

Abstract
Repetitive transcranial magnetic stimulation (rTMS) is a noninvasive tool used to manipulate activity in specific neural circuits of the human brain. Clinical studies suggest that, in some patients with major depression, rTMS has the potential to alleviate symptoms that may be related to functional abnormalities in a frontocingulate circuit. This paper reviews the rationale for the use of rTMS in this context. The following topics are discussed: symptoms and cognition in major depression, with special emphasis on the initiation of speech; neuroimaging studies of depression; rTMS as treatment for depression; structure and function of the mid-dorsolateral frontal and anterior cingulate cortices; and combined TMS/positron emission tomography studies of frontocortical connectivity.
Medical subject headings: brain; cognition; depression; electroconvulsive therapy; frontal lobe; speech; tomography, emission computed; transcranial magnetic stimulation
Abstract
La stimulation magnétique transcrânienne répétitive (SMTr) est un outil non effractif utilisé pour manipuler l'activité de circuits neuraux précis du cerveau humain. Les études cliniques indiquent que chez certains patients aux prises avec une dépression majeure, la SMTr peut atténuer les symptômes pouvant être reliés à des anomalies fonctionnelles dans un circuit frontocingulaire. Cette communication passe en revue la justification de l'utilisation de la SMTr dans ce contexte. On aborde les sujets suivants : symptômes et cognition en cas de dépression majeure, avec accent sur l'initiation de la parole; étude de la dépression par neuro-imagerie; SMTr comme traitement de la dépression; structure et fonction des cortex cingulaires frontal et antérieur mésodorsolatéral; et étude de la connectivité frontocorticale par SMT et tomographie par émission de positrons combinées.
Introduction
Major depression is a common disorder with a lifetime prevalence in the general population of about 13% in men and 21% in women.1 Despite continuing advances in the development of antidepressant drugs, the condition of about 30% of patients remains refractory to drug treatment2 and may require electroconvulsive therapy (ECT). Although ECT as currently practised is a relatively safe procedure, it nevertheless requires general anesthesia, muscular relaxation and induction of a seizure, and it involves side effects such as memory disturbances.3 Over the past 5 years, repetitive transcranial magnetic stimulation (rTMS) of the mid-dorsolateral frontal cortex (MDLFC, also described as the dorsolateral prefrontal cortex [PFC]) has been established as a less invasive alternative to ECT. Its effectiveness may be related to the functional abnormalities in the left mid-dorsolateral and anterior cingulate cortices often observed in patients with depression. The relevant mechanisms of action are, however, still unknown. At least 2 possibilities exist. First, rTMS may modulate activity in the specific neural circuits (e.g., frontocingulate system) that mediate a given group of symptoms. Second, rTMS effects may be the result of a facilitation of monoaminergic neurotransmission. This review focuses on neuroimaging studies that address these 2 neural mechanisms that may underlie TMS-induced changes in depression-like symptoms, with a particular emphasis on the role of the anterior cingulate cortex (ACC) in the initiation of speech. The following 5 topics are reviewed:
Symptoms and cognition in major depression
Neuroimaging studies of depression
rTMS as treatment for depression
Structure and function of the MDLFC and the ACC
Combined TMS/positron emission tomography (PET) studies of frontocortical connectivity
Symptoms and cognition in major depression
The defining symptom of major depression is that of depressed mood; patients appear sad and often speak of feeling “low” or “down.” The patient's everyday behaviour is consistent with a state of decreased drive and motivation; patients eat less and do not engage in, or enjoy to a lesser extent (hence anhedonia), social activities, such as sports, seeing friends and going out. The cognitive performance of depressed patients is impaired in many domains, including learning, memory and attention, as well as executive, motor and perceptual functions.4,5 In terms of motor function, the speech of depressed patients is often slow, quiet and monotonous and infrequently initiated.5 Analyses of speech acoustics have revealed the presence of quantitative and qualitative differences between the speech of depressed patients and control subjects, including speech pitch, loudness and rate.6,7,8,9,10,11,12 The wide range of deficits observed in depression is compatible with disease- related changes in motivation and executive processes and, by implication, with abnormalities in the neural systems mediating these functions.
A number of neuropsychological studies have documented poor performance of patients with major depression on tasks that test executive functions. For example, Purcell et al13 administered the Cambridge Neuropsychological Test Automated Battery (CANTAB) to 20 patients (18–52 yr) with unipolar depression and found significant impairment on intradimensional and extradimensional set shifting and on “subsequent movement times” in the Tower of London task, the latter suggesting “that motor slowness in depression occurs when sustained motor responses to changing stimuli are required.”13 Using the Stroop task, several authors found increased sensitivity to interference in patients with major depression,14,15,16although Austin et al17 reported negative findings. Furthermore, several studies found that depressed patients were slower than control subjects when simply reading the names of colours15,18 and displayed reduced verbal fluency,18,19,20 which may suggest impairment in the initiation of retrieval and/or utterance of words.
On the basis of the findings described here, patients with major depression appear to suffer from a deficit in 2 aspects of volition, namely, the ability to resist interference and the ability to initiate actions.
Neuroimaging studies of depression
Early influential studies that examined resting cerebral glucose metabolism and blood flow with PET in patients with depression found hypometabolism and hypoperfusion localized to the left MDLFC (Fig. 1).21,22,23,24,25,26,27,28,29,30,31 The discovery that decreased neural activity in the MDLFC was correlated with severity of depression or was reversed upon recovery from depression22,25,26 was believed to support a critical role for this region in the pathophysiology of depression. These initial imaging findings were confirmed by later PET studies32,33,34,35 and extended by electroencephalography studies that demonstrated increased alpha power (alpha power is thought to be inversely related to neural activity) in left frontal regions of the brains of depressed patients.36,37,38,39
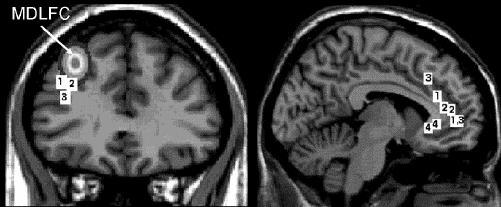
Fig. 1: Comparison of brain locations (in standardized stereotaxic space) identified as hypometabolic or hypoperfused in previous imaging studies of depression (labelled 2,27 323 and 428) with those targeted in our transcranial magnetic stimulation (TMS)/positron emission tomography (PET) studies (labelled 129,30,31). The location of the mid-dorsolateral frontal cortex (MDLFC) is also indicated; in repetitive TMS (rTMS) studies of depression, this location is typically defined as a region located 5 cm anterior to the primary motor cortex (M1). Here, we established the “average” location of the MDLFC as follows: magnetic resonance images (MRIs) obtained in 152 healthy subjects were used to label the brain locations located 5 cm in front of the left M1 in each subject and to transform these “MDLFC” labels from native to standardized space. In the case of the anterior cingulate cortex (ACC), projections of brain locations onto a single sagittal slice were used.
The apparent depression-related left frontal hypometabolism or hypoperfusion was initially extended to include the ACC, with studies of cerebral blood flow and metabolism also demonstrating reduced activity in both supracallosal (areas 24á, 24b́, 24ć and 32́ as defined by Vogt et al40), perigenual (areas 24a, 24b, 24c and 32) and subcallosal (area 25) regions of the ACC (Fig. 1).23,25,28,41,42 As was the case for the lateral PFC, blood flow and metabolic abnormalities in ventral perigenual regions of the ACC (approximately perigenual area 24 below z = 0) were thought to be related to the reduction in grey-matter volume observed in this region in depressed patients.28,42 Unlike in the PFC, however, after correcting for the partial volume effect of the reduced grey matter using a computer simulation, it was later concluded that metabolic activity in the ventral perigenual ACC was actually increased in depressed patients relative to control subjects.43 The finding of depression-related increases in neural activity in the perigenual ACC has since been replicated35 and, importantly, much research has demonstrated that neural activity in this region decreases in response to antidepressant treatment and is positively correlated with severity of depression,43,44,45,46 but see Mayberg et al for opposite findings.33 Although less commonly found, increased glucose metabolism has been observed in the subcallosal ACC of depressed patients,32,47 and recovery from depression has been associated with metabolic decreases in this region.33,48
Whereas early studies also cited decreased neural activity in dorsal regions of the ACC during depression,23,25 more recent work has not confirmed these findings. Instead, research has found that neural activity in the supracallosal and dorsal perigenual ACC is also increased in depression35,49,50 and that hyperperfusion and hypermetabolism in these regions predict antidepressant response.27,47,51,52 Note, however, that opposite findings were reported by Brody et al53 and by Ketter et al.54
Altogether, these studies highlight 2 important issues:
the involvement of the MDLFC and ACC in depression; and
the reversibility of depression-related “abnormalities” in the MDLFC and ACC with successful treatment.
rTMS as treatment for depression
TMS is a noninvasive technique that allows investigators to stimulate specific cortical regions of the human brain through the skull and, in turn, induce short-term (milliseconds) and long-term (minutes to hours) changes in behaviour.55 The duration of the effect depends on the stimulation mode, and it is well characterized in the case of single-pulse and paired-pulse stimulation of the primary motor cortex.56 When applied in trains of pulses, namely, as repetitive TMS or rTMS, stimulation typically interferes with specific cognitive operations presumably executed by the stimulated cortical region during the stimulation train. But several studies have now demonstrated that rTMS may also facilitate, or modulate, behaviour well beyond the actual stimulation.57,58 The most common instance of such long-lasting effects is that of rTMS-induced changes in mood observed in both healthy and depressed subjects after the stimulation of the frontal cortex. In the early 1990s, several investigators initiated studies of rTMS-induced changes of mood in both healthy volunteers and in patients with major depression. In these studies, rTMS was applied at different locations (left and right MDLFC and the vertex), with different stimulation parameters (0.3–20 Hz, 1-s to 10-s trains, 20 trains/session) and for a variable total amount of stimulation (up to 20 daily sessions). In most depression studies, rTMS is applied over the left MDLFC. To target this region, the coil is typically positioned 5 cm in front of the scalp location from which single-pulse TMS elicits muscle twitches in the contralateral hand (i.e., M1). As can be seen in Figure 1, the average location (in Talairach space) of the MDLFC defined in this way is about 10–15 mm dorsal to the hypometabolic regions revealed in the neuroimaging studies reviewed in the previous section (Dr. Irena Rektorova, Masaryk University, Brno, Czech Republic, and Dr. Tomáš Paus: unpublished data, 1998).
In healthy subjects, George et al59 and Pascual-Leone et al60 described significant increases in sadness following rTMS applied over the left MDLFC; no changes were found when stimulating over the midline (5 cm anterior to the leg M1). In the study by Pascual-Leone et al,60 stimulation was delivered with a Cadwell High Speed Magnetic Stimulator (Cadwell Laboratories, Inc., Kennewick, Wash.) and a large water-cooled figure-eight coil using the following parameters: 110% motor threshold, 10 Hz, 5-s trains, 10 trains every 25 s for a total of 500 pulses at each site.
Since the first case report of potential treatment effects of rTMS on depression,61 a number of studies have sought to establish that rTMS indeed alleviates the symptoms of depression.62,63,64 Pascual-Leone et al65 and George et al66 carried out crossover placebo-controlled rTMS studies in patients with major depression and observed significant decreases in scores on the Hamilton Depression Rating Scale (HDRS); these decreases were observed only when the left MDLFC was stimulated. Similar findings were reported also by others in placebo-controlled67 and open-label68,69,70 studies. Grunhaus et al69 randomly assigned 40 patients with drug-refractory major depression (with or without psychosis) to either ECT (n = 20) or rTMS (n = 20) groups; ECT included 7–14 sessions, and rTMS involved 20 sessions of left MDLFC stimulation (10 Hz, 2-s or 6-s trains, 20 trains/session, 90% motor threshold). At the end of the treatment, both groups showed a statistically significant decrease in HDRS scores (ECT: 61%, 16/20 responders; rTMS: 40%, 9/20 responders). The most important finding of this study was that of a differential treatment response of patients with psychosis and those without psychosis. The patients without psychosis benefited equally from the ECT and rTMS treatments, whereas the patients with psychosis showed only small rTMS effects (patients without psychosis: ECT: 45%, 6/10 responders; rTMS: 53%, 7/11 responders; patients with psychosis: ECT: 73%, 10/10 responders; rTMS: 28%, 2/9 responders).
The number of patients studied in published reports is relatively small. In order to pool all the available data, Avery et al have compiled a database of rTMS trials carried out in over 20 centres.71 As of April 2003, a total of 523 patients with drug-refractory depression had received rTMS treatment. In most studies (88%), the left MDLFC was the most effective site of stimulation, high-frequency stimulation (> 1 Hz) was the most common (98%) mode of rTMS, and 10 daily sessions was the most frequent number of treatments. In these studies, the mean before-versus-after decrease in the HDRS scores was 34% (range 15%–62%). In 2 studies (n = 85), a low-frequency (1 Hz) rTMS was applied over the right MDLFC with a resulting 41% mean decrease in the HDRS scores. These data are consistent with published reports and confirm the efficacy of rTMS treatment of depression.
Structure and function of the MDLFC and the ACC
As pointed out earlier, the left MDLFC and the ACC have been implicated in the pathophysiology of depression. Furthermore, the left MDLFC is the most common target for rTMS treatment of depression and, based on our TMS/PET studies,29,72 rTMS of the left MDLFC modulates the blood-flow response in the ACC. For these reasons, we shall provide a brief overview of the corticocortical connectivity and functions of these 2 cortical regions.
Major subdivisions of the human prefrontal cortex have been recently re-examined by Petrides and Pandya (Fig. 2.73,74 The MDLFC occupies the middle frontal and superior frontal gyri and comprises cytoarchitectonic areas 46 and 9/46 (middle frontal gyrus) and area 9 (superior frontal gyrus). Major corticocortical connections of the MDLFC are bidirectional and link this region with posterior neocortical areas that are involved in the processing of visual (prestriate and inferior temporal cortices), auditory (superior temporal cortex) and somatosensory (parietal cortex) information.73 Importantly, the MDLFC also possesses reciprocal connections with the anterior and, to a lesser extent, posterior cingulate cortices.75,76,77 Patients with unilateral lesions involving the lateral frontal cortex show impairment on tasks that typically require the subject to organize and plan sequences of responses, select appropriate strategies and monitor self-generated actions.78,79,80 In the search for a critical contribution of the MDLFC to such a host of executive processes, Petrides and collaborators carried out a series of lesion and imaging studies in human and nonhuman primates.78 In these studies, they used different variations of self-ordered and externally ordered working-memory tasks and concluded that the MDLFC “is a specialized region for the on-line monitoring and manipulation of cognitive representations within working memory” (p. 167).78
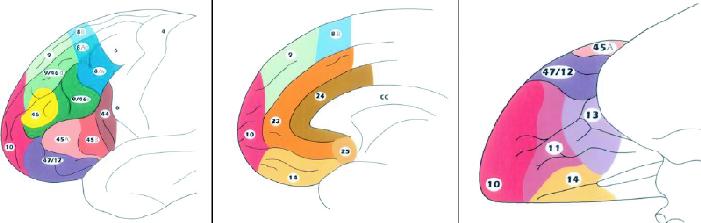
Fig. 2: Schematic diagrams of the lateral (left), medial (middle) and inferior (right) surfaces of the human frontal lobe to illustrate its cytoarchitectonic parcellation. Reproduced with permission from Elsevier.74
The human ACC also represents a structurally and functionally heterogeneous region.81 At a gross morphological level, we have subdivided the ACC region into the caudal and rostral, supracallosal and subcallosal, and limbic and paralimbic subdivisions (Fig. 3).81,82,83,84,85,86 Several cytoarchitecturally distinct regions have been identified within area 24, including limbic areas 24a and 24b and paralimbic area 24c in humans40,74,87 and monkeys.74,75,88,89,90 Different cortical zones can also be distinguished along the rostral–caudal plane of the human ACC, resulting in further subdivisions of areas 24 and 32 as they curve around the genu of the corpus callosum.40,86 Cytoarchitectonic area 25 occupies the most posterior portion of the subcallosal ACC and borders orbitofrontal area 12. At a functional level, a distinction has been made between the supracallosal or “cognitive” and the subcallosal or “visceral” parts of the ACC.27,40 In the monkey, these regions show somewhat distinct patterns of connectivity. Areas 24 and 32 are predominantly supracallosal and are interconnected with the MDLFC.88,91 Subcallosal area 25 is interconnected with the posterior orbitofrontal area 13,88 hypothalamus,92 ventral striatum93 and periaqueductal grey,94 and has been implicated in the control of respiration, blood pressure and other autonomic functions.95,96
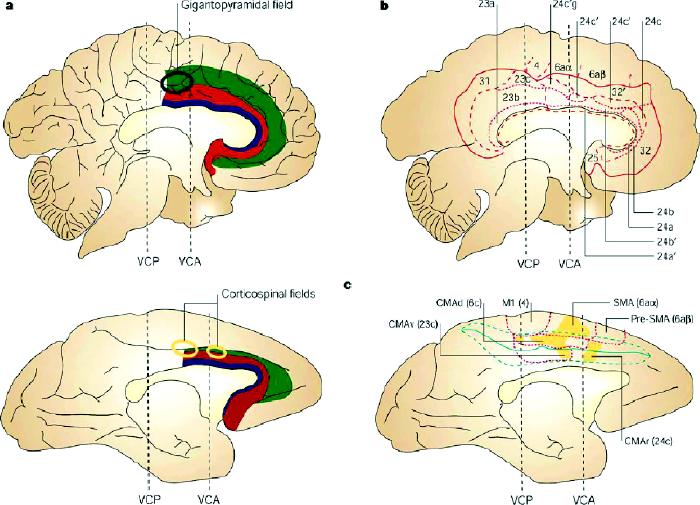
Fig. 3: Cytoarchitectonic subdivisions of human and monkey cingulate cortex.
A: Along a rostrocaudal axis, the cingulate cortex can be divided into (1) a posterior region (areas 23, 26, 29, 30, 31) characterized mostly by a granular type of cortex and (2) an anterior agranular region (areas 24, 25, 32, 33). The anterior cingulate cortex is an agranular type of cortex (i.e., layer IV is absent) with a prominent and deeply stained layer V. Area 32 has an incipient granular layer IV. A ventrodorsal distinction, based on the degree of laminar differentiation, sets apart the old periallocortical areas adjacent to the corpus callosum (area 33) from the proisocortical region (areas 24, 25) and the paralimbic region on the upper bank of the cingulate sulcus and in the paracingulate gyrus (area 32). In addition to these main rostrocaudal and ventrodorsal distinctions, subtle variations in cytoarchitecture define further subdivisions of area 32, often reflecting structural features of the adjacent neocortical areas.86 The approximate position of the corticospinal fields is indicated relative to the vertical plane passing through the anterior commissure (VCA). VCP = vertical plane passing through the posterior commissure.
B: Cytoarchitectonic areas superimposed on the flat map of the medial wall of the human brain. The bold lines outline the cingulate region, the thinner dashed lines show the borders between the cingulate areas (e.g., between areas 24 and 23), and the dotted lines indicate the borders between subdivisions of each area (e.g., between areas 24b and 24c).
C: Location of the motor areas on the medial wall of the monkey brain. The dotted lines show the boundaries of the cytoarchitectonic areas. Shaded areas correspond to the territory of origin of corticospinal projections to cervical and upper thoracic segments. M1 = primary motor cortex, SMA = supplementary motor area, CMAr = rostral cingulate motor area, CMAd = caudal cingulate motor area, dorsal bank, CMAv = caudal cingulate motor area, ventral bank. Reproduced with permission from Macmillan Magazines Ltd. (www.nature.com/reviews) (Nat Rev Neurosci 2001;2:417-24).81
In the context of depression, the role of the supracallosal ACC in volitional control of behaviour, in general, and speech and vocalization, in particular, are emphasized. In the monkey, lesions to the most rostral part of the ACC were shown to reduce condition-specific vocal output.97,98,99 In humans, bilateral cingulate lesions initially give rise to akinetic mutism100,101 and, on recovery, speech output characterized by monotonous intonation and a low frequency of spontaneous utterances.102 In our neuroimaging studies of the ACC,103 the role of the ACC in effortful as compared with automatic control of speech has been established. In a number of studies, a robust “activation” of the human ACC was observed during the performance of Stroop tasks with verbal responses.104,105,106 More recently, we have shown that neural activity in the ACC was correlated with paralinguistic characteristics of speech production during sad affect.107
Combined TMS/PET studies of frontocortical connectivity
As reviewed earlier, changes in mood observed in both healthy and depressed subjects following the stimulation of the frontal cortex are the most common instances of long-lasting effects induced by repetitive TMS. The neural mechanisms of these effects are unknown but, as described earlier, at least 2 possibilities exist:
rTMS may modulate activity in the specific neural circuits (e.g., frontocingulate system) that mediate a given group of symptoms; and
rTMS effects may be the result of a facilitation of monoaminergic neurotransmission.
We can assess both potential mechanisms by combining TMS with PET.
The first type of study uses 15O-H2O to measure regional distribution of cerebral blood flow (CBF) as an index of the net amount of excitatory postsynaptic neurotransmission.55 One such TMS/PET experiment, carried out in our laboratory, was aimed at studying corticocortical connectivity of the MDLFC and its modulation by brief periods of rTMS (Fig. 4A).29,108 In this study, the target region was chosen based on a probabilistic location of the left MDLFC (x = –40, y = 32, z = 30), as revealed by a previous PET study of verbal working memory (Fig. 4B). Importantly, the left prefrontal region that displays hypometabolism in depression described by Bench et al23 is close to this target (Fig. 1). Using the inverse MRInative–MRITalairach transformation matrix, x, y and z coordinates of this location were calculated for each individual's “native” brain space and a figure-eight coil was positioned over this location using frameless stereotaxy; during the study, the coil was held with a rigid arm mounted at the back of the scanner's gantry. The transmission scan was carried out and used not only for the attenuation corrections but also for the verification of coil positioning (Fig. 4C).109 Six 60-second water-bolus (15O-H2O) emission scans were acquired afterward: 2 baseline scans with no TMS applied and 4 TMS scans during which 30 pairs of pulses were administered with intensity at the individual's motor threshold (Fig. 4A). White noise (90 dB) was played over insert earphones during all scans to attenuate the coil-generated clicks. The double-pulse TMS was applied during the scans at 0.5-Hz frequency to provide us with a measure of cortical excitability and connectivity of the left MDLFC. In addition, we investigated the putative modulatory effect of high-frequency rTMS on MDLFC excitability and connectivity. To do this, we applied 2 series of rTMS between the first and last TMS scans; the following TMS parameters were used for each series: 15 1-second trains; 10 pulses in each train (i.e., 10 Hz); 10-second between-train intervals; intensity at motor threshold (Fig. 4A). The stimulation site was identical for the double-pulse TMS and rTMS. In response to the double-pulse TMS applied before rTMS, CBF decreased both at the stimulation site and in several distal regions presumably connected to the site, including the ACC (Fig. 4D, Fig. 4E). Although the precise mechanisms are unknown, such CBF decreases most likely reflect a net decrease in excitatory synaptic activity mediated by TMS-induced release of γ-aminobutyric acid (GABA).110,111,112 Following the 2 series of rTMS, this “suppression” response was reversed, resulting in double-pulse-induced increases in CBF that were maximal during the last TMS scan (Fig. 4F). Using correlational analysis, a network of cortical regions was revealed in which the blood-flow response to double-pulse TMS covaried with that at the stimulation site, including the contralateral MDLFC and the ACC (Fig. 4G). Overall, this study demonstrated that a mere 30 trains (300 pulses) of 10-Hz rTMS can induce subtle changes in cortical excitability and connectivity of the stimulated region. Such a putative reversal of the initial “inhibitory” response to low-frequency TMS is akin to the phenomenon of “long-term transformation,” namely, the transformation of the hyperpolarizing GABA-mediated inhibitory postsynaptic potentials into depolarizing responses as observed in vitro.113,114
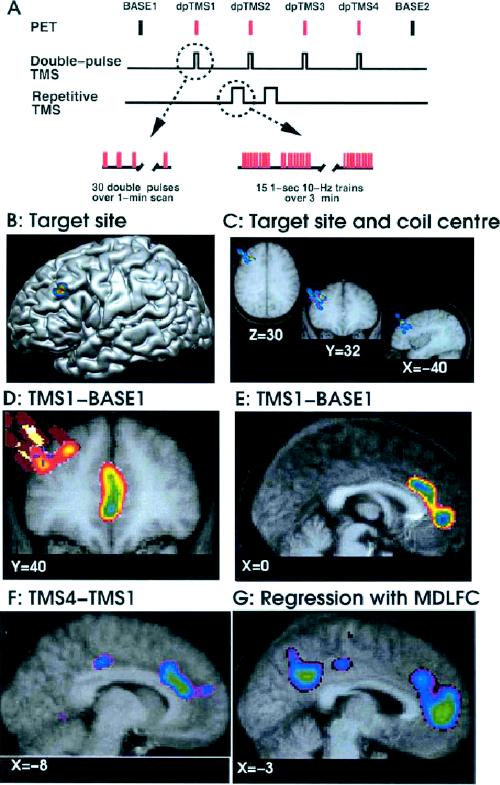
Fig. 4: Modulation of corticocortical connectivity by rTMS. The flowchart (A) indicates the sequence of events during the TMS/PETstudies; the PET scans were repeated every 10 min (Base, no TMS applied; dpTMS, double-pulse TMS). The target site (B) within the left MDLFC was selected from a previous blood-flow activation study by Petrides et al;108 the “peak” is located just above the left inferior frontal sulcus. The proximity of the target site (crosshair) and the coil centre derived from the transmission scans in the 8 subjects (colour “bars”) demonstrate the successful positioning of the coil with frameless stereotaxy (C). After subtraction (D–F) and regression (G) analyses of blood-flow data, the images depict the exact locations of statistically significant decreases (D, E) and increases (F) in blood flow and significant positive correlation with blood flow at the stimulation site (G). The thresholded maps of t- statistic values (t > 3.0 or t < –3.0) are superimposed on coronal (D) and sagittal (E–G) sections through the average MRI of the 8 subjects. All images are aligned within the standardized stereotaxic space. Reproduced with permission from Blackwell Publishing (Eur J Neurosci 2001;14:1405-11).29
The second type of study uses PET ligands that allow the investigator to measure the regional release of neurotransmitters. The measurement of TMS-induced release of specific neurotransmitters is particularly attractive, because it allows us to delineate the neurochemical pathways involved in mediating behavioural and treatment effects of TMS. We have begun this research by studying TMS-induced release of dopamine; 11C-raclopride was employed to measure the release of dopamine in the human striatum in response to rTMS of the left MDLFC, with the left occipital cortex being used as a control site.30 On 2 successive days, 3 series of 15 10-Hz trains of rTMS were applied 10 minutes apart with a circular coil at either of the 2 sites. 11C-raclopride was injected immediately after the end of the last rTMS series and the tracer uptake in the brain was measured over the next 60 minutes. Voxel-wise 11C-raclopride binding potential was calculated using a simplified reference tissue method115,116 to generate statistical parametric images of change in binding potential.117 This analysis revealed a significant decrease in binding potential in the left caudate nucleus after rTMS of the left MDLFC, as compared with rTMS of the occipital cortex (Fig. 5).30,31,118,119 Such a reduction in 11C-raclopride binding potential is indicative of an increase in extracellular dopamine concentration.120,121 It is likely that this TMS-induced focal release of dopamine in the ipsilateral caudate nucleus is mediated by excitatory corticostriatal projections known to originate in high density in the primate prefrontal cortex and to synapse at the vicinity of nigrostriatal dopaminergic nerve terminals. More recently, we used an identical stimulation and imaging protocol but changed the stimulation site: the left primary motor cortex was stimulated instead of the left MDLFC.31 The results of this study were clear: dopamine was released in the ipsilateral putamen rather than the caudate nucleus (Fig. 5). This finding is consistent with the organization of the corticostriatal loops, namely, the existence of dense projections from the prefrontal and primary motor cortices targeting, respectively, the head of the caudate nucleus and the putamen (Fig. 5). Overall, these studies confirm the feasibility of using PET to investigate TMS-induced changes in specific neurotransmitter systems of the human brain and open up new avenues for studies of the pathophysiology of neurologic and psychiatric disorders.
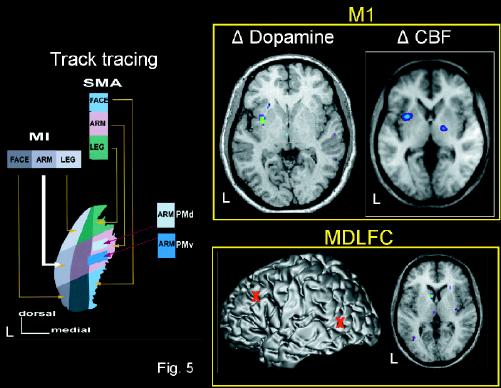
Fig. 5: Dopamine release induced by rTMS of the prefrontal cortex. Left: organization of corticostriatal projections to the monkey putamen.118
Top right: changes in dopamine release31 and cerebral blood flow (CBF)119 in the human putamen after rTMS was applied over the left primary motor cortex (M1). Bottom right: changes in dopamine release30 in the human caudate nucleus after rTMS was applied over the left MDLFC. Location (red markers) of the 2 stimulation sites, the left MDLFC and the left occipital cortex, on the MRI of 1 subject in stereotaxic space. PMd = dorsal promoter, PMv = ventral promoter. Reproduced with permission from Springer-Verlag (Exp Brain Res 1998; 120:114-28).118
Studies of long-term (days, weeks) effects of rTMS are crucial for our understanding of the neural mechanisms that mediate some of the treatment effects of rTMS. Several reports have been published in which single-photon emission computed tomography (SPECT)122,123,124 or PET125,126 were used to measure neural changes before and after rTMS treatment of major depression; the treatment typically consisted of 10 days of high-frequency (10–20 Hz) rTMS applied daily over the left MDLFC. In most of these studies, such treatment resulted in the increase of perfusion or metabolism in the prefrontal cortex. Kimbrell et al126and Speer et al125 employed both high-frequency (20 Hz) and low-frequency (1 Hz) stimulation and observed, respectively, increases and decreases in CBF and/or glucose metabolism in the frontal cortex and other brain regions. Interpretation of previous studies is somewhat limited, however, because of the lack of detailed information about the exact coil position in relation to the underlying cortical regions and, in many cases, the absence of adequate control stimulation or a comparison group. Furthermore, in most of the published studies, the blood-flow or metabolic measurements were acquired during a resting baseline. In order to facilitate interpretation of the long-term effects of rTMS, it may be useful to evaluate the effect of rTMS applied between scans on cortical excitability and connectivity by “probing” neural activity with TMS stimuli applied during the scanning (see earlier). Overall, it is likely that future imaging studies will provide useful information vis-à-vis potential therapeutic effects of rTMS in depression, as well as in other psychiatric and neurologic disorders.127,128 The combination of TMS and imaging may, in this context, lead to improvement of the treatment protocols as well as an increased understanding of the pathophysiology of these brain disorders.
Previous articles in this series
Lesch KP. Gene–environment interaction and the genetics of depression. J Psychiatry Neurosci 2004; 29(3):174-84.
Barden N. Implication of the hypothalamic–pituitary–adrenal axis in the physiopathology of depression. J Psychiatry Neurosci 2004;29(3):185-93.
Malberg J. Implications of adult hippocampal neurogenesis in antidepressant action. J Psychiatry Neurosci 2004;29(3):196-205.
Blier P, Gobbi G, Haddjeri N, Santarelli L, Mathew G, Hen R. Impact of substance P receptor antagonism on the serotonin and norepinephrine systems: relevance to the antidepressant/anxiolytic response. J Psychiatry Neurosci 2004;29(3):208-18.
Acknowledgments
We thank our colleagues at the Montreal Neurological Institute for their contributions to the ongoing transcranial magnetic stimulation/positron emission tomography studies of cortical excitability and connectivity. Dr. Paus's research is supported by the Canadian Institutes of Health Research, the Canadian Foundation for Innovation and the Natural Sciences and Engineering Research Council of Canada. Cadwell Laboratories Inc. and Rogue Research Inc. donated equipment and services to the laboratory.
Footnotes
Competing interests: None declared.
Correspondence to: Dr. Tomáš Paus, Cognitive Neuroscience Unit/Neuropsychology Department, Montreal Neurological Institute, 3801 University St., Montréal QC H3A 2B4; fax 514 398-1338; tomas@bic.mni.mcgill.ca
Submitted July 4, 2003; Revised Feb. 6, 2004; Accepted Feb. 24, 2004
References
- 1.Blazer DG, Kessler RC, McGonagle KA, Swartz MS. The prevalence and distribution of major depression in a national community sample: the National Comorbidity Survey. Am J Psychiatry 1994;151:979-86. [DOI] [PubMed]
- 2.Hirschfeld RM. Efficacy of SSRIs and newer antidepressants in severe depression: comparison with TCAs. J Clin Psychiatry 1999; 60:326-35. [DOI] [PubMed]
- 3.American Psychiatric Association. The practice of electroconvulsive therapy. New York: Oxford University Press; 1990.
- 4.Murphy FC, Sahakian BJ, Carroll RE. Cognitive impairment in depression: psychological models and clinical issues. In: Ebert D, Ebmeier KP, editors. Advances in biological psychiatry. Vol 19 of New models for depression series. New York : Karger; 1998. p. 1-33.
- 5.Sobin C, Sackheim HA. Psychomotor symptoms of depression. Am J Psychiatry 1997;154:4-17. [DOI] [PubMed]
- 6.Flint AJ, Black SE, Campbell-Taylor I, Gailey GF, Levinton C. Abnormal speech articulation, psychomotor retardation, and subcortical dysfunction in major depression. J Psychiatr Res 1993; 27:285-7. [DOI] [PubMed]
- 7.Godfrey HPD, Knight RG. The validity of actometer and speech activity measures in the assessment of depressed patients. Br J Psychiatry 1984;145:159-63. [DOI] [PubMed]
- 8.Nilsonne A. Speech characteristics as indicators of depressive illness. Acta Psychiatr Scand 1988;77:253-63. [DOI] [PubMed]
- 9.Stassen HH, Kuny S, Hell D. The speech analysis approach to determining onset of improvement under antidepressants. Eur Neuropsychopharmacol 1998;8:303-10. [DOI] [PubMed]
- 10.McKenna FP, Lewis C. A speech rate measure of laboratory induced affect: the role of demand characteristics revisited. Br J Clin Psychol 1994;33:345-51. [DOI] [PubMed]
- 11.Garcia-Toro M, Talavera JA, Saiz-Ruiz J, Gonzalez A. Prosody impairment in depression measured through acoustical analysis. J Nerv Ment Dis 2000;188:824-9. [DOI] [PubMed]
- 12.Alpert M, Pouget ER, Silva RR. Reflections of depression in acoustic measures of the patient's speech. J Affect Disord 2001; 66: 59-69. [DOI] [PubMed]
- 13.Purcell R, Maruff P, Kyrios M, Pantelis C. Neuropsychological function in young patients with unipolar major depression. Psychol Med 1997;27:1277-85. [DOI] [PubMed]
- 14.Lemelin S, Baruch P, Vincent A, Everett J, Vincent P. Distractibility and processing resource deficit in major depression. Evidence for two deficient attentional processing models. J Nerv Ment Dis 1997;185:542-8. [DOI] [PubMed]
- 15.Schatzberg AF, Posener JA, DeBattista C, Kalehzan BM, Rothschild AJ, Shear PK. Neuropsychological deficits in psychotic versus nonpsychotic major depression and no mental illness. Am J Psychiatry 2000;157:1095-100. [DOI] [PubMed]
- 16.Trichard C, Martinot JL, Alagille M, Masure MC, Hardy P, Ginestet D, et al. Time course of prefrontal lobe dysfunction in severely depressed in-patients: a longitudinal neuropsychological study. Psychol Med 1995;25:79-85. [DOI] [PubMed]
- 17.Austin MP, Mitchell P, Wilhelm K, Parker G, Hickie I, Brodaty H, et al. Cognitive function in depression: A distinct pattern of frontal impairment in melancholia? Psychol Med 1999;29:73-85. [DOI] [PubMed]
- 18.Degl'Innocenti A, Agren H, Backman L. Executive deficits in major depression. Acta Psychiatr Scand 1998;97:182-8. [DOI] [PubMed]
- 19.Portefield T, Cook M, Deary IJ, Ebmeier KP. Neuropsychological function and diurnal variation in depression. J Clin Exp Neuropsychol 1997;19:906-13. [DOI] [PubMed]
- 20.Fossati P, Amar G, Raoux N, Ergis AM, Allilaire JF. Executive functioning and verbal memory in young patients with unipolar depression and schizophrenia. Psychiatry Res 1999;89:171-87. [DOI] [PubMed]
- 21.Baxter LR Jr, Phelps ME, Mazziotta JC, Schwartz JM, Gerner RH, Selin CE, et al. Cerebral metabolic rates for glucose in mood disorders. Studies with positron emission tomography and fluorodeoxyglucose F18. Arch Gen Psychiatry 1985;42:441-7. [DOI] [PubMed]
- 22.Baxter LR Jr, Schwartz JM, Phelps ME, Mazziotta JC, Guze BH, Selin CE, et al. Reduction of prefrontal cortex glucose metabolism common to three types of depression. Arch Gen Psychiatry 1989;46:243-50. [DOI] [PubMed]
- 23.Bench CJ, Friston KJ, Brown RG, Scott LC, Franckowiak RS, Dolan RJ. The anatomy of melancholia-focal abnormalities of cerebral blood flow in major depression. Psychol Med 1992; 22: 607-15. [DOI] [PubMed]
- 24.Bench CJ, Friston KJ, Brown RG, Scott LC, Franckowiak RS, Dolan RJ. Regional cerebral blood flow in depression measured by positron emission tomography: the relationship with clinical dimensions. Psychol Med 1993;23:579-90. [DOI] [PubMed]
- 25.Bench CJ, Frackowiak RS, Dolan RJ. Changes in cerebral blood flow on recovery from depression. Psychol Med 1995;25:247-61. [DOI] [PubMed]
- 26.Martinot JL, Hardy P, Feline A, Huret JD, Mazoyer B, Attar-Levy D, et al. Left prefrontal glucose hypometabolism in the depressed state: a confirmation. Am J Psychiatry 1990;147:1313-7. [DOI] [PubMed]
- 27.Mayberg HS, Brannan SK, Mahurin RK, Jerabek PA, Brickman JS, Tekell JL, et al. Cingulate function in depression: a potential predictor of treatment response. Neuroreport 1997;8:1057-61. [DOI] [PubMed]
- 28.Drevets W, Price J, Simpson J, Todd R, Reich T, Vannier M, et al. Subgenual prefrontal cortex abnormalities in mood disorders. Nature 1997;386:824-7. [DOI] [PubMed]
- 29.Paus T, Castro-Alamancos M, Petrides M. Cortico-cortical connectivity of the human mid-dorsolateral frontal cortex and its modulation by repetitive transcranial magnetic stimulation. Eur J Neurosci 2001;14:1405-11. [DOI] [PubMed]
- 30.Strafella A, Paus T, Barrett J, Dagher A. Repetitive transcranial magnetic stimulation of the human prefrontal cortex induces dopamine release in the caudate nucleus. J Neurosci 2001;21:1-4. [DOI] [PMC free article] [PubMed]
- 31.Strafella AP, Paus T, Fraraccio M, Dagher A. Striatal dopamine release induced by repetitive transcranial magnetic stimulation of the human motor cortex. Brain 2003;126:2609-15. [DOI] [PubMed]
- 32.Buchsbaum MS, Wu J, Siegel BV, Hackett E, Trenary M, Abel L, et al. Effect of sertraline on regional metabolic rate in patients with affective disorder. Biol Psychiatry 1997;41:15-22. [DOI] [PubMed]
- 33.Mayberg HS, Brannan SK, Tekell JL, Silva JA, Mahurin RK, McGinnis S, et al. Regional metabolic effects of fluoxetine in major depression: serial changes and relationship to clinical response. Biol Psychiatry 2000;48:830-43. [DOI] [PubMed]
- 34.Kennedy SH, Evans KR, Kruger S, Mayberg HS, Meyer JH, McCann S, et al. Changes in regional brain glucose metabolism measured with positron emission tomography after paroxetine treatment of major depression. Am J Psychiatry 2001; 158:899-905. [DOI] [PubMed]
- 35.Videbech P, Ravnkilde B, Pedersen TH, Hartvig H, Egander A, Clemmensen K, et al. The Danish PET/depression project: clinical symptoms and cerebral blood flow. A regions-of-interest analysis. Acta Psychiatr Scand 2002;106:35-44. [DOI] [PubMed]
- 36.Henriques JB, Davidson RJ. Left frontal hypoactivation in depression. J Abnorm Psychol 1991;100:535-45. [DOI] [PubMed]
- 37.Bruder GE, Fong R, Tenke CE, Leite P, Towey JP, Stewart JE, et al. Regional brain asymmetries in major depression with or without an anxiety disorder: a quantitative electroencephalographic study. Biol Psychiatry 1997;41:939-48. [DOI] [PubMed]
- 38.Debener S, Beauducel A, Nessler D, Brocke B, Heilemann H, Kayser J. Is resting anterior EEG alpha asymmetry a trait marker for depression? Findings for healthy adults and clinically depressed patients. Neuropsychobiology 2000;41:31-7. [DOI] [PubMed]
- 39.Davidson RJ, Pizzagalli D, Nitschke JB, Putnam K. Depression: perspectives from affective neuroscience. Ann Rev Psychol 2002; 53:545-74. [DOI] [PubMed]
- 40.Vogt BA, Nimchinsky EA, Vogt LJ, Hof PR. Human cingulate cortex: surface features, flat maps, and cytoarchitecture. J Comp Neurol 1995;359:490-506. [DOI] [PubMed]
- 41.Mayberg HS, Lewis PJ, Regenold W, Wagner HN. Paralimbic hypoperfusion in unipolar depression. J Nucl Med 1994;35:929-34. [PubMed]
- 42.Ongur D, Drevets WC, Price J. Glial reduction in the subgenual prefrontal cortex in mood disoders. Proc Natl Acad Sci U S A 1998;95:13290-5. [DOI] [PMC free article] [PubMed]
- 43.Drevets WC. Prefrontal cortical-amygdalar metabolism in major depression. Ann N Y Acad Sci 1999;877:614-37. [DOI] [PubMed]
- 44.Osuch EA, Ketter TA, Kimbrell TA, George MS, Benson BE, Willis MW, et al. Regional cerebral metabolism associated with anxiety symptoms in affective disorder patients. Biol Psychiatry 2000;48:1020-3. [DOI] [PubMed]
- 45.Brody AL, Saxena S, Mandelkern MA, Fairbanks LA, Ho ML, Baxter LR. Brain metabolic changes associated with symptom factor improvement in major depressive disorder. Biol Psychiatry 2001;50:171-8. [DOI] [PubMed]
- 46.Drevets WC, Bogers W, Raichle ME. Functional anatomical correlates of antidepressant drug treatment assessed using PET measures of regional glucose metabolism. Eur Neuropsychopharmacol 2002;12:527-44. [DOI] [PubMed]
- 47.Wu J, Buchsbaum MS, Gillin JC, Tang C, Cadwell S, Wiegand M, et al. Prediction of antidepressant effects of sleep deprivation by metabolic rates in the ventral anterior cingulate and medial prefrontal cortex. Am J Psychiatry 1999;156:1149-58. [DOI] [PubMed]
- 48.Mayberg HS, Liotti M, Brannan SK, McGinnis S, Mahurin RK, Jerabek PA, et al. Reciprocal limbic-cortical function and negative mood: converging PET findings in depression and normal sadness. Am J Psychiatry 1999;156:675-82. [DOI] [PubMed]
- 49.Dunn RT, Kimbrell TA, Ketter TA, Frye MA, Willis MW, Luckenbaugh DA, et al. Principal components of the Beck Depression Inventory and regional cerebral metabolism in unipolar and bipolar depression. Biol Psychiatry 2002;51:387-99. [DOI] [PubMed]
- 50.Liotti M, Mayberg HS, McGinnis S, Brannan SL, Jerabek P. Unmasking disease-specific cerebral blood flow abnormalities: mood challenge in patients with remitted unipolar depression. Am J Psychiatry 2002;159:1830-40. [DOI] [PubMed]
- 51.Pizzagalli D, Pascual-Marqui RD, Nitschke JB, Oakes TR, Larson CL, Abercrombie HC, et al. Anterior cingulate activity as a predictor of degree of treatment response in major depression: evidence from brain electrical tomography analysis. Am J Psychiatry 2001;158:405-15. [DOI] [PubMed]
- 52.Stefurak TL, Mikulis D, Mayberg H. Deep brain stimulation associated dysphoria and cortico-limbic changes detected by fMRI. Mov Disord 2001;16(Suppl 1):S54-5.
- 53.Brody AL, Saxena S, Silverman DH, Alborzian S, Faribanks LA, Phelps ME, et al. Brain metabolic changes in major depressive disorder from pre- to post treatment with paroxetine. Psychiatry Res 1999;91:127-39. [DOI] [PubMed]
- 54.Ketter TA, Kimbrell TA, George MS, Willis MW, Benson BE, Danielson A, et al. Baseline cerebral hypermetabolism associated with carbamazepine response, and hypometabolism with nimodipine response in mood disorders. Biol Psychiatry 1999; 46: 1364-74. [DOI] [PubMed]
- 55.Paus T. Combination of transcranial magnetic stimulation with brain imaging. In: Mazziotta J, Toga A, editors. Brain mapping: the methods. 2nd ed. San Diego: Academic Press; 2002. p. 691-705.
- 56.Rothwell JC, Hallett M, Berardelli A, Eisen A, Rossini P, Paulus W. Magnetic stimulation: motor evoked potenials. Electroencephalogr Clin Neurophysiol 1999;52:97-103. [PubMed]
- 57.Pascual-Leone A, Tarazona F, Catala MD. Applications of transcranial magnetic stimulation in studies on motor learning. Electroencephalogr Clin Neurophysiol 1999;51:157-61. [PubMed]
- 58.Kosslyn SM, Pascual-Leone A, Felician O, Camposano S, Keenan HP, Thompson WL. The role of area 17 in visual imagery: convergent evidence from PET and rTMS. Science 1999; 284: 167-70. [DOI] [PubMed]
- 59.George MS, Wasserman EM, Williams WA, Steppel J, Pascual-Leone A, Basser P, et al. Changes in mood and hormone levels after rTMS of the prefrontal cortex. J Neuropsychiatry Clin Neurosci 1996;8:172-80. [DOI] [PubMed]
- 60.Pascual-Leone A, Dolores-Catala M, Pascual-Leone Pascual A. Lateralized effect of rapid rate transcranial magnetic stimulation of the prefrontal cortex on mood. Neurology 1996;46:499-502. [DOI] [PubMed]
- 61.Hoflich G, Kasper S, Hufnagel A, Ruhrmann S, Moller HJ. Application of transcranial magnetic stimulation in treatment of drug-resistant major depression: a report of two cases. Hum Psychopharmacol 1993;8:361-5.
- 62.McNamara B, Ray JL, Arthurs OJ, Boniface S. Transcranial magnetic stimulation for depression and other psychiatric disorders. Psychol Med 2001; 31:1141-6. [DOI] [PubMed]
- 63.Burt T, Lisanby SH, Sackeim HA. Neuropsychiatric applications of transcranial magnetic stimulation: a meta analysis. Int J Neuropsychopharmacol 2002;5:73-103. [DOI] [PubMed]
- 64.Gershon AA, Dannon PN, Grunhaus L. Transcranial magnetic stimulation in the treatment of depression. Am J Psychiatry 2003; 160:835-45. [DOI] [PubMed]
- 65.Pascual-Leone A, Rubio B, Pallardo F, Catale MD. Rapid-rate transcranial magnetic stimulation of the left dorsolateral prefrontal cortex in drug-resistant depression. Lancet 1996;348:233-7. [DOI] [PubMed]
- 66.George MS, Wasserman EM, Kimbrell TA, Little JT, Williams WE, Danielson AL, et al. Mood improvement following daily left prefrontal repetitive magnetic stimulation in patients with depression: a placebo-controlled crossover trial. Am J Psychiatry 1997;154:1752-6. [DOI] [PubMed]
- 67.Berman RM, Narasimhan M, Sanacora G, Miano AP, Hoffman RE, Hu XS, et al. A randomized clinical trial of repetitive transcranial magnetic stimulation in the treatment of major depression. Biol Psychiatry 2000;47:332-7. [DOI] [PubMed]
- 68.Triggs WJ, Karin JM, Greer R, Rossi F, Bowers D, Kortenkamp S, et al. Effects of left frontal TMS on depressed mood, cognition and corticomotor threshold. Biol Psychiatry 1999;45:1440-6. [DOI] [PubMed]
- 69.Grunhaus L, Dannon PN, Schreiber S, Dolberg OH, Amiaz R, Ziv R, et al. Repetitive transcranial magnetic stimulation is as effective as electroconvulsive therapy in the treatment of nondelusional major depressive disorder: an open study. Biol Psychiatry 2000;47:314-24. [DOI] [PubMed]
- 70.Avery DH, Claypoole K, Robinson L, Neumaier PJF, Dunner DL, Scheele L, et al. Repetitive transcranial magnetic stimulation in the treatment of medication-resistant depression: preliminary data. J Nerv Ment Dis 1999;187:114-7. [DOI] [PubMed]
- 71.The Avery-George-Holtzheimer database of rTMS depression studies. Available: www.ists.unibe.ch/TMSAvery.htm (accessed 2004 June 11).
- 72.Barrett J, Della-Maggiore V, Chouinard PA, Paus T. Mechanisms of action underlying the effect of repetitive transcranial magnetic stimulation on mood: behavioral and brain imaging studies. Neuropsychopharmacology 2004;29:1172-89. [DOI] [PubMed]
- 73.Petrides M, Pandya DN. Dorsolateral prefrontal cortex: comparative cytoarchitectonic analysis in the human and the macaque brain and corticocortical connection patterns. Eur J Neurosci 1999;11:1011-36. [DOI] [PubMed]
- 74.Petrides M, Pandya DN. Comparative architectonic analysis of the human and the macaque frontal cortex. In: Boller F, Grafman J, editors. Handbook of neuropsychology. Amsterdam: Elsevier; 1994. p. 17-58.
- 75.Vogt BA, Pandya DN. Cingulate cortex of the rhesus monkey: II. Cortical afferents. J Comp Neurol 1987;262:271-89. [DOI] [PubMed]
- 76.Barbas H. Architecture and cortical connections of the prefrontal cortex in the rhesus monkey. In: Chauvel P, Delgado-Escueta AV, editors. Advances in neurology. Vol 57. New York: Raven Press; 1992. p. 91-115. [PubMed]
- 77.Bates JF, Goldman-Rakic PS. Prefrontal connections of medial motor areas in the rhesus monkey. J Comp Neurol 1993;336:211-28. [DOI] [PubMed]
- 78.Petrides M. Mapping prefrontal cortical systems for the control of cognition. In: Mazziotta JC, Toga AW, editors. Brain mapping: the systems. San Diego: Academic Press; 2000. p. 159-75.
- 79.Milner B, Petrides M. Behavioral effects of frontal-lobe lesions in man. Trends Neurosci 1984;7:403-7.
- 80.Milner B. Some effects of frontal lobectomy in man. In: Warren JM, Akert K, editors. The frontal granular cortex and behavior. New York: McGraw-Hill; 1964. p. 313-34.
- 81.Paus T. Primate anterior cingulate cortex: where motor control, drive and cognition interface. Nat Rev Neurosci 2001;2:417-24. [DOI] [PubMed]
- 82.Paus T, Koski L, Caramanos Z, Westbury C. Regional differences in the effects of task difficulty and motor output on blood flow response in the human anterior cingulate cortex: a review of 107 PET activation studies. Neuroreport 1998;9:R37-47. [DOI] [PubMed]
- 83.Koski L, Paus T. Functional connectivity of the anterior cingulate cortex within the human frontal lobe: a brain-mapping meta-analysis. Exp Brain Res 2000;133:55-65. [DOI] [PubMed]
- 84.Paus T, Tomaiuolo F, Otaky N, MacDonald D, Petrides M, Atlas J, et al. Human cingulate and paracingulate sulci: pattern, variability, asymmetry, and probabilistic map. Cereb Cortex 1996; 6:207-14. [DOI] [PubMed]
- 85.Paus T, Otaky N, Caramanos Z, MacDonald D, Zijdenbos A, D'Avirro D, et al. In vivo morphometry of the intrasulcal gray matter in the human cingulate, paracingulate, and superior-rostral sulci: hemispheric asymmetries, gender differences and probability maps. J Comp Neurol 1996;376:664-73. [DOI] [PubMed]
- 86.Sarkissov S, Filimonoff J, Kononova E, Preobraschenskaja I, Kukuew L. Atlas of the cytoarchitectonics of the human cerebral cortex. Moscow: Medzig; 1955.
- 87.Zilles K, Schlaug G, Matelli M, Luppino G, Schleicher A, Qu M, et al. Mapping of human and macaque sensorimotor areas by integrating architectonic, transmitter receptor, MRI and PET data. J Anat 1995;187:515-37. [PMC free article] [PubMed]
- 88.Barbas H, Pandya DN. Architecture and intrinsic connections of the prefrontal cortex in the rhesus monkey. J Comp Neurol 1989; 286:353-75. [DOI] [PubMed]
- 89.Matelli M, Luppino G, Rizzolatti G. Architecture of superior and mesial area 6 and the adjacent cingulate cortex in the macaque monkey. J Comp Neurol 1991;311:445-62. [DOI] [PubMed]
- 90.Vogt BA, Pandya DN, Rosene DL. Cingulate cortex of the rhesus monkey: I. Cytoarchitecture and thalamic afferents. J Comp Neurol 1987;262:256-70. [DOI] [PubMed]
- 91.Morecraft RJ, Geula C, Mesulam MM. Architecture of connectivity within a cingulo-fronto-parietal neurocognitive network for directed attention. Arch Neurol 1993;50:279-84. [DOI] [PubMed]
- 92.Ongur D, An X, Price J. Prefrontal cortical projections to the hypothalamus in macaque monkeys. J Comp Neurol 1998; 401: 480-505. [PubMed]
- 93.Kunishio K, Haber S. Primate cingulostriatal projection: limbic striatal versus sensorimotor striatal input. J Comp Neurol 1994;350:337-56. [DOI] [PubMed]
- 94.An X, Bandler R, Ongur D, Price J. Prefrontal cortical projections to longitudinal columns in the midbrain periaqueductal grey in macaque monkeys. J Comp Neurol 1998;401:455-79. [PubMed]
- 95.Kaada B. Cingulate, posterior orbital, anterior insular and temporal pole cortex. In: Field J, Magoun H, Hall V, editors. Neurophysiology. Vol. 2. Washington: American Physiological Society; 1960. p. 1345-72.
- 96.Neafsey E. Prefrontal cortical control of the autonomic nervous system: anatomical and physiological observations. Prog Brain Res 1990;85:147-65. [DOI] [PubMed]
- 97.Aitken PG. Cortical control of conditioned and spontaneous vocal behavior in rhesus monkeys. Brain Lang 1981;13:171-84. [DOI] [PubMed]
- 98.McLean PD, Newman JD. Role of the midline frontocortical cortex in production of the isolation call of squirrel monkeys. Brain Res 1988;450:111-23. [DOI] [PubMed]
- 99.Sutton D, Larson C, Lindeman RC. Neocortical and limbic lesion effects on primate phonation. Brain Res 1974;71:61-75. [DOI] [PubMed]
- 100.Barris RW, Shuman HR. Bilateral anterior cingulate gyrus lesions. Syndrome of the anterior cingulate gyri. Neurology 1953; 3: 44-52. [DOI] [PubMed]
- 101.Nielsen JM, Jacobs LL. Bilateral lesions of the anterior cingulate gyri. Report of case. Bull Los Angel Neuro Soc 1951;16:231-4. [PubMed]
- 102.Jurgens U, Von Cramon D. On the role of the anterior cingulate cortex in phonation: a case report. Brain Lang 1982;15:234-48. [DOI] [PubMed]
- 103.Paus T, Petrides M, Evans AC, Meyer E. Role of the human anterior cingulate cortex in the control of oculomotor, manual, and speech responses: a positron emission tomography study. J Neurophysiol 1993;70:453-69. [DOI] [PubMed]
- 104.Pardo JV, Pardo PJ, Janer KW, Raichle ME. The anterior cingulate cortex mediates processing selection in the Stroop attentional conflict paradigm. Proc Natl Acad Sci U S A 1990;87:256-9. [DOI] [PMC free article] [PubMed]
- 105.Bench CJ, Frith CD, Grasby PM, Friston KJ, Paulesu E, Frackowiak RS, et al. Investigations of the functional anatomy of attention using the Stroop test. Neuropsychologia 1993;31:907-22. [DOI] [PubMed]
- 106.Taylor SF, Kornblum S, Minoshima S, Oliver LM, Koeppe RA. Changes in medial cortical blood flow with a stimulus-response compatibility task. Neuropsychologia 1994;32:249-55. [DOI] [PubMed]
- 107.Barrett J, Pike GB, Paus T. The role of the anterior cingulate cortex in pitch variation during sad affect. Eur J Neurosci 2004; 19: 458-64. [DOI] [PubMed]
- 108.Petrides M, Alivisatos B, Meyer E, Evans AC. Functional activation of the human frontal cortex during the performance of verbal working memory tasks. Proc Natl Acad Sci U S A 1993; 90: 878-82. [DOI] [PMC free article] [PubMed]
- 109.Paus T, Wolforth M. Transcranial magnetic stimulation during PET: reaching and verifying the target site. Hum Brain Mapp 1998;6:399-402. [DOI] [PMC free article] [PubMed]
- 110.Ziemann U, Lonnecker S, Steinhoff BJ, Paulus W. Effects of antiepileptic drugs on motor cortex excitability in humans: a transcranial magnetic stimulation study. Ann Neurol 1996; 40: 367-78. [DOI] [PubMed]
- 111.Werhahn KJ, Kunesch E, Noachtar S, Benecke R, Classen J. Differential effects on motorcortical inhibition induced by blockade of GABA uptake in humans. J Physiol 1999;517:591-7. [DOI] [PMC free article] [PubMed]
- 112.Avoli M, Hwa G, Louvel J, Kurcewicz I, Pumain R, Lacaille JC. Functional and pharmacological properties of GABA-mediated inhibition in the human neocortex. Can J Physiol Pharmacol 1997;75:526-34. [PubMed]
- 113.Sun MK, Nelson TJ, Alkon DL. Functional switching of GABAergic synapses by ryanodine receptor activation. Proc Natl Acad Sci U S A 2000;97:12300-5. [DOI] [PMC free article] [PubMed]
- 114.Sun M, Dahl D, Alkon DL. Heterosynaptic transformation of GABAergic gating in the hippocampus and effects of carbonic anhydrase inhibition. J Pharmacol Exp Ther 2001;296:811-7. [PubMed]
- 115.Lammertsma AA, Hume SP. Simplified reference tissue model for PET receptor studies. Neuroimage 1996;4:153-8. [DOI] [PubMed]
- 116.Gunn RN, Lammertsma A, Hume SP, Cunningham VJ. Parametric imaging of ligand-receptor binding in PET using a simplified reference region model. Neuroimage 1997;6:279-87. [DOI] [PubMed]
- 117.Aston JA, Gunn RN, Worsley KJ, Ma Y, Evans AC, Dagher A. A statistical method for the analysis of positron emission tomography neuroreceptor ligand data. Neuroimage 2000;12:245-56. [DOI] [PubMed]
- 118.Takada M, Tokuno H, Nambu A, Inase M. Corticostriatal projections from the somatic motor areas of the frontal cortex in the macaque monkey: segregation versus overlap of input zones from the primary motor cortex, the supplementary motor area, and the premotor cortex. Exp Brain Res 1998;120:114-28. [DOI] [PubMed]
- 119.Chouinard PA, Van Der Werf YD, Leonard G, Paus T. Modulating neural networks with transcranial magnetic stimulation applied over the dorsal premotor and primary motor cortices. J Neurophysiol 2003;90:1071-83. [DOI] [PubMed]
- 120.Laruelle M, Iyer RN, Al-Tikriti MS, Zea-Ponce Y, Malison R, Zoghbi SS, et al. Microdialysis and SPECT measurements of amphetamine-induced dopamine release in nonhuman primates. Synapse 1997;25:1-14. [DOI] [PubMed]
- 121.Endres CJ, Kolachana BS, Saunders RC, Su T, Weinberger D, Breier A, et al. Kinetic modelling of [11C] raclopride: combined PET-microdialysis studies. J Cereb Blood Flow Metab 1997; 9:932-42. [DOI] [PubMed]
- 122.Peschina W, Conca A, Konig P, Fritzsche H, Beraus W. Low frequency rTMS as an add-on antidepressive strategy: heterogeneous impact on 99mTc-HMPAO and 18 F-FDG uptake as measured simultaneously with the double isotope SPECT technique. Pilot study. Nucl Med Commun 2001;22:867-73. [DOI] [PubMed]
- 123.Zheng XM. Regional cerebral blood flow changes in drug-resistant depressed patients following treatment with transcranial magnetic stimulation: a statistical parametric mapping analysis. Psychiatry Res 2000;100:75-80. [DOI] [PubMed]
- 124.Teneback CC, Nahas Z, Speer AM, Molloy M, Stallings LE, Spicer KM, et al. Changes in prefrontal cortex and paralimbic activity in depression following two weeks of daily left prefrontal TMS. J Neuropsychiatry Clin Neurosci 1999;11:426-35. [DOI] [PubMed]
- 125.Speer AM, Kimbrell TA, Wassermann EM, Repella JD, Willis MW, Herscovitch P, et al. Opposite effects of high and low frequency rTMS on regional brain activity in depressed patients. Biol Psychiatry 2000;48:1133-41. [DOI] [PubMed]
- 126.Kimbrell TA, Little JT, Dunn RT, Frye MA, Greenberg BD, Wasserman EM, et al. Frequency dependence of antidepressant response to left prefrontal repetitive transcranial magnetic stimulation (rTMS) as a function of glucose metabolism. Biol Psychiatry 1999;46:1603-13. [DOI] [PubMed]
- 127.Post A, Keck ME. Transcranial magnetic stimulation as a therapeutic tool in psychiatry: What do we know about the neurobiological mechanisms? J Psychiatr Res 2001;35:193-215. [DOI] [PubMed]
- 128.Wassermann EM, Lisanby SH. Therapeutic application of repetitive transcranial magnetic stimulation: a review. Clin Neurophysiol 2001;112:1367-77. [DOI] [PubMed]