspeptin facilitates sustained firing through inhibiting an apamin-insensitive IsAHP in GnRH neurons (via PKC). The physiological significance is that although single action potential-generated calcium influx is sufficient to spark the release of classical neurotransmitters, burst firing or tetanic stimulation is required for the release of neuropeptides as first eloquently shown by Wakerly and Lincoln in vivo [102] and then in vitro by others [103-105].
Role of pacemaker currents in Kiss1 neurons
Kiss1 neurons in the AVPV/PeN appear to be presynaptic pacemaker neurons that drive GnRH neurons. At least 50% of GnRH neurons receive monosynaptic input from the AVPV/PeN neurons, some of which have been identified as Kiss1 neurons [106]. Importantly, higher frequency stimulation of AVPV/PeN neurons elicits a delayed excitatory response in GnRH neurons, which can be blocked by the kisspeptin inhibitory peptide 318 [106]. Recently, the development of Kiss1-CreGFP knock-in mice has allowed the targeting of GFP-expressing Kiss 1 neurons for more detailed analysis [107] (see [30] for review). Both Ih and IT are prominently expressed in Kiss1 neurons [14,34], and the Kiss1 neurons also express the corresponding critical transcripts, HCN1 and Cav3.1 [14]. In particular, CaV3.1 calcium channels are highly expressed in AVPV/PeN Kiss1 neurons, and are exceptionally sensitive to E2 [14] (and Zhang et al., manuscript in preparation). The presence of a robust T-type calcium current is essential for the high-frequency rebound bursting that is manifested following a hyperpolarizing stimulus in these neurons [14].
In addition, Kiss1 neurons express a persistent Na+ current (INaP) that is activated ∼10 mV negative to the threshold for the transient sodium current in a membrane potential range in which few voltage-gated channels are activated and neuronal input resistance is high [14]. Thus, INaP tends to amplify the depolarization induced by T-type calcium channels. Most importantly in terms of providing excitatory drive to GnRH neurons, an E2 treatment that produces a LH surge, significantly increases both Ih and IT in AVPV/PeN Kiss1 neurons by 3.4- and 6-fold, respectively [14,34]. AVPV/PeN Kiss1 neurons rest at a relatively positive resting membrane potential of -55 mV. Thus, some sort of robust inhibitory synaptic input is necessary for reaching hyperpolarized membrane potentials to recruit a critical mass of Cav3.1 (by removing their inactivation) and HCN1 channels for initiating burst generation [14,34].
Indeed, GABA via GABAB receptors (Gαi.o-coupled to GIRK channels), can provide the hyperpolarizing stimulus for generating burst firing [14]. GABA neurons are abundant in the hypothalamus, and the majority of Kiss1 neurons in the AVPV/PeN express GAD 67 (glutamic acid decarboxylase, the GABA synthesizing enzyme) [108]. As such, Kiss1 neurons may themselves be an endogenous source of GABA, whose action could be auto-synaptic and perhaps be responsible for hyperpolarizing Kiss1 neurons via GABAB receptors. Although further studies are needed to elucidate all of the physiological mechanisms underlying high-frequency burst firing of AVPV/PeN Kiss1 neurons, it is clear that these neurons express the critical channels and receptors that permit E2-dependent burst firing. Therefore, Kiss1 neurons, similar to thalamocortical neurons, express the critical pacemaker conductances, and as such Kiss1 neurons have the capacity to generate bursting activity and excite GnRH neurons, which supports the idea that Kiss1 neurons provide the excitatory drive for GnRH surge secretion.
Summary
What is critical for the control of GnRH neuronal excitability and ultimately the control of fertility is the hypothalamic circuitry. This “circuitry” not only includes the synaptic input to kisspeptin and GnRH neurons, but also the effects of circulating E2, which conveys vital feedback information about reproductive states, on the excitability of these neurons. All AVPV/PeN kisspeptin neurons express the endogenous burst-generating conductances (Ih, and IT, and INaP) that allow these vital neurons to generate burst firing and release kisspeptin, which in turn excites GnRH neurons that also express Ih, IT and INaP [22,32,48,106]. A future challenge is not only to identify all of key channels and signaling molecules and how they are regulated by E2, but also how these channels fit into the kisspeptin-GnRH neuronal circuitry for generation of burst firing and peptide release.
Acknowledgments
The authors thank current and previous members of their laboratories who contributed to the work described in the present review. We also thank Dr. Casey C Nestor for helpful comments on earlier version of the manuscript, and Ms. Marina V. Rulevskaya and Ms. Uyen-Vy Navarro for their excellent technical support. The work from the authors' laboratories was supported by PHS grants NS 43330, NS 38809 and DK 68098.
References
- 1.Terasawa E, Rodriguez JS, Bridson WE, Wiegand SJ. Factors influencing the positive feedback action of estrogen upon luteinizing hormone surge in the ovariectomized guinea pig. Endocrinology. 1979;104:680–686. doi: 10.1210/endo-104-3-680. [DOI] [PubMed] [Google Scholar]
- 2.Caraty A, Locatelli A, Martin GB. Biphasic response in the secretion of gonadotrophin-releasing hormone in ovariectomized ewes injected with oestradiol. J Endocrinol. 1989;123:375–382. doi: 10.1677/joe.0.1230375. [DOI] [PubMed] [Google Scholar]
- 3.Chappell PE, Levine JE. Stimulation of gonadotropin-releasing hormone surges by estrogen. I Role of hypothalamic progesterone receptors. Endocrinology. 2000;141:1477–1485. doi: 10.1210/endo.141.4.7428. [DOI] [PubMed] [Google Scholar]
- 4.Spergel DJ, Kruth U, Hanley DF, Sprengel R, Seeburg PH. GABA-and glutamate activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 1999;19:2037–2050. doi: 10.1523/JNEUROSCI.19-06-02037.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Suter KJ, Song WJ, Sampson TL, Wuarin JP, Saunders JT, Dudek FE, Moenter SM. Genetic targeting of green fluorscent protein to gonadotropin-releasing hormone neurons: Characterization of whole-cell electrophysiological properties and morphology. Endocrinology. 2000;141:412–419. doi: 10.1210/endo.141.1.7279. [DOI] [PubMed] [Google Scholar]
- 6.Fujioka H, Suzuki M, Yamanouchi K, Ohta A, Nagashima H, Kato M, Nishihara M. Generation of transgenic rats expressing enhanced green fluorescent protein in gonadotropin-releasing hormone neurons. J Reprod Dev. 2003;49:523–529. doi: 10.1262/jrd.49.523. [DOI] [PubMed] [Google Scholar]
- 7.Smith JT, Cunningham MJ, Rissman EF, Clifton DK, Steiner RA. Regulation of Kiss1 gene expression in the brain of the female mouse. Endocrinology. 2005;146:3686–3692. doi: 10.1210/en.2005-0488. [DOI] [PubMed] [Google Scholar]
- 8.Han SK, Gottsch ML, Lee KJ, Popa SM, Smith JT, Jakawich SK, Clifton DK, Steiner RA, Herbison AE. Activation of gonadotropin-releasing hormone neurons by kisspeptin as a neuroendocrine switch for the onset of puberty. J Neurosci. 2005;25:11349–11356. doi: 10.1523/JNEUROSCI.3328-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Liu X, Lee K, Herbison AE. Kisspeptin excites gonadotropin-releasing hormone (GnRH) neurons through a phospholipase C/calcium-dependent pathway regulating multiple ion channels. Endocrinology. 2008;149:4605–4614. doi: 10.1210/en.2008-0321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pielecka-Fortuna J, Chu Z, Moenter SM. Kisspeptin acts directly and indirectly to increase gonadotropin-releasing hormone neuron activity and its effects are modulated by estradiol. Endocrinology. 2008;149:1979–1986. doi: 10.1210/en.2007-1365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang C, Roepke TA, Kelly MJ, Rønnekleiv OK. Kisspeptin depolarizes gonadotropin-releasing hormone neurons through activation of TRPC-like cationic channels. J Neurosci. 2008;28:4423–4434. doi: 10.1523/JNEUROSCI.5352-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kelly MJ, Rønnekleiv OK. Electrophysiological analysis of neuroendocrine neuronal activity in hypothalamic slices. In: Levine JE, editor. Methods in Neurosciences: Pulsatility in Neuroendocrine Systems. San Diego: Academic Press, Inc.; 1994. pp. 47–67. [Google Scholar]
- 13.Bosch MA, Zhang C, Rønnekleiv OK. Regulation of endogenous conductances in GnRH neurons by estrogens. Brain Res. 2010;1364:25–34. doi: 10.1016/j.brainres.2010.08.096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhang C, Tonsfeldt KJ, Qiu J, Bosch MA, Kobayashi K, Steiner RA, Kelly MJ, Rønnekleiv OK. Molecular mechanisms that drive estradiol-dependent burst firing of Kiss1 neurons in the rostral periventricular preoptic area. Am J Physiol Endocrinol Metab. 2013;305:E1384–E1397. doi: 10.1152/ajpendo.00406.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McCormick DA, Huguenard JR. A model of the electrophysiological properties of thalamocortical relay neurons. J Neurophysiol. 1992;68:1384–1400. doi: 10.1152/jn.1992.68.4.1384. [DOI] [PubMed] [Google Scholar]
- 16.Lüthi A, McCormick DA. H-current: properties of a neuronal and network pacemaker. Neuron. 1998;21:9–12. doi: 10.1016/s0896-6273(00)80509-7. [DOI] [PubMed] [Google Scholar]
- 17.Kim D, Song I, Keum S, Lee T, Jeong MJ, Kim SS, McEnery MW, Shin HS. Lack of the burst firing of thalamocortical relay neurons and resistance to absence seizures in mice lacking alpha(1G) T-type Ca(2+) channels. Neuron. 2001;31:35–45. doi: 10.1016/s0896-6273(01)00343-9. [DOI] [PubMed] [Google Scholar]
- 18.Kelly MJ, Wagner EJ. GnRH neurons and episodic bursting activity. Trends Endocrinol Metab. 2002;13:409–410. doi: 10.1016/s1043-2760(02)00698-7. [DOI] [PubMed] [Google Scholar]
- 19.Lyons DJ, Horjales-Araujo E, Broberger C. Synchronized network oscillations in rat tuberoinfundibular dopamine neurons: swith to tonic discharge by thyrotropin-releasing hormone. Neuron. 2010;65:217–229. doi: 10.1016/j.neuron.2009.12.024. [DOI] [PubMed] [Google Scholar]
- 20.Chen X, Iremonger K, Herbison A, Kirk V, Sneyd J. Regulation of electrical bursting in a spatiotemporal model of a GnRH neuron. Bull Math Biol. 2013;75:1941–1960. doi: 10.1007/s11538-013-9877-7. [DOI] [PubMed] [Google Scholar]
- 21.Moenter SM, Anthony DR, Pitts GR, Nunemaker CS. Mechanisms underlying episodic gonadotropin-releasing hormone secretion. Front Neuroendocrinol. 2003;24:79–93. doi: 10.1016/s0091-3022(03)00013-x. [DOI] [PubMed] [Google Scholar]
- 22.Chu Z, Moenter SM. Physiologic regulation of a tetrodotoxin-sensitive sodium influx that mediates a slow afterdepolarization potential in gonadotropin-releasing hormone neurons: possible implications for the central regulation of fertility. J Neurosci. 2006;26:11961–11973. doi: 10.1523/JNEUROSCI.3171-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Popa SM, Clifton DK, Steiner RA. The role of kisspeptins and GPR54 in the neuroendocrine regulation of reproduction. Annu Rev Physiol. 2008;70:14.1–14.26. doi: 10.1146/annurev.physiol.70.113006.100540. [DOI] [PubMed] [Google Scholar]
- 24.Colledge WH. Kisspeptins and GnRH neuronal signalling. Trends Endocrinol Metab. 2009;3:115–121. doi: 10.1016/j.tem.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 25.Lee K, Duan W, Sneyd J, Herbison AE. Two slow calcium-activated afterhyperpolarization currents control burst firing dynamics in gonadotropin-releasing hormone neurons. J Neurosci. 2010;30:6214–6224. doi: 10.1523/JNEUROSCI.6156-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Herbison AE, Moenter SM. Depolarising and hyperpolarising actions of GABAA receptor activation on gonadotrophin-releasing hormone neurons: towards an emerging consensus. J Neuroendocrinology. 2011;23:557–569. doi: 10.1111/j.1365-2826.2011.02145.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pielecka-Fortuna J, DeFazio RA, Moenter SM. Voltage-gated Potassium currents are targets of diurnal changes in estradiol feedback reglation and Kisspeptin action on Gonadotropin-releasing hormone neurons in mice. Biol Repro. 2011;85:987–995. doi: 10.1095/biolreprod.111.093492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chu Z, Tomaiuolo M, Bertram R, Moenter SM. Two types of burst firing in gonadotropin-releasing hormone neurones. J Neuroendocrinology. 2012;24:1065–1077. doi: 10.1111/j.1365-2826.2012.02313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Frazão R, Cravo RM, Donato J, Jr, Ratra DV, Clegg DJ, Elmquist JK, Zigman JM, Williams KW, Elias CF. Shift in kiss1 cell activity requires estrogen receptor α. J Neurosci. 2013;33:2807–2820. doi: 10.1523/JNEUROSCI.1610-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Piet R, de Croft S, Liu X, Herbison AE. Electrical properties of kisspeptin neurons and their regulation of GnRH neurons. Front Neuroendocrinol. 2014 doi: 10.1016/j.yfrne.2014.05.006. [DOI] [PubMed] [Google Scholar]
- 31.Huguenard JR, Prince DA. A novel T-type current underlies prolonged Ca2+-dependent burst firing in GABAergic neurons of rat thalamic reticular nucleus. J Neurosci. 1992;12:3804–3817. doi: 10.1523/JNEUROSCI.12-10-03804.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhang C, Bosch MA, Rick EA, Kelly MJ, Rønnekleiv OK. 17β-estradiol regulation of T-type calcium channels in gonadotropin-releasing hormone neurons. J Neurosci. 2009;29:10552–10562. doi: 10.1523/JNEUROSCI.2962-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chu Z, Takagi H, Moenter SM. Hyperpolarization-activated currents in gonadotropin-releasing hormones (GnRH) neurons contribute to intrinsic excitability and are regulated by gonadal steroid feedback. J Neurosci. 2010;30:13373–13383. doi: 10.1523/JNEUROSCI.1687-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Piet R, Boehm U, Herbison AE. Estrous cycle plasticity in the hyperpolarization-activated current Ih is mediated by circulating 17β-estradiol in preoptic area kisspeptin neurons. J Neurosci. 2013;33:10828–10839. doi: 10.1523/JNEUROSCI.1021-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Perez-Reyes E, Cribbs LL, Daud A, Lacerda AE, Barclay J, Williamson MP, Margaret F, Rees M, Lee JH. Molecular characterization of a neuronal low-voltage-activated T-type calcium channel. Nature. 1998;391:896–900. doi: 10.1038/36110. [DOI] [PubMed] [Google Scholar]
- 36.Lee JH, Daud AN, Cribbs LL, Lacerda AE, Pereverzev A, Klockner U, Schneider T, Perez-Reyes E. Cloning and expression of a novel member of the low voltage-activated T-type calcium channel family. J Neurosci. 1999;19:1912–1921. doi: 10.1523/JNEUROSCI.19-06-01912.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chemin J, Monteil A, Perez-Reyes E, Bourinet E, Nargeot J, Lory P. Specific contribution of human T-type calcium channel isotypes (alpha(1G), alpha(1H) and alpha(1I)) to neuronal excitability. J Physiol. 2002;540:3–14. doi: 10.1113/jphysiol.2001.013269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kato M, Ui-Tei K, Watanabe M, Sakuma Y. Characterization of voltage-gated calcium currents in gonadotropin-releasing hormone neurons tagged with green fluorescent protein in rats. Endocrinology. 2003;144:5118–5125. doi: 10.1210/en.2003-0213. [DOI] [PubMed] [Google Scholar]
- 39.Cueni L, Canepari M, Lujan R, Emmenegger Y, Watanabe M, Bond CT, Franken P, Adelman JP, Luthi A. T-type ca2+ channels, sk2 channels and sercas gate sleep-related oscillations in thalamic dendrites. Nature Neurosci. 2008;11:683–692. doi: 10.1038/nn.2124. [DOI] [PubMed] [Google Scholar]
- 40.Liu S, Shipley MT. Multiple conductances cooperatively regulate spontaneous bursting in mouse olfactory bulb external tufted cells. J Neurosci. 2008;28:1625–1639. doi: 10.1523/JNEUROSCI.3906-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sun J, Chu Z, Moenter SM. Diurnal in vivo and rapid in vitro effects of estadiol on voltage-gated calcium channels in gonadotropin-releasing hormone neurons. J Neurosci. 2010;30:3912–3923. doi: 10.1523/JNEUROSCI.6256-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nunemaker CS, DeFazio RA, Moenter SM. Calcium current subtypes in GnRH neurons. Biol Reprod. 2003;69:1914–1922. doi: 10.1095/biolreprod.103.019265. [DOI] [PubMed] [Google Scholar]
- 43.Spergel DJ. Calcium and small-conductance calcium-activated potassium channels in gonadotropin-releasing hormone neurons before, during, and after puberty. Endocrinology. 2007;148:2383–2390. doi: 10.1210/en.2006-1693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tanaka N, Ishii H, Yin C, Koyama M, Sakuma Y, Kato M. Voltage-gated Ca2+ channel mRNAs and T-type Ca2+ currents in rat gonadotropin-releasing hormone neurons. J Physiol Sci. 2010;60:195–204. doi: 10.1007/s12576-010-0085-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bosch MA, Tonsfeldt KJ, Rønnekleiv OK. mRNA expression of ion channels in GnRH neurons: subtype-specific regulation by 17β-Estradiol. Mol Cell Endocrinol. 2013;367:85–97. doi: 10.1016/j.mce.2012.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Christian CA, Mobley JL, Moenter SM. Diurnal and estradiol-dependent changes in gonadotropin-releasing hormone neuron firing activity. Proc Natl Acad Sci USA. 2005;102:15682–15687. doi: 10.1073/pnas.0504270102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lagrange AH, Rønnekleiv OK, Kelly MJ. Estradiol-17β and μ-opioid peptides rapidly hyperpolarize GnRH neurons: A cellular mechanism of negative feedback? Endocrinology. 1995;136:2341–2344. doi: 10.1210/endo.136.5.7720682. [DOI] [PubMed] [Google Scholar]
- 48.Zhang C, Bosch MA, Levine JE, Rønnekleiv OK, Kelly MJ. Gonadotropin-releasing hormone neurons express K(ATP) channels that are regulated by estrogen and responsive to glucose and metabolic inhibition. J Neurosci. 2007;27:10153–10164. doi: 10.1523/JNEUROSCI.1657-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hille B. Potassium channels and chloride channels. In: Hille B1, editor. Ion channels of excitable membranes. Sunderland, MA: Sinaur; 2001. pp. 131–165. [Google Scholar]
- 50.Wu M, Dumalska I, Morozova E, Van den Pol AN, Alreja M. Melanin-concentrating hormone directly inhibits GnRH neurons and blocks kisspeptin activation, linking energy balance to reproduction. Proc Natl Acad Sci USA. 2009;106:17217–17222. doi: 10.1073/pnas.0908200106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang C, Bosch MA, Rønnekleiv OK, Kelly MJ. GABAB receptor mediated inhibition of GnRH neurons is suppressed by kisspeptin-GPR54 signaling. Endocrinology. 2009;150:2388–2394. doi: 10.1210/en.2008-1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Xu J, Kirigiti MA, Cowley MA, Grove KL, Smith MS. Suppression of basal spontaneous gonadotropin-releasing hormone neuronal activity during lactation: role of inhibitory effects of neuropeptide Y. Endocrinology. 2009;150:333–340. doi: 10.1210/en.2008-0962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bosch MA, Kelly MJ, Rønnekleiv OK. Distribution, neuronal co-localization and 17β-E2 modulation of small conductance calcium-activated K+ channel (SK3) mRNA in the guinea pig brain. Endocrinology. 2002;143:1097–1107. doi: 10.1210/endo.143.3.8708. [DOI] [PubMed] [Google Scholar]
- 54.Kato M, Tanaka N, Usui S, Sakuma Y. SK channel blocker apamin inhibits slow afterhyperpolarization currents in rat gonadotropin-releasing hormone neurones. J Physiol. 2006;574(2):431–442. doi: 10.1113/jphysiol.2006.110155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang Y, Kuehl-Kovarik MC. Flufenamic acid modulates multiple currents in gonadotropin-releasing hormone neurons. Brain Res. 2010;1353:94–105. doi: 10.1016/j.brainres.2010.07.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zhang C, Rønnekleiv OK, Kelly MJ. Kisspeptin inhibits a slow afterhyperpolarization current via protein kinase C and reduces spike-frequency adaptation in GnRH neurons. Am J Physiol Endocrinol Metab. 2013;304:E1237–E1244. doi: 10.1152/ajpendo.00058.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hiraizumi Y, Nishimura I, Ishii H, Tanaka N, Takeshita T, Sakuma Y, Kato M. Rat GnRH neurons exhibit large conductance voltage- and Ca2+-Activated K+ (BK) currents and express BK channel mRNAs. J Physiol Sci. 2008;58:21–29. doi: 10.2170/physiolsci.RP013207. [DOI] [PubMed] [Google Scholar]
- 58.Liu X, Herbison AE. Small-conductance calcium-activated potassium channels control excitability and firing dynamics in gonadotropin-releasing hormone (GnRH) neurons. Endocrinology. 2008;149:3598–3604. doi: 10.1210/en.2007-1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Chu Z, Andrade J, Shupnik MA, Moenter SM. Differential regulation of gonadotropin-releasing hormone neuron activity and membrane properties by acutely applied estradiol: dependence on dose and estrogen receptor subtype. J Neurosci. 2009;29:5616–5627. doi: 10.1523/JNEUROSCI.0352-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sah P, Faber ESL. Channels underlying neuronal calcium-activated potassium currents. Prog Neurobiol. 2002;66:345–353. doi: 10.1016/s0301-0082(02)00004-7. [DOI] [PubMed] [Google Scholar]
- 61.Kelly MJ, Rønnekleiv OK, Eskay RL. Identification of estrogen-responsive LHRH neurons in the guinea pig hypothalamus. Brain Res Bull. 1984;12:399–407. doi: 10.1016/0361-9230(84)90112-6. [DOI] [PubMed] [Google Scholar]
- 62.Condon TP, Rønnekleiv OK, Kelly MJ. Estrogen modulation of the α1-adrenergic response of hypothalamic neurons. Neuroendo. 1989;50:51–58. doi: 10.1159/000125201. [DOI] [PubMed] [Google Scholar]
- 63.Zhang C, Kelly MJ, Rønnekleiv OK. 17β-estradiol rapidly increases ATP-sensitive potassium channel activity in gonadotropin-releasing hormone neurons via a protein kinase signaling pathway. Endocrinology. 2010;151:4477–4484. doi: 10.1210/en.2010-0177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lagrange AH, Rønnekleiv OK, Kelly MJ. Modulation of G protein-coupled receptors by an estrogen receptor that activates protein kinase A. Mol Pharmacol. 1997;51:605–612. doi: 10.1124/mol.51.4.605. [DOI] [PubMed] [Google Scholar]
- 65.Qiu J, Bosch MA, Tobias SC, Grandy DK, Scanlan TS, Rønnekleiv OK, Kelly MJ. Rapid signaling of estrogen in hypothalamic neurons involves a novel G protein-coupled estrogen receptor that activates protein kinase C. J Neurosci. 2003;23:9529–9540. doi: 10.1523/JNEUROSCI.23-29-09529.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Qiu J, Bosch MA, Tobias SC, Krust A, Graham S, Murphy S, Korach KS, Chambon P, Scanlan TS, Rønnekleiv OK, Kelly MJ. A G protein-coupled estrogen receptor is involved in hypothalamic control of energy homeostasis. J Neurosci. 2006;26:5649–5655. doi: 10.1523/JNEUROSCI.0327-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Rønnekleiv OK, Zhang C, Kelly MJ. Estradiol and kisspeptin modulation of gonadotropin-releasing hormone (GnRH) neuronal excitability. In: Armstong WE, Tasker JG, editors. Neurophysiology of Neuroendocrine Neurons. Hoboken, NJ: John Wiley and Sons, Ltd.; 2014. pp. 301–321. [Google Scholar]
- 68.Weatherill PJ, Wilson APM, Nicholson RI, Davies P, Wakeling AE. Interaction of the antioestrogen ICI 164,384 with the oestrogen receptor. J Ster Bioc Mol Biol. 1988;30:263–266. doi: 10.1016/0022-4731(88)90103-3. [DOI] [PubMed] [Google Scholar]
- 69.Romanò N, Lee K, Ábrahám IM, Jasoni CL, Herbison AE. Non-classical estrogen modulation of presynaptic GABA terminals modulates calcium dynamics in gonadotropin-releasing hormone (GnRH) neurons. Endocrinology. 2008;149:5335–5344. doi: 10.1210/en.2008-0424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Abraham IM, Han SK, Todman MG, Korach KS, Herbison AE. Estrogen receptor beta mediates rapid estrogen actions on gonadotropin-releasing hormone neurons in vivo. J Neurosci. 2003;23:5771–5777. doi: 10.1523/JNEUROSCI.23-13-05771.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Terasawa E, Schanhofer WK, Keen KL, Luchansky LL. Intracellular Ca2+ oscillations in luteinizing hormone-releasing hormone neurons derived from the embryonic olfactory placode of the rhesus monkey. J Neurosci. 1999;19:5898–5909. doi: 10.1523/JNEUROSCI.19-14-05898.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Richter TA, Keen KL, Terasawa E. Synchronization of Ca2+ oscillations among primate LHRH neurons and nonneuronal cells in vitro. J Neurophysiol. 2002;88:1559–1567. doi: 10.1152/jn.2002.88.3.1559. [DOI] [PubMed] [Google Scholar]
- 73.Temple JL, Laing E, Sunder A, Wray S. Direct action of estradiol on gonadotropin-releasing hormone-1 neuronal activity via a transcription-dependent mechanism. J Neurosci. 2004;24:6326–6333. doi: 10.1523/JNEUROSCI.1006-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Temple JL, Wray S. BSA-estrogen compounds differentially alter gonadotropin-releasing hormone-1 neuronal activity. Endocrinology. 2005;146:558–563. doi: 10.1210/en.2004-1117. [DOI] [PubMed] [Google Scholar]
- 75.Kenealy BP, Keen KL, Rønnekleiv OK, Terasawa E. STX, a novel nonsteroidal estrogenic compound, induces rapid action in primate GnRH neuronal calcium dynamics and peptide release. Endocrinology. 2011;152:182–191. doi: 10.1210/en.2011-0097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Abe H, Keen KL, Terasawa E. Rapid action of estrogens on intracellular calcium oscillations in primate LHRH-1neurons. Endocrinology. 2008;149:1155–1162. doi: 10.1210/en.2007-0942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kenealy BP, Keen KL, Terasawa E. Rapid action of estradiol in primate GnRH neurons: the role of estrogen receptor alpha and estrogen receptor beta. Steroids. 2011;76:861–866. doi: 10.1016/j.steroids.2011.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Constantin S, Caligioni CS, Stojilkovic S, Wray S. Kisspeptin-10 facilitates a plasma membrane-driven calcium oscillator in gonadotropin-releasing hormone-1 neurons. Endocrinology. 2009;150:1400–1412. doi: 10.1210/en.2008-0979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kotani M, Detheux M, Vandenbogaerde A, Communi D, Vanderwinden JM, Le Poul E, Brezillon S, Tyldesley R, Suarez-Huerta N, Vandeput F, Blanpain C, Schiffmann SN, Vassart G, Parmentier M. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J Biol Chem. 2001;276:34631–34636. doi: 10.1074/jbc.M104847200. [DOI] [PubMed] [Google Scholar]
- 80.Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O'Rahilly S, Carlton MBL, Crowley WF, Aparicio SAJR, Colledge WH. The GPR54 gene as a regulator of puberty. N Engl J Med. 2003;349:1614–1627. doi: 10.1056/NEJMoa035322. [DOI] [PubMed] [Google Scholar]
- 81.d'Anglemont de Tassigny X, Fagg LA, Dixon JPC, Day K, Leitch HG, Hendrick AG, Zahn D, Franceschini I, Caraty A, Carlton MBL, Aparicio SAJR, Colledge WH. Hypogonadotropic hypogonadism in mice lacking a functional KiSS 1 gene. Proc Natl Acad Sci USA. 2007;104:10714–10719. doi: 10.1073/pnas.0704114104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gottsch ML, Cunningham MJ, Smith JT, Popa SM, Acohido BV, Crowley WF, Seminara S, Clifton DK, Steiner RA. A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology. 2004;145:4073–4077. doi: 10.1210/en.2004-0431. [DOI] [PubMed] [Google Scholar]
- 83.Kinoshita M, Tsukamura H, Adachi S, Matsui H, Uenoyama Y, Iwata K, Yamada S, Inoue K, Ohtaki T, Matsumoto H, Maeda KI. Involvement of central metastin in the regulation of preovulatory luteinizing hormone surge and estrous cyclicity in female rats. Endocrinology. doi: 10.1210/en.2005-0195. [DOI] [PubMed] [Google Scholar]
- 84.Simerly RB, McCall LD, Watson SJ. Distribution of opioid peptides in the preoptic region: immunohistochemical evidence for a steroid-sensitive enkephalin sexual dimorphism. J Comp Neurol. 1988;276:442–459. doi: 10.1002/cne.902760309. [DOI] [PubMed] [Google Scholar]
- 85.Wagner EJ, Rønnekleiv OK, Bosch MA, Kelly MJ. Estrogen biphasically modifies hypothalamic GABAergic function concomitantly with negative and positive control of luteinizing hormone release. J Neurosci. 2001;21:2085–2093. doi: 10.1523/JNEUROSCI.21-06-02085.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Jackson GL, Kuehl D. Gamma-aminobutyric acid (GABA) regulation of GnRH secretion in sheep. Reproduction. 2002;59:15–24. [PubMed] [Google Scholar]
- 87.DeFazio RA, Heger S, Ojeda SR, Moenter SM. Activation of A-type gamma-aminobutyric receptors excites gonadotropin-releasing hormone neurons. Mol Endocrinol. 2002;16:2872–2891. doi: 10.1210/me.2002-0163. [DOI] [PubMed] [Google Scholar]
- 88.Smith JT, Popa SM, Clifton DK, Hoffman GE, Steiner RA. Kiss1 neurons in the forebrain as central processors for generating the preovulatory luteinizing hormone surge. J Neurosci. 2006;26:6687–6694. doi: 10.1523/JNEUROSCI.1618-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Christian CA, Moenter SM. Estradiol induces diurnal shifts in GABA transmission to gonadotropin-releasing hormone neurons to provide a neural signal for ovulation. J Neurosci. 2007;27:1913–1921. doi: 10.1523/JNEUROSCI.4738-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogen receptor-α and -β mRNA in the rat central nervous system. J Comp Neurol. 1997;388:507–525. doi: 10.1002/(sici)1096-9861(19971201)388:4<507::aid-cne1>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 91.Wintermantel TM, Campbell RE, Porteous R, Bock D, Gröne HJ, Todman MG, Korach KS, Greiner E, Perez CA, Schultz G, Herbison AE. Definition of estrogen receptor pathway critical for estrogen positive feedback to gonadotropin-releasing hormone neurons and fertility. Neuron. 2006;52:271–280. doi: 10.1016/j.neuron.2006.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Clarkson J, d'Anglemont de Tassigny X, Moreno AS, Colledge WH, Herbison AE. Kisspeptin-GPR54 signaling is essential for preovulatory gonadotropin-releasing hormone neuron activation and the luteinizing hormone surge. J Neurosci. 2008;28:8691–8697. doi: 10.1523/JNEUROSCI.1775-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Wiegand SJ, Terasawa E, Bridson WE. Persistent estrus and blockade of progesterone-induced LH release follows lesions which do not damage the suprachiasmatic nucleus. Endocrinology. 1978;102:1645–1648. doi: 10.1210/endo-102-5-1645. [DOI] [PubMed] [Google Scholar]
- 94.Rønnekleiv OK, Kelly MJ. Plasma prolactin and luteinizing hormone profiles during the estrous cycle of the female rat: effects of surgically induced persistent estrus. Neuroendo. 1988;47:133–141. doi: 10.1159/000124903. [DOI] [PubMed] [Google Scholar]
- 95.Petersen SL, Barraclough CA. Suppression of spontaneous LH surges in estrogen-treated ovariectomized rats by microimplants of antiestrogens into the preoptic brain. Brain Res. 1989;484:279–289. doi: 10.1016/0006-8993(89)90371-5. [DOI] [PubMed] [Google Scholar]
- 96.Ma YJ, Kelly MJ, Rønnekleiv OK. Pro-gonadotropin-releasing hormone (ProGnRH) and GnRH Content in the preoptic area and the basal hypothalamus of anterior medial preoptic nucleus/suprachiasmatic nucleus-lesioned persistent estrous rats. Endocrinology. 1990;127:2654–2664. doi: 10.1210/endo-127-6-2654. [DOI] [PubMed] [Google Scholar]
- 97.Kroll H, Bolsover S, Hsu J, Kim SH, Bouloux PM. Kisspeptin-evoked calcium signals in isolated primary rat gonadotropin-releasing hormone neurones. Neuroendo. 2011;93:114–120. doi: 10.1159/000321678. [DOI] [PubMed] [Google Scholar]
- 98.Zhang C, Bosch MA, Rønnekleiv OK, Kelly MJ. Kisspeptin activation of TRPC4 channels in female GnRH neurons requires PIP2 depletion and cSrc kinase activation. Endocrinology. 2013;154:2772–2783. doi: 10.1210/en.2013-1180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zhang XB, Spergel DJ. Kisspeptin inhibits high-voltage activated Ca2+ channels in GnRH neurons via multiple Ca2+ influx and release pathways. Neuroendo. 2012;96:68–80. doi: 10.1159/000335985. [DOI] [PubMed] [Google Scholar]
- 100.Odell AF, Scott JL, Van Helden DF. Epidermal growth factor induces tyrosine phosphorylation, membrane insertion, and activation of transient receptor potential channel 4. J Biol Chem. 2012;280:37974–37987. doi: 10.1074/jbc.M503646200. [DOI] [PubMed] [Google Scholar]
- 101.Birnbaumer L. The TRPC class of ion channels: a critical review of their roles in slow, sustained increases in intracellular Ca2+ concentrations. Annu Rev Pharmacol Toxicol. 2009;49:395–426. doi: 10.1146/annurev.pharmtox.48.113006.094928. [DOI] [PubMed] [Google Scholar]
- 102.Wakerley JB, Lincoln DW. The milk-ejection reflex of the rat: a 20- to 40- fold acceleration in the firing of paraventricular neurones during oxytocin release. J Endo. 1973;57:477–493. doi: 10.1677/joe.0.0570477. [DOI] [PubMed] [Google Scholar]
- 103.Bicknell RJ. Optimizing release from peptide hormone secretory nerve terminals. J exp Biol. 1988;139:51–65. doi: 10.1242/jeb.139.1.51. [DOI] [PubMed] [Google Scholar]
- 104.Shakiryanova D, Tully A, Hewes RS, Deitcher DL, Levitan ES. Activity-dependent liberation of synaptic neuropeptide vesicles. Nat Neurosci. 2005;8:173–178. doi: 10.1038/nn1377. [DOI] [PubMed] [Google Scholar]
- 105.Masterson SP, Li J, Bickford ME. Frequencey-dependent release of substance P mediates heterosynaptic potentiation of glutamatergic synaptic responses in the rat visual thalamus. J Neurophysiol. 2010;104:1758–1767. doi: 10.1152/jn.00010.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Liu X, Porteous R, d'Anglemont de Tassigny X, Colledge WH, Millar R, Petersen SL, Herbison AE. Frequency-dependent recruitment of fast amino acid and slow neuropeptide neurotransmitter release controls gonadotropin-releasing hormone neuron excitability. J Neurosci. 2011;31:2421–2430. doi: 10.1523/JNEUROSCI.5759-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Gottsch ML, Popa SM, Lawhorn JK, Qiu J, Tonsfeldt KJ, Bosch MA, Kelly MJ, Rønnekleiv OK, Sanz E, McKnight GS, Clifton DK, Palmiter RD, Steiner RA. Molecular properties of Kiss1 neurons in the arcuate nucleus of the mouse. Endocrinology. 2011;152:4298–4309. doi: 10.1210/en.2011-1521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Cravo RM, Margatho LO, Osborne-Lawrence S, Donato J, Jr, Atkin S, Bookout AL, Rovinsky S, Frazão R, Lee CE, Gautron L, Zigman JM, Elias CF. Characterization of Kiss1 neurons using transgenic mouse models. Neurosci. 2011;173:37–56. doi: 10.1016/j.neuroscience.2010.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]