

ABSTRACT
Klebsiella pneumoniae is an urgent public health threat because of resistance to carbapenems, antibiotics of last resort against Gram-negative bacterial infections. Despite the fact that K. pneumoniae is a leading cause of pneumonia in hospitalized patients, the bacterial factors required to cause disease are poorly understood. Insertion site sequencing combines transposon mutagenesis with high-throughput sequencing to simultaneously screen thousands of insertion mutants for fitness defects during infection. Using the recently sequenced K. pneumoniae strain KPPR1 in a well-established mouse model of pneumonia, insertion site sequencing was performed on a pool of >25,000 transposon mutants. The relative fitness requirement of each gene was ranked based on the ratio of lung to inoculum read counts and concordance between insertions in the same gene. This analysis revealed over 300 mutants with at least a 2-fold fitness defect and 69 with defects ranging from 10- to >2,000-fold. Construction of 6 isogenic mutants for use in competitive infections with the wild type confirmed their requirement for lung fitness. Critical fitness genes included those for the synthesis of branched-chain and aromatic amino acids that are essential in mice and humans, the transcriptional elongation factor RfaH, and the copper efflux pump CopA. The majority of fitness genes were conserved among reference strains representative of diverse pathotypes. These results indicate that regulation of outer membrane components and synthesis of amino acids that are essential to its host are critical for K. pneumoniae fitness in the lung.
IMPORTANCE
Klebsiella pneumoniae is a bacterium that commonly causes pneumonia in patients after they are admitted to the hospital. K. pneumoniae is becoming resistant to all available antibiotics, and when these infections spread to the bloodstream, over half of patients die. Since currently available antibiotics are failing, we must discover new ways to treat these infections. In this study, we asked what genes the bacterium needs to cause an infection, since the proteins encoded by these genes could be targets for new antibiotics. We identified over 300 genes that K. pneumoniae requires to grow in a mouse model of pneumonia. Many of the genes that we identified are found in K. pneumoniae isolates from throughout the world, including antibiotic-resistant forms. If new antibiotics could be made against the proteins that these genes encode, they may be broadly effective against K. pneumoniae.
INTRODUCTION
Klebsiella pneumoniae is a pathogenic Gram-negative bacterium that is a member of the Enterobacteriaceae family. While the bacterium is normally found in the flora of human skin, mouth, and intestinal tract, it is also well characterized as an opportunistic pathogen (1, 2). K. pneumoniae infections are associated with hospitalized patients with a weakened immune system. The bacterium causes a wide range of human diseases that include urinary tract infections, pyogenic liver abscess (PLA), and pneumonia (1, 3). Due to the alarming increase in carbapenem-resistant Enterobacteriaceae (CRE), of which K. pneumoniae comprises the majority of infections, the Centers for Disease Control (CDC) have designated these bacteria as an urgent threat to public health (4).
Because of the decreasing efficacy of antibiotics against K. pneumoniae, new treatment modalities must be developed, and a promising approach is through identifying bacterial factors required to cause disease. The few virulence factors identified to date include capsule and lipopolysaccharide (LPS), which vary by structure and serotype, and adhesins and siderophores, which vary in frequency among clinical isolates (1, 5–7). To develop novel therapies that will be broadly effective against K. pneumoniae, conserved targets must be identified.
Barriers to identifying new K. pneumoniae virulence factors include limitations with genetic sequencing techniques and the difficulties in manipulating the K. pneumoniae chromosome. In recent years, newer approaches such as transposon insertion site sequencing (InSeq) have allowed for greatly enhanced genetic screening. This method uses high-throughput sequencing to determine the frequency and chromosomal location of thousands of transposon mutations from a large pool of mutants in a single experiment (8, 9).
InSeq requires a fully sequenced genome and confirmation of mutant phenotypes by either simultaneously constructing an indexed transposon mutant library or constructing mutants in selected genes that elicit fitness defects. Because of advances in genetic sequencing, a number of high-quality reference sequences of K. pneumoniae are now available (10–13). However, many of the standard methods for gene replacement in Escherichia coli, such as suicide vector conjugation and linear DNA homologous recombination, are less efficient in K. pneumoniae or are complicated by intrinsic and acquired antibiotic resistance (14). The Lambda Red recombinase system, which is incredibly efficient in E. coli (15), could greatly increase the efficiency of gene recombination in K. pneumoniae (16). These advances now provide the ability to rigorously screen the genome for novel genes that contribute to virulence or in vivo fitness. Using this new sequencing approach and the Lambda Red recombinase system, we have identified numerous novel K. pneumoniae fitness genes required to infect the lung in a mouse model of pneumonia.
RESULTS
Transposon library construction, inoculation, and validation.
K. pneumoniae KPPR1 causes acute pneumonia complicated by bacteremia in a well-established murine model (5, 6). To facilitate InSeq analysis, the KPPR1 genome was sequenced to generate a single, gapless contig and the genes contained in it were annotated (10). The genome contains 5,191 predicted genes, including 25 rRNA, 85 tRNA, and 5,081 protein-coding sequences. To identify fitness factors required during lung infection, a library of ~25,000 transposon mutants was constructed to provide 99% genome coverage. Colony PCR and Southern blotting of representative transconjugants indicated that the library contained single, random transposon insertions at varied locations in the chromosome without integration of the vector (see Fig. S1 in the supplemental material). This library was inoculated into mice using an aspiration model of pneumonia at a dose of 1.4 × 106 CFU (5, 10, 17). After 24 h, the mean bacterial density increased to 5 × 106 CFU in the lung (Fig. 1A). To quantify and map each transposon onto the chromosome, genomic DNA was extracted from the inoculum and lung homogenates, and transposon junction fragments were generated. These fragments were sequenced using an Illumina HiSeq2500 instrument, mapped to the chromosome, and enumerated based on read count (18). Each sample had at least 68 million reads corresponding to 25,346 unique transposon insertions within genes (see Table S1), meeting the coverage goal as stated above. Of 5,191 predicted genes, 4,312 had at least one insertion detected, whereas 879 genes did not. Because the transconjugants were required to grow to a colony for inclusion in the library, genes without insertions may be essential. However, a larger mutagenesis library would be required to estimate more precisely the number of essential genes (8).
FIG 1 .

InSeq analysis indicates that fitness genes are important during pneumonia based on differential transposon read counts between the inoculum and mouse lung. Mice (n = 5) inoculated with 1.4 × 106 CFU of a pool of ~25,000 transposon mutants for 24 h were sacrificed and cultured on LB agar for CFU (A) or for DNA extraction, Illumina sequencing of transposon junctions, and read alignment with the KPPR1 genome to map and count transposon insertions. The locations and frequencies of transposon insertions in ilvD (B) and VK055_468 (C) are shown.
InSeq analysis identifies hundreds of potential K. pneumoniae lung fitness factors.
To identify in vivo fitness genes, the numbers of transposon insertion reads within each gene were compared between the lung and inoculum pools. Of genes with insertions, 3,880 had at least one shared transposon between the inoculum and lung samples (see Data Set S1 in the supplemental material) and were used for subsequent analysis. Since input and output data are “paired” in nature for each insertion, a P value was calculated for each insertion using an exact Poisson test and then combined into a single P value for each gene using Fisher’s method. This method is similar to an approach called CEDER, which uses information from each exon of a eukaryotic gene to determine if there is consistent differential expression in RNA transcriptome sequencing (RNA-Seq) data for a given gene (19). Therefore, we call this P value a CEDER P value. Genes were sorted based on their fitness factor (total insertion counts in the inoculum/total insertion counts in the lung) and their CEDER P value (see Data Set S3). Inspection of transposon numbers and counts in individual genes suggested that the CEDER P value could be used as a selection criterion for true-positive results of identification of fitness genes by InSeq. For example, ilvD, required for branched-chain amino acid (BCAA) synthesis, had a P value of 0 based on 18 insertions in the input and 13 in the lung with 11 shared insertions having an average fitness factor of 192 (Fig. 1B). In contrast, VK055_468, a putative transcriptional regulatory gene, had a CEDER P value of 2e−14 based on a 186-fold fitness factor but only one insertion (Fig. 1C).
To confirm fitness defects and test the predictive power of CEDER, seven potential fitness genes identified by InSeq analysis were selected for experimental validation by coinfection. Six genes (rfaH, ilvC, ilvD, copA, aroE, and VK055_4417) with a CEDER P value of 0 and a fitness factor of >10 were tested compared to VK055_468 (P = 2e−14; fitness factor of 168). These genes were mutated using Lambda Red recombinase to replace the entire coding sequence with a kanamycin resistance gene (15). Mice were then infected as described in Materials and Methods, but with a 1:1 ratio of the wild type (WT) and each mutant. Based on the bacterial density of each strain after 24 h, the competitive index was calculated. All six mutants with a CEDER P value of 0 had a fitness defect in the lung compared to the wild type (Fig. 2), ranging from 1.4-fold (VK055_4417) to 11,694-fold (rfaH). In contrast, VK055_468 did not have a fitness defect compared to the wild type (Fig. 2). Therefore, genes with a CEDER P value of >0 were not characterized further. In total, 1272 genes had a P value of 0 (top 25 shown in Table 1), including 69 genes with a fitness factor of >10 and 333 genes with a fitness factor of >2. There were also 452 genes with a fitness factor of <0.5 and 97 genes with a fitness factor of <0.1, suggesting that insertion mutants increase lung fitness. Since the goal of the study was to identify mutants with decreased lung fitness, mutants overrepresented in the lung were not pursued further.
FIG 2 .

Competitive infection against the wild-type strain confirms the fitness defect of mutants selected by InSeq. Insertion-deletion mutants of six genes selected and one gene not selected by CEDER analysis (VK055_468) of InSeq fitness were constructed using Lambda Red recombinase and used to coinfect mouse lungs with the wild type (WT), or WT carrying pACYC184 where noted, at a 1:1 ratio in a total inoculum of 1 × 106 CFU. Mean competitive index (log10) for each mutant compared to the wild type at 24 h is shown. *, P < 0.05; **, P < 0.01; ***, P < 0.001, by one-sample t test on log10-transformed data (≥4 mice per group); ###, P < 0.001 by two-sample t test.
TABLE 1 .
Top 25 K. pneumoniae KPPR1 lung fitness genesa
| Locus ID (VK055_x) | Gene | Product | Ratio (input/lung) |
|---|---|---|---|
| 5014 | Capsule assembly Wzi family protein | 2820.95 | |
| 3141 | rfaH | Transcriptional activator | 2696.77 |
| 5096 | Hypothetical protein | 2486.08 | |
| 3202 | ilvC | Ketol-acid reductoisomerase | 2248.61 |
| 3832 | argR | Arginine repressor | 1140.74 |
| 5025 | Undecaprenyl-phosphate glucose phosphotransferase | 840.55 | |
| 5012 | galF | Regulatory protein | 704.99 |
| 3206 | ilvE | Branched-chain amino acid aminotransferase | 581.10 |
| 3515 | Rhodanese-like domain protein | 508.09 | |
| 4417 | MarR family protein | 442.14 | |
| 4811 | purF | Amidophosphoribosyltransferase | 378.16 |
| 4619 | purL | Phosphoribosylformylglycinamidine synthase | 312.03 |
| 1194 | trpD | Anthranilate synthase component II | 301.31 |
| 4135 | serA | d-3-Phosphoglycerate dehydrogenase; provisional | 248.61 |
| 2495 | leuC | 3-Isopropylmalate isomerase subunit, dehydratase component | 246.77 |
| 3142 | tatC | Twin arginine-targeting protein translocase | 219.96 |
| 3205 | ilvD | Dihydroxy acid dehydratase | 203.61 |
| 5023 | Hypothetical protein | 172.11 | |
| 4883 | rcsB | Transcriptional regulator | 126.39 |
| 2215 | phoR | Phosphate regulon sensor kinase | 110.92 |
| 4579 | pheA | Bifunctional chorismate mutase/prephenate dehydratase | 94.55 |
| 3368 | 2-Oxo-3-deoxygalactonate 6-phosphate aldolase | 90.01 | |
| 2084 | copA | Copper-translocating P-type ATPase | 69.70 |
| 3086 | purH | Phosphoribosylaminoimidazole carboxamide formyltransferase | 66.84 |
| 3791 | aroE | Dehydroshikimate reductase | 61.93 |
Experimentally validated genes are shown in boldface.
rfaH is required for serum resistance.
RfaH is an antiterminator that specifically promotes the transcription of long virulence operons, including capsule and lipopolysaccharide (LPS) synthesis genes, in E. coli and Salmonella enterica serotype Typhimurium (20–22). In KPPR1, an rfaH mutant is severely attenuated in the lung (>10,000-fold compared to wild type), and the fitness of the mutant is significantly restored by complementation with the rfaH gene ligated into pACYC184 (Fig. 2). This mutant forms a small colony compared to the wild type, suggesting a defect in capsule biosynthesis. Indeed, India ink staining of bacteria delineated copious capsule in KPPR1 but no visible capsule in the rfaH mutant (Fig. 3A and B). To assess mucoviscosity, a phenotype associated with hypervirulent capsular serotypes in K. pneumoniae clinical isolates, low-speed centrifugation was performed and the optical density of the supernatant was measured (23, 24). Whereas the wild-type suspension remained turbid, the rfaH mutant pelleted readily, and complementation restored mucoviscosity significantly (Fig. 3D). Both capsule and O antigen of LPS can contribute to resistance to complement-mediated killing (25), which has been correlated with the ability to cause patient infections (1). Incubation of the rfaH mutant in serum demonstrated a >1,000-fold loss in viability, greater than that for the susceptible control strain KP297, whereas there was no loss of viability in the wild type (Fig. 4). Complementation restored serum resistance. In RPMI medium with 10% heat-inactivated serum, complement is disabled and K. pneumoniae grows robustly but requires the siderophore enterobactin or its glycosylated derivative salmochelin to acquire iron; yersiniabactin does not support growth under this condition (5, 26). Whereas the wild type replicates robustly, the rfaH mutant had a moderate growth defect comparable to the loss of salmochelin expression (iroA ybtS mutant) but not as severe as the siderophore null entB ybtS mutant (Fig. 5). In contrast, the rfaH mutant grew well in RPMI medium without added serum (see Fig. S2A in the supplemental material). Together, these data indicate that rfaH is required for wild-type capsule production, to resist complement-mediated serum killing, and for maximal growth in iron-limited serum.
FIG 3 .

The rfaH mutant is deficient in extracellular capsule. (A to C) India ink staining of K. pneumoniae KPPR1 (WT) (A) and isogenic rfaH (B) and aroE (C) mutants after overnight culture in LB broth is shown by phase-contrast microscopy, in which the presence of capsule is indicated by an area of negative staining around the bacteria. Magnification, ×1,000. (D) Mucoviscosity, as measured by optical density (OD600) after centrifugation for 5 min at 1,000 × g with a starting turbidity of 1.0, is shown as the mean and standard deviation from 6 biological replicates. *, P < 0.0001 by ANOVA with Fisher’s LSD test compared to the wild type; #, P < 0.0001 compared to both WT carrying pACYC and the rfaH mutant carrying pRfaH.
FIG 4 .

The K. pneumoniae rfaH and aroE mutants are susceptible to serum killing. Viability of K. pneumoniae KPPR1 (WT) and isogenic mutants, without (A) or containing pACYC184 with or without rfaH (B), after a 3-h incubation in human serum is shown as mean log10 CFU/ml ± standard deviation from at least two replicates per group. *, P < 0.0001 by ANOVA with Fisher’s LSD test compared to the wild type.
FIG 5 .

The K. pneumoniae rfaH mutant has a mild growth defect in heat-inactivated serum. K. pneumoniae KPPR1 (WT) and isogenic mutant growth (log10 CFU/ml) after overnight incubation of a 1 × 103-CFU/ml inoculum in RPMI medium supplemented with 10% heat-inactivated serum is shown as the mean ± standard deviation from 4 independent experiments. *, P < 0.05; **, P < 0.01; ****, P < 0.0001, by ANOVA with Fisher’s LSD test.
K. pneumoniae requires branched-chain amino acid synthesis for full virulence.
The ilv locus encodes enzymes for synthesis of the branched-chain amino acids isoleucine and valine and the precursors for leucine (27). ilvC and ilvD mutants had 100- and 66-fold defects in competition with the wild type (Fig. 2), respectively, indicating that branched-chain amino acid synthesis is required for fitness in the lung. To confirm that the ilvC and ilvD mutants are auxotrophs for branched-chain amino acids, their growth in minimal medium was compared to that of the wild type (Fig. 6). Neither mutant grew significantly, but growth of both was restored by supplementation of the medium with the branched-chain amino acids isoleucine and valine at 10 mM concentrations. These data indicate that synthesis of branched-chain amino acids is required for lung fitness.
FIG 6 .

K. pneumoniae ilvC and ilvD mutants are unable to replicate in minimal medium without branched-chain amino acids. Optical density (OD600) of K. pneumoniae KPPR1 (WT) and ilvC and ilvD mutants in M9 minimal medium with or without 10 mM isoleucine (I) and valine (V) is shown as the mean from triplicate samples representative of 3 independent experiments.
aroE is required for serum resistance.
Disruption of the aroE gene, which encodes shikimate dehydrogenase required for aromatic amino acid synthesis, causes a 13-fold decrease in fitness in competitive infection (Fig. 2). Unlike for ilvC and ilvD mutants, the aroE mutant is not defective for growth in minimal medium (see Fig. S2B in the supplemental material), perhaps due to compensation by the ydiB-encoded quinate/shikimate dehydrogenase (10, 28). However, the aroE mutant does have a moderate defect in serum survival (Fig. 4) and a defect in mucoviscosity (Fig. 3D), despite production of capsule as measured by India ink (Fig. 3C). These data indicate that aroE is dispensable for bacterial growth in vitro but is required to evade complement-mediated serum killing.
copA is required to prevent copper toxicity.
Mutation of the copA gene, encoding a P-type ATPase copper efflux pump (29), causes a 37-fold fitness defect in competitive infection with the wild type (Fig. 2). To determine whether deletion of copA increases susceptibility to copper toxicity, as seen in E. coli, the wild type and copA mutant were incubated in M9 minimal medium with increasing amounts of copper (5 to 25 µM). We found that CFU are significantly lower in copA mutants on average (P = 0.02) and the rate of decreasing CFU with increasing Cu concentration is significantly higher in a copA mutant (P = 0.0003) than in the wild type (WT) (Fig. 7A). Complementation with a plasmid-expressed copy of copA restored copper resistance to the mutant (Fig. 7B). In E. coli, copper toxicity occurs through inactivation of dehydratases such as IlvD in the branched-chain synthesis pathway (30). The ilvD mutant was more resistant to copper than the copA mutant, suggesting additional cellular targets of copper toxicity (Fig. 7C). However, the addition of branched-chain amino acids significantly protected the copA mutant (Fig. 7C). These data indicate that copper resistance and branched-chain amino acid metabolism pathways may overlap in their contribution to K. pneumoniae lung fitness.
FIG 7 .
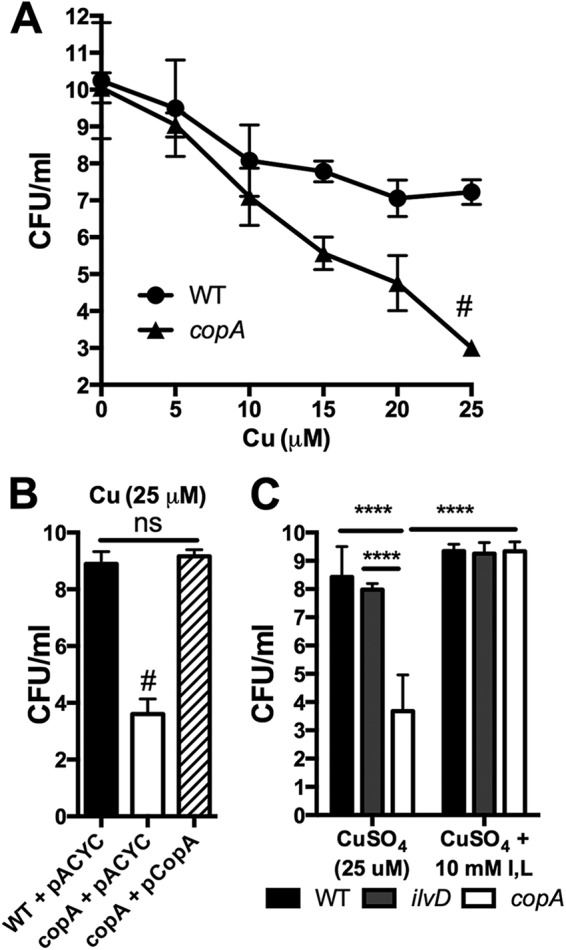
The K. pneumoniae copA mutant has increased sensitivity to copper. (A) Viability of a 1 × 108-CFU inoculum of K. pneumoniae WT and copA mutant after an 18-h incubation in M9 broth with concentrations of cupric sulfate indicated is shown as mean ± standard deviation of results from at least 3 independent experiments. Based on a linear regression model (modeling log-transformed CFU as a function of the Cu concentration, group [WT or copA], and interaction between concentration and group), the copA mutant has a significantly faster-decreasing rate over concentrations than the wild type (#). (B) Viability of K. pneumoniae harboring pACYC184 (pACYC) or pACYC184 with copA (pCopA) after incubation in 25 µM CuSO4 is shown as the mean ± standard deviation from three independent experiments. #, P < 0.0001 compared to both strains by ANOVA with Fisher’s LSD test; ns, not significant. (C) Viability of K. pneumoniae mutants as indicated in 25 µM CuSO4 with or without 10 mM isoleucine and valine is shown as the mean ± standard deviation from at least three independent experiments. ****, P < 0.0001 by ANOVA with Fisher’s LSD.
InSeq identifies conserved genes as critical fitness factors for K. pneumoniae lung infection.
To define broad pathways that may contribute to K. pneumoniae fitness in the lung, gene enrichment analysis was performed. The KEGG classification of the 333 genes in which mutants were >2-fold underrepresented in the lung was determined, and the representation of each class was compared to the total gene set in the KPPR1 genome as annotated based on KEGG assignment. The pathways for 2-oxocarboxylic acid metabolism; valine, leucine, and isoleucine biosynthesis; and biosynthesis of amino acids were significantly overrepresented among genes identified as fitness factors by InSeq (corrected P values of <0.01), indicating the critical importance of branched-chain amino acid synthesis during lung infection.
The products of conserved fitness genes could serve as targets for novel therapeutics against K. pneumoniae. To identify conserved genes in other clinically significant isolates, the KPPR1 genome was compared to the following reference sequences: MGH78578, an isolate from a patient with pneumonia; NTUH-K2044, an isolate that caused a PLA; NJST258_1, a carbapenem-resistant (KPC) ST258 isolate; and Kp342, a plant endophyte. All combinations of shared, full-length chromosomal genes were analyzed, and overlapping gene sets were tabulated (Fig. 8A; see also Data Set S3 in the supplemental material). KPPR1 shares a conserved set of 3,865 genes (quadrant X; 76% of predicted protein-encoding genes) with all strains; 121 genes with human isolates MGH78578, NTUH-K2044, and NJST258_1 but not the plant endophyte Kp342 (quadrant XIV); and 20, 89, 40, and 27 genes exclusively with MGH78578 (quadrant II), NTUH-K2044 (quadrant XV), NJST258_1 (quadrant IV), and Kp342 (quadrant VIII), respectively. To assess the conservation of the top fitness genes identified by InSeq analysis, the 69 genes with a fitness factor of >10 were mapped to the regions of overlap between KPPR1 and 4 reference genomes. In fact, 60 of 68 mapped genes were conserved between all K. pneumoniae isolates (Fig. 8B), and all 6 fitness genes verified by targeted mutation are conserved. These data indicate that InSeq analysis detects lung fitness genes conserved in reference strains causing nosocomial pneumonia, pyogenic liver abscess, and carbapenem-resistant infections.
FIG 8 .

K. pneumoniae conserved genes are fitness factors during pneumonia. (A) The number of genes shared between KPPR1 (orange), pneumonia isolate MGH78578 (green), pyogenic liver abscess isolate NTUH-K2044 (pink), carbapenem-resistant isolate NJST258_1 (blue), and plant endophyte Kp342 (yellow) is shown with the number of genes in each quadrant (labeled by Roman numerals) indicated as calculated by CloVR comparative software. (B) Based on InSeq analysis, 60 of 69 genes with a fitness factor of >10 were conserved in all five isolates (green), 2 were present in human isolates but not Kp342 (yellow), and 2 were shared with hypervirulent NTUH-K2044 (red).
DISCUSSION
By performing InSeq analysis of K. pneumoniae KPPR1, leveraging high-quality genome sequence and a well-established animal model, this study identified over 300 fitness genes important in lung infection. Detection of known virulence genes such as the wzi capsule gene validated this approach. Adaptation of CEDER analysis for ranking of fitness genes, based on the concordance of read count ratios from multiple transposon insertions within genes, provided a reliable list of fitness genes that were experimentally validated. These validation experiments demonstrated that aromatic and branched-chain amino acid synthesis pathways, copper efflux, and global virulence gene regulators such as RfaH are critical for K. pneumoniae to cause pneumonia. The identification of genes such as rfaH, with a profound contribution to lung fitness, indicates the benefit of applying InSeq even when prior forward genetic screens have been performed (6, 7). Enrichment analysis further substantiated amino acid synthesis as central to lung fitness, and comparative genomics indicated that many critical fitness genes are highly conserved in K. pneumoniae. The large set of fitness genes in KPPR1 newly identified by InSeq should accelerate progress in defining the virulence strategies of K. pneumoniae.
Enrichment analysis and experimental validation of InSeq-selected genes indicate that K. pneumoniae must synthesize amino acids that the host cannot. Since essential amino acids are acquired through diet, it does not necessarily follow that these amino acids would be limiting in the lung. However, based on measurements in human subjects, the concentrations of these amino acids are significantly lower in pulmonary epithelial lining fluid than in plasma (31). InSeq identified mutants with decreased fitness in pathways that synthesize the host’s essential amino acids histidine (hisG); isoleucine, valine, and leucine (ilvABCDEN and leuABC); methionine/cysteine (metACBFL); phenylalanine and tryptophan (aroE, pheA, and trpD); and threonine (thrBC) (see Data Set S2 in the supplemental material) (32). This indicates that synthesis of amino acids that are essential to its host is also critical for K. pneumoniae to infect the lung.
Branched-chain amino acid metabolism may be critical for K. pneumoniae lung fitness for two reasons: nutrient limitation and copper toxicity. In contrast to reduced branched-chain amino acid levels, copper is increased in human lungs compared to serum (33). Our data show that K. pneumoniae must synthesize branched-chain amino acids in the lung, but in E. coli, this pathway is blocked by copper through its inhibition of dehydratases such as IlvD (30). In K. pneumoniae, the copA mutant is exquisitely sensitive to copper but is protected by exogenous isoleucine and valine (Fig. 7C), consistent with this mechanism of copper toxicity. Sulfonylurea herbicides that inhibit acetohydroxy acid synthase in the branched-chain amino acid synthesis pathway can protect mice from Pseudomonas aeruginosa and Burkholderia pseudomallei pneumonia, but K. pneumoniae is predicted to contain a resistant isozyme (34). The nine K. pneumoniae synthesis genes identified by InSeq (ilvABCDEN and leuABC) could provide novel targets to block branched-chain amino acid (BCAA) synthesis.
In contrast to ilv mutants, the aroE mutant is not auxotrophic in minimal medium but has reduced fitness in the lung. In E. coli, the YdiB quinate/shikimate dehydrogenase can perform the same function as AroE, albeit with lower efficiency (28). The fact that the aroE mutant is susceptible to serum killing suggests a specific defect that affects complement resistance. Salmonella enterica aro mutants are also attenuated in vivo and are susceptible to serum killing in addition to their auxotrophy (35). The mechanism for the increased serum sensitivity of the K. pneumoniae aroE mutant is unclear.
In addition to synthesizing building blocks for replication despite nutrient limitations, K. pneumoniae must also evade host defenses to cause disease. Polysaccharide capsule is a critical virulence factor of K. pneumoniae that evades complement-mediated killing and opsonophagocytosis. Detection of the capsular assembly gene wzi as the most significant fitness gene by InSeq analysis reinforces the critical role of capsule. Phenotypic assays indicate that rfaH is important in capsular synthesis and is required for survival in vivo and in serum. S. Typhimurium and uropathogenic E. coli rfaH mutants are also severely attenuated in animal models, corresponding to defects in E. coli capsular synthesis and LPS synthesis in both (20, 36). RfaH appears to be a critical virulence factor that is conserved among K. pneumoniae strains and with other pathogenic Enterobacteriaceae.
Treatment of K. pneumoniae infections has become increasingly challenging due to carbapenem and extended-spectrum β-lactamase resistance (4). Therefore, novel therapeutic approaches are needed to fight these common and life-threatening nosocomial infections. Targets for antimicrobials should be conserved within a species or a group of closely related species at a minimum, have a low potential for toxicity or disruption of human metabolism, and ideally not have a preexisting resistance mechanism. This study identifies 60 genes that are conserved among diverse representatives of K. pneumoniae and have a 10- to 10,000-fold defect when disrupted, and many are in prokaryote-specific functions such as synthesis of amino acids that humans do not make or transcription of polycistronic RNAs (rfaH). Through careful study of their function during infection, some of these gene products could be pursued as targets for novel antimicrobials.
MATERIALS AND METHODS
Bacterial strains and media.
K. pneumoniae KPPR1 (10) and isogenic mutants were cultured in Luria-Bertani (LB) broth at 37°C with shaking or 30°C on agar (Becton, Dickinson and Company, Sparks, MD) supplemented with kanamycin (25 µg/ml), spectinomycin (50 µg/ml), chloramphenicol (20 µg/ml), and rifampin (30 µg/ml) as indicated.
Murine pneumonia model.
Six- to 8-week-old C57BL/6 mice (Jackson Laboratory, Bar Harbor, ME) were anesthetized with isoflurane and inoculated retropharyngeally with 1 × 106 CFU of pooled K. pneumoniae transposon mutants. LB broth-grown cultures were centrifuged, resuspended, and diluted in phosphate-buffered saline (PBS) to a concentration of 2 × 107 CFU, and 50 µl of the suspension was administered. After 24 h, mice were euthanized by CO2 asphyxiation. To determine bacterial density, lungs and spleens were removed, homogenized in 1 ml PBS, and cultured on LB agar with appropriate antibiotics. To determine competitive indices, mice (at least 4 per group) were infected as described above with 50 µl of a 1:1 mixture of wild-type and mutant K. pneumoniae strains in a total inoculum of 1 × 106 CFU. After 24 h, mice were euthanized, lungs were homogenized in 1 ml PBS, wild-type CFU were measured by culture on LB plates supplemented with rifampin, and mutant CFU were measured on LB plates with rifampin and kanamycin. The competitive index was calculated as (mutant lung CFU/wild-type lung CFU)/(mutant inoculum CFU/wild-type inoculum CFU).
Transposon library construction and sequencing.
The transposon library was constructed by conjugation of the pSAM_Cam plasmid, a modified version of pSAM_Ec (37) containing the chloramphenicol acetyltransferase gene from pKD3 (15), into KPPR1 (see Text S1 in the supplemental material for detailed methods). Insertion sequencing (InSeq; Sequence Read Archive ID SRP051394) was performed as previously described (18).
Construction and complementation of mutants.
Lambda Red mutagenesis was performed as previously described (15). To make electrocompetent cells, K. pneumoniae containing the pKD46 plasmid (15), modified to encode spectinomycin resistance (a gift from Chris Alteri), was cultured overnight in LB broth containing spectinomycin at 30°C with shaking. The following day, cultures were diluted 1:100 in LB broth with spectinomycin and 50 mM l-arabinose and cultured at 30°C until reaching a reading of optical density at 600 nm (OD600) of 0.5 to 0.6 (approximately 4 h). Cultures were placed on ice for at least 30 min and centrifuged in sterile cold bottles at 8,000 × g for 15 min at 4°C. The supernatant was decanted, and bacteria were serially washed and centrifuged in ice-cold sterile volumes of 50 ml 1 mM HEPES at pH 7.4 (Invitrogen, Carlsbad, CA), 50 ml distilled water (dH2O), and 20 ml 10% glycerol. Pellets were resuspended in ice-cold sterile 10% glycerol at a final density of 2 × 1010 to 3 × 1010 CFU/ml and stored at −80°C in 50-µl aliquots.
To generate null mutants, E. coli BW25141 containing the pKD4 plasmid was cultured overnight in LB broth containing 50 µg/ml kanamycin (MP Biomedicals, Santa Ana, CA) at 30°C. The pKD4 plasmid was isolated using a Spin Miniprep kit (Qiagen, Valencia, CA). Oligonucleotide primers were then designed with 60-bp homology flanking the region targeted for deletion added to 5′ ends of P1 and P2 sites of the pKD4 kanamycin resistance cassette (see Table S2 in the supplemental material). The targeting fragment was generated by PCR consisting of 95°C for 5 min; 30 cycles of 95°C for 1 min, 58°C for 1 min, and 72°C for 1 min; and 72°C for 5 min. PCR products were pooled and purified using a Qiagen PCR purification kit. PCR products were digested overnight at 37°C with DpnI, and 10 µl was added to the electrocompetent pKD46 cells, gently mixed, and incubated on ice for 10 min. The mixture was electroporated using a 0.1-cm-gap cuvette (Fisher Scientific; catalog no. FB101) at 1.8 kV, 400 Ω, 25 µF, with a Bio-Rad Micropulser (Bio-Rad, Hercules, CA), and cells were recovered with SOC medium and incubated overnight at 30°C with shaking in sterile culture tubes. Next, cells were spun down, resuspended in LB broth, plated onto LB broth plates containing kanamycin, and incubated at 37°C overnight. Transformants were restreaked on LB agar and confirmed by colony PCR using flanking primers (see Table S2).
To complement the rfaH and copA mutants, PCR products containing the open reading frame and upstream sequence (see Table S2 in the supplemental material) were inserted into pCR 2.1 by TOPO TA cloning (Life Technologies, Carlsbad, CA) and transferred to pACYC184 by ligation after digestion with Xbal and HindIII. The resulting complementation plasmid or pACYC184 alone was introduced into WT or mutant K. pneumoniae by electroporation.
Serum growth assay.
RPMI medium (Invitrogen, Grand Island, NY), supplemented with 10% (vol/vol) heat-inactivated pooled human serum, was inoculated with 1 × 103 CFU/ml of an overnight culture of K. pneumoniae and incubated overnight in a final volume of 100 µl in 96-well plates at 37°C with 5% CO2. To determine bacterial density, samples were serially diluted and plated on LB agar (Thermo Fisher Scientific, Waltham, MA) with appropriate antibiotics.
Growth curves.
K. pneumoniae strains were cultured overnight in LB broth. On the following day, cultures were incubated in LB or M9 (Invitrogen) medium with or without isoleucine and valine supplementation (Sigma-Aldrich, St. Louis, MO) at a starting density of 2.6 × 107 CFU/ml and cultured for 8 h at 37°C. Absorbance readings at 600 nm were taken every 15 min using an Eon microplate spectrophotometer with Gen5 software (BioTek, Winooski, VT).
Capsule staining.
K. pneumoniae strains were cultured overnight in LB broth. On the following day, 10 µl of culture was added to slides and allowed to air dry. The dried bacteria were then heat fixed to the slide (3 min at 56°C); 10 µl of India ink (Becton, Dickinson and Company, Sparks, MD) diluted 1:3 in PBS was added and wet mounted with a coverslip. Superfrost slides (Thermo Fisher Scientific, Waltham, MA) were viewed and imaged using an Axioplan2 phase-contrast microscope (Zeiss, Irvine, CA) with a SPOT diagnostic camera and software (SPOT Imagine Solutions, Sterling Heights, MI).
Mucoviscosity assay.
Overnight cultures were pelleted by centrifugation at 9,400 × g and resuspended in PBS to an OD600 of ~1. The suspensions were centrifuged for 5 min at 1,000 × g, and the OD600 of the supernatants was measured. Final readings were normalized to the OD600 of the wild type before centrifugation.
Serum killing assay.
K. pneumoniae strains were cultured overnight in LB broth. On the second day, cultures were diluted to 2.5 × 105 CFU/ml in non-heat-inactivated human serum and incubated for 3 h at 37°C. Aliquots at t = 0 and t = 3 h were plated for CFU on LB agar with appropriate antibiotics.
Copper sensitivity assay.
Sensitivity to copper was measured by washing 3 ml of stationary-phase cultures twice with PBS and then centrifuging them at 10,000 × g and resuspending them in M9 minimal medium. Next, 2-ml reaction mixtures were prepared with defined concentrations of copper sulfate (98% pure; Sigma, St. Louis, MO) in M9 medium with bacterial cultures normalized to an OD600 of 0.2. Reaction mixtures were incubated for 18 h at 37°C in six-well plates. Following incubation, CFU were enumerated by quantitative culture.
Genome comparison and gene enrichment analysis.
To identify genes shared between KPPR1 (GenBank identifier [ID] CP009208) and reference K. pneumoniae isolates MGH78578 (GenBank ID CP000647), NTUH-K2044 (GenBank ID AP006725), NJST258_1 (GenBank ID CP006923.1), and Kp342 (GenBank ID CP000964), reference genomes were compared by CloVR comparative software (on the DIAG computing platform), generating clusters of genes shared between genomes. The number of genes shared between each genome was tabulated, and a Venn diagram was generated with free R (v.3.1.1) software and the Venn diagram package. To identify biological pathways associated with lung fitness, enrichment analysis was performed using KOBAS 2.0 (38).
Statistical analysis.
To quantify the significance of each gene as a potential fitness factor, we first tested the difference in counts between inoculum and lung at each transposon insertion using an exact Poisson test and then combined the insertion-level P values into a P value for each gene using Fisher’s method (using R 3.1.1 software) in a modification of CEDER analysis (19). A small P value indicates an important fitness factor. To test whether the copA mutant is more sensitive to copper than the wild type, a linear regression model was used (the relation between log-transformed CFU and Cu concentration is approximately linear). Statistical analysis of CFU data was performed on log-transformed data by a one-sample t test for competitive index and a two-sample t test or analysis of variance (ANOVA) with Fisher’s least significant difference (LSD) test as indicated using Prism 6 (GraphPad Software, La Jolla, CA).
Ethics statement.
This study was performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals (39). The University of Michigan Institutional Animal Care and Use Committee (IACUC) approved this research (PRO00005795).
SUPPLEMENTAL MATERIAL
Supplemental methods. Download
Transposon insertion sites and counts. Download
Fitness ratio and CEDER analysis of K. pneumoniae KPPR1 genes. Download
Gene intersection lists. Download
pSAM_Cam is suicidal in K. pneumoniae, and transposon insertion is random. (a) Relevant features of pSAM_Cam. (b) To verify that pSAM_Cam was suicidal in K. pneumoniae, colony PCR was performed using primers that anneal within the plasmid backbone (top) and primers that anneal within the Kan cassette (bottom). The presence of pSAM_Cam generates a 999-bp PCR product from the plasmid backbone and a 1,338-bp product from the Kan cassette. The presence of the transposon-containing Kan cassette inserted into K. pneumoniae generates a 1,338-bp product. (c) To assess the randomness of the transposon mutant library, genomic DNA from Rifr Kanr colonies was harvested, digested with EcoRI, and subjected to Southern blotting. The variation in sizes of fragments containing the Kan cassette indicates that the transposon is inserted in a different region of the chromosome for each of the mutants tested. Download
The aroE and rfaH mutants do not have growth defects in M9 or RPMI medium. (A) CFU per milliliter after inoculation with 1 × 103 CFU and overnight incubation in RPMI medium is shown as mean ± standard deviation from two independent experiments. (B) Optical density of K. pneumoniae KP4 and isogenic mutants during incubation in M9 minimal medium at 37°C is shown as mean ± standard deviation from triplicate cultures and is representative of three independent experiments. Download
Read counts from InSeq analysis.
Primers used in this study.
ACKNOWLEDGMENTS
This research was supported in part by the University of Michigan Medical School Host Microbiome Initiative. Research using CloVR was conducted on the National Science Foundation funded MRI-R2 project #DBI-0959894.
We thank Victoria Holden for assistance with bacterial plating, Sargurunathan Subashchandrabose for scientific discussions about the project, the University of Michigan DNA Sequencing Core for consultation and Illumina HiSeq sequencing, and the Center for Live Cell Imaging for microscope training and access.
Footnotes
Citation Bachman MA, Breen P, Deornellas V, Mu Q, Zhao L, Wu W, Cavalcoli JD, Mobley HLT. 2015. Genome-wide identification of Klebsiella pneumoniae fitness genes during lung infection. mBio 6(3):e00775-15. doi:10.1128/mBio.00775-15.
REFERENCES
- 1.Podschun R, Ullmann U. 1998. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev 11:589–603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Maroncle N, Balestrino D, Rich C, Forestier C. 2002. Identification of Klebsiella pneumoniae genes involved in intestinal colonization and adhesion using signature-tagged mutagenesis. Infect Immun 70:4729–4734. doi: 10.1128/IAI.70.8.4729-4734.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hsieh PF, Lin TL, Lee CZ, Tsai SF, Wang JT. 2008. Serum-induced iron-acquisition systems and TonB contribute to virulence in Klebsiella pneumoniae causing primary pyogenic liver abscess. J Infect Dis 197:1717–1727. doi: 10.1086/588383. [DOI] [PubMed] [Google Scholar]
- 4.Centers for Disease Control and Prevention 2013. Antibiotic resistance threats in the United States. Centers for Disease Control and Prevention, Atlanta, GA. [Google Scholar]
- 5.Bachman MA, Oyler JE, Burns SH, Caza M, Lépine F, Dozois CM, Weiser JN. 2011. Klebsiella pneumoniae yersiniabactin promotes respiratory tract infection through evasion of lipocalin 2. Infect Immun 79:3309–3316. doi: 10.1128/IAI.05114-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lawlor MS, Hsu J, Rick PD, Miller VL. 2005. Identification of Klebsiella pneumoniae virulence determinants using an intranasal infection model. Mol Microbiol 58:1054–1073. doi: 10.1111/j.1365-2958.2005.04918.x. [DOI] [PubMed] [Google Scholar]
- 7.Struve C, Forestier C, Krogfelt KA. 2003. Application of a novel multi-screening signature-tagged mutagenesis assay for identification of Klebsiella pneumoniae genes essential in colonization and infection. Microbiology 149:167–176. doi: 10.1099/mic.0.25833-0. [DOI] [PubMed] [Google Scholar]
- 8.Langridge GC, Phan M-D, Turner DJ, Perkins TT, Parts L, Haase J, Charles I, Maskell DJ, Peters SE, Dougan G, Wain J, Parkhill J, Turner AK. 2009. Simultaneous assay of every Salmonella typhi gene using one million transposon mutants. Genome Res 19:2308–2316. doi: 10.1101/gr.097097.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Goodman AL, McNulty NP, Zhao Y, Leip D, Mitra RD, Lozupone CA, Knight R, Gordon JI. 2009. Identifying genetic determinants needed to establish a human gut symbiont in its habitat. Cell Host Microbe 6:279–289. doi: 10.1016/j.chom.2009.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Broberg CA, Wu W, Cavalcoli JD, Miller VL, Bachman MA. 2014. Complete genome sequence of Klebsiella pneumoniae strain ATCC 43816 KPPR1, a rifampin-resistant mutant commonly used in animal, genetic, and molecular biology studies. Genome Announc 2(5):e00924-14. doi: 10.1128/genomeA.00924-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wu KM, Li LH, Yan JJ, Tsao N, Liao TL, Tsai HC, Fung CP, Chen HJ, Liu YM, Wang JT, Fang CT, Chang SC, Shu HY, Liu TT, Chen YT, Shiau YR, Lauderdale TL, Su IJ, Kirby R, Tsai SF. 2009. Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis. J Bacteriol 191:4492–4501. doi: 10.1128/JB.00315-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.DeLeo FR, Chen L, Porcella SF, Martens CA, Kobayashi SD, Porter AR, Chavda KD, Jacobs MR, Mathema B, Olsen RJ, Bonomo RA, Musser JM, Kreiswirth BN. 2014. Molecular dissection of the evolution of carbapenem-resistant multilocus sequence type 258 Klebsiella pneumoniae. Proc Natl Acad Sci U S A 111:4988–4993. doi: 10.1073/pnas.1321364111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fouts DE, Tyler HL, Deboy RT, Daugherty S, Ren Q, Badger JH, Durkin AS, Huot H, Shrivastava S, Kothari S, Dodson RJ, Mohamoud Y, Khouri H, Roesch LF, Krogfelt KA, Struve C, Triplett EW, Methé BA. 2008. Complete genome sequence of the N2-fixing broad host range endophyte Klebsiella pneumoniae 342 and virulence predictions verified in mice. PLoS Genet 4:e1000141. doi: 10.1371/journal.pgen.1000141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wei D, Wang M, Shi J, Hao J. 2012. Red recombinase assisted gene replacement in Klebsiella pneumoniae. J Ind Microbiol Biotechnol 39:1219–1226. doi: 10.1007/s10295-012-1117-x. [DOI] [PubMed] [Google Scholar]
- 15.Datsenko KA, Wanner BL. 2000. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A 97:6640–6645. doi: 10.1073/pnas.120163297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Huang T-W, Lam I, Chang H-Y, Tsai SF, Palsson BO, Charusanti P. 2014. Capsule deletion via a λ-red knockout system perturbs biofilm formation and fimbriae expression in Klebsiella pneumoniae MGH 78578. BMC Res Notes 7:13. doi: 10.1186/1756-0500-7-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Clarke L, Carbon J. 1976. A colony bank containing synthetic Col El hybrid plasmids representative of the entire E. coli genome. Cell 9:91–99. doi: 10.1016/0092-8674(76)90055-6. [DOI] [PubMed] [Google Scholar]
- 18.Goodman AL, Wu M, Gordon JI. 2011. Identifying microbial fitness determinants by insertion sequencing using genome-wide transposon mutant libraries. Nat Protoc 6:1969–1980. doi: 10.1038/nprot.2011.417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wan L, Sun F. 2012. CEDER: accurate detection of differentially expressed genes by combining significance of exons using RNA-Seq. IEEE/ACM Trans Comput Biol Bioinform 9:1281–1292. doi: 10.1109/TCBB.2012.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nagy G, Danino V, Dobrindt U, Pallen M, Chaudhuri R, Emödy L, Hinton JC, Hacker J. 2006. Down-regulation of key virulence factors makes the Salmonella enterica serovar Typhimurium rfaH mutant a promising live-attenuated vaccine candidate. Infect Immun 74:5914–5925. doi: 10.1128/IAI.00619-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gao Q, Xu H, Wang X, Zhang D, Ye Z, Gao S, Liu X. 2013. RfaH promotes the ability of the avian pathogenic Escherichia coli O2 strain E058 to cause avian colibacillosis. J Bacteriol 195:2474–2480. doi: 10.1128/JB.02074-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Burmann BM, Knauer SH, Sevostyanova A, Schweimer K, Mooney RA, Landick R, Artsimovitch I, Rösch P. 2012. An α helix to β barrel domain switch transforms the transcription factor RfaH into a translation factor. Cell 150:291–303. doi: 10.1016/j.cell.2012.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lin T-L, Yang F-L, Yang A-S, Peng H-P, Li T-L, Tsai M-D, Wu S-H, Wang J-T. 2012. Amino acid substitutions of MagA in Klebsiella pneumoniae affect the biosynthesis of the capsular polysaccharide. PLoS One 7:e46783. doi: 10.1371/journal.pone.0046783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fang C-T, Chuang YP, Shun CT, Chang SC, Wang JT. 2004. A novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J Exp Med 199:697–705. doi: 10.1084/jem.20030857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Merino S, Camprubí S, Albertí S, Benedí VJ, Tomás JM. 1992. Mechanisms of Klebsiella pneumoniae resistance to complement-mediated killing. Infect Immun 60:2529–2535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bachman MA, Miller VL, Weiser JN. 2009. Mucosal lipocalin 2 has pro-inflammatory and iron-sequestering effects in response to bacterial enterobactin. PLoS Pathog 5:e1000622. doi: 10.1371/journal.ppat.1000622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Subashchandrabose S, LeVeque RM, Wagner TK, Kirkwood RN, Kiupel M, Mulks MH. 2009. Branched-chain amino acids are required for the survival and virulence of Actinobacillus pleuropneumoniae in swine. Infect Immun 77:4925–4933. doi: 10.1128/IAI.00671-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Michel G, Roszak AW, Sauvé V, Maclean J, Matte A, Coggins JR, Cygler M, Lapthorn AJ. 2003. Structures of shikimate dehydrogenase AroE and its paralog YdiB. A common structural framework for different activities. J Biol Chem 278:19463–19472. doi: 10.1074/jbc.M300794200. [DOI] [PubMed] [Google Scholar]
- 29.Rensing C, Fan B, Sharma R, Mitra B, Rosen BP. 2000. CopA: an Escherichia coli Cu(I)-translocating P-type ATPase. Proc Natl Acad Sci U S A 97:652–656. doi: 10.1073/pnas.97.2.652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Macomber L, Imlay JA. 2009. The iron-sulfur clusters of dehydratases are primary intracellular targets of copper toxicity. Proc Natl Acad Sci U S A 106:8344–8349. doi: 10.1073/pnas.0812808106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hofford JM, Milakofsky L, Pell S, Fish JE, Peters SP, Pollice M, Vogel WH. 1997. Levels of amino acids and related compounds in bronchoalveolar lavage fluids of asthmatic patients. Am J Respir Crit Care Med 155:432–435. doi: 10.1164/ajrccm.155.2.9032174. [DOI] [PubMed] [Google Scholar]
- 32.John AM, Bell JM. 1976. Amino acid requirements of the growing mouse. J Nutr 106:1361–1367. [DOI] [PubMed] [Google Scholar]
- 33.Shafeeq S, Yesilkaya H, Kloosterman TG, Narayanan G, Wandel M, Andrew PW, Kuipers OP, Morrissey JA. 2011. The cop operon is required for copper homeostasis and contributes to virulence in Streptococcus pneumoniae. Mol Microbiol 81:1255–1270. doi: 10.1111/j.1365-2958.2011.07758.x. [DOI] [PubMed] [Google Scholar]
- 34.Kreisberg JF, Ong NT, Krishna A, Joseph TL, Wang J, Ong C, Ooi HA, Sung JC, Siew CC, Chang GC, Biot F, Cuccui J, Wren BW, Chan J, Sivalingam SP, Zhang LH, Verma C, Tan P. 2013. Growth inhibition of pathogenic bacteria by sulfonylurea herbicides. Antimicrob Agents Chemother 57:1513–1517. doi: 10.1128/AAC.02327-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sebkova A, Karasova D, Crhanova M, Budinska E, Rychlik I. 2008. Aro mutations in Salmonella enterica cause defects in cell wall and outer membrane integrity. J Bacteriol 190:3155–3160. doi: 10.1128/JB.00053-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nagy G, Dobrindt U, Schneider G, Khan AS, Hacker J, Emödy L. 2002. Loss of regulatory protein RfaH attenuates virulence of uropathogenic Escherichia coli. Infect Immun 70:4406–4413. doi: 10.1128/IAI.70.8.4406-4413.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wiles TJ, Norton JP, Russell CW, Dalley BK, Fischer KF, Mulvey MA. 2013. Combining quantitative genetic footprinting and trait enrichment analysis to identify fitness determinants of a bacterial pathogen. PLoS Genet 9:e1003716. doi: 10.1371/journal.pgen.1003716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Xie C, Mao X, Huang J, Ding Y, Wu J, Dong S, Kong L, Gao G, Li CY, Wei L. 2011. KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res 39:W316–W322. doi: 10.1093/nar/gkr483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.National Research Council 2011. Guide for the care and use of laboratory animals, 8th ed. National Academies Press, Washington, DC. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental methods. Download
Transposon insertion sites and counts. Download
Fitness ratio and CEDER analysis of K. pneumoniae KPPR1 genes. Download
Gene intersection lists. Download
pSAM_Cam is suicidal in K. pneumoniae, and transposon insertion is random. (a) Relevant features of pSAM_Cam. (b) To verify that pSAM_Cam was suicidal in K. pneumoniae, colony PCR was performed using primers that anneal within the plasmid backbone (top) and primers that anneal within the Kan cassette (bottom). The presence of pSAM_Cam generates a 999-bp PCR product from the plasmid backbone and a 1,338-bp product from the Kan cassette. The presence of the transposon-containing Kan cassette inserted into K. pneumoniae generates a 1,338-bp product. (c) To assess the randomness of the transposon mutant library, genomic DNA from Rifr Kanr colonies was harvested, digested with EcoRI, and subjected to Southern blotting. The variation in sizes of fragments containing the Kan cassette indicates that the transposon is inserted in a different region of the chromosome for each of the mutants tested. Download
The aroE and rfaH mutants do not have growth defects in M9 or RPMI medium. (A) CFU per milliliter after inoculation with 1 × 103 CFU and overnight incubation in RPMI medium is shown as mean ± standard deviation from two independent experiments. (B) Optical density of K. pneumoniae KP4 and isogenic mutants during incubation in M9 minimal medium at 37°C is shown as mean ± standard deviation from triplicate cultures and is representative of three independent experiments. Download
Read counts from InSeq analysis.
Primers used in this study.