

Abstract
Background
Estrogen receptors alpha and beta (ERα and ERβ) differentially activate genes with AP-1 elements. ERα activates AP-1 targets via activation functions with estrogens (the AF-dependent pathway), whereas ERβ, and a short version of ERα (ERα DBD-LBD) activate only with anti-estrogens (AF-independent pathway). The DNA binding domain (DBD) plays an important role in both pathways, even though neither pathway requires ERE recognition.
Results
Mutations of a highly conserved DBD lysine (ERα.K206A/G), lead to super-activation of AP-1 through activation function dependent pathways, up to 200 fold. This super-activity can be elicited either through ER AFs 1 or 2, or that of a heterologous activation function (VP16). The homologous substitution in ERβ, K170A, or in ERα DBD-LBD leads to estrogen-dependent AP-1 activation and loss of the usually potent anti-estrogen effects. Each of numerous K206 substitutions in ERα, except K206R, eliminates anti-estrogen activation and this loss correlates perfectly with a loss of ability to titrate a repressive function from the RU486 bound progesterone receptor.
Conclusion
We conclude that ER DBDs contain a complex regulatory function that influences ligand activation profiles at AP-1. This function, which requires the integrity of the conserved lysine, both allows for activation at AP-1 with anti-estrogens (with ERβ and ERα DBD-LBD), and prevents ERα from becoming superactive at AP-1 with estrogens. We discuss the possibility that a repressor interaction with the DBD both mediates the AF-independent pathway and dampens the AF dependent pathway. Mutations in the conserved lysine might, by this model, disrupt the binding or function of the repressor.
Background
Estrogen receptors alpha and beta (ERα and ERβ) classically activate transcription by binding to cognate estrogen response elements (EREs). The receptors also activate expression of target genes through alternate pathways, e.g. through AP-1 and CRE-like response elements [1,2]. AP-1/CRE-like elements bind Jun and related transcription factors but not ERs [2,3]. ER action at these alternate elements appears increasingly to be important. For example, ER/AP-1 pathways underlie estrogen activation of collagenase, Cyclin D1 and IGF-1genes [1,2,4]. The relative contributions of classic and alternate pathways to a given gene response in vivo have not yet been directly explored. However, there is suggestive evidence that AP-1 alternate pathways play an important role in estrogen-dependent proliferation [1,5-7].
There are two distinct ER/AP-1 pathways: one activated by estrogens, the other activated by anti-estrogens [1,3,8]. The estrogen-activated ERα/AP-1 pathway is mediated by ERα's two activation functions: the constitutive AF-1 in the N'terminal domain (NTD), and the estrogen-activated AF-2 in the C-terminal ligand binding domain (LBD) [8]. The ligand dependent AF-2 is functional in virtually all cell types but is blocked by anti-estrogens. Cells of endometrial origin also support strong AF-1 activity at AP-1 which is allowed by some anti-estrogens most notably tamoxifen (Tam) but not by raloxifene or ICI 182,780. These latter two "pure" anti-estrogens do not permit AF-1 activity most likely because they allow for efficient binding of the corepressor N-CoR [9]. It is inferred therefore, that when these pure anti-estrogens activate AP-1 mediated transcription (with ERβ or ERα DBD-LBD) they do so through an AF-independent mechanism possibly one that involves N-CoR binding since a mutation that eliminates N-CoR binding eliminates this anti-estrogen activation [9].
To summarize our previous work, we have identified two pathways by which the ER can activate transcription at AP-1 elements, AF dependent and AF independent. Three classes of ligands can be distinguished since the AP-1 targets can be activated: (1) By estrogens through an AF-1 and -2 dependent pathway; (2) By Tam through AF-1, and partly through the AF-independent activity, and (3) By "pure" anti-estrogens (ICI and Raloxifene) that activate only through an AF-independent pathway [9]. The latter pathway is most prominent in the presence of ERβ and certain ERαNTD-deleted mutants [8].
The mechanisms that underlie the AF-independent activation have not been elucidated. However, we have suggested that anti-estrogen-ER complexes increase AP-1 activity by functionally inactivating an unidentified co-repressor normally associated with the AP-1 complex and/or its chromatin environment [3,8]. The lack of AF involvement in this pathway leaves open the question as to which portions of the ER contribute to this mode of anti-estrogen action through AP-1. Here we present data that a domain generally not considered to contain transcriptional activation functions, the DNA binding domain (DBD), plays a critical role in regulating the balance between different pathways of ER action at AP-1 sites.
We noted that nuclear receptor DBDs contain a highly conserved lysine (lys) residue at the base of the first zinc binding motif (Fig. 1A). In the context of the glucocorticoid receptor (GR) this residue is required for GR's ability to inhibit AP-1 mediated transcriptional activation. Its mutation to gly or ala converts the GR from an inhibitor to a stimulator of the AP-1 activity in response to agonist [10,11]. We noted in a preliminary report that the equivalent ERα mutation (ERα.K206A) potentiates estrogen action at AP-1 sites, and at hybrid AP-1/cAMP response elements (CREs) in the context of much larger promoter, Cyclin D1 [2]. Here we report that mutations of this residue show complex effects on the ligand profile of ER action at AP-1 sites and propose that DBD interacting proteins could regulate the type of ER pathway used at alternate response elements.
Figure 1.

ERαK206A and K206G mutations lead to super-stimulation at AP-1 sites through the estrogen pathway. (A) Location of K206 at the base of the first zinc binding domain in the ERα DBD. (B) ERαK206A and ERα G400V.K206A ligand profiles in HeLa cells. The panel shows results of HeLa cell transfections in which activity of ERα, and ERα G400V with or without the K206A mutation, were assessed at the Coll73-Luc reporter (shown in schematic above), in the presence of a range of ligands. Note: ERα refers to the fully wild type human estrogen receptor (Fomerly pHEG0). All mutations in this manuscript are in this background unless performed in the human ERα that contains a G400V mutation in the ligand binding domain. This latter construct is designated ERαG400V (Formerly pHE0). "No ER" indicates cells transfected exactly as in all others in the experiment except that an empty expression vector (pSG5) is transfected instead of the same vector containing receptor cDNA, as indicated. Vehicle treatments (No H.) are represented by white bars, ICI by light grey bars, Ral by medium grey bars, Tam by dark grey bars and E2 by black bars. The figure represents a single representative experiment with standard deviations calculated from multiple wells. (C, D) ERα.K206A and ERα.K206G are both superactive at AP-1 sites, but not EREs. (C) The panel shows results from a transfection in which activities of ERα with and without the K206A/G mutations were assessed at an AP-1 site (Coll73-Luc reporter). (D) Results of a similar transfection in which activities of the same ERs were assessed at an ERE (ERE-Luc). The panel represents a single experiment with average results obtained from triplicate wells.
Results
ERα.K206A and K206G are superactive at AP-1 sites but not at EREs
We first mutated the conserved ERα lysine residue at the base of the first zinc finger to ala or gly (ERα.K206A or K206G, Fig. 1A). Introduction of these mutations was motivated by reports that mutations in the homologous residue convert rat and human GRs from inhibitors to activators at AP-1 regulated genes [10,11]. Data in Fig. 1B confirm that both E2-liganded ERα.K206A and ERα.G400V.K206A enhance the activity of an AP-1 responsive reporter gene (Coll73-Luc) more efficiently than their K206 counterparts in HeLa cells. Also apparent was the inhibitory nature of the anti-estrogen ligands used in presence of the K206A mutation, vide infra for further discussion.
ERα.K206A also showed increased activity in the absence of exogenous ligands. Fully wild type ERα. G400 (ERα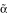 designated as ERα throughout the remainder of the manuscript unless otherwise indicated) exhibits variable levels of constitutive activity in cell culture because of very low levels of estrogens in serum [12]. This phenomenon probably accounts for the increased ability of unliganded ERα bearing the K206A mutation to enhance AP-1 dependent transcription because a mutation (G400V) that slightly reduces ERα affinity for ligand [12] abolished the constitutive activity exhibited by the fully wild type receptor, without affecting overall E2 response (compare ERαG400 to ERαG400V with and with out the K206A mutation, Fig. 1B). ERα.K206G also showed marked ability to enhance AP-1 activity in response to E2 (Fig. 1C). As mentioned above, the degree of constitutive activity displayed by ERα is variable as seen by comparing Fig 1C to 1B. However, regardless of the amount of constitutive activity, treatment with E2 always increased the estrogen response in the presence of the ERα.K206. Additionally, it exhibited enhanced activity at the Cyclin D1 promoter (not shown, [2]). Curiously, of all mutations tested, be they DBD deletions, chimeras, or substitution of K206 to other residues, only K206A/G sustained superactivity at AP-1 (Figs. 1C, 5A and data not shown). In distinction, at an ERE, estrogen-bound ERα.K206A and G showed mildly reduced to normal levels of activity (Fig. 1D). Thus, K206A and K206G mutations enhance ERα and ERαG400V activity at AP-1 and CRE-like elements but not at EREs.
designated as ERα throughout the remainder of the manuscript unless otherwise indicated) exhibits variable levels of constitutive activity in cell culture because of very low levels of estrogens in serum [12]. This phenomenon probably accounts for the increased ability of unliganded ERα bearing the K206A mutation to enhance AP-1 dependent transcription because a mutation (G400V) that slightly reduces ERα affinity for ligand [12] abolished the constitutive activity exhibited by the fully wild type receptor, without affecting overall E2 response (compare ERαG400 to ERαG400V with and with out the K206A mutation, Fig. 1B). ERα.K206G also showed marked ability to enhance AP-1 activity in response to E2 (Fig. 1C). As mentioned above, the degree of constitutive activity displayed by ERα is variable as seen by comparing Fig 1C to 1B. However, regardless of the amount of constitutive activity, treatment with E2 always increased the estrogen response in the presence of the ERα.K206. Additionally, it exhibited enhanced activity at the Cyclin D1 promoter (not shown, [2]). Curiously, of all mutations tested, be they DBD deletions, chimeras, or substitution of K206 to other residues, only K206A/G sustained superactivity at AP-1 (Figs. 1C, 5A and data not shown). In distinction, at an ERE, estrogen-bound ERα.K206A and G showed mildly reduced to normal levels of activity (Fig. 1D). Thus, K206A and K206G mutations enhance ERα and ERαG400V activity at AP-1 and CRE-like elements but not at EREs.
Figure 5.

Anti-estrogen stimulation is highly sensitive to mutations at K206. (A) Results of a HeLa cell transfection in which the phenotype of ERα and ERα.K206A are compared with other ERα.K206 mutants. Note that ERα.K206A shows superactivity in the presence of E2 and little or no activity in the presence of anti-estrogens. Other K206 mutations, with the exception of K206 mutation to a like amino acid (K206R), convert anti-estrogen stimulation to repression despite the lack of E2-associated superactivity. (B) Alterations in ERα steady state levels do not account for altered phenotypes. HeLa cells were transfected with 10 μg of each expression vector, induced with appropriate ligands for 24 hrs and processed for Western blotting. The panels show individual exposures of autoradiograms of Western blots of HeLa cell extracts transfected with individual ERα mutants or DBD-LBD and its K206A equivalent.
ERα enhances AP-1 activity in the presence of E2 in a variety of cell lines ([3] and references therein). ERα also enhances AP-1 activity in the presence of the partial agonist/antagonist Tam in cell lines that support AF-1 activity at AP-1 sites [1,7]. Since the K206A mutation enhances estrogen activity, we determined whether the ERα.K206A mutation would also amplify Tam action at AP-1. Consistent with our previous results [1], endogenous ERα activated AP-1 in the presence of both E2 and Tam in Ishikawa cells (Figs. 2A insert) whereas endogenous ERα activated the AP-1 responsive reporter only in the presence of E2 in MCF-7 breast cells, which do not support AF-1 activity at AP-1 (Fig. 2B, insert). Co-transfection of both cell types with the K206A mutation amplified these responses. In Ishikawas the K206A mutation enhanced both Tam and E2 response, whereas in MCF-7s ERα.K206A primarily enhanced the E2 response. The MCF-7 cell response is like that seen in HeLas (Fig. 1B), which also have very little AF-1 activity. Enhanced tamoxifen response was also supported by MDA-MB-435 (a breast cell line; Fig. 3A) and chicken embryo fibroblasts (CEFs; data not shown) both of which support AF-1 activity. We conclude that the ERα.K206A mutation amplifies E2 responses at AP-1 sites and also amplifies the agonist properties of the mixed agonist/antagonist Tam in cells that support AF-1 activity at AP-1 sites.
Figure 2.

ERαK206A super-stimulates in cells that contain endogenous ERs. Both Ishikawa (A) and MCF-7 cells (B) were transfected with Coll73-Luc and treated with ligands as indicated in Fig. 1B. The insets show the effect of treatment with endogenous receptor only. The main graphs show ligand effect in the presence of the indicated transfected ERα (5 μg).
Figure 3.

ERαK206A super-stimulation requires activation functions. (A) Truncation of AF-1 abolishes Tam response to ERα.K206A at AP-1 sites in MDA-MB-453 cells, which support AF-1 activity. (B) In HeLa cells, mutation of AF-2, also in the ERα.K206A background, abrogates K206A activity. Cells were transfected with ERs bearing K206A mutants in the context of full-length receptor, or the ERα DBD-LBD truncation lacking the NTD, which contains AF-1. (C) HeLa cells were transfected with ERα.DBD-LBD or its K206A equivalent, or a VP16-DBD-LBD fusion or its K206A equivalent. Note that E2, but not Tam, responses are amplified by the K206A mutation in the context of the DBD-LBD truncation but E2 and Tam responses are amplified by the K206A mutation in the context of the VP16-DBD-LBD fusion. Results of a representative experiment are shown.
ERα.K206A activity is dependent on linked activation functions, either endogenous or exogenous
We next determined whether the increased activity elicited by the ERα K206A mutation was truly dependent on ER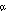 activation functions. As indicated above, the ERα.K206A mutation amplifies Tam responses at AP-1 sites in cells that support AF-1 activity. To further test the dependence of Tam's effect on AF-1, we evaluated the effect of an ERα AF-1 truncation (DBD-LBDα.K206A). This mutation completely abolished Tam response in MDA-MB-453 cells (Fig. 3A). The DBD-LBDαK206A also exhibited diminished E2 response (which in turn, was abolished by mutation of AF-2; not shown). Similar results were also obtained in Ishikawa, MDA-MB-231 and CEF cells, all cells known to support AF-1 activity (not shown). Thus, ERαK206A-dependent Tam effects at AP-1 sites require AF-1. In these conditions AF-1 contributes to overall level of E2 activation, as well, perhaps by synergizing with AF-2.
activation functions. As indicated above, the ERα.K206A mutation amplifies Tam responses at AP-1 sites in cells that support AF-1 activity. To further test the dependence of Tam's effect on AF-1, we evaluated the effect of an ERα AF-1 truncation (DBD-LBDα.K206A). This mutation completely abolished Tam response in MDA-MB-453 cells (Fig. 3A). The DBD-LBDαK206A also exhibited diminished E2 response (which in turn, was abolished by mutation of AF-2; not shown). Similar results were also obtained in Ishikawa, MDA-MB-231 and CEF cells, all cells known to support AF-1 activity (not shown). Thus, ERαK206A-dependent Tam effects at AP-1 sites require AF-1. In these conditions AF-1 contributes to overall level of E2 activation, as well, perhaps by synergizing with AF-2.
As suggested by the data above, the K206A mutation exerts its effects via both AFs 1 and 2 in the presence of E2 (Figs. 3A). To test for the contribution of AF-2, we used a double hydrophobic mutation within the ERα LBD C-terminal helix 12 (M543A;L544A) that completely abolishes ERα AF-2 activity at classical EREs [13]. We performed this experiment in HeLa cells, which only support minimal AF-1 activity. This mutation completely abolished ERα.K206A action at AP-1 sites in HeLa cells (Fig. 3B). Thus, ERα.K206A enhances AP-1 activity in HeLa cells in a manner that is completely dependent on AF-2. These observations indicate that the enhancement of AP-1 activity by the K206A ERα DBD is dependent on linked activation functions AF-1 and AF-2.
We then investigated whether the K206A mutation would amplify the activity of a heterologous activation function. Substitution of a strong heterologous activation function for the ERα N-terminus (VP16) failed to enhance Tam response at AP-1 sites in HeLa cells. In contrast, fusion of the same activation function to the DBD-LBDαK206A N-terminus produced robust Tam activation (Fig 3C). Thus, the VP16 activation function when fused to the ER DBD is nearly inactive at AP-1unless the DBD carries the K206A mutation. This observation suggests that the K206A mutation may relieve a repressive function mediated through the DBD exerted on linked activation functions when ERα activates at AP-1. This is further explored below.
The conserved lysine is required for AF-independent anti-estrogen action at AP-1 sites
ERβ enhances AP-1 activity in the presence of anti-estrogens, but not estrogens ([5] and Fig. 4A). We have suggested that these effects require ERβ contacts with corepressors one of which is N-CoR [3,8,9,14]. Since the ERα and ERβ DBDs are highly conserved, we examined the effect of mutating the conserved lysine (ERβ.K170) on ERβ action at AP-1 sites. In marked contrast to wild type ERβ, the ERβ.K170A eliminated anti-estrogen enhanced AP-1 activity and strongly enhanced E2 AP-1 activity (Fig. 4A). Thus, the ERβ.K170A mutation completely reverses the ligand profile of ERβ action at AP-1 sites.
Figure 4.

The ERβ.K170A mutation reverses ERβ ligand activation profile at AP-1. (A) Results of a HeLa cell transfection in which activity of ERβ (530 amino acid isoform) and an equivalent K170A mutation (shown in schematic at left) are compared at an AP-1 responsive reporter (Coll73-Luc). (B) Results of a similar HeLa cell transfection in which the full ligand activation profile of the ERα DBD-LBD region and its K206A equivalent are compared at AP-1 sites.
ERα truncations that lack portions of the NTD also show potent anti-estrogen responses at AP-1 sites [8]. Fig. 4B shows that in the absence of the NTD, the K206A mutation abolished potent anti-estrogen effects and, as for ERβ (Fig. 4A), allowed for enhanced activation with E2. Thus, K > A mutations of the conserved lysine residue at the base of the first zinc binding motif both enhance E2/AF-dependent and eliminate anti-estrogen/AF-independent activity of ER α and β at AP-1.
Most substitutions of the K206 residue abolish anti-estrogen activation without conferring super-activation at AP-1
As noted earlier both K206A and K206G were superactive at AP-1/CRE targets. We next explored a gamut of substitutions at the K206 residue to see whether they would behave similarly. Surprisingly, K206A or K206G were the sole mutations capable of generating superactivity in the estrogen pathway (Fig. 1B &1C and 5A). All other amino acid conversions either left the ability of ERα to stimulate AP-1 activity relatively intact (K206H, F), or blocked ERα activity (K206P, E, and L; Fig. 5A). These data suggest that mutations in the ER DBD fall into three classes with respect to their effects on the AF-dependent responses at AP-1 sites: those that super-stimulate (A, G), those that permit relatively normal stimulation (H, F) and those that fail to stimulate (P, E, and L). These findings point to considerable complexity in the DBD's affects on AF-dependent ER activity at AP-1 sites.
In stark contrast to the mixture of phenotypes in the presence of estrogen, with one exception, all mutations of the K206 residue eliminated the AF-independent anti-estrogen activation (Fig. 5A). The sole exception was K206R, mutation to a like amino acid, which maintained anti-estrogen activity (Fig. 5A). In addition to mutations at the K206 residue, we have observed that other changes in this domain, such as deletion or point mutation of an individual zinc finger lead to a similar loss of anti-estrogen activation without creating super-activity with estrogen (data not shown). These observations underscore the hypothesis that the two ER/AP-1 pathways, estrogen and anti-estrogen, are completely distinct.
The effects of K206 substitutions on anti-estrogen activation at AP-1 sites are not a consequence of decreased ERα protein levels. Fig. 5B confirms that ERα steady state levels are reduced by ICI and Ral and increased by Tam. The K206A mutation did modestly increase full-length ERα steady state levels in the presence of E2. However, similar increases were observed with the other ERα mutations that do not show superactivity at AP-1 sites (such as ERα.K206H). Lastly, protein levels of the N'terminal domain (AF-1) deletion mutants with and without the K206A mutation were virtually indistinguishable (Fig. 5B). Thus, the dramatic loss of ICI and raloxifene responses exhibited by the DBD-LBDα.K206A mutant (Fig. 4B) cannot be explained by reduced protein levels (Fig. 5B). Overall, our results suggest that effects of the K206A mutation on steady state levels of ERα are unlikely to explain ERα.K206A behavior at AP-1.
Substitutions at K206 that abolish the AF-independent anti-estrogen activation at AP-1, also abolish the ability of the anti-estrogen bound ERα to titrate repressors from the progesterone receptor
We previously proposed that AF-independent anti-estrogen activation at AP-1 sites involves functional inactivation of corepressors [3,8,9]. ERα-anti-estrogen complexes can titrate corepressors from another antagonist-bound steroid receptor, the PR-RU486 complex, and thereby increase its activity ([9,15] and Fig. 6, insert). We therefore tested whether the K206A substitutions would affect the ability of ERα to sequester corepressors from the PR-RU486 complex. Data in Fig. 6 confirm that ERα-anti-estrogen complexes derepress the PR-RU486 complex at a PRE. Strikingly, they further indicate that these effects are abolished by nearly all K206 substitutions the sole exception being ERα.K206R. This particular mutation (K > R) was also the only mutation that preserved activation through the ERα/AP-1 anti-estrogen pathway (Fig. 5A). Thus, we conclude that there is a perfect correlation between the ability of K206 substitutions to eliminate AF-independent anti-estrogen action at AP-1 and to eliminate the ability of ERα to titrate repressors from PR.
Figure 6.

K206 mutations abrogate the ability of ERα to titrate a repressive function from PR. (Insert) The experimental system: HeLa cells were co-transfected with a TAT3-Luc reporter that contains three progestin response elements (PREs), a PR expression vector (5 μg), and a CMV β-galactosidase internal control (2 μg). Transfections also included ERα, ERα K206 mutants or empty pSG5 vector ("No ER"; 5 μg each). (Graph) All cells were treated with the PR antagonist RU486 (10-6 M), which promotes recruitment of repressive functions to DNA by PR. The repressive functions are competed for by the ER-bound anti-estrogens, as indicated. The data is expressed as relative luciferase activity, where the ratio of luciferase activity to β-galactosidase activity in the absence of ER ligand is set to a value of 1.
The K > A similarly affects TR transcriptional activity and the ability of the receptor to interact with a repressor
To extend these studies, we examined the effect of a K > A mutation of the analogous residue in thyroid hormone receptor-alpha (TRα), which is known to interact strongly with a co-repressor complex that includes N-CoR or SMRT. Fig. 7A confirms our previous observation that unliganded chicken TRα enhances AP-1 activity and that addition of T3 abolishes unliganded TRα activity and further represses AP-1 activity below basal [16]. Introduction of an alanine substitution at the K206 homologue (TRα.K72A; Fig. 7A) abolished unliganded TRα activity, and actually allowed unliganded TRα to suppress AP-1 activity below basal, just as the ERα.K206A mutation abolished anti-estrogen activation and allowed anti-estrogens to suppress AP-1 activity below basal. Thus, the K72A mutation allows TRα to enhance AP-1 dependent transcription in the presence of T3, just as ERα.K206A allows ERα to activate AP-1 activity in response to estradiol, and equivalent K > A mutations allow other nuclear receptors to enhance AP-1 activity in response to their cognate ligands [10,11,17].
Figure 7.

TRα.K72 behaves similarly to ERα.K206A. (A) The K72 mutation eliminates unliganded and enhances liganded stimulation by TRα. HeLa cells were transfected as above using TRα and TRα.K72A, and then treated with 100 nM T3. (B) The K72A mutation abrogates unliganded TRα's ability to titrate a repressive function. The experiment was performed as in Fig. 6, except that cells were transfected with TRα (5 ug) and treated with 100 nM T3 instead of ER and ER ligands, respectively. (C) The K72A mutation enhances TRα's ability to inhibit transcription from classic thyroid response elements (TREs). Cells were transfected with 1 ug TRα and 2 ug TRE2-LUC reporter.
We then explored whether the K72A mutation would also affect TRα functional interactions with a corepressor complex at the PRE. Fig. 7B shows that unliganded TRα derepresess the activity of the PR-RU486 complex, but that TRα-K72A did not. Moreover, TRα.K72A showed increased ability to suppress basal transcriptional activity of a reporter gene that contains a classic thyroid response element, which binds the TR (Fig. 7C), an effect known to be mediated by recruitment of a repressive complex. Together, our results are consistent with our hypothesis that the K > A mutation abolishes the ability of nuclear receptors to functionally eliminate repressive influences on their ability to mediate transcriptional regulation.
Discussion
ER influences on physiological and pathological processes depends on regulation of genes through classic EREs and alternate response elements, such as AP-1/CRE elements, and the subsequent interplay of their gene products. As described in the Introduction, it is important to understand how ERs regulate genes with AP-1/CRE elements because this type of response element appears to activate growth factor expression (e.g. IGF I, and Cyclin D1), as well as underlie ERα effects on cell proliferation [7]. ER action at AP-1 involves two pathways: AF-dependent, active in the presence of estrogens; and AF-independent, active with "pure" anti-estrogens such as raloxifene and ICI, and exhibited prominently by ERβ or ERα truncations missing the N-terminal domain.
In this study, we examined the role of the DBD in ER action at AP-1 sites. In particular, we focused on a lysine residue at the base of the first zinc binding motif, which is highly conserved throughout the nuclear receptor family. Mutation of this residue to ala or gly enhances activity through the ER/AP-1 AF-dependent pathway but eliminates activation through the "pure" anti-estrogen pathway. This interconversion is most clearly seen in the context of either the ERα deleted of its NTD (K206A), or in the context of ERβ (K170A; Fig. 4). Bjornstrom and Sjoberg have also studied the role of the ERβ DBD in anti-estrogen activation at AP-1 and have similarly found that mutations widely spaced through the DBD eliminate anti-estrogen activation and confer some activation with estrogen. The K170 residue was not tested in that study and no superactivation was observed [18]. Their observations are consistent with our model of two exclusive mechanisms at AP-1 and the role of the DBD in controlling choice between the mechanisms.
The homologous mutation in TRα (K72A; Fig 7) exerts a highly similar phenotype to that of ERs α and β. It converts unliganded TRα stimulation at AP-1 to repression of basal transcription and converts liganded TRα from inhibiting AP-1 activity to stimulating it. Thus, the pathways affected by the ERα.K206A mutation are likely to be utilized by other members of the nuclear receptor superfamily, as well. In addition, the ERα.K206A mutation amplifies ERα action at AP-1 sites with a ligand profile that resembles that of endogenous ERα in MCF-7 and Ishikawa cells (Fig. 2). Taken together these data suggests that the K206A and G mutations amplify pathways that are likely to be physiologically relevant across nuclear receptors.
While it may seem paradoxical that mutation of a single residue in the DBD could lead to such disparate ligand effects, super-stimulation in the presence of estrogens and complete disruption of stimulation in the presence of anti-estrogens, the findings strongly corroborate our previous proposal that estrogen and anti-estrogen effects are mediated by completely distinct (and mutually exclusive) mechanisms. Consideration of these models suggest a hypothesis to explain some of the complex behaviors of our ER DBD mutants. First, we originally suggested that in the AF-dependent pathway, estrogen-liganded ERα forms part of an activation complex at the promoter, which enhances AP-1 activity (Fig. 8 AF-dependent, left). The super-activity of K206A/G suggests that the full powers of the ER activation domains at AP-1 sites are restrained by interaction with the DBD and an as yet unidentified repressor, or repressive complex. The K206A/G mutations eliminate this interaction and thus permit superstimulation (Fig 8, AF-dependent, right). Second, we have proposed that AF-independent activity elicited from the wild type ER at AP-1 results from functional repressor inactivation, be it titration, sequestration, or another process (Fig 8, AF-independent, Left). Again, introduction of the K206A mutation abrogates the interaction of ERs with an as yet unidentified repressor. Therefore, ER is no longer able to de-repress AP-1 activity. This model of the anti-estrogen pathway is consistent with the concordance of results elicited by the behavior of a wide array of point mutations at this residue at an AP-1 site (Fig. 5A) and the behavior of the same mutations in the repressor-titration assay (Fig. 6). Thus, the behavior of the K206A mutation at the AF-independent pathway can be explained by suggesting that it blocks functional inactivation of a repressor with an ER. Such a repressor, or repressive complex, need not be the same in AF-dependent and AF-independent pathways.
Figure 8.

Model of ER action at AP-1 sites. The underlying mechanisms of estrogen and anti-estrogen pathways are distinct. In the AF-dependent/Estrogen Pathway: the balance of co-activator and co-repressor functions is altered by the introduction of the K206A mutation. Loss of ER: repressor interactions lead to unopposed, potent, co-activator stimulation (right). In the AF-independent or anti-estrogen pathway, ERs titrate, or functionally inactivate, a repressive function associated with the promoter (left). The K206A mutation renders the ER unable to do so. The star represents a mutation at ERα K206: K > A/G in the AF-dependent pathway; or K > any mutation except arg, in the AF-independent pathway.
There is precedence for DBD: co-regulator interactions. Jun dimerization protein 2 binds the PR DBD [19], four proteins have been found to interact with the AR DBD [20], proteins have been found that associate with TR and RXR DBDs [21], and lastly, HET/SAFb associates with the ER DBD [22]. Perhaps some of these co-regulators, or others like them, are involved in ER action at AP-1. Whatever the co-regulator, the complex range of behaviors of ERα.K206 mutations suggests that they affect correspondingly complex functions in the DBD. We are trying to identify the interacting factors, be they repressive or not, by testing candidate factors, and screening for interacting proteins using a modified yeast two-hybrid system and biochemical techniques.
Although the inactivation of a repressive complex or function that associates with nuclear receptor DBDs is our working hypothesis, we cannot absolutely rule out that the mutation could also alter a post-translational modification required for agonist or anti-estrogen behavior at an AP-1 site. One possibility we entertained was that the K206 lysine was a target for acetylation. However, the finding that the K206R mutation leaves the anti-estrogen/AF-independent pathway intact brings acetylation as a mechanism into doubt. This conclusion is further supported by the finding that ERα K206 is not acetylated ([23], and W.L. Kraus, personal communication). To date, we have no information on other post-translational modifications that might be altered, such as phosphorylation or ubiquitination. Additionally, we do not yet know if the K > A conversion alters sub-cellular localization. We have only begun to determine the effects of the K206A mutation on localization by tagging ERα.K206A with GFP. We found no gross alteration in ER localization (data not shown). This question of altered sub-cellular localization, however, deserves further inquiry.
One wonders which physiologic or pathophysiologic processes are regulated by this highly conserved lysine. We have generated transgenic mice that express the K206A mutant under the regulation of either a keratin or a mouse mammary tumor virus promoter. Both develop a hyperproliferative response in the targeted tissues that over time correlate with tumor development (Manuscript in preparation). This is in keeping with previous findings that genes associated with growth (e.g., IGF-1) and proliferative regulation (e.g., Cyclin D1) are regulated by the ER through AP-1 or AP-1 like elements [2,4]. We are unaware of a naturally occurring germline or somatic mutations associated with squamous or breast carcinomas that involve the homologous, highly conserved lysine. However, a naturally occurring mutation in the Vitamin D Receptor (VDR) at this conserved site (K45; mutated to K45E) has been associated with a hereditary form of vitamin D resistant rickets [24]. Therefore, even though we cannot point to this mutation in a given human breast carcinoma, or growth disorder of squamous epithelium, data from our transgenic mice and the discovery of a disease associated with the homologous lysine in a different nuclear receptor (Vitamin D resistant rickets) underscore the potential importance that this residue plays in physiologic or pathophysiologic mechanisms. In fact, we hope that our work spurs a quest for mutations of this highly conserved lysine in tumors, or other pathologies.
Conclusions
Our data indicate that the ER DBD acts as a regulatory switch that dictates the ligand activation profile at AP-1 sites and thereby distinguishes the AF-dependent (estrogen) from the AF-independent (anti-estrogen) pathways. What are the factors that might bind the ER DBD and influence ER action at AP-1 sites? Data from three experiments described here corroborate our hypothesis that the factors are part of a repressive complex: 1) The K > A mutation is able to unmask the activity of a heterologous activation function (VP16; Fig. 3C), 2) Conversion of K206 to any residue except the like amino acid, arg, eliminates the AF-independent pathway (Fig. 5A), and 3) the profile of activity elicited by the same mutations parallels their behavior in the repressor titration assay (Fig. 6).
Regardless of the actual mechanism, the degree of super-stimulation elicited by ERα.K206A is unlikely to be benign in normal growth and development. In fact, data from our ERα.K206A transgenic mice bear this out (Manuscript in preparation). The extraordinary degree of ER-stimulated AP-1 activity possible likely needs to be restrained over a wide range of cellular conditions, though it may be selectively exposed in some. Given the potential importance of such mechanisms, it is likely that the proteins that mediate these effects will represent valuable targets for drugs designed specifically to dampen ER stimulated growth. We are presently searching for proteins that associate with the ER DBD in a manner influenced by mutations of this highly conserved lysine, in the hopes that they will direct novel drug development.
Methods
Cell lines and ER ligands
HeLa, Ishikawa, MCF-7, and MDA-MB-453 cells were grown in Dulbecco's Modified Eagle's/F-12 Coon's modified medium (Sigma), with 15 mM Hepes, L-glutamine (0.438 g/liter), NaHCO3 (1.38 g/liter), 10% iron supplemented calf serum (Sigma) which we screen for negligible E2 activity, and penicillin/streptomycin as previously described [5,8]. E2 was purchased from Sigma (St. Louis, Mo). ICI 182,780 was a gift from Dr. Alan Wakeling (Astra/Zeneca, Macclesfieled, UK). Ral was a gift from Dr. Stefan Nilsson (Karo Bio AB Huddinge, Sweden).
Plasmids
The following expression vectors were previously described: ERα: G400, the fully wild type and G400V which contains the indicated G > V conversion in the LBD of ERα. These vectors have previously been termed HEG0 and HE0 respectively [12,25]; ERα.K206A [2], the ERβ 530 amino acid isoform [8], and PR [26].
ERαG400V.K206A was generated by D. Barry Starr, using site directed mutagenesis [27]. ERα mAF-2.K206A was prepared by ligating a BglII fragment from a mutated version of pSG5-ERαLBD [28] into BglII digested ERαK206A. ERα.K206 was mutated to ala in the context of ERαG400V and ERα DBD-LBD mutations by site directed mutagenesis of HE0 using the Stratagene Quickchange kit. ERβ.K170A was made similarly. VP16-ER-DBD-LBD and the K206A equivalent (both in ERα) were generated by PCR amplification of ERα amino acids 164–595 using ERα and ERαK206A as templates. The primers were designed with EcoRI and SalI sites. The amplified cDNA sequences were digested with these enzymes and ligated downstream (in frame) of a cDNA encoding the strong HSV VP16 activation domain in expression vector pVP16 (Clontech). Primer sequences and PCR conditions are available on request.
Reporter genes Coll73-Luc [1], ERE-Luc [5], and GAL4-RE5-Luc (GK1) [29] have been previously described. The PR (and GR) responsive reporter gene TAT3-Luc (3xPRE) was a gift from K. Yamamoto (UCSF) and is described in [30].
Transfections
Transfections were performed as described [2,5,8]. Cells were grown to an approximate density of 5 × 104 cells/cm2 on 10cm diameter tissue culture plates. The cells were trypsinized and resuspended in a single 0.4-cm gap electroporation cuvette in 0.5 ml electroporation buffer (PBS supplemented with 10% glucose and 10 μg/ml BioBrene detergent; Applied Biosystems, Foster City, CA). Transfections to determine ER affects on AP-1 activity contained 2 μg of Coll73-Luc and actin β-galactosidase reporters and 5 μg ER vector or empty vector controls. Transfections to determine ER effects on classical response elements contained 2 μg of the appropriate reporter, an actin-β-galactosidase control, and 1 μg ER expression vector. Transfections to determine ER affects on PR-RU486 complex activity contained 5 μg of TAT3-Luc reporter, 2 μg of CMV-β-galactosidase and 5 μg each of PR and ER expression vectors. Transfected cells were resuspended in standard growth medium and plated on 12-well dishes. Ligands were then added at the following final concentrations (ICI 10-7 M, Ral 10-7 M, Tam 5 × 10-6 M, E2 10-7 M, RU486 10-6 M).
Western Blots
10 cm dishes of HeLa cells (transfected with 10 μg ER expression vector and 2 μg CMV-β-galactosidase to permit detection of equivalent transfection efficiency) were treated with ligands for 24 hrs, chilled on ice, washed with ice-cold PBS, then incubated in lysis buffer (0.2% Triton X-100, 150 mM NaCl, 50 mM Tris pH 7.8, 1 mM EDTA, and 1/1000 dilution Novagen protease inhibitor cocktail) for 10 mins. They were then scraped and centrifuged at 4°C in an Eppendorf bench top centrifuge at maximum speed for 15 mins. Protein concentration in cell extract was determined by standard methods. The concentration of each sample was normalized by addition of cold lysis buffer to a final value of approximately 5 mg total protein per ml. 20–30 μg of proteins were separated by SDS-PAGE. Proteins were transferred to a wet Immuno-Blot PVDF membrane (Bio-Rad, Hercules CA), overnight at 90 mA, 30 V, using a standard transfer apparatus. The membrane was then incubated in 5% non-fat milk in PBS-T (1 × PBS, 0.1% Tween-20) for 1 hour and washed twice in PBS-T for 10 minutes at room temperature. The primary ERα antibody used in this study was HC-20 (Santa Cruz Biotechnology, Inc.) directed against the ERα C-terminus. Primary antibody was diluted 1:2000 in PBS-T and incubated with the membrane for 1 hour, followed by PBS-T washes, 1 × 15 minutes and then 2 × 5 minutes. The membrane was incubated for 45 minutes with horse-radish peroxidase conjugated anti-rabbit IgG (Santa Cruz Biotechnology, Inc.) diluted at 1:2000 in PBS-T, followed by PBS-T washes (1 × 15 minutes and 4 × 5 minutes). After the last wash, the membrane was developed with a standard ECL kit (Amersham-Pharmacia Biotech), covered with Saran wrap and exposed to X-ray film.
Abbreviations
AF-1 Activation Function 1
AF-2 Activation Function 2
AP-1 Activator Protein-1
CEF Chicken Embryo Fibroblasts
CRE cAMP Response Element
DBD DNA binding Domain
ERα Estrogen Receptor-alpha, fully wild type: G400
ERαG400V Estrogen Receptor-alpha with LBD mutation: G400V
ERβ Estrogen Receptor-beta
ERE Estrogen Response Element
GR Glucocorticoid Receptor
ICI Imperial Chemical Industries #182,780
LBD Ligand Binding Domain
N-CoR Nuclear receptor Corepressor
NTD Amino (N)-terminal domain
PR Progesterone Receptor
PRE Progesterone Response Element
Ral Raloxifene (Evista)
SMRT Silencing mediator of retinoid and thyroid receptors
Tam Tamoxifen (Nolvadex)
Competing interests
None declared for any authors except PK who is a consultant, former member of the board of directors, and has significant financial interests in KaroBio AB, a Swedish company that develops pharmaceuticals that target the ER and other nuclear receptors.
Authors' contributions
RU Made the original observation that ERα.K206A superstimulated the ER/AP-1 pathway, performed experiments while a post-doctoral fellow in PJK's lab, and finished the project in her own lab at the University of Virginia, which included writing the manuscript.
PW Provided considerable intellectual guidance through out the project. He provided significant input in crafting the manuscript.
RP Provided intellectual insight and designed and performed several experiements, in particular the VP16 fusion experiment (Fig. 3C).
PN Performed many experiments under the guidance of PW
CV & HF Performed many experiments under the guidance of PW and PJK.
PK Oversaw the entire project and provided the majority of funding.
Acknowledgments
Acknowledgements
Some of these data were initially presented at the 1998 & 2000 Nuclear Receptor Keystone meetings, and the 2001 Cold Spring Harbor meeting. We also thank Carol M. Anderson and Kristen Hilty for their excellent technical support in the early part of the project. This work was supported by K08 DK 02335 and R03 DK56236 (R.M.U.) and DK51083 and CA80210 to (P.J.K.).
Contributor Information
Rosalie M Uht, Email: ruht@virginia.edu.
Paul Webb, Email: webbp@itsa.ucsf.edu.
Phuong Nguyen, Email: pnguyen@diabetes.ucsf.edu.
Richard H Price Jr, Jr, Email: r.price@amgen.com.
Cathleen Valentine, Email: valentin@medicine.ucsf.edu.
Helene Favre, Email: favrehe@itsa.ucsf.edu.
Peter J Kushner, Email: kushner@itsa.ucsf.edu.
References
- Webb Paul, Lopez Gabriela N., Uht Rosalie M., Kushner Peter J. Tamoxifen activation of the estrogen receptor/AP-1 pathway: potential origin for the cell-specific estrogen-like effects of antiestrogens. Mol Endocrinol. 1995;9:443–456. doi: 10.1210/me.9.4.443. [DOI] [PubMed] [Google Scholar]
- Liu MM, Albanese C, Anderson CM, Hilty K, Webb P, Uht RM, Price R. H., Jr., Pestell RG, Kushner PJ. Opposing action of estrogen receptors alpha and beta on cyclin D1 gene expression. J Biol Chem. 2002;277:24353–24360. doi: 10.1074/jbc.M201829200. [DOI] [PubMed] [Google Scholar]
- Kushner PJ, Agard DA, Greene GL, Scanlan TS, Shiau AK, Uht RM, Webb P. Estrogen receptor pathways to AP-1. Steroid Biochemistry & Molecular Biology. 2000;74:311–317. doi: 10.1016/S0960-0760(00)00108-4. [DOI] [PubMed] [Google Scholar]
- Umayahara Y, Kawamori R, Watada H, Imano E, Iwama N, Morishima T, Yamasaki Y, Kajimoto Y, Kamada T. Estrogen regulation of the insulin-like growth factor I gene transcription involves an AP-1 enhancer. J Biol Chem. 1994;269:16433–16442. [PubMed] [Google Scholar]
- Paech K, Webb P, Kuiper GG, Nilsson S, Gustafsson J, Kushner PJ, Scanlan TS. Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sites [see comments] Science. 1997;277:1508–1510. doi: 10.1126/science.277.5331.1508. [DOI] [PubMed] [Google Scholar]
- Philips Alexandre, Teyssier Catherine, Galtier Florence, Rivier-Covas Corinne, Rey Jean-Marc, Rochefort Henri, Chalbos Dany. FRA-1 Expression Level Modulates Regulation of Activator protein-1 Activity by Estradiol in Breast Cancer Cells. Mol Endocrinol. 1998;12:973–985. doi: 10.1210/me.12.7.973. [DOI] [PubMed] [Google Scholar]
- Shang Y, Brown M. Molecular determinants for the tissue specificity of SERMs. [see comments.] Science. 2002;295:2465–2468. doi: 10.1126/science.1068537. [DOI] [PubMed] [Google Scholar]
- Webb P, Nguyen P, Valentine C, Lopez GN, Kwok GR, McInerney E, Katzenellenbogen BS, Enmark E, Gustafsson J-Å, Nilsson S, Kushner PJ. The Estrogen Receptor Enhances AP-1 Activity by two Distinct Mechanisms with Different Requirements for Receptor Transactivation Functions. Mol Endocrinol. 1999;13:1672–1685. doi: 10.1210/me.13.10.1672. [DOI] [PubMed] [Google Scholar]
- Webb P, Nguyen P, Kushner PJ. Differential SERM effects on corepressor binding dictate ERalpha activity in vivo. J Biol Chem. 2003;278:6912–6920. doi: 10.1074/jbc.M208501200. [DOI] [PubMed] [Google Scholar]
- Yang-Yen Hsin-Fang, Chambard Jean-Claude, Sun Yu-Lin, Smeal Tod, Schmidt Thomas J., Drouin Jacques, Karin Michael. Transcriptional interference between c-Jun and the glucocorticoid receptor: mutual inhibition of DNA binding due to direct protein-protein interaction. Cell. 1990;62:1205–1215. doi: 10.1016/0092-8674(90)90396-V. [DOI] [PubMed] [Google Scholar]
- Starr D.Barry, Matsui William, R. Thomas Jay, Yamamoto Keith R. Intracellular receptors use a common mechanism to interpret signaling information at response elements. Genes & Dev. 1996;10:1271–1283. doi: 10.1101/gad.10.10.1271. [DOI] [PubMed] [Google Scholar]
- Tora L, Mullick A, Metzger D, Ponglikitmongkol M, Park I, Chambon P. The cloned human oestrogen receptor contains a mutation which alters its hormone binding properties. EMBO J. 1989;8:1981–1986. doi: 10.1002/j.1460-2075.1989.tb03604.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danielian PS, White R, Lees JA, Parker MG. Identification of a conserved region required for hormone dependent transcriptional activation by steroid hormone receptors. EMBO J. 1992;11:1025–1033. doi: 10.1002/j.1460-2075.1992.tb05141.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb P, Valentine C, Nguyen P, Price R. H., Jr., Marimuthu A, West BL, Baxter JD, Kushner PJ. ERbeta Binds N-CoR in the Presence of Estrogens via an LXXLL-like Motif in the N-CoR C-terminus. Nucl Recept. 2003;1:4. doi: 10.1186/1478-1336-1-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Xun, Jeyakumar M, Petukhov Sergei, Bagchi Milan K. A Nuclear Receptor Corepressor Modulates Transcriptional Activity of Antagonist-Occupied Steroid Hormone Receptor. Mol Endocrinol. 1998;12:513–524. doi: 10.1210/me.12.4.513. [DOI] [PubMed] [Google Scholar]
- Lopez G, Schaufele F, Webb P, Holloway JM, Baxter JD, Kushner PJ. Positive and negative modulation of Jun action by thyroid hormone receptor at a unique AP1 site. Mol Cell Biol. 1993;13:3042–3049. doi: 10.1128/mcb.13.5.3042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaaskelainen Tiina, Pirskanen Asta, Ryhanen Sanna, Palvimo Jorma J., Deluca Hector F., Maenpaa Pekka. Functional interference between AP-1 and the vitamin D receptor on osteocalcin gene expression in human osteosarcoma cells. Eur J Biochem. 1994;224:11–20. doi: 10.1111/j.1432-1033.1994.tb19989.x. [DOI] [PubMed] [Google Scholar]
- Bjornstrom L, Sjoberg M. Mutations in the estrogen receptor DNA-binding domain discriminate between the classical mechanism of action and cross-talk with Stat5b and activating protein 1 (AP-1) J Biol Chem. 2002;277:48479–48483. doi: 10.1074/jbc.C200570200. [DOI] [PubMed] [Google Scholar]
- Wardell SE, Boonyaratanakornkit V, Adelman JS, Aronheim A, Edwards DP. Jun dimerization protein 2 functions as a progesterone receptor N-terminal domain coactivator. Mol Cell Biol. 2002;22:5451–5466. doi: 10.1128/MCB.22.15.5451-5466.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janne OA, Moilanen AM, Poukka H, Rouleau N, Karvonen U, Kotaja N, Hakli M, Palvimo JJ. Androgen-receptor-interacting nuclear proteins. Biochem Soc Trans. 2000;28:401–405. [PubMed] [Google Scholar]
- Mathur Mukul, Tucker Philip W., Samuels Herbert H. PSF is a Novel Corepressor that Mediates its Effect through Sin3A and the DNA Binding Domain of Nuclear Hormone Receptors. Mol Cell Biol. 2001;21:2298–2311. doi: 10.1128/MCB.21.7.2298-2311.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oesterreich S, Zhang QP, Hopp T, Fuqua SAW, Michaelis M, Zhao HH, Davie JR, Osborne CK, Lee AV. Tamoxifen-bound estrogen receptor (ER) strongly interacts with the nuclear matrix protein HET/SAF-B, a novel inhibitor of ER-mediated transactivation. Mol Endocrinol. 2000;14:369–381. doi: 10.1210/me.14.3.369. [DOI] [PubMed] [Google Scholar]
- Wang C, Fu M, Angeletti RH, Siconolfi-Baez L, Reutens AT, Albanese C, Lisanti MP, Katzenellenbogen BS, Kato S, Hopp T, Fuqua SA, Lopez GN, Kushner PJ, Pestell RG. Direct acetylation of the estrogen receptor alpha hinge region by p300 regulates transactivation and hormone sensitivity. J Biol Chem. 2001;276:18375–18383. doi: 10.1074/jbc.M100800200. [DOI] [PubMed] [Google Scholar]
- Kahlen J-P, Carlberg C. Allosteric interaction of the 1a,25-dihydroxyvitamin D3 receptor and the retinoid X receptor on DNA. NAR. 1997;25:4307–4313. doi: 10.1093/nar/25.21.4307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green Stephen, Walter Philippe, Kumar Vijay, Krust Andree, Bornert Jean-Marc, Argos Patrick, Chambon Pierre. Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature. 1986;320:134–139. doi: 10.1038/320134a0. [DOI] [PubMed] [Google Scholar]
- Uht Rosalie M., Anderson Carol M., Webb Paul, Kushner Peter J. Transcriptional Activities of Estrogen and Glucocorticoid Receptors Are Functionally Integtrated at the AP-1 Response Element. Endocrinology. 1997;138:2900–2908. doi: 10.1210/en.138.7.2900. [DOI] [PubMed] [Google Scholar]
- Kunkel TA. Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc Natt Acad Sci, U S A. 1985;82:488–492. doi: 10.1073/pnas.82.2.488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez GN, Webb P, Shinsako JH, Baxter JD, Greene GL, Kushner PJ. Titration by estrogen receptor activation function-2 of targets that are downstream from coactivators. Mol Endocrinol. 1999;13:897–909. doi: 10.1210/me.13.6.897. [DOI] [PubMed] [Google Scholar]
- Webb P, Nguyen P, Shinsako J, Anderson C, Feng W, Nguyen MP, Chen D, Huang SM, Subramanian S, McKinerney E, Katzenellenbogen BS, Stallcup MR, Kushner PJ. Estrogen receptor activation function 1 works by binding p160 coactivator proteins. Mol Endocrinol. 1998;12:1605–1618. doi: 10.1210/me.12.10.1605. [DOI] [PubMed] [Google Scholar]
- Iniguez-Lluhi JA, Pearce D. A common motif within the negative regulatory regions of multiple factors inhibits their transcriptional synergy. Mol Cell Biol. 2000;20:6040–6050. doi: 10.1128/MCB.20.16.6040-6050.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]