

Abstract
Secondary phloem and xylem tissues are produced through the activity of vascular cambium, the cylindrical secondary meristem which arises among the primary plant tissues. Most dicotyledonous species undergo secondary development, among them Arabidopsis. Despite its small size and herbaceous nature, Arabidopsis displays prominent secondary growth in several organs, including the root, hypocotyl and shoot. Together with the vast genetic resources and molecular research methods available for it, this has made Arabidopsis a versatile and accessible model organism for studying cambial development and wood formation. In this review, we discuss and compare the development and function of the vascular cambium in the Arabidopsis root, hypocotyl, and shoot. We describe the current understanding of the molecular regulation of vascular cambium and compare it to the function of primary meristems. We conclude with a look at the future prospects of cambium research, including opportunities provided by phenotyping and modelling approaches, complemented by studies of natural variation and comparative genetic studies in perennial and woody plant species.
INTRODUCTION
Plant vasculature forms a network of interconnected cells spanning the plant's body in an organized manner, from the root tip immersed deep within the soil to the highest tree-tops. The vascular system of multicellular land plants fulfills two main functions, long distance transport and mechanical support. Xylem cells, with thick secondary cell walls rich in lignin, cellulose and hemicellulose, are mainly responsible for providing support to the plant, as well as bulk transport of water, nutrients and minerals from the root system to the shoot. Phloem mediates the shoot-to-root transport of the autotrophic energy source, photoassimilates, as well as signaling molecules, such as plant hormones and peptides.
In comparison with animals, plants possess an extraordinary ability for post-embryonic growth and development, which occur throughout a plant's life. Plant growth arises from mitotic cell divisions taking place in growth foci called meristems. The earliest (primary) meristems are of embryonic origin, such as the root apical meristem (RAM) and shoot apical meristem (SAM), which contribute to root and shoot elongation, respectively. These meristems produce the primary plant body, including the primary vasculature. The vascular anatomy of Arabidopsis primary roots and shoots differs. In the primary shoot, the vasculature is located in separate collateral vascular bundles with primary xylem towards the pith parenchyma cells (Fig. 1). In roots, the vascular tissue is arranged in a bisymmetric pattern; primary xylem forms a central axis flanked by two poles of primary phloem (Fig. 1). Procambial cells intervene between the primary xylem and phloem in both root and shoot vasculature; at the onset of secondary growth, these begin to divide periclinally (parallel to the plant axis/surface), giving rise to secondary xylem (inwards), secondary phloem (outwards), and a secondary meristem called vascular cambium, which forms a continuous ring in an organ-specific manner (Fig. 1, discussed in detail later). The vascular cambium is responsible for the lateral (secondary) growth of plants, a process which must be carefully regulated in order to ensure holistic development of the plant vasculature.
Figure 1.

Schematic representation of vascular organization in the Arabidopsis inflorescence stem (A–C) and root (D–F).
The hypocotyl corresponds to the upper part of the root (D). Xylem=blue, (pro)cambium=yellow, phloem=red.
Meristematic cells are small, cytoplasmic and undifferentiated. As these cells divide, the outermost cells are pushed away from the meristem, where they cease division, initiate turgor-driven cell expansion and differentiate into specialized cell types. The balance between cell proliferation and differentiation into other cell types is crucial for meristem indeterminacy, and it is evident that both of these aspects of growth are under genetic control. The developmental mechanisms governing cell division and identity in the early Arabidopsis embryo are also present in the postembryonic root meristem to at least some degree (reviewed in De Rybel et al., 2014a). Similarly, researchers have discovered an increasing array of features shared between the regulation of primary and secondary meristems (reviewed by Miyashima et al., 2013; Jouannet et al., 2014). It is an intriguing possibility that throughout plant life, indeterminate growth may be sustained by the same hormones and genes or perhaps by functionally overlapping sets of hormones and genes. Here, we describe recent advances in our knowledge of secondary growth in Arabidopsis.
Secondary vasculature in trees and Arabidopsis
Since the rise of Arabidopsis as a model plant species (Meyerowitz, 1989), the development of secondary growth has been investigated in the Arabidopsis root (Dolan et al., 1993), hypocotyl (Busse and Evert, 1999a; Chaffey et al., 2002) and shoot (inflorescence stem) (Lev-Yadun, 1994). After a period of secondary growth, all three organs establish prominent secondary xylem consisting of water-conducting vessels, xylem fibers and xylem parenchyma cells similar to angiosperm trees. The vascular cell types in the secondary xylem of the Arabidopsis hypocotyl are similar to those in poplar, albeit smaller in size and lacking the radial vascular rays of parenchyma cells which mediate lateral transport within tree xylem (Fig. 2) (Chaffey et al., 2002), although the formation of ray-like cells has been observed in Arabidopsis stems under weight-induced conditions (see below for details) (Mazur and Kurczynska, 2012). Secondary phloem contains sieve-elements and their companion cells, phloem fibers and phloem parenchyma.
Figure 2.
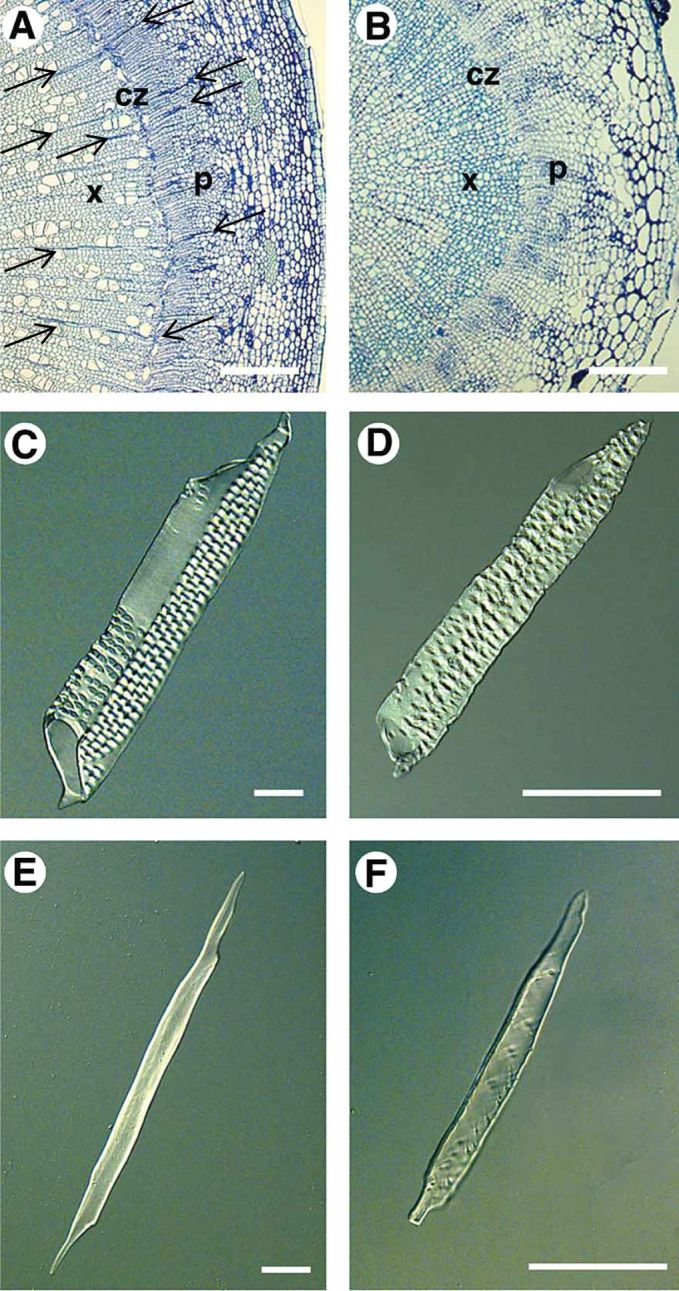
Comparison of the poplar stem and the Arabidopsis hypocotyl.
Cross sections of the poplar stem (A) and Arabidopsis hypocotyl (B) fixed and embedded in methacrylate resin, sectioned at 2 µm and stained with TBO. Maceration of the secondary xylem of poplar (C, E) and Arabidopsis (D, F), showing similarity in the structure of vessel elements (C, D) and fibres (E, F). CZ, cambial zone; P, phloem; X, xylem; vascular rays are marked by arrows. Scale bars equal (A) 450 µm (B) 160 µm, and (C–F) 25 µm. Figure reprinted and modified from Chaffey et al., 2002 with permission. Copyright © 2002 Physiologia Plantarum.
ONTOGENY OF VASCULAR CAMBIUM
Vascular meristems generate cells which differentiate into xylem and phloem. The apical meristems in the shoot and root contain procambium, the primary vascular meristem. Vascular tissue in the primary root and hypocotyl originates from embryonic provascular tissue, whereas shoot vascular tissue, located in vascular bundles, is derived from the shoot apical meristem (Fig. 3). In Arabidopsis and other species which undergo secondary growth, a lateral vascular meristem called cambium develops mainly from the procambium embedded between the differentiated xylem and phloem. In the shoot, the cambium between the vascular bundles arises from parenchyma and endodermis tissues. Consequently, the complete ring of vascular cambium is formed early on in root/hypocotyl, whereas in shoot (inflorescence stem) the formation of a closed cambial circle is a late event, occurring only after initiation of the interfascicular cambium between the vascular bundles.
Figure 3.

The ontogeny of vascular cell types in Arabidopsis.
Embryogenesis gives rise to provascular tissues within the seedling axis and preprocambial strands in the cotyledons. The apical (pro)meristems are also of embryonic origin. Vascular cells in the RAM are collectively called procambium, and they produce primary xylem and phloem/procambium cell lineages. The SAM produces procambium which develops into vascular bundles in the shoot. Activation of procambial cell divisions in the fascicular regions marks the onset of secondary growth in the root/hypocotyl and shoot.
Preprocambium precedes the development of vasculature in the cotyledons and true leaves. Preprocambial cells are parenchymatic cells which develop into a network of elongated procambial cells and further differentiate into functional vascular bundles containing xylem, phloem and cambium (Scarpella and Meijer, 2004). However, usually very little secondary growth takes place in the leaf veins.
Embryogenesis
Primary root anatomy is established during embryogenesis by a tightly regulated developmental program with little variation in terms of cell numbers and patterning (Dolan et al., 1993). Polarity is already established in the first division of the zygote into an apical cell and basal cell, and the provascular tissue is established by the division of four provascular initials during early embryogenesis (Scheres et al., 1994). A computational model recently predicted that the initial geometry of the embryo contributes to vascular patterning (De Rybel et al., 2014b). The cylindrical pattern along the embryo axis is predetermined to form vasculature containing xylem, phloem and (pro)cambium postembryonically in the roots and hypocotyl (Fig. 3). Notably, although embryos do not contain functional xylem or phloem cells (Busse and Evert, 1999b), several genes regulating root/hypocotyl vascular differentiation are already expressed prior to germination. Altogether, these findings indicate that vascular patterning precedes the formation of vasculature during embryo development.
Embryogenesis is hormonally regulated, and auxin, in particular, plays a major role (Friml et al., 2003). MONOPTEROS/AUXIN RESPONSE FACTOR5 (MP/ARF5; At1g19850) is among the earliest markers of provascular tissue in the embryo, and embryos of mp mutants have severely reduced provascular tissue (Hardtke and Berleth, 1998). Several TARGETS OF MONOPTEROS (TMOs) have been identified in the vascular tissues, including the basic helix-loop-helix (bHLH) transcription factor TMO5/bHLH32 (At3g25710) and the DNA-BINDING WITH ONE ZINC FINGER (DOF) family transcription factor TMO6 (At5g60200) (Schlereth et al., 2010). TMO5 interacts with LONESOME HIGHWAY (LHW/bHLH156; At2g27230), another bHLH transcription factor, and together they promote periclinal divisions in the vascular tissue of the embryonic and postembryonic root. Absence of the TMO5 or LHW family members results in a dramatic reduction in periclinal divisions within the vasculature, and therefore in the number of the vascular cell files (Ohashi-Ito and Bergmann, 2007; De Rybel et al., 2013; Ohashi-Ito et al., 2013). Remarkably, 78% of Arabidopsis genes are expressed during the course of embryogenesis, and 55% in the mature embryo (Xiang et al., 2011). In the near future, marker genes identified for each embryonic tissue can be used to create high-resolution transcriptome maps of embryogenesis to study the mechanisms governing the earliest cell-fate decisions in plant life (Palovaara et al., 2013).
Primary root vasculature development
Embryogenesis predefines the tiers of initials surrounding the rarely-dividing quiescent center (QC) cells in the RAM; upon germination, these produce new cells for the elongating primary root cell types (van den Berg et al., 1997). The initials (stem cells) and QC form the stem cell niche. Xylem cell lineages are specified early, deriving directly from the vascular initials, whereas phloem and the intervening procambium lineages are specified higher up in the meristem as a result of asymmetric periclinal divisions (Mähönen et al., 2000). Differentiated primary vascular tissue consists of proto- and metaxylem, proto- and metaphloem, and intervening procambium.
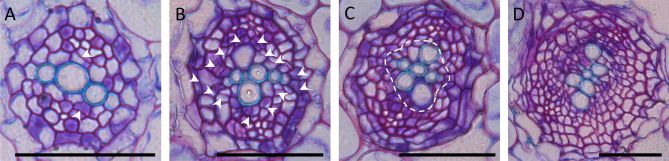
Several factors have been shown to regulate cellular identity and bilateral symmetry within the stele. Periclinal divisions in the stele increase the number of cell files. Whereas the four to five xylem initials touching the QC undergo very few or no periclinal divisions, procambium and phloem initials divide multiple times longitudinally (Mähönen et al., 2000). These periclinal cell divisions are reduced in the wooden leg (wol) mutant, which has a mutation in cytokinin receptor CYTOKININ RESPONSE 1/ ARABIDOPSIS HISTIDINE KINASE 4 (CRE1/AHK4; At2g01830) (Mähönen et al., 2000; Inoue et al., 2001). In wol, all cells within the vasculature differentiate into protoxylem (Scheres et al., 1995; Mähönen et al., 2000). A similar all-protoxylem phenotype can be observed in mutant combinations lacking various components of the cytokinin signaling pathway (Hutchison et al., 2006; Mähönen et al., 2006a; Yokoyama et al., 2007; Argyros et al., 2008), while cytokinin treatments have been shown to inhibit protoxylem formation (Mähönen et al., 2006b). Taken together, these results demonstrate that cytokinins have a dual role during vascular development; they act both as a promoter of periclinal cell divisions and an inhibitor of protoxylem differentiation (Mähönen et al., 2000; Mähönen et al., 2006a; Mähönen et al., 2006b). ARABIDOPSIS HISTIDINE PHOSPHOTRANSFER PROTEIN 6 (AHP6; At1g80100), expressed in protoxylem and adjacent pericycle cells, promotes protoxylem development by negatively regulating cytokinin signaling (Mähönen et al., 2006a). AHP6 expression is auxin-dependent, and loss of AHP6 function results in expansion of the expression domains of cytokinin-response genes ARABIDOPSIS RESPONSE REGULATOR 5 (ARR5; At3g48100) and ARR15 (At1g74890) from the procambium into the protoxylem position (Mähönen et al., 2006a; Bishopp et al., 2011a). Cytokinin signaling, in turn, regulates auxin availability. High cytokinin signalling in the procambium promotes the efflux of auxin from the procambium into the xylem axis by stimulating lateralization of PINFORMED 1 (PIN1; At1g73590) protein and by increasing the expression of the laterally localized PIN7 (At1g23080), and perhaps also PIN3 (At1g70940) (Bishopp et al., 2011a). The mutually inhibitory interaction between cytokinin and auxin in adjacent locations maintains the bisymmetric vascular pattern in the primary root (Bishopp et al., 2011a). Recently, a connection between the auxin-MP-TMO5/LHW and the cytokininAHP6 pathways was identified. The TMO5/LHW dimer promotes the expression of the xylem precursor-specific genes LONELY GUY 3 (LOG3; At2g37210) and LOG4 (At3g53450), which encode enzymes catalyzing the final reaction step of cytokinin biosynthesis (De Rybel et al., 2014b; Ohashi-Ito et al., 2014). Since TMO5/LHW also promotes the expression of AHP6, the protoxylem precursor cells have low cytokinin signaling levels despite being the site of cytokinin synthesis, and therefore display a reduced rate of periclinal cell division. However, cytokinin is able to move from the xylem precursor cells to the neighboring procambial cells where it activates the cytokinin signaling pathway and thus promotes periclinal cell division (Mähönen et al., 2006a; De Rybel et al., 2014b; Ohashi-Ito et al., 2014).
Primary xylem formation is also specified by radial signaling between the stele and endodermis leading to degradation of the HOMEODOMAIN-LEUCINE ZIPPER (HD-ZIP) class III transcription factor mRNAs by microRNA165/166 (miR165/166; encoded by At1g01183, At4g00885, At2g46685, At3g61897, At5g08712, At5g08717, At5g41905, At5g43603 and At5g63715) (Carlsbecker et al., 2010; Miyashima et al., 2011). The GRAS family transcription factor SHORT ROOT (SHR; At4g37650) moves through plasmodesmata from the stele to the adjacent endodermis, where it promotes the expression of another GRAS family transcription factor, SCARECROW(SCR; At3g54220) (Helariutta et al., 2000; Nakajima et al., 2001; Carlsbecker et al., 2010; Vatén et al., 2011; Miyashima et al., 2011). SCR in turn, promotes the expression of microRNA165/166, which moves in the opposite direction, into the stele, to restrict the HD-ZIP III expression domain (Carlsbecker et al., 2010; Miyashima et al., 2011). The HD-ZIP III genes guide xylem development and determine the number of cell files in the stele in a dosage-dependent manner. Loss of all five HD-ZIP III genes results in a complete lack of xylem accompanied by an increased number of procambial cell files, while low levels of HD-ZIP III expression induce protoxylem, and overexpression leads to metaxylem formation (Carlsbecker et al., 2010). AT-HOOK MOTIF NUCLEAR LOCALIZED PROTEIN3 (AHL3; At4g25320) and AHL4 (At5g51590), which also move to the xylem axis from the surrounding tissues, regulate the boundary between xylem and procambium (Zhou et al., 2013). Altogether, the morphogenesis of the root vascular bundle is seen to be regulated by antagonistic networks of laterally moving intercellular signals, including auxin, cytokinins, mobile transcription factors, and microRNAs.
Development of the shoot vascular cambium
In the Arabidopsis inflorescence stem, the vascular cambium develops in two different anatomical regions, within the vascular bundles and between them (Fig. 1 A–C; Fig. 4). Fascicular cambium forms when the procambial cells between the primary xylem and phloem inside the vascular bundles start to divide, whereas interfascicular cambium develops between the bundles. The formation of fascicular cambium precedes the interfascicular cambium; when the latter arises, it completes the circle of shoot vascular cambium.
Figure 4.

The progression of secondary development in the Arabidopsis inflorescence stem.
Anatomy at A) the base of a mature Col stem, B) 5 cm height, C) the base of a 6-cm-long inflorescence stem, and D) the base of a 20 cm long inflorescence stem. Interfascicular cambium marked with an arrow. E–G) The formation of the interfascicular meristem starts when the parenchymous and/or endodermal cells located to the both sides of the fascicular cambial cells begin to divide (arrows). The cell divisions gradually extend towards the middle of the interfascicular region (long double arrows) until the ring of cambial cells divisions is closed, ifr interfascicular region, vb vascular bundle; dotted arrows mark borders between fascicular and interfascicular areas. Scale bar in (A) 500 µm; (B–D) 100 µm; (E and G) 20 µm; (F) 50 µm. Reprinted with permission from Lens et al., 2012 (A–D) and from Mazur et al., 2014 (E–G). Copyright © 2011 Lens et al. and © 2014 Springer-Verlag Wien, respectively.
The interfascicular cambium, and therefore the complete cambial ring, develops only in the most basal region of the inflorescence stem (Lev-Yadun, 1994; Altamura et al., 2001; Little et al., 2002; Sehr et al., 2010; Paul-Victor and Rowe, 2011; Suer et al., 2011; Agusti et al., 2011a; Agusti et al., 2011b). In the upper parts of the stem, only the fascicular cambium is active, and it gives rise to moderate secondary growth inside the vascular bundles. In general, the fascicular cambium is more active in Arabidopsis; it produces more secondary tissues than the interfascicular cambium. The formation of the interfascicular meristem starts when the parenchyma cells (and later, endodermal cells higher up) located on both sides of the fascicular cambial cells begin to divide (Fig. 4). The cell divisions gradually extend towards the middle of the interfascicular region until the ring of cambial cells is closed (Mazur et al., 2014). In the lower parts of the stem, the parenchymous cells located between the vascular bundles and inwards from the interfascicular cambium differentiate into interfascicular fibers (Fig. 4C–D). These fibers, which are not produced through the activity of interfascicular cambium, presumably provide further support for the erect stem by complementing the moderate amount of secondary xylem produced by the interfascicular cambium.
Vascular cambium development in the roots and hypocotyl
During the primary development of the root, the bisymmetric vascular pattern of a central xylem axis flanked by procambial cells and primary phloem poles on both sides is established. Secondary growth initiates with the division of the procambium cells proximal to primary xylem soon after primary development, often even before the secondary cell walls of the metaxylem vessels have matured (Baum et al., 2002). Periclinal cell divisions are initiated in the root procambium approximately five days after germination (Fig. 5 A). A continuous cambial ring is formed as the pericycle cells start dividing as well (Fig. 5 B–C) (Busse and Evert, 1999a). During the transit stage, actively dividing cambium produces secondary xylem inwards and secondary phloem outwards, resulting in a radially symmetric vascular pattern in the root. Secondary growth in the hypocotyl follows the same principles as in the root, since the primary anatomy is similar. Ongoing secondary growth within the root stele forces the non-dividing outer cell layers to peel off. Thereafter, the root's outer protective layer is replaced by a barrier of cork cells (phellem) produced by a lateral meristem called cork cambium (phellogen), which originates from the pericycle. The cork cambium also produces parenchyma (phelloderm) cells inwards. Collectively, these cell types are called periderm.
Figure 5.
Root secondary growth activation in Col-0 roots from 5-days-old (left) to 15-days-old (right).
Periclinal cell divisions in the activating (pro)cambium (A) and in both cambium and pericycle (B) are marked with arrowheads, and the completed vascular cambium cylinder with a dashed line (C). Root diameter is further increased through the cell division activity of the vascular cambium (D). Cross-sections are stained with toluidine blue, scale bar represents 50 µm. (Pictures by Riccardo Siligato).
The hypocotyl has been presented as an interesting model of secondary growth because of the early cessation of elongation followed by activation of cambial cell divisions (Sibout et al., 2008; Ragni and Hardtke, 2014). Lateral growth of the hypocotyl has two distinct phases: phase I, where equal amounts of secondary xylem and phloem are produced, and phase II, called xylem expansion, where more xylem than phloem tissue is produced (Sibout et al., 2008; Ragni et al., 2011). The cell-type composition of the xylem tissue also differs between the two phases; xylem produced during the first phase consists of xylem vessels and parenchyma cells, whereas xylem vessels and fibers form during the second phase. The transition between the phases occurs at the onset of flowering, and gibberellin has been identified as a mobile signal from the shoot to the hypocotyl that triggers this event (Sibout et al., 2008; Ragni et al., 2011). Phase II can also be distinguished during secondary root growth, where, as in the hypocotyl, the secondary xylem produced during late development contains xylem fibers in addition to xylem vessels. Overall, secondary growth in the roots and hypocotyl can be divided into stages of activation, transition, and a radially symmetric pattern, which take place in a developmental gradient in roots (Fig. 5 A–D, also Fig. 11 A) due to ongoing axial growth, which is lacking in hypocotyls.
Figure 11.
Wood anatomy of the roots of selected Brassicaceae species.
A) Secondary growth in the root of Arabidopsis thaliana, Columbia accession. B) The root of the small annual plant Thlaspi perfoliatum. Similarly to secondary development in Arabidopsis, vessels and parenchyma cells are present in the centre of the root, with a ring of lignified tissues at the outer layers. C) The root of the small annual plant, Capsella bursa-pastoris. Vessels are arranged in multiple ring patterns. D) Distinct annual rings with marginal parenchyma and no vascular rays in the stem of the dwarf perennial shrub Ptilotrichum spinosum. E) Large vascular rays in the root of the perennial herb Cardaria draba. F) Dilated rays in the root of the perennial herb, Cardamine alpine. Picture (A) by Marcelo Pace; B–F) reprinted with permission from Schweingruber et al., 2011. Copyright © 2011 Springer-Verlag Berlin Heidelberg.
MOLECULAR REGULATORS OF ROOT AND HYPOCOTYL SECONDARY GROWTH
The plant hormone cytokinin is required for cambial growth in Arabidopsis roots (Matsumoto-Kitano et al., 2008). The quadruple isopentenyltransferase mutant ipt1,3,5,7 (IPT1; At1g68460, IPT3; At3g63110, IPT5; At5g19040, IPT7; At3g23630), which is defective in cytokinin biosynthesis, lacks cambium activation and secondary growth in the root (and shoot, see later section) (Fig. 6). The root phenotype can be rescued by external cytokinin application or by grafting a wild type shoot to act as a cytokinin source for the mutant root (Matsumoto-Kitano et al., 2008). Interestingly, the ipt1,3,5,7 shoot phenotype is also restored by a wild-type root, supporting the idea of systemic cytokinin translocation between root and shoot. Recently, an ATP-cassette binding (ABC) transporter subfamily protein G14 (AtABCG14; At1g31770) required for root-to-shoot translocation of cytokinins was isolated based on an expression pattern similar to IPT3, as well as its transcriptional response to cytokinin (Ko et al., 2014). Systemic rootwards trans-location of cytokinin in phloem (Bishopp et al., 2011b) and shootwards transport in xylem (Ko et al., 2014) may therefore act as an important signal co-ordinating growth in different plant organs.
Figure 6.
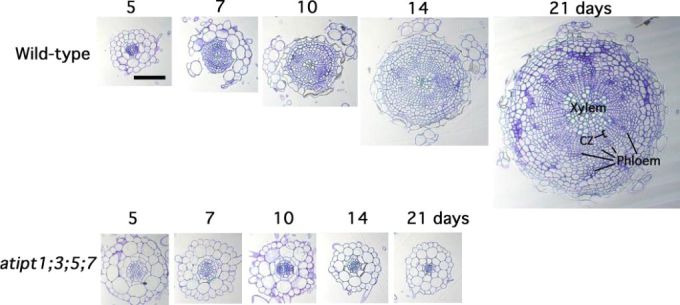
Secondary growth of roots in WT and atipt1;3;5;7.
Typical cross sections of primary roots at the basal region of 5-, 7-, 10-, 14-, and 21-day-old WT (top row) and atipt1;3;5;7 (bottom row) are shown. During thickening growth in WT, the original cortex and epidermis break away. The numbers of days after sowing is indicated. Scale bar: 0.1 mm. Reprinted with permission from Matsumoto-Kitano et al., 2008. Copyright © 2008 by The National Academy of Sciences of the USA.
Vascular development requires an intricate coordination between the genetically separate processes of cambial cell proliferation and differentiation, which has been elegantly shown during the characterization of the TDIF (TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR)-TDR (TDIF RECEPTOR) ligand-receptor pair and its downstream targets. In a Zinnia (Zinnia elegans L.) cell culture system, mesophyll cells transdifferentiate into xylem cells (tracheary elements; TE) when grown in a medium supplemented with auxin and cytokinin (Fukuda, 1997). Furthermore, the premature TEs were found to affect the transdifferentiation of other cells, indicative of cell-to-cell signaling (Motose et al., 2001). The dodecapeptide TDIF was isolated from an auxin-containing culture medium fraction which, when inoculated into a fresh Zinnia culture, was able to strongly inhibit TE differentiation while promoting cell divisions (Ito et al., 2006). The TDIF peptide is endogenously encoded in Arabidopsis by CLAVATA3 (CLV3)/EMBRYO SURROUNDING REGION (CLE) genes CLE41 (At3g24770) and CLE44 (At4g13195), and a close homolog with similar activity is encoded by CLE42 (At2g34925) (Ito et al., 2006). TDIF treatment causes proliferation of procambium in the Arabidopsis hypocotyl and leaf veins, along with reduced xylem differentiation indicated by a discontinuous leaf vein phenotype (Hirakawa et al., 2008). The TDIF receptor TDR/PXY (At5g61480), a subclass XI member of the leucine-rich repeat receptor-like kinase (LRR-RLK) gene family, was identified by a screen for TDIF insensitivity, and TDR was shown to bind specifically to TDIF/CLE41/CLE44 (Hirakawa et al., 2008). The tdr mutant exhibits an occasional loss of procambial cells and therefore the appearance of xylem adjacent to phloem in the hypocotyl (Hirakawa et al., 2008), a phenotype previously described in stems of a mutant named phloem intercalated with xylem (pxy) (Fisher and Turner, 2007), a procambium-expressed LRR-RLK allelic to tdr. Histological analysis revealed CLE41 promoter activity in the phloem and adjacent pericycle cells in roots and hypocotyls, whereas the CLE44 promoter was also active in endodermis (Hirakawa et al., 2008). However, the CLE41/44 expression pattern does not overlap with TDR/PXY expression, which is procambium specific. Together with recognition of TDIF (and to lesser extent CLE42) by the TDIF peptide antibody around the primary phloem and procambium, this has led to a model involving non-cell autonomous (intercellular) signaling between vascular cell types. TDIF is secreted from the phloem and perceived in the procambium by TDR/PXY, which suppresses the differentiation of secondary xylem and promotes cambium identity (Hirakawa et al., 2008). A phloem-derived peptide signal therefore acts as a polarized positional cue to guide the orientation of periclinal cambial cell divisions. Ectopic overexpression of CLE41 and its homolog CLE42 disrupted the periclinal division pattern in hypocotyls, causing drastically altered vascular patterning, including intermixed xylem and phloem domains; by contrast, phloem-localized CLE41 expression increased the vascular cell number while maintaining a normal vascular pattern (Etchells and Turner, 2010). The cle41-1 mutant has reduced stele width in the hypocotyl (Hirakawa et al., 2010).
TDIF/CLE41/CLE44 belongs to the functionally distinguishable B-type CLE peptides, which, unlike the A-type CLE peptides, do not affect primary root growth (Ito et al., 2006; Whitford et al., 2008). Interestingly, type A and B CLE peptides, in particular CLE6 (At2g31085) and CLE41, have synergistic effects in promoting hypocotyl stele size by increasing the cambial cell number and inhibiting xylem and phloem differentiation, which also causes abnormal vascular organization (Whitford et al., 2008). This suggests that peptides other than CLE41 may potentially regulate cambial activity via yet unidentified receptors.
The mediation of intercellular signaling by the TDIF-TDR/PXY ligand-receptor pair is a familiar motif from the SAM, where the peptide encoded by CLV3 (At2g27250) is secreted from stem cells and recognized primarily by the LRR-RLK CLAVATA1 (CLV1; At1g75820), the receptor-like protein CLV2 (At1g65380) and the serine/threonine kinase CORYNE (CRN; At5g 13290) receptor complexes which are expressed in the organizing center. This leads to downregulation of the WUSCHEL (WUS; At2g17950) homeobox transcription factor, a positive regulator of stem cell maintenance (reviewed by Miyashima et al., 2013; Sparks et al., 2013). The WUSCHEL-RELATED HOMEOBOX4 (WOX4; At1g46480) transcription factor was identified as the WOX family member most responsive to the TDIF peptide in Arabidopsis seedlings (Hirakawa et al., 2010). Expression of WOX4 is readily detected in the procambium and cambium, while in one week-old hypocotyls weak expression is also present in the phloem and pericycle (Hirakawa et al., 2010). The TDIF peptide is able to induce discontinuous xylem strands in the wox4 mutant but not in tdr/pxy, indicating that the xylem differentiation pathway is independent of WOX4 (Hirakawa et al., 2010). WOX4 is required for TDIF-induced (pro)cambial proliferation, but overexpression of WOX4 alone was not sufficient to increase cambial cell divisions (Hirakawa et al., 2010). Moreover, another WUSCHEL-homolog, WOX14 (At1g20700), acts redundantly with WOX4 (Etchells et al., 2013).
TDIF-dependent xylem inhibition is redundantly mediated by glycogen synthase kinase 3 proteins (GSK3s), especially BRASSINOSTEROID-INSENSITIVE 2 (BIN2; At4g18710), which interacts with TDR (Kondo et al., 2014). TDR-BIN2 interaction at the plasma membrane increases BIN2 kinase activity, leading to inactivation of the BIN2 substrate, the transcription factor BRI1-EMS SUPPRESSOR 1 (BES1; At1g19350), which is a positive regulator of xylem differentiation (Kondo et al., 2014). The target genes of WOX4 and BES1, the transcription factors regulating cambial cell proliferation and inhibiting secondary xylem differentiation, respectively, are currently unknown.
Mutants affected in secondary growth may be divided into two main categories: i) mutants with increased/decreased cambial proliferation but without changes in the pattern of the secondary vasculature, and ii) mutants with altered vasculature patterning. The ipt1,3,5,7 and wox4 mutants belong to the first category, the tdr/pxy mutant to the latter. Isolated mutants with altered cambial development display quite mild phenotypes in general, which may indicate that several parallel networks and/or functionally homologous genes provide robustness to plant secondary development. The tdr/pxy mutant has increased expression of several ETHYLENE RESPONSE FACTORS (ERFs), which led to the discovery that ethylene signaling acts in parallel to the TDR/PXY signaling pathway in maintaining cambial cell divisions without affecting the vascular pattern in Arabidopsis shoots and hypocotyls (Etchells et al., 2012). The receptor-like kinase ERECTA (ER; At2g26330) has been shown to restrict xylem expansion in phase II of secondary growth in Arabidopsis hypocotyls (Ragni et al., 2011) and to enhance the intercalation of xylem and phloem in pxy stem vascular bundles, suggesting that ER-signaling (discussed in detail in the section about secondary stem growth) interacts with PXY-dependent vascular organization (Etchells et al., 2013). ER and its close homolog ERECTA-LIKE 1 (ERL1; At5g62230) also contribute to the maintenance of (pro)cambium cell layers in the inflorescence stems (Uchida and Tasaka, 2013). In addition, ER and ERL1 regulate stem elongation (Uchida and Tasaka, 2013). PXY has also two homologs, PXY-LIKE 1 (PXL1,At1g08590) and PXL2 (At4g28650), which act at least partially redundantly with PXY (Fisher and Turner, 2007; Etchells et al., 2013).
Altogether, the TDIF-TDR-WOX4 and TDIF-TDR-GSK3s-BES1 pathways are known to interact with other signaling cascades, including plant hormones (Fig. 7). Earlier studies have shown that brassinosteroids promote xylem differentiation (Caño-Delgado et al., 2004). As BIN2 and BES1 are also core components of the brassinosteroid signaling pathway, future studies are needed to link brassinosteroids to the regulation of vascular cambium. Cross-talk between cytokinin and CLE signaling may be a general feature in plant development, as cytokinin signaling in primary roots was enhanced by CLE peptides (Kondo et al., 2011), though this has not yet been explored in the context of vascular cambium. The TDIF and CLE42 peptides are also involved in the formation of axillary branches (Yaginuma et al., 2011) and lateral roots (Cho et al., 2014) (Fig. 7). Furthermore, an LRR-RLK transcript coexpressed with PXY, PXY/TDR-CORRELATED GENES1 (PXC1, At2g36570), is required for xylem fiber formation in stem secondary vasculature (Wang et al., 2013). Another RLK, XYLEM INTERMIXED WITH PHLOEM/C-TERMINALLY ENCODED PEPTIDE RECEPTOR 1 (XIP1/CEPR1; At5g49660) prevents the ectopic lignification of phloem in stems (Bryan et al., 2012). RLKs therefore seem to function in several aspects of secondary growth in a tissue-specific manner. Identifying their ligands and role in intercellular signaling will further increase our understanding of the coordination of secondary growth.
Figure 7.

Model of peptide signaling regulating secondary vascular tissues.
Peptide ligands produced in the endodermis (EPFL4, EPFL6) and phloem (CLE41/CLE44/TDIF and CLE42) are secreted and mediate intercellular signaling by binding to their receptors in phloem and cambium, respectively. Auxin has been shown to promote TDIF-TDR-WOX signaling, while the WOX genes are negative regulators of ethylene-mediated cambium proliferation. The TDIF-peptide simultaneously inhibits xylem differentiation, thereby affecting vascular organization. See text for details.
The Class III HD-ZIP transcription factors are involved in vascular development from the early globular embryo to secondary growth (Baima et al., 2001; Prigge et al., 2005). PHABULOSA (PHB; At2g34710), PHAVOLUTA (PHV; At1g30490), REVOLUTA/INTERFASCICULAR FIBERLESS (REV/IFL; At5g60690), ARABIDOPSIS THALIANA HOMEOBOX PROTEIN15/CORONA (ATHB15/CNA; At1g52150) and ATHB8 (At4g32880) may have synergistic, distinct or antagonistic roles in plant development, depending on the age and tissue (Prigge et al., 2005). Ilegems et al. (2010) concluded that the Class III HD-ZIPs promote axial elongation and tracheary element differentiation, therefore canalizing auxin flow. The KANADI (KAN) genes (KAN1; At5g16560, KAN2; At1g32240, KAN3; At4g17695, KAN4; At5g42630) are GARP transcription factors which seem to act antagonistically to the HD-ZIP III genes. KAN1 is expressed in the early globular embryo, later becoming restricted to the hypocotyl periphery and abaxial side of cotyledon primordia (Kerstetter et al., 2001). Over-expression of KAN1 hinders vascular development in the hypocotyl, likely by disturbing the central-periphery symmetry required for vascular development (Eshed et al., 2001; Kerstetter et al., 2001). KAN1 was shown to inhibit procambium and consequently vascular development under the procambial ATHB15 promoter in the Arabidopsis shoot, root and hypocotyl, whereas the kan1kan2kan3kan4 mutant exhibits premature secondary growth and an increase in the procambial cell number (Ilegems et al., 2010). Several lines of evidence suggest that KAN1 is a negative regulator of auxin transport, specifically PIN1 (At1g73590) expression (Izhaki and Bowman, 2007; Ilegems et al., 2010).
The low number of genes thus far identified playing a role in the regulation of cambial morphogenesis suggests that there may yet be abundant genetic regulators of secondary growth undiscovered. In the future, both forward and reverse genetic approaches may prove useful in the isolation of these factors.
REGULATION OF SHOOT SECONDARY GROWTH
Hormonal regulation of vascular cambium development in the Arabidopsis shoot
In the shoot, almost all known plant hormones have been shown to participate in the regulation of secondary development (reviewed by Brackmann and Greb, 2014). Among the various hormones, the contribution of auxin to cambial development is the most thoroughly studied. Through hormone and inhibitor treatments and mutant studies, basipetal (rootwards) auxin transport has been proven to be pivotal in the regulation of cambial activity in the Arabidopsis inflorescence stem. As the shoot tip represents a major auxin source, development of the vascular cambium can be inhibited through decapitation of the inflorescence stem (Little et al., 2002). The establishment and activity of both fascicular and interfascicular cambium can subsequently be reinstated by treating the decapitated stems with auxin (Little et al., 2002). Similarly, local treatment of the inflorescence stem with the auxin transport inhibitor NPA leads to the accumulation of auxin above the treatment position and stimulates cambial activity (Little et al., 2002; Suer et al., 2011). Genetic marker studies have shown that auxin accumulation in the interfascicular regions precedes the establishment of the interfascicular cambium. Expression of the DR5::GUS auxin response marker is induced early during the initiation of interfascular cambium by auxin treatment (Mazur et al., 2014). The marker first appears at the sides of the vascular bundle, next to the fascicular cambium, from where it gradually extends towards the middle of the interfascicular region (Mazur et al., 2014). Auxin accumulation is followed by expression of the PIN1 auxin transporter, after which the first periclinal cell divisions mark the onset of the interfascicular cambium (Mazur et al., 2014). In addition to PIN1, PIN3 has also been shown to have a high expression level in secondarily thickened stems (Gälweiler et al., 1998; Agusti et al., 2011a). Accordingly, in both the pin1 and pin3 loss-of-function mutants the initiation and activity of interfascicular cambium are delayed and reduced (Agusti et al., 2011a). Interestingly, even though the auxin transport capacity of pin1 mutants is strongly reduced (Okada et al., 1991; Gälweiler et al., 1998) and the inflorescence stem has transformed into a round pin-like structure, fascicular cambium development still appears to take place relatively normally inside the vascular bundle closest to the cauline leaf (Gälweiler et al., 1998). Presumably, PIN1 acts partly redundantly with some other auxin transporters, including PIN3, during the establishment and activity of vascular cambium. The proximity of a leaf, which acts as an auxin source, can therefore compensate for some defects in basipetal auxin transport. The initiation and activity of interfascicular cambium is also impaired in several auxin signaling and biosynthesis mutants, including the highly auxin insensitive auxin resistant 1 (axr1; At1g05180) (Agusti et al., 2011a), further supporting the role of auxin in the regulation of cambial activity. Together, these results demonstrate that basipetal auxin transport along the stem is positively correlated with both the initiation and activity of the vascular cambium.
In addition to auxin, cytokinin has also emerged as a pivotal regulator of both fascicular and interfascicular cambium development. As in the root, the strongest evidence for the significance of cytokinin signaling comes from the phenotype of the quadruple ipt mutant, in which cytokinin levels are severely decreased (Matsumoto-Kitano et al., 2008). The stem diameter is greatly reduced in this dwarf mutant, together with the number and size of the vascular bundles (Matsumoto-Kitano et al., 2008). Furthermore, the atipt3 single mutant, which has moderately decreased levels of cytokinins, has a narrower stem and fewer vascular cells inside the bundles, whereas the length of inflorescence stem remains similar to WT. This demonstrates that reduced cambial activity in this cytokinin biosynthesis mutant is not an indirect effect of impaired apical growth or the stunted size of the inflorescence stem. Further support for the role of cytokinin signaling in secondary development is provided by the single, double and triple loss-of-function mutants of the cytokinin receptors CRE1, AHK2 (At5g35750) and AHK3 (At1g27320), which also show reduced cambial activity (Mähönen et al., 2006b; Hejatko et al., 2009). However, as the mutants have a shorter stem than WT (Higuchi et al., 2004), the difference in vascular development may partly be an indirect effect of reduced growth.
In addition to the canonical cytokinin receptors, an atypical Arabidopsis two-component histidine kinase, CYTOKININ-INDEPENDENT 1 (CKI1; At2g47430) (Kakimoto, 1996), also participates in the regulation of cambial development (Hejatko et al., 2009). CKI1 can initiate the cytokinin signaling phosphorelay (Mähönen et al., 2006b) and induce cytokinin responses independently of cytokinin, and therefore does not represent a true cytokinin receptor (Yamada et al., 2001). CKI1 expression has been detected in the vascular tissues of the inflorescence stem, and its ectopic overexpression in Arabidopsis increases the number of cambial cells inside the vascular bundles (Hejatko et al., 2009). Consistent with this, the number of cambial cells was reduced in RNAi lines with reduced levels of CKI1 expression (Hejatko et al., 2009). It remains to be determined whether cytokinin regulates PIN expression and controls the auxin transport and distribution pattern in the shoot in a manner similar to during primary development in roots.
In addition to auxin and cytokinin, strigolactones have recently emerged as regulators of cambial activity. Highly branched Arabidopsis loss-of-function mutants for a strigolactone biosynthesis gene (MORE AXILLARY BRANCHES 1, MAX1; At2g26170) and for a positive regulator of strigolactone response (MAX2; At2g42620) display delayed and reduced interfascicular cambium activity (Agusti et al., 2011a). Furthermore, interfascicular cell divisions can be induced in both WT and max1 stems through treatment with a synthetic strigolactone, GR24 (Agusti et al., 2011a). The treatment does not repress the outgrowth of rosette branches in max1 mutant, demonstrating that strigolactone can stimulate secondary growth independently from its effect on shoot branching. There are indications that strigolactone signaling acts downstream of auxin in the regulation of secondary growth; the induction of interfascicular tissue production by NPA-treatment was strongly reduced in the max1 mutant, and the response was not further impaired in the highly auxin-insensitive axr1 max1 double mutant. It is currently not known what molecular pathways are the downstream targets of strigolactone signaling.
Based on mutant studies, the role of ethylene in the regulation of cambial activity appears to be more complex than the consistently stimulative role described for other hormones. Initiation of interfascicular cambium is reduced in the loss-of-function mutant of the ETHYLENE RESPONSE FACTOR 104 (ERF104; At5g61600), a positive downstream regulator of ethylene signaling (Sehr et al., 2010), whereas ethylene overproducer mutants (eto1; At3g15770 and eto2; At5g65800) display increased fascicular and interfascicular cambial activity (Etchells et al., 2012). Together, these results indicate that ethylene has a positive effect on cambial activity. By contrast, no vascular phenotype was observed in the inflorescence stems of either the ethylene insensitive2 (ein2; At5g03280) mutant, where ethylene signaling is assumed to be absent (Alonso et al., 1999), or the dominant negative ethylene insensitive receptor mutant ethylene response 1-3d (etr1-3d; At1g66340) (Bleecker et al., 1988; Etchells et al., 2012). These results together indicate that another molecular pathway acts in parallel with ethylene signaling during secondary development; see below for a discussion of the interaction between the PXY/WOX pathway and ethylene signaling.
Brassinosteroid signaling has a well-established role in regulating primary vascular patterning in the Arabidopsis shoot. Mutants deficient for brassinosteroid biosynthesis (constitutive photomorpogenic dwarf, cpd; At5g05690), perception (brassinosteroid insensitive 1, bri1; At4g39400) or signaling (bin2) display a reduced number of vascular bundles, whereas transgenic lines or mutations leading to elevated brassinosteroid hormone levels, such as the DWARF4 (DWF4; At3g50660) overexpressor, or signaling levels, such as the gain-of-function mutants bes1-d and brassinazole-resistant1-d (bzr1-d; At1g75080), increase the bundle number (Ibañes et al., 2009). Whether brassinosteroid signaling also has a role in the regulation of cambial activity is less well understood. DIMINUTO 1 (DIM1; At3g19820), a protein involved in brassinosteroid biosynthesis (Hossain et al., 2012), is expressed in both fascicular and interfascicular vasculature, most strongly in the developing xylem tissue (Hossain et al., 2012). DIM1 overexpressor lines appears to produce extra secondary xylem, whereas the lignification of xylem cells and interfascicular fibers, together with the number and size of xylem vessels, was reduced in DIM1 silencer lines (Hossain et al., 2012). These results indicate that brassinosteroid signaling may contribute to the regulation of secondary xylem production. However, the silencer plants were severely stunted, which may have indirectly affected the size of the vasculature.
In addition to these hormones, jasmonate (JA) signaling also contributes to the regulation of secondary growth in the Arabidopsis inflorescence stem (Sehr et al., 2010). Jasmonate treatment stimulates activity of the interfascicular cambium, and the jasmonate signaling genes CORONATINE INSENSITIVE 1 (COI1; At2g39940), MYC2 (At1g32640), JASMONATE-ZIM-DOMAIN PROTEIN 7 (JAZ7; At2g34600) and JAZ10 (At5g13220) are expressed in the secondary inflorescence stem. COI1 encodes a jasmonate receptor, whereas the JAZ genes act as repressors of JA signaling, and MYC2 is a transcription factor upregulating JA response genes (Sheard et al., 2010). In the loss-of-fuction jaz10 mutant, interfascicular cambium forms earlier and its activity is increased, whereas the initiation of interfascicular cambium was delayed in the coi1 and myc2 loss-of-function mutants (Sehr et al., 2010). Taken together, these results demonstrate a positive role for JA signaling in the regulation of interfascicular cambium initiation and activity.
Mechanosensory regulation of the shoot vascular cambium
Plants are known to able to adapt their developmental processes to a range of mechanical signals, including wind and touch. Plant growth response to repeated touching or bending, called thigmomorphogenesis (Jaffe, 1973), commonly results in shorter, thicker plants which are presumably more resistant to mechanical stress (Telewski and Jaffe, 1986; Coutand et al., 2008). However, an opposite response is seen in many plant species, including Arabidopsis, where mechanical stress instead inhibits radial growth. Repeated brushing and bending of the Arabidopsis inflorescence stem inhibits apical growth and does not stimulate cambial activity; in general, mechanically perturbed plants are thinner and more flexible than control plants, with thinner cell wall fibers (Paul-Victor and Rowe, 2011). The perturbed plants showed less evidence of fascicular cambial activity than the control plants, although this reduction may be a secondary effect of reduced apical growth (Paul-Victor and Rowe, 2011).
Even though an understanding of the molecular regulation underlying developmental responses to biophysical forces is still largely lacking, progress has been made through studies of the plant mechanostimulus sensory system. At least part of the response to mechanical stimuli in plants is mediated through Ca2+ signals, and MscS-like (MSL) and mid1-complementing activity (MCA) proteins have been identified as potential calcium permeable mechanosensitive channels (reviewed by Kurusu et al., 2013). These channels are thought to be responsible for sensing osmotic shock, touch and gravity. Furthermore, the touch-induced decrease in inflorescence stem elongation has been shown to be a jasmonate (JA) hormone-mediated response in Arabidopsis. Repetitive touch treatments have no effect on inflorescence elongation in the loss-of-function allene oxide synthase (aos; At5g42650) mutant, which is defective in jasmonate biosynthesis (Chehab et al., 2012). Similar results were obtained from the mutation of two other JA pathway genes, JASMONATE RESISTANT 1 (JAR1; At2g46370), which is required to generate the bioactive form of the hormone, and COI1, which encodes the JA-coreceptor (Chehab et al., 2012). Mechanostimulation by touching the inflorescence stem induces the expression of a JA signaling repressor, JAZ10, indicating that down-regulated jasmonate signaling may play a role in the inhibition of cambial activity in response to mechanical stress (Sehr et al., 2010). Ethylene signaling does not appear to be required for thigmomorphogenesis in Arabidopsis, as the ethylene-insensitive mutants, etr1 and ein2 maintain the reduced stem elongation response under wind treatment (Johnson et al., 1998).
In contrast to the negative effect of repeated bending on cambial activity, mechanical forces also play a positive role in the regulation of Arabidopsis secondary development. Development of secondary xylem at the base of the inflorescence stem is positively correlated with the height of the stem, indicating that the weight of the stem is able to stimulate cambial activity (Ko et al., 2004). Furthermore, formation of interfascicular cambium can be induced by weight treatment in decapitated stems (Fig. 8) (Ko et al., 2004). The weight-based increase in cambial activity is auxin dependent, as an auxin source, in the form of either applied auxin (Ko et al., 2004) or axillary branches (Mazur et al., 2014), is required for the activation of cambial development. In keeping with this, weight treatment of the stem stimulates basipetal auxin transport (Ko et al., 2004). However, initiation of the interfascicular cambium along the stem does not proceed at the same rate as the increase in weight of the growing shoot (Sehr et al., 2010), indicating that the dynamics between shoot weight and cambial activity are not completely linear. Interestingly, it has been reported by Mazur and Kurczynska (2012) that weight-treatment was able to induce the formation of ray-like structures in the inflorescence stem. Simple rays were detected in tangential cross-sections (Fig. 9), though it remains to be determined whether these ray-like structures are capable of transverse transport and whether Arabidopsis is also capable of developing rays during normal growth.
Figure 8.
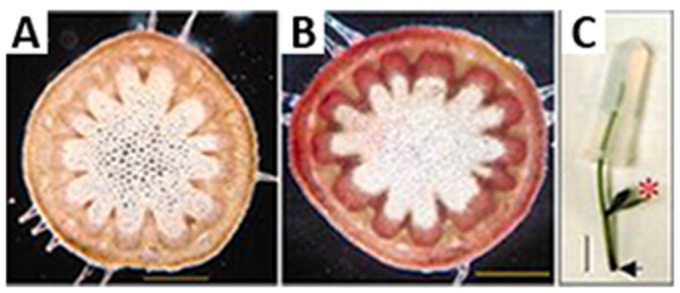
Weight treatment promotes secondary development in the Arabidopsis inflorescence stem.
A) Anatomy at the base of an intact 11 cm tall plant. B) Weight-treatment of a decapitated 5 cm tall plant has induced more secondary growth at the base of the stem than seen in (A). C) Weight (2.5 g) was applied to the top part of the decapitated stem. Phloroglucinol-HCl staining in A and C. Scale bars 0.5 mm (A and B); 1 cm (C). Reprinted with permission from Ko et al., 2004. Copyright © 2004 American Society of Plant Biologists.
Figure 9.
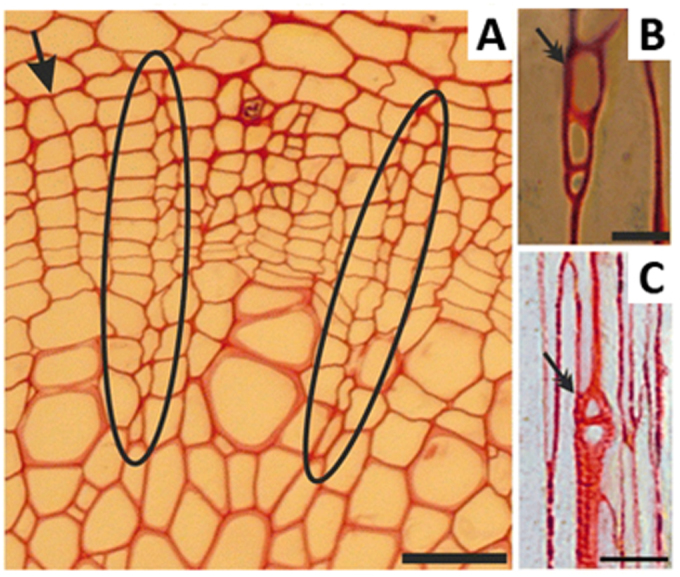
Weight-induced formation of vascular rays in the Arabidopsis inflorescence stem.
Two rays (circled) extend across the vascular cambium. The simple rays (marked by double arrows) are one cell wide (uniseriate). Reprinted with permission from Mazur and Kurczynska, 2012. Copyright © 2011 Mazur and Kurczynska.
Receptor kinase signaling pathways active during shoot secondary growth
As in the root and hypocotyl, the CLE-PXY-WOX pathway has emerged as an important regulator of cambial activity in the inflorescence stem. PXY is expressed in fascicular and interfascicular cambial cells (Etchells and Turner, 2010), and the spatial organization inside vascular bundles is disturbed in the pxy loss-of-function mutant, with some phloem cells adjacent to or interspersed within the xylem tissue (Fig. 10 A–B; F–G; K–L; P–Q) (Fisher and Turner, 2007). The shape of the vascular bundles is also affected; the bundles are more flat than in the WT, narrower radially and wider tangentially (Fisher and Turner, 2007). Instead of developing in the middle of the dividing cambial cell, the cell division planes form at aberrant and relatively random positions, leading to the development of irregularly shaped and curved xylem vessels (Etchells and Turner, 2010). Whereas single loss-of-function mutants of the PXY homologs pxl1 or pxl2 do not exhibit any obvious stem phenotype, the double-mutant and the combinations with pxy generate a more severe vascular phenotype than pxy alone (Fig. 10 A–E), indicating that these genes act redundantly in the regulation of vascular organization (Fisher and Turner, 2007). However, the pxl1 pxl2 pxy triple mutant does not exhibit an enhanced phenotype compared with the double mutants, indicating a somewhat complex functional redundancy between the three genes.
Figure 10.

Vascular anatomy of inflorescence stems in various secondary development mutant and transgenic lines.
(A–E) Xylem (xy) and phloem (ph) are intercalated in the pxy mutant, and the phenotype is enhanced by the pxl1 and pxl2 mutations. (A) WT Col; (B) pxy; (C) pxl1 pxl2; (D) pxy pxl1; (E) pxy pxl2. F–J) wox4 and wox4 wox14 double mutants have less secondary development but show no vascular organisation defects. (F) WT Col; (G) pxy; (H) wox4; (I) wox4 wox14; (J) pxy wox4. K–O) In comparison to wild type and pxy, erf109 erf018, pxy erf109 and pxy erf109 erf018 vascular tissue demonstrates a reduction in size. x is xylem, pc is procambium, ph is phloem. (K) WT Col; (L) pxy; (M) pxy erf109; (N) pxy erf018; (O) pxy erf109 erf018. P–T) Vascular organisation is perturbed in pxy and the phenotype is more severe in pxy er lines. (P) WT Col; (Q) pxy; (R) er; (S) pxy er; and (T) er erl1 erl2. U–W) Brackets enclose cambial cells; arrowheads mark phloem (Ph) cells directly touching xylem (Xy) cells. The dotted line in (W) indicates discontinuity of xylem regions. (U) WT Col; (V) epfl4 epfl6; (W) epfl4 epfl6 tdr. X–Z) Compared with WT, mol1 and rul1 show increased and decreased secondary growth, respectively. Brackets enclose cambial cells. (X) WT Col; (Y) mol; (Z) rul. AA–EE) Vascular bundles of rev-10d (BB) and rev-δmiRNA (CC) stems are often radialized and amphivasal, with xylem tissue (arrowheads) surrounding phloem tissue (ph). Vascular bundles in kan1 kan2 kan3 stems (DD) also exhibit an amphivasal pattern, with xylem (arrowheads) surrounding phloem (ph). rev phb phv displays a radialized amphicribral vascular bundle, where phloem (arrowheads) is surrounding xylem tissue (xy). (AA) WT Ler; (BB) the semidominant gain-of-function REV allele, rev-10d; (CC) miRNA resistant rev-δmiRNA; (DD) kan1 kan2 kan3; (EE) rev phb phv. FF–HH) Gaps in the bp vascular ring (arrows) are associated with stripes of ectopically lignified epidermal/cortical tissue. Arrowheads mark premature lignification of phloem fiber cells in primary vascular bundles. bop1 bop2 bp is similar to wild type. (FF) WT Col; (GG) bp; (HH) bop1 bop2 bp. Scale bars (A–E) (F–J) (K–O) (P–T) (X–Z) 50 µm; (U–W) 40 µm; (FF–HH) 100 µm. (A–E) reprinted with permission from Fisher and Turner, 2007; (F–J) from Etchells et al., 2013; (K–O) from Etchells et al., 2012; (P–T) from Etchells et al., 2013; (U–W) from Uchida and Tasaka, 2013; (X–Z) from Agusti et al., 2011b; (AA–EE) from Emery et al., 2003; (FF–HH) from Khan et al., 2012b. Copyright © 2007 Elsevier (A–E), © 2013 Development (F–J, P–T), © 2012 Etchells et al. (K–O), © 2013 Uchida and Tasaka (U–W), © 2011 Agusti et al. (X–Z), © 2003 Elsevier Science Ltd. (AA–EE) and © 2012 American Society of Plant Biologists (FF–HH).
Partially pxy-like phenotypes have also been detected in the missense and loss-of-function mutants of another LRR-RLK, XYLEM INTERMIXED WITH PHLOEM (XIP1/CEPR1) (Bryan et al., 2012). XIP1 is expressed in the vasculature, and the shoots of xip mutants show ectopic differentiation of cells with xylem characteristics inside the phloem tissues. Furthermore, differentiation of adjacent phloem and xylem cells is sometimes detected in the fascicular cambium (Bryan et al., 2012). Interestingly, XIP1 was recently identified to play a role in systemic nitrogen-demand signaling (Tabata et al., 2014). The potential partial redundancy between PXY and XIP1 functions, together with the possible link of vascular differentiation to nitrogen perception, remains to be studied.
As was described above, WOX4, a WUSCHEL-RELATED HOMEOBOX gene, acts downstream of PXY in the regulation of cambial cell proliferation. As in the root and hypocotyl, both fascicular and interfascicular cambial activity was reduced in the stem of the wox4 loss-of-function mutant (Fig. 10 F–J) (Suer et al., 2011). Consistent with this, WOX4 is expressed along the circumference of the vascular cambium, in both the fascicular and interfascicular regions (Suer et al., 2011). Auxin signaling appears to interact with the WOX4 pathway during cambial development. Based on in situ and reporter gene expression analyses, auxin response marker (DR5:GFP) expression precedes WOX4 expression during interfascicular cambium initiation (Suer et al., 2011). Furthermore, WOX4 expression is induced in response to NPA-mediated auxin accumulation (Suer et al., 2011). It has recently been shown that WOX4 acts redundantly with WOX14 in the regulation of cell division, and that even though the number of cells in vascular bundles is further reduced in the wox4 wox14 double mutant, the general organization of the bundles is not affected (Fig. 10 I) (Etchells et al., 2013). Interestingly, cambial activity is not completely abolished even in the double mutant, indicating that although the number of cambial cell divisions is reduced, WOX4 and WOX14 are not required for the establishment of the cambial meristem. Auxin may therefore stimulate cambial cell divisions partly through the WOX4 pathway, but it appears that some other auxin-mediated pathways activate the initiation of cambial meristem.
As in the root and hypocotyl, there is evidence that the PXY/ WOX pathway interacts not only with auxin but also with ethylene in the shoot vascular cambium (Etchells et al., 2012). Expression of several members of the AP2/ERF family of transcription factors is elevated in both pxy and wox4 mutant backgrounds, indicating that their expression is suppressed by the PXY/WOX4 pathway (Etchells et al., 2012). Among the upregulated genes are ERF109 (At4g34410) and ERF018 (At1g74930), which are expressed in the vascular bundles. Whereas single mutants of pxy or the erf genes did not show a significant decrease in the number of cells per vascular bundle, a reduction was seen in the pxy erf109 double mutant, and the number was further reduced in the pxy erf018 erf109 triple mutant (Fig. 10 K–O) (Etchells et al., 2012). Furthermore, even though the ein2 single mutant does not exhibit a vascular phenotype, the vascular cell number is severely reduced in the pxy ein2 double mutant. Together with the results of the ethylene biosynthesis mutant studies described above, these results confirm that ethylene signaling stimulates cambial activity, and that the PXY/WOX4-pathway normally represses this ethylene signaling -mediated pathway (Etchells et al., 2012). Since ethylene is well-known to regulate developmental responses to various environmental factors (Merchante et al., 2013), it is possible that ethylene controlled cambial growth is induced under certain physical conditions.
Besides its connection to the auxin and ethylene hormonal signaling, the PXY/WOX pathway also interacts with various other receptor kinase-mediated pathways, among them the LRR receptor-like kinase ERECTA (ER). ER has a general effect on inflorescence architecture; er loss-of-function mutants shoots have reduced size and compact stature (Torii et al., 1996; Bundy et al., 2012). Both ER and the closely related ERECTA-LIKE 1 (ERL1) are expressed in the phloem and xylem parenchyma tissue inside the vascular bundles of the inflorescence stem (Uchida and Tasaka, 2013). The single loss-of-function mutants does not have any evident vascular phenotype, but the double mutant er erl1 has abnormal vascular organization resembling the pxy mutant, where some phloem and xylem cells are directly in contact with each other, interrupting the continuity of the fascicular cambium (Fig. 10 P–T) (Uchida and Tasaka, 2013). This phenotype can be complemented through phloem-specific expression of ER. Both PXY and ER contribute to the regulation of vascular bundle organization; in the pxy er double mutant, the shape of the vascular bundles is flattened and the xylem and phloem tissues are more severely intercalated than in the pxy single mutant (Etchells et al., 2013; Uchida and Tasaka, 2013). In fact, the pxy mutant was originally identified in the Landsberg erecta background with a mutated ER gene, and the intercalated tissue phenotype is milder in Columbia background.
The EPIDERMAL PATTERNING FACTOR LIKE 4 (EPFL4; At4g14723) and EPFL6 (At2g30370) peptides have been demonstrated to act as ligands of the ER proteins in the regulation of shoot elongation (Abrash et al., 2011; Uchida et al., 2012). However, despite their high expression level in the endodermal layer of the inflorescence stem (Uchida et al., 2012), no vascular phenotype was observed in the epfl4 epfl6 double mutant (Uchida and Tasaka, 2013). By contrast, the epfl4 epfl6 pxy triple mutant displays a phenotype resembling er pxy, with flattened vascular bundles and intercalated vascular tissues (Fig. 10 U–W). Expression of either EPFL4 or EPFL6 in the endodermal tissue rescues the phenotype to the level of the pxy single mutant. These results indicate that endodermis-derived EPFL4 and EPFL6 peptides are perceived in phloem by the receptors ER/ERL1. Based on the lack of a vascular phenotype in epfl4 epfl6, it is possible that EPFL4 and EPFL6 act redundantly with some other ligands in the regulation of ER proteins (Uchida and Tasaka, 2013).
In summary, in the WOX/PXY pathway, WOX4 regulates the cell division rate together with WOX14, whereas PXY functions with ER mainly in the regulation of vascular tissue patterning (Etchells et al., 2013). Interestingly, ER, ERL1 and ERL2 (At5g07180) were recently shown to regulate auxin transport in the SAM during leaf initiation; they enhance the expression of PIN1 in the vasculature of forming leaf primordia (Chen et al., 2013). It remains to be studied whether the ER family proteins have any analogous function during cambial development.
In addition to the PXY and ER family genes, two other receptor-like kinases, MORE LATERAL GROWTH1 (MOL1; At5g51350) and REDUCED IN LATERAL GROWTH1 (RUL1; At5g05160) have emerged as regulators of cambial activity (Agusti et al., 2011b). The mol1 loss-of-function mutant displays substantially enhanced formation of secondary vascular tissue in both fascicular and interfascicular regions compared with wildtype. By contrast, tissue formation in the interfascicular regions is decreased in the rul1 loss-of-function mutant, reflecting a reduction in cambium activity (Fig. 10 X–Z). MOL1 therefore appears to act as a repressor and RUL1 as an activator of secondary growth in the inflorescence stem (Agusti et al., 2011b). Analysis of the mol1 rul1 double mutant revealed a wild-type-like rate of vascular tissue production in the interfascicular regions, indicating that the two mutations can compensate for each other. RUL1, PXY and WOX4 expression were upregulated in the mol1 background, whereas the level of WOX4 was almost unchanged in rul1. These results indicate that MOL1 may function upstream of RUL1 and the PXY-WOX4 pathways, even though their function in the either same or parallel pathways remains to be demonstrated.
Arabidopsis mutants with enhanced shoot secondary development
Direct screens for mutants with altered secondary vascular development have identified a limited number of mutants with enhanced formation of vascular tissue in the interfascicular regions. These phenotypes culminate in the early establishment of a continuous ring of vascular cambium. high cambial activity2 (hca2; At5g62940), a gain-of-function mutant of the Dof transcription factor DOF5.6/HCA2, shows lateral expansion of vascular bundles and a severe reduction of the the interfascicular area between them (Guo et al., 2009). HCA2 is highly expressed in the cambium, phloem and interfascicular parenchyma, supporting its role in the regulation of cambial development. The dof5.6 loss-of-function mutant does not display any detectable vascular phenotype, indicating that the gene may be redundant with some other Dof transcription factors. Another mutant with a similar phenotype is continuous vascular ring (cov1; At2g20120) (Parker et al., 2003). Recently, COV1 was identified to be a trans-Golgi network localized membrane protein, and the cov1 mutant was shown to exhibit abnormal Golgi morphology and defects in vacuolar protein sorting (Shirakawa et al., 2014). The connection between membrane trafficking and vascular cambium development remains to be clarified. Yet another similar phenotype has been seen in the high cambial activity (hca) mutant (Pineau et al., 2005), although the causative mutation, which is not allelic to either HCA2 or COV1, has not yet been identified. Interestingly, all three mutants with an early continuous vascular ring phenotype display impaired inflorescence growth and a reduced stem diameter, indicating that the early induction of secondary growth is potentially related to their stunted stature.
Transcription factor mediated regulation of cambial development in the Arabidopsis shoot
The Class III HOMEODOMAIN LEUCINE-ZIPPER (HD-ZIP III) transcription factors have emerged as important regulators of vasculature organization and polarity in the shoot, as in the root and hypocotyl (reviewed by Sanchez et al., 2012). As mentioned above, Arabidopsis has five HD-ZIP III genes: REVOLUTA (REV/ IFL), PHABULOSA (PHB/ATHB14), PHAVOLUTA (PHV/ATHB9), CORONA (CNA/ATHB15) and ATHB8. Their expression is controlled through microRNA165/166-mediated mRNA degradation; loss of a microRNA binding site leads to gain-of-function mutations with ectopic and enhanced expression patterns. The functions of the five genes are highly interconnected; in different combinations they display various redundant and antagonistic interactions. Adding to the complexity, members of the KANADI GARP transcription factor family (KAN1-3) antagonistically regulate the activity of the HD-ZIP III genes. The KANADI genes are expressed in the developing phloem, whereas the HD-ZIP III genes are present in the developing xylem (Emery et al., 2003).
In the stem, the function of REV has received the most study. The loss of microRNA regulation in rev gain-of-function mutants leads to a dramatic change in vascular organization; the bundles develop in a radialized amphivasal pattern, where xylem surrounds the phloem tissue (Fig. 10 AA–CC) (Zhong et al., 1999; Emery et al., 2003; Zhong and Ye, 2004; Zhou et al., 2007). In addition to the compromised anatomy, the position of bundles is also affected; instead of forming a ring at the periphery of the stem, they are located more internally, being present even in the pith of the stem (Emery et al., 2003; Zhong and Ye, 2004). Additionally, the development of secondary cell wall in the interfascicular fibers is severely reduced (Zhong and Ye, 2004; Zhou et al., 2007). The kan1 kan2 kan3 triple loss-of-function mutant stem phenocopies the amphivasal vascular bundle phenotype of the rev gain-of-function mutant (Fig. 10 DD), demonstrating the antagonistic relationship between REV and the KANADI genes in the regulation of vascular bundle organization (Emery et al., 2003).
In the rev loss-of-function mutant, the organization of the vascular bundles is normal, but the differentiation of interfascicular fibers is impaired (Zhong and Ye, 2001). Single phb, phv or athb8 loss-of-function mutants do not have any distinct vascular phenotypes in the stem (Baima et al., 2001; Emery et al., 2003), indicating the redundancy between the HD-ZIP III genes. However, the combination of either phb or phv with rev makes the size and position of vascular bundles more erratic (Prigge et al., 2005). The triple loss-of-function mutant rev phb phv has a dramatic phenotype; the mutant lacks an apical meristem and produces only a single radialized cotyledon with a radialized amphicribral vascular bundle, where the phloem is surrounding xylem tissue (Fig. 10 EE) (Emery et al., 2003). Reversed vascular organization is observed in the radialiazed leaves of the phb gain-of-function mutant; in extreme cases the leaves lack a vascular strand, whereas less affected leaves have radialized amphivasal bundles (McConnell and Barton, 1998).
The distribution of vascular bundles is also disturbed in the cna loss-of-function mutant, reflecting the importance of HD-ZIP III genes for vascular organization. The phenotype is more severe in the cna phb phv triple mutant, where the stem diameter is increased and internal vascular bundles develop in the pith (Prigge et al., 2005). The defects in both the rev and cna single mutants are partially suppressed in the rev cna athb8 triple mutant, which has normally positioned bundles and differentiatied interfascicular fibers (Prigge et al., 2005). Interestingly, ectopic overexpression of ATHB8 under the 35S promoter enhances fascicular and interfascicular cambium activity (Baima et al., 2001). Additionally, the shape of the vascular bundles is flattened and many pith cells are ectopically lignified (Baima et al., 2001). This phenotype indicates that ATHB8 may act as a positive regulator of cambial activity and xylem development.
The HD-ZIP III pathway has been connected to auxin signaling. In the loss-of-function rev/ifl mutant, the PIN1 and PIN2 (At5g57090) auxin transporters are expressed at lower levels, and the mutant displays reduced auxin transport along the inflorescence stem (Zhong and Ye, 2001). In addition, chromatin immunoprecipitation studies have shown that REV upregulates several auxin biosynthesis, transport and response genes, including PIN1, while KAN1 downregulates many of them (Merelo et al., 2013; Huang et al., 2014). However, activity of both the fascicular and interfascicular cambium appears to proceed relatively normally in the rev stem (Talbert et al., 1995), indicating that REV itself is not necessary for secondary growth and that there is still sufficient auxin transport taking place for cambial development.
Altogether, these results demonstrate that the HD-ZIP III and KANADI genes are central for the regulation of the positioning and organization of the vasculature. Their role in the regulation of cambial activity is more difficult to decipher, since the pleiotropic nature of the loss-of-function and overexpression phenotypes makes it challenging to dissect any direct effect on secondary development. Furthermore, the connections between the five HD-ZIP III genes are highly complex. The results indicate that REV and CNA both act partly redundantly with PHB and PHV during vascular development, but are also partly antagonistically towards each other. Further complicating interpretation is the fact that the REV gain-of-function and loss-of-function mutants each produce radialized vascular bundles and impaired interfascicular fiber differentiation in certain mutant combinations; it seems that a fine-tuned balance of REV function is required for normal vascular organization and development.
In addition to the WOX, DOF, ERF, HD-ZIP III and KAN gene families, several other transcription factors contribute to the regulation of shoot vascular development. Three interacting homeodomain transcription factors, the KNOX protein BREVIPEDICELLUS/KNOTTED-LIKE FROM ARABIDOPSIS THALIANA 1 (BP/KNAT1; At4g08150), SHOOT MERISTEMLESS (STM; At1g62360) and the BELL family protein PENNYWISE/ BELLRINGER/VAANAMA (PNY/BLR/VAN; At5g02030) function in the regulation of Arabidopsis shoot architecture and vascular development (Bhatt et al., 2004; Etchells et al., 2012; Khan et al., 2012a; Khan et al., 2012b; Liebsch et al., 2014). It is well-established that various KNOX and BELL family proteins interact during meristem development; these interactions determine their subcellular localization and DNA binding affinity (reviewed by Hay and Tsiantis, 2010). Different heterodimer combinations of KNOX/BELL transcription factors may regulate specific downstream genes.
The shoot of the bp loss-of-function mutant has sectors which are defective in the differentiation of vascular tissues. Within these sectors, the cells inside the vascular bundles are small and often lack secondary cell walls; interfascicular fibers are also absent (Fig. 10 FF–GG). These sectors are associated with stripes of ectopically lignified cortical tissue and appear as gaps in the vascular ring (Khan et al., 2012a; Khan et al., 2012b). Vascular bundles with deficient xylem fiber differentiation also frequently appear in the shoot of the weak stm loss-of-function mutant (Liebsch et al., 2014). By contrast, single loss-of-function mutants of pny have an increased number of vascular bundles which appear relatively normal (Smith and Hake, 2003; Etchells et al., 2012). The bp phenotype is enhanced in the bp pny double mutant, where very few cells inside the defective sectors have secondary walls. Interestingly, secondary growth is stimulated in both the normal and defective areas in the mutant, indicating that BP and PNY function as redundant repressors of this excessive cell proliferation. Since vascular growth is reduced in bp pny rev triple mutant, REV may be connected to BP-PNY-mediated regulation (Etchells et al., 2012). However, it is also possible that reduced auxin transport in the rev background may have an indirect inhibitory effect on growth in bp pny.
It has been shown that the defects in vascular tissue differentiation in bp and pny are linked to the misexpression of two lateral organ boundary genes, BLADE-ON-PETIOLE1 (BOP1; At3g57130) and BOP2 (At2g41370), which appear to function downstream of BP-STM-PNY in an antagonistic fashion. BOP1 and BOP2 are normally expressed only in the pedicel axils, but in the bp pny mutant their expression pattern expands from the pedicel to the main stem. Similarly, BOP1 and BOP2 expression is also upregulated in the weak stm mutant (Liebsch et al., 2014). The defective sectors in bp are rescued in the bop1 bop2 bp triple mutant (Fig. 10 FF–HH), and the vascular bundle number is normalized in the bop1 bop2 pny mutant (Khan et al., 2012b). It would be interesting to study whether the loss of BOP1 and BOP2 function can also rescue the excessive cell proliferation in the bp pny double mutant or if this phenotype is independent of the two genes.
XYLEM CELL DIFFERENTIATION DURING CAMBIAL DEVELOPMENT
Xylem cell differentiation consists of four partially distinct processes: secondary cell wall deposition, programmed cell death (PCD), autolysis, and lignification (reviewed in Escamez and Tuominen, 2014). A few NAC-domain transcription factors (VASCULAR RELATED NAC DOMAIN; VND) have been identified as master regulators of xylem differentiation capable of switching on the developmental program. VND7 (At1g71930) induces protoxylem differentiation and VND6 (At5g62380) metaxylem differentiation (Kubo et al., 2005), while SECONDARY WALL-ASSOCIATED NAC DOMAIN PROTEIN 1/NAC SECONDARY WALL THICKENING PROMOTING FACTOR 3 (SND1/NST3; At1g32770), together with NST1 (At2g46770), promotes fiber differentiation (Mitsuda et al., 2007) (reviewed in Zhang et al., 2011).
The NAC master regulators turn on transcription factors that regulate downstream developmental processes, among them the MYB transcription factors MYB46 (At5g12870) and MYB83 (At3g08500), which regulate secondary cell wall synthesis and deposition (Ohashi-Ito et al., 2010; Zhong et al., 2010; Zhong and Ye, 2012) in both tracheary elements and fibers (Zhong et al., 2007; McCarthy et al., 2009). The induction of PCD, characterized by the burst of vacuolar membrane and arrested cytoplasmic streaming, occurs after the deposition of secondary cell wall is completed. Since xylary fibers do not die in Arabidopsis (Bollhöner et al., 2012), PCD takes place only in the tracheary elements. The timing of PCD is regulated by the polyamine thermospermine, which is synthesized by ACAULIS5 (ACL5; At5g19530) (Muñiz et al., 2008). PCD is followed by autolysis of the protoplast and weakening of the primary cell wall (reviewed in Escamez and Tuominen, 2014). VND6 is known to induce expression of the cysteine proteases XYLEM CYSTEINE PEPTIDASE 1 (XCP1; At4g35350) and XCP2 (At1g20850) (Ohashi-Ito et al., 2010), which participate in autolysis during tracheary element differentiation (Avci et al., 2008). Lignification is the last step of xylem cell differentiation. Monolignols are stored within the vacuole and released during PCD to polymerize into the cell wall; the monolignol polymerization process is partly non-cell autonomous and occurs mainly after PCD (Pesquet et al., 2013). Very recently, systematic protein-DNA network analysis revealed the signaling network regulating secondary cell wall synthesis in Arabidopsis (Taylor-Teeples et al., 2015).
NATURAL VARIATION IN BRASSICACEAE SECONDARY DEVELOPMENT
Arabidopsis thaliana belongs to the Brassicaceae family, which has a large number of species (over 3000) and a worldwide distribution. The family includes species with different lifestyles, including annual and perennial herbaceous plants, dwarf shrubs and shrubs. Brassicaceae species with economic importance include numerous vegetables and flowering plants, among them horseradish (Armoracia rusticana), broccoli, brussel sprouts, cauliflower, cabbage, collards, cale (all cultivare of Brassica oleracea), rutabaga and canola oil (B. napus), mustard (B. nigra), turnip (B. rapa), wasabi (Eutrema japonicum), and many more.
The diversity of Brassicaceae provides ample opportunity to compare different traits between related species and to learn about the evolution and adaptiveness of the studied traits. An example of successful interspecies research is the study of composite leaf development in Cardamine hirsuta, a close relative of Arabidopsis (Hay and Tsiantis, 2006; Barkoulas et al., 2008; Blein et al., 2008; Vlad et al., 2014). Arabidopsis has lost this leaf form during its evolution and has simple leaves; by comparing the two species, it was possible to study the evolutionary history and potential adaptiveness of these two forms. A similar approach could be applied to root development research, since a diverse array of wood anatomies is present among the Brassicaceae (Fig. 11) (Schweingruber et al., 2011). Many, like Arabidopsis, are rayless in normal conditions, but several form either thin or wide (multiseriate) vascular rays. Annual growth rings are often present in perennial species, and vessels can be either mostly solitary or arranged in clusters (Lens et al., 2012).
Interestingly, the developmental response to wind exposure appears to vary between species of Brassicaceae. The stem of some species elongates in response to wind, whereas others, including Arabidopsis, decrease in height. A similar difference is seen in cambial activity; some species respond by getting thicker, whereas others grow less (Murren and Pigliucci, 2005). Comparative studies between different Brassicaceae species could provide information about the adaptive value of these different responses to mechanical stress, as well as the molecular mechanisms behind them.
Arabidopsis has the capacity to be genetically engineered to have a woody and perennial lifestyle, reflecting the evolutionary history of different lifestyles in Brassicaceae. A double loss-of-function mutant for two Arabidopsis MADS box genes, SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1; At2g45660) and FRUITFULL (FUL; At5g60910) shows massive secondary xylem accumulation in the inflorescence stem (Fig. 12) (Melzer et al., 2008). Both of these genes play a role in controlling flowering time; SOC1 integrates signals for photoperiod, temperature, gibberellins and age, while FUL regulates floral meristem identity, together with carpel and leaf development. Together, they affect both flowering time and meristem determinacy. The double soc1 ful mutant displays a shrub-like phenotype due to continuous growth of vegetative aerial rosettes. It also exhibits other features of perennial plants, including a prolonged lifetime, the co-occurrence of vegetative and reproductive meristems and recurrent flowering cycles (Melzer et al., 2008). Interestingly, despite the extensive secondary growth, no vascular rays were detected in the soc1 ful stem (Melzer et al., 2008). This contrasts with the weight-induced ray formation discussed above. Despite the lack of rays, the wood of soc1 ful Arabidopsis looks similar to several naturally shrubby Brassicaceae species (Fig. 11; Fig. 12) (Melzer et al., 2008; Lens et al., 2012), providing an opportunity to study the features leading to woodiness in herbaceous species.
Figure 12.

Perennial features in the soc1 ful Arabidopsis mutant.
A) Comparison of a 2-month-old Col wild-type plant (left) and an 8-month-old soc1 ful mutant (right). The mutant has many aerial rosettes at the base and shows floral reversion at inflorescence meristems. B) Anatomy of the base of a mature Col inflorescence stem. C) Anatomy of a mature soc1 ful stem illustrating extensive secondary growth. D) Close-up of the soc1 ful stem. The stem has one annual ring; no vascular rays were detected. Scale bars 500 µm. (A) reprinted with permission from Melzer et al., 2008; (B–D) from Lens et al., 2012. Copyright © 2008 Nature Publishing Group and © 2011 Lens et al., respectively.
Interestingly, a continuous ring of vascular cambium is established very early in the inflorescence stem of the double mutant. This indicates that the enhanced secondary development results not only from an increase in life span, but that SOC1 and FUL may also act as more direct repressors of secondary growth. The possibility of engineering a herbaceous plant into to a perennial woody growth habit by modifying just a few of its genes reflects the evolutionary flexibility between the woody and herbaceous plant lifestyles.
STEM CELLS AND CAMBIUM
Stem cells are located in the meristems, where they maintain the undifferentiated state of the other meristamatic cells. A classical stem cell niche consists of a group of cells called an organizing center which keeps the adjacent stem cells from differentiating. A good example of this is the stem cell system in the root meristem, where the quiescent center (QC) functions as an organizing center and maintains the undifferentiated state of the surrounding stem cells (van den Berg et al., 1997). Organizing center identity is promoted by QC-specific expression of the WOX5 (WUSCHEL-RELATED HOMEOBOX 5; At3g11260) homeodomain gene (Sarkar et al., 2007); another member of the same gene family, WUS, promotes organizing center identity in shoot apical meristem (Schoof et al., 2000; Brand et al., 2000). As described above, it was recently discovered that two other members of the same gene family, WOX4 and WOX14, are expressed in the cambium, and their mutants show reduced cambial growth (Hirakawa et al., 2010; Ji et al., 2010; Etchells et al., 2013). These results raise the possibility that WOX4 and WOX14 are stem cell factors in cambium. However, WOX4 and WOX14 seem to have a broad expression domain within the cambium, rather than being restricted to a narrow area as would be typical for factors in a stem cell niche. Additionally, the ontogeny of cambial cells and the xylem and phloem cells produced by the cambium is not fully understood, making it difficult to build a framework for the stem cell system and its regulators (such as WOX4 and WOX14) in cambium. Nonetheless, classical microscopic studies have shed some light on cambial organization. Early on, researchers noted that the cambium consists of a few layers of ultrastructurally similar, narrow, thin-walled, elongated cells, making it difficult to assign specific functions to the cells in each layer. Despite these technical hurdles, researches have generally agreed that there is a single layer of cambial initials producing xylem mother cells inwards and phloem mother cells outwards from the cambium (reviewed in Larson, 1994; Lachaud et al., 1999). The prospect for the future is to assess these classical observations with modern methods and combine them with knowledge of gene functions in the cambium.
FUTURE OUTLOOK
In recent years, our understanding of the development and function of vascular cambium in Arabidopsis has advanced greatly. Even though important progress has been made, some pivotal aspects of cambium function remain unknown, among them the position and nature of the cambium stem cells and the difference between fascicular and interfascicular cambium development. One of the major challenges for secondary development research is that several genes required for the function of vascular cambium are already active during embryo development and also function in the apical meristems during primary development. It is therefore challenging to dissect their function specifically during the initiation and activity of cambial meristem separate from potentially indirect effects of early developmental defects. The study of vascular phenotypes derived from inducible overexpressor and silencing constructs driven under strictly cambium specific promoters will be highly informative, as it will enable at least partial separation of the effects during primary and secondary development. These approaches will allow us to identify the specific differences between the development and function of primary and secondary meristems, even though, as has been discussed above, they share several analogous signaling pathways.
Another emerging aspect of secondary development research, complementing the inducible mutants, is computer-assisted phenotyping (Sankar et al., 2014), which will enable us to detect and quantify subtle differences in the pattern and number of secondary tissues from a large quantities of cross-sections. This approach will facilitate the characterisation of mutant phenotypes and natural ecotypes. In addition to automated phenotyping, computer-based modelling approaches can be developed in the future to help us understand and interpret the molecular regulation behind the formation of diverse cambial patterns.
However, even though Arabidopsis is a versatile and accessible model organism for studies of cambial development and wood formation, as discussed above, it still lacks several features present in perennial and woody plants. Due to its small size and short life-span, it has no need for the formation of annual growth rings (often with distinctive earlywood-latewood characteristics). Even in biannual Arabidopsis, the inflorescence stem only develops during the second year and lives for one growing season, and therefore has no need to undergo an annual cambial activity—dormancy cycle. Arabidopsis secondary growth also does not require the development of vascular rays, which facilitate radial transport across woody tissue. Interestingly, structures resembling simple vascular rays can be induced to form in the Arabidopsis inflorescence stem under weight-treatment, even though they do not appear to be required for sustaining prolonged secondary growth in Arabidopsis mutants with perennial features. It appears that Arabidopsis has the molecular machinery for ray production available for activation under certain circumstances. This, together with the ease of transforming Arabidopsis into a perennial through the mutation of just two genes, underscores the evolutionary flexibility of woody versus herbaceous plant development.
Some of these features can be studied in different Brassicaceae species, which display various wood formation traits. However, none of them represents a de facto tree; they are all of relatively modest size and stature, reaching a shrub-like phenotype at best. Arabidopsis studies can therefore also be complemented through research into multiple wood formation related traits in full-sized tree species. These features include the annual activity-dormancy cycle of cambium, the production of early vs latewood during the growth period, the development and function of complex vascular rays consisting of several specialized cell types, and the formation of tension wood and heartwood. The comparison of the molecular mechanisms behind traits present in either herbaceous or tree species will help us understand the defining differences between the perennial woody and annual herbaceous plant lifestyle.
ACKNOWLEDGEMENTS
We are grateful to Riccardo Siligato and Marcelo Pace for providing pictures and to Sedeer El-Showk for proofreading the text. T.B., Y.H. and A.P.M. were funded by the Academy of Finland Centre of Excellence programme. Additionally, financial support for K.N. was provided by the Academy Postdoctoral Researcher grant and Natural Resources Institute Finland (Luke); for T.B and A.P.M by the Academy Research Fellowship grant and University of Helsinki; and for Y.H. by the Gatsby Foundation, University of Helsinki, the European Research Council Advanced Investigator Grant Symdev and Tekes (the Finnish Funding Agency for Technology and Innovation).
Footnotes
Citation: Kaisa Nieminen, Tiina Blomster, Ykä Helariutta and Ari Pekka Mähönen. (2015) Vascular Cambium Development. The Arabidopsis Book 11:e0177. doi:10.1199/tab.0177
elocation-id: e0177
*These authors contributed equally.
First published on May 21, 2015: e0177. doi: 10.1199/tab.0177
REFERENCES
- Abrash E.B., Davies K.A., Bergmann D.C. Generation of signaling specificity in Arabidopsis by spatially restricted buffering of ligand-receptor interactions. Plant Cell. 2011;23:2864–2879. doi: 10.1105/tpc.111.086637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agusti J., Herold S., Schwarz M., Sanchez P., Ljung K., Dun E.A., Brewer P.B., Beveridge C.A., Sieberer T., Sehr E.M., Greb T. Strigolactone signaling is required for auxin-dependent stimulation of secondary growth in plants. Proc. Natl. Acad. Sci. USA. 2011a;108:20242–20247. doi: 10.1073/pnas.1111902108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agusti J., Lichtenberger R., Schwarz M., Nehlin L., Greb T. Characterization of transcriptome remodeling during cambium formation identifies MOL1 and RUL1 as opposing regulators of secondary growth. PLoS Genet. 2011b;7:e1001312. doi: 10.1371/journal.pgen.1001312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alonso J.M., Hirayama T., Roman G., Nourizadeh S., Ecker J.R. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science. 1999;284:2148–2152. doi: 10.1126/science.284.5423.2148. [DOI] [PubMed] [Google Scholar]
- Altamura M.M., Possenti M., Matteucci A., Baima S., Ruberti I., Morelli G. Development of the vascular system in the inflorescence stem of Arabidopsis. New Phytol. 2001;151:381–389. [Google Scholar]
- Argyros R.D., Mathews D.E., Chiang Y.H., Palmer C.M., Thibault D.M., Etheridge N., Argyros D.A., Mason M.G., Kieber J.J., Schaller G.E. Type B response regulators of Arabidopsis play key roles in cytokinin signaling and plant development. Plant Cell. 2008;20:2102–2116. doi: 10.1105/tpc.108.059584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avci U., Petzold H.E., Ismail I.O., Beers E.P., Haigler C.H. Cysteine proteases XCP1 and XCP2 aid micro-autolysis within the intact central vacuole during xylogenesis in Arabidopsis roots. Plant J. 2008;56:303–315. doi: 10.1111/j.1365-313X.2008.03592.x. [DOI] [PubMed] [Google Scholar]
- Baima S., Possenti M., Matteucci A., Wisman E., Altamura M.M., Ruberti I., Morelli G. The Arabidopsis ATHB-8 HD-Zip protein acts as a differentiation-promoting transcription factor of the vascular meristems. Plant Physiol. 2001;126:643–655. doi: 10.1104/pp.126.2.643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkoulas M., Hay A., Kougioumoutzi E., Tsiantis M. A developmental framework for dissected leaf formation in the Arabidopsis relative Cardamine hirsuta. Nat. Genet. 2008;40:1136–1141. doi: 10.1038/ng.189. [DOI] [PubMed] [Google Scholar]
- Baum S.F., Dubrovsky J.G., Rost T.L. Apical organization and maturation of the cortex and vascular cylinder in Arabidopsis thaliana (Brassicaceae) roots. Am. J. Bot. 2002;89:908–920. doi: 10.3732/ajb.89.6.908. [DOI] [PubMed] [Google Scholar]
- Bhatt A.M., Etchells J.P., Canales C., Lagodienko A., Dickinson H. VAAMANA—a BEL1-like homeodomain protein, interacts with KNOX proteins BP and STM and regulates inflorescence stem growth in Arabidopsis. Gene. 2004;328:103–111. doi: 10.1016/j.gene.2003.12.033. [DOI] [PubMed] [Google Scholar]
- Bishopp A., Help H., El-Showk S., Weijers D., Scheres B., Friml J., Benková E., Mähönen A.P., Helariutta Y. A mutually inhibitory interaction between auxin and cytokinin specifies vascular pattern in roots. Curr. Biol. 2011a;21:917–926. doi: 10.1016/j.cub.2011.04.017. [DOI] [PubMed] [Google Scholar]
- Bishopp A., Lehesranta S., Vatén A., Help H., El-Showk S., Scheres B., Helariutta K., Mähönen A.P., Sakakibara H., Helariutta Y. Phloem-transported cytokinin regulates polar auxin transport and maintains vascular pattern in the root meristem. Curr. Biol. 2011b;21:927–932. doi: 10.1016/j.cub.2011.04.049. [DOI] [PubMed] [Google Scholar]
- Bleecker A.B., Estelle M.A., Somerville C., Kende H. Insensitivity to ethylene conferred by a dominant mutation in Arabidopsis thaliana. Science. 1988;241:1086–1089. doi: 10.1126/science.241.4869.1086. [DOI] [PubMed] [Google Scholar]
- Blein T., Pulido A., Vialette-Guiraud A., Nikovics K., Morin H., Hay A., Johansen I.E., Tsiantis M., Laufs P. A conserved molecular framework for compound leaf development. Science. 2008;322:1835–1839. doi: 10.1126/science.1166168. [DOI] [PubMed] [Google Scholar]
- Bollhöner B., Prestele J., Tuominen H. Xylem cell death: emerging understanding of regulation and function. J. Exp. Bot. 2012;63:1081–1094. doi: 10.1093/jxb/err438. [DOI] [PubMed] [Google Scholar]
- Brackmann K., Greb T. Long- and short-distance signaling in the regulation of lateral plant growth. Physiol. Plant. 2014;151:134–141. doi: 10.1111/ppl.12103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brand U., Fletcher J.C., Hobe M., Meyerowitz E.M., Simon R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science. 2000;289:617–619. doi: 10.1126/science.289.5479.617. [DOI] [PubMed] [Google Scholar]
- Bryan A.C., Obaidi A., Wierzba M., Tax F.E. XYLEM INTERMIXED WITH PHLOEM1, a leucine-rich repeat receptor-like kinase required for stem growth and vascular development in Arabidopsis thaliana. Planta. 2012;235:111–122. doi: 10.1007/s00425-011-1489-6. [DOI] [PubMed] [Google Scholar]
- Bundy M.G., Thompson O.A., Sieger M.T., Shpak E.D. Patterns of cell division, cell differentiation and cell elongation in epidermis and cortex of Arabidopsis pedicels in the wild type and in erecta. PLoS One. 2012;7:e46262. doi: 10.1371/journal.pone.0046262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busse J.S., Evert R.F. Vascular Differentiation and Transition in the Seedling of Arabidopsis thaliana (Brassicaceae). Int. J. Plant Sci. 1999a;160:241–251. [Google Scholar]
- Busse J.S., Evert R.F. Pattern of differentiation of the first vascular elements in the embryo and seedling of Arabidopsis thaliana. Int. J. Plant Sci. 1999b;160:1–13. [Google Scholar]
- Caño-Delgado A., Yin Y., Yu C., Vafeados D., Mora-Garcia S., Cheng J.C., Nam K.H., Li J., Chory J. BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development. 2004;131:5341–5351. doi: 10.1242/dev.01403. [DOI] [PubMed] [Google Scholar]
- Carlsbecker A., Lee J.Y., Roberts C.J., Dettmer J., Lehesranta S., Zhou J., Lindgren O., Moreno-Risueno M.A., Vatén A., Thitamadee S., Campilho A., Sebastian J., Bowman J.L., Helariutta Y., Benfey P.N. Cell signalling by microRNA165/6 directs gene dose-dependent root cell fate. Nature. 2010;465:316–321. doi: 10.1038/nature08977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaffey N., Cholewa E., Regan S., Sundberg B. Secondary xylem development in Arabidopsis: a model for wood formation. Physiol. Plant. 2002;114:594–600. doi: 10.1034/j.1399-3054.2002.1140413.x. [DOI] [PubMed] [Google Scholar]
- Chehab E.W., Yao C., Henderson Z., Kim S., Braam J. Arabidopsis touch-induced morphogenesis is jasmonate mediated and protects against pests. Curr. Biol. 2012;22:701–706. doi: 10.1016/j.cub.2012.02.061. [DOI] [PubMed] [Google Scholar]
- Chen M.K., Wilson R.L., Palme K., Ditengou F.A., Shpak E.D. ERECTA family genes regulate auxin transport in the shoot apical meristem and forming leaf primordia. Plant Physiol. 2013;162:1978–1991. doi: 10.1104/pp.113.218198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho H., Ryu H., Rho S., Hill K., Smith S., Audenaert D., Park J., Han S., Beeckman T., Bennett M.J., Hwang D., De Smet I., Hwang I. A secreted peptide acts on BIN2-mediated phosphorylation of ARFs to potentiate auxin response during lateral root development. Nat. Cell Biol. 2014;16:66–76. doi: 10.1038/ncb2893. [DOI] [PubMed] [Google Scholar]
- Coutand C., Dupraz C., Jaouen G., Ploquin S., Adam B. Mechanical stimuli regulate the allocation of biomass in trees: demonstration with young Prunus avium trees. Ann. Bot. 2008;101:1421–1432. doi: 10.1093/aob/mcn054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Rybel B., Möller B., Yoshida S., Grabowicz I., Barbier de Reuille P., Boeren S., Smith R.S., Borst J.W., Weijers D. A bHLH complex controls embryonic vascular tissue establishment and indeterminate growth in Arabidopsis. Dev. Cell. 2013;24:426–437. doi: 10.1016/j.devcel.2012.12.013. [DOI] [PubMed] [Google Scholar]
- De Rybel B., Breda A.S., Weijers D. Prenatal plumbing-vascular tissue formation in the plant embryo. Physiol. Plant. 2014a;151:126–133. doi: 10.1111/ppl.12091. [DOI] [PubMed] [Google Scholar]
- De Rybel B., Adibi M., Breda A.S., Wendrich J.R., Smit M.E., Novák O., Yamaguchi N., Yoshida S., Van Isterdael G., Palovaara J., Nijsse B., Boekschoten M.V., Hooiveld G., Beeckman T., Wagner D., Ljung K., Fleck C., Weijers D. Plant development. Integration of growth and patterning during vascular tissue formation in Arabidopsis. Science. 2014b;345:1255215. doi: 10.1126/science.1255215. [DOI] [PubMed] [Google Scholar]
- Dolan L., Janmaat K., Willemsen V., Linstead P., Poethig S., Roberts K., Scheres B. Cellular organisation of the Arabidopsis thaliana root. Development. 1993;119:71–84. doi: 10.1242/dev.119.1.71. [DOI] [PubMed] [Google Scholar]
- Emery J.F., Floyd S.K., Alvarez J., Eshed Y., Hawker N.P., Izhaki A., Baum S.F., Bowman J.L. Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 2003;13:1768–1774. doi: 10.1016/j.cub.2003.09.035. [DOI] [PubMed] [Google Scholar]
- Escamez S., Tuominen H. Programmes of cell death and autolysis in tracheary elements: when a suicidal cell arranges its own corpse removal. J. Exp. Bot. 2014;65:1313–1321. doi: 10.1093/jxb/eru057. [DOI] [PubMed] [Google Scholar]
- Eshed Y., Baum S.F., Perea J.V., Bowman J.L. Establishment of polarity in lateral organs of plants. Curr. Biol. 2001;11:1251–1260. doi: 10.1016/s0960-9822(01)00392-x. [DOI] [PubMed] [Google Scholar]
- Etchells J.P., Turner S.R. The PXY-CLE41 receptor ligand pair defines a multifunctional pathway that controls the rate and orientation of vascular cell division. Development. 2010;137:767–774. doi: 10.1242/dev.044941. [DOI] [PubMed] [Google Scholar]
- Etchells J.P., Provost C.M., Turner S.R. Plant vascular cell division is maintained by an interaction between PXY and ethylene signalling. PLoS Genet. 2012;8:e1002997. doi: 10.1371/journal.pgen.1002997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etchells J.P., Provost C.M., Mishra L., Turner S.R. WOX4 and WOX14 act downstream of the PXY receptor kinase to regulate plant vascular proliferation independently of any role in vascular organisation. Development. 2013;140:2224–2234. doi: 10.1242/dev.091314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher K., Turner S. PXY, a receptor-like kinase essential for maintaining polarity during plant vascular-tissue development. Curr. Biol. 2007;17:1061–1066. doi: 10.1016/j.cub.2007.05.049. [DOI] [PubMed] [Google Scholar]
- Friml J., Vieten A., Sauer M., Weijers D., Schwarz H., Hamann T., Offringa R., Jürgens G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature. 2003;426:147–153. doi: 10.1038/nature02085. [DOI] [PubMed] [Google Scholar]
- Fukuda H. Programmed cell death during vascular system formation. Cell Death Differ. 1997;4:684–688. doi: 10.1038/sj.cdd.4400310. [DOI] [PubMed] [Google Scholar]
- Guo Y., Qin G., Gu H., Qu L.J. Dof5.6/HCA2, a Dof transcription factor gene, regulates interfascicular cambium formation and vascular tissue development in Arabidopsis. Plant Cell. 2009;21:3518–3534. doi: 10.1105/tpc.108.064139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gälweiler L., Guan C., Müller A., Wisman E., Mendgen K., Yephremov A., Palme K. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science. 1998;282:2226–2230. doi: 10.1126/science.282.5397.2226. [DOI] [PubMed] [Google Scholar]
- Hardtke C.S., Berleth T. The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 1998;17:1405–1411. doi: 10.1093/emboj/17.5.1405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay A., Tsiantis M. The genetic basis for differences in leaf form between Arabidopsis thaliana and its wild relative Cardamine hirsuta. Nat. Genet. 2006;38:942–947. doi: 10.1038/ng1835. [DOI] [PubMed] [Google Scholar]
- Hay A., Tsiantis M. KNOX genes: versatile regulators of plant development and diversity. Development. 2010;137:3153–3165. doi: 10.1242/dev.030049. [DOI] [PubMed] [Google Scholar]
- Hejatko J., Ryu H., Kim G.T., Dobešová R., Choi S., Choi S.M., Souček P., Horák J., Pekárová B., Palme K., Brzobohatý B., Hwang I. The histidine kinases CYTOKININ-INDEPENDENTI and ARABIDOPSIS HISTIDINE KINASE2 and 3 regulate vascular tissue development in Arabidopsis shoots. Plant Cell. 2009;21:2008–2021. doi: 10.1105/tpc.109.066696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helariutta Y., Fukaki H., Wysocka-Diller J., Nakajima K., Jung J., Sena G., Hauser M.T., Benfey P.N. The SHORT-ROOT gene controls radial patterning of the Arabidopsis root through radial signaling. Cell. 2000;101:555–567. doi: 10.1016/s0092-8674(00)80865-x. [DOI] [PubMed] [Google Scholar]
- Higuchi M., Pischke M.S., Mähönen A.P., Miyawaki K., Hashimoto Y., Seki M., Kobayashi M., Shinozaki K., Kato T., Tabata S., Helariutta Y., Sussman M.R., Kakimoto T. In planta functions of the Arabidopsis cytokinin receptor family. Proc. Natl. Acad. Sci. USA. 2004;101:8821–8826. doi: 10.1073/pnas.0402887101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirakawa Y., Shinohara H., Kondo Y., Inoue A., Nakanomyo I., Ogawa M., Sawa S., Ohashi-Ito K., Matsubayashi Y., Fukuda H. Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proc. Natl. Acad. Sci. USA. 2008;105:15208–15213. doi: 10.1073/pnas.0808444105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirakawa Y., Kondo Y., Fukuda H. TDIF peptide signaling regulates vascular stem cell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell. 2010;22:2618–2629. doi: 10.1105/tpc.110.076083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hossain Z., McGarvey B., Amyot L., Gruber M., Jung J., Hannoufa A. DIMINUTO 1 affects the lignin profile and secondary cell wall formation in Arabidopsis. Planta. 2012;235:485–498. doi: 10.1007/s00425-011-1519-4. [DOI] [PubMed] [Google Scholar]
- Huang T., Harrar Y., Lin C., Reinhart B., Newell N.R., Talavera-Rauh F., Hokin S.A., Barton M.K., Kerstetter R.A. Arabidopsis KANADI1 acts as a transcriptional repressor by interacting with a specific cis-element and regulates auxin biosynthesis, transport, and signaling in opposition to HD-ZIPIII factors. Plant Cell. 2014;26:246–262. doi: 10.1105/tpc.113.111526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchison C.E., Li J., Argueso C., Gonzalez M., Lee E., Lewis M.W., Maxwell B.B., Perdue T.D., Schalier G.E., Alonso J.M., Ecker J.R., Kieber J.J. The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling. Plant Cell. 2006;18:3073–3087. doi: 10.1105/tpc.106.045674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibañes M., Fábregas N., Chory J., Caño-Delgado A.I. Brassinosteroid signaling and auxin transport are required to establish the periodic pattern of Arabidopsis shoot vascular bundles. Proc. Natl. Acad. Sci. USA. 2009;106:13630–13635. doi: 10.1073/pnas.0906416106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ilegems M., Douet V., Meylan-Bettex M., Uyttewaal M., Brand L., Bowman J.L., Stieger P.A. Interplay of auxin, KANADI and Class III HD-ZIP transcription factors in vascular tissue formation. Development. 2010;137:975–984. doi: 10.1242/dev.047662. [DOI] [PubMed] [Google Scholar]
- Inoue T., Higuchi M., Hashimoto Y., Seki M., Kobayashi M., Kato T., Tabata S., Shinozaki K., Kakimoto T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature. 2001;409:1060–1063. doi: 10.1038/35059117. [DOI] [PubMed] [Google Scholar]
- Ito Y., Nakanomyo I., Motose H., Iwamoto K., Sawa S., Dohmae N., Fukuda H. Dodeca-CLE peptides as suppressors of plant stem cell differentiation. Science. 2006;313:842–845. doi: 10.1126/science.1128436. [DOI] [PubMed] [Google Scholar]
- Izhaki A., Bowman J.L. KANADI and class III HD-Zip gene families regulate embryo patterning and modulate auxin flow during embryogenesis in Arabidopsis. Plant Cell. 2007;19:495–508. doi: 10.1105/tpc.106.047472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaffe M.J. Thigmomorphogenesis: The response of plant growth and development to mechanical stimulation. With special reference to Bryonia dioica. Planta. 1973;114:143–157. doi: 10.1007/BF00387472. [DOI] [PubMed] [Google Scholar]
- Ji J., Strable J., Shimizu R., Koenig D., Sinha N., Scanlon M.J. WOX4 promotes procambial development. Plant Physiol. 2010;152:1346–1356. doi: 10.1104/pp.109.149641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson K.A., Sistrunk M.L., Polisensky D.H., Braam J. Arabidopsis thaiiana responses to mechanical stimulation do not require ETR1 or EIN2. Plant Physiol. 1998;116:643–649. doi: 10.1104/pp.116.2.643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jouannet V., Brackmann K., Greb T. (Pro)cambium formation and proliferation: two sides of the same coin? Curr. Opin. Plant Biol. 2014;23:54–60. doi: 10.1016/j.pbi.2014.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kakimoto T. CKI1, a histidine kinase homolog implicated in cytokinin signal transduction. Science. 1996;274:982–985. doi: 10.1126/science.274.5289.982. [DOI] [PubMed] [Google Scholar]
- Kerstetter R.A., Bollman K., Taylor R.A., Bomblies K., Poethig R.S. KANADI regulates organ polarity in Arabidopsis. Nature. 2001;411:706–709. doi: 10.1038/35079629. [DOI] [PubMed] [Google Scholar]
- Khan M., Tabb P., Hepworth S.R. BLADE-ON-PETIOLE1 and 2 regulate Arabidopsis inflorescence architecture in conjunction with homeobox genes KNAT6 and ATH1. Plant. Signal. Behav. 2012a;7:788–792. doi: 10.4161/psb.20599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan M., Xu M., Murmu J., Tabb P., Liu Y., Storey K., McKim S.M., Douglas C.J., Hepworth S.R. Antagonistic interaction of BLADE-ON-PETIOLE1 and 2 with BREVIPEDICELLUS and PENNYWISE regulates Arabidopsis inflorescence architecture. Plant Physiol. 2012b;158:946–960. doi: 10.1104/pp.111.188573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko D., Kang J., Kiba T., Park J., Kojima M., Do J., Kim K.Y., Kwon M., Endler A., Song W.Y., Martinoia E., Sakakibara H., Lee Y. Arabidopsis ABCG14 is essential for the root-to-shoot translocation of cytokinin. Proc. Natl. Acad. Sci. USA. 2014;111:7150–7155. doi: 10.1073/pnas.1321519111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko J.H., Han K.H., Park S., Yang J. Plant body weight-induced secondary growth in Arabidopsis and its transcription phenotype revealed by whole-transcriptome profiling. Plant Physiol. 2004;135:1069–1083. doi: 10.1104/pp.104.038844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondo Y., Hirakawa Y., Kieber J.J., Fukuda H. CLE peptides can negatively regulate proto xylem vessel formation via cytokinin signaling. Plant Cell Physiol. 2011;52:37–48. doi: 10.1093/pcp/pcq129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondo Y., Ito T., Nakagami H., Hirakawa Y., Saito M., Tamaki T., Shirasu K., Fukuda H. Plant GSK3 proteins regulate xylem cell differentiation downstream of TDIF-TDR signalling. Nat. Commun. 2014;5:3504. doi: 10.1038/ncomms4504. [DOI] [PubMed] [Google Scholar]
- Kubo M., Udagawa M., Nishikubo N., Horiguchi G., Yamaguchi M., Ito J., Mimura T., Fukuda H., Demura T. Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 2005;19:1855–1860. doi: 10.1101/gad.1331305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurusu T., Kuchitsu K., Nakano M., Nakayama Y., Iida H. Plant mechanosensing and Ca2+transport. Trends Plant Sci. 2013;18:227–233. doi: 10.1016/j.tplants.2012.12.002. [DOI] [PubMed] [Google Scholar]
- Lachaud S., Catesson A.M., Bonnemain J.L. Structure and functions of the vascular cambium. C. R. Acad. Sci. III. 1999;322:633–650. doi: 10.1016/s0764-4469(99)80103-6. [DOI] [PubMed] [Google Scholar]
- Larson P.R. The Vascular Cambium: Development and Structure. Springer: 1994. p. 725. [Google Scholar]
- Lens F., Smets E., Melzer S. Stem anatomy supports Arabidopsis thaliana as a model for insular woodiness. New Phytol. 2012;193:12–17. doi: 10.1111/j.1469-8137.2011.03888.x. [DOI] [PubMed] [Google Scholar]
- Lev-Yadun S. Induction of schlereid differentiation in the pith of Arabidopsis thaliana (L.) Heyhn. J. Exp. Bot. 1994;45:1845–1849. [Google Scholar]
- Liebsch D., Sunaryo W., Holmlund M., Norberg M., Zhang J., Hall H.C., Helizon H., Jin X., Helariutta Y., Nilsson O., Polie A., Fischer U. Class I KNOX transcription factors promote differentiation of cambial derivatives into xylem fibers in the Arabidopsis hypocotyl. Development. 2014;141:4311–4319. doi: 10.1242/dev.111369. [DOI] [PubMed] [Google Scholar]
- Little C.H.A., MacDonald J.E., Olsson O. Involvement of indole-3-acetic acid in fascicular and interfascicular cambial growth and interfascicular extraxylary fiber differentiation in Arabidopsis thaliana inflorescence stems. Int. J. Plant Sci. 2002;16:519–529. [Google Scholar]
- Matsumoto-Kitano M., Kusumoto T., Tarkowski R., Kinoshita-Tsujimura K., Václavíková K., Miyawaki K., Kakimoto T. Cytokinins are central regulators of cambial activity. Proc. Natl. Acad. Sci. USA. 2008;105:20027–20031. doi: 10.1073/pnas.0805619105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazur E., Kurczynska E.U. Rays, intrusive growth, and storied cambium in the inflorescence stems of Arabidopsis thaliana (L.) Heynh. Protoplasma. 2012;249:217–220. doi: 10.1007/s00709-011-0266-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazur E., Kurczyńska E.U., Friml J. Cellular events during interfascicular cambium ontogenesis in inflorescence stems of Arabidopsis. Protoplasma. 2014;251:1125–1139. doi: 10.1007/s00709-014-0620-5. [DOI] [PubMed] [Google Scholar]
- McCarthy R.L., Zhong R., Ye Z.H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009;50:1950–1964. doi: 10.1093/pcp/pcp139. [DOI] [PubMed] [Google Scholar]
- McConnell J.R., Barton M.K. Leaf polarity and meristem formation in Arabidopsis. Development. 1998;125:2935–2942. doi: 10.1242/dev.125.15.2935. [DOI] [PubMed] [Google Scholar]
- Melzer S., Lens F., Gennen J., Vanneste S., Rohde A., Beeckman T. Flowering-time genes modulate meristem determinacy and growth form in Arabidopsis thaliana. Nat. Genet. 2008;40:1489–1492. doi: 10.1038/ng.253. [DOI] [PubMed] [Google Scholar]
- Merchante C., Alonso J.M., Stepanova A.N. Ethylene signaling: simple ligand, complex regulation. Curr. Opin. Plant Biol. 2013;16:554–560. doi: 10.1016/j.pbi.2013.08.001. [DOI] [PubMed] [Google Scholar]
- Merelo P., Xie Y., Brand L., Ott F., Weigel D., Bowman J.L., Heisler M.G., Wenkel S. Genome-wide identification of KANADI1 target genes. PLoS One. 2013;8:e77341. doi: 10.1371/journal.pone.0077341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyerowitz E.M. Arabidopsis, a useful weed. Cell. 1989;56:263–269. doi: 10.1016/0092-8674(89)90900-8. [DOI] [PubMed] [Google Scholar]
- Mitsuda N., Iwase A., Yamamoto H., Yoshida M., Seki M., Shinozaki K., Ohme-Takagi M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell. 2007;19:270–280. doi: 10.1105/tpc.106.047043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyashima S., Koi S., Hashimoto T., Nakajima K. Non-cellautonomous microRNA165 acts in a dose-dependent manner to regulate multiple differentiation status in the Arabidopsis root. Development. 2011;138:2303–2313. doi: 10.1242/dev.060491. [DOI] [PubMed] [Google Scholar]
- Miyashima S., Sebastian J., Lee J.Y., Helariutta Y. Stem cell function during plant vascular development. EMBO J. 2013;32:178–193. doi: 10.1038/emboj.2012.301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motose H., Fukuda H., Sugiyama M. Involvement of local intercellular communication in the differentiation of zinnia mesophyll cells into tracheary elements. Planta. 2001;213:121–131. doi: 10.1007/s004250000482. [DOI] [PubMed] [Google Scholar]
- Muñiz L., Minguet E.G., Singh S.K., Pesquet E., Vera-Sirera F., Moreau-Courtois C.L., Carbonell J., Blázquez M.A., Tuominen H. ACAULIS5 controls Arabidopsis xylem specification through the prevention of premature cell death. Development. 2008;135:2573–2582. doi: 10.1242/dev.019349. [DOI] [PubMed] [Google Scholar]
- Murren C.J., Pigliucci M. Morphological responses to simulated wind in the genus Brassica (Brassicaceae): allopolyploids and their parental species. Am. J. Bot. 2005;92:810–818. doi: 10.3732/ajb.92.5.810. [DOI] [PubMed] [Google Scholar]
- Mähönen A.P., Bonke M., Kauppinen L., Riikonen M., Benfey P.N., Helariutta Y. A novel two-component hybrid molecule regulates vascular morphogenesis of the Arabidopsis root. Genes Dev. 2000;14:2938–2943. doi: 10.1101/gad.189200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mähönen A.P., Bishopp A., Higuchi M., Nieminen K.M., Kinoshita K., Törmäkangas K., Ikeda Y., Oka A., Kakimoto T., Helariutta Y. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science. 2006a;311:94–98. doi: 10.1126/science.1118875. [DOI] [PubMed] [Google Scholar]
- Mähönen A.P., Higuchi M., Törmäkangas K., Miyawaki K., Pischke M.S., Sussman M.R., Helariutta Y., Kakimoto T. Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis. Curr. Biol. 2006b;16:1116–1122. doi: 10.1016/j.cub.2006.04.030. [DOI] [PubMed] [Google Scholar]
- Nakajima K., Sena G., Nawy T., Benfey P.N. Intercellular movement of the putative transcription factor SHR in root patterning. Nature. 2001;413:307–311. doi: 10.1038/35095061. [DOI] [PubMed] [Google Scholar]
- Ohashi-Ito K., Bergmann D.C. Regulation of the Arabidopsis root vascular initial population by LONESOME HIGHWAY. Development. 2007;134:2959–2968. doi: 10.1242/dev.006296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohashi-Ito K., Oda Y., Fukuda H. Arabidopsis VASCULAR-RELATED NAC-DOMAIN6 directly regulates the genes that govern programmed cell death and secondary wall formation during xylem differentiation. Plant Cell. 2010;22:3461–3473. doi: 10.1105/tpc.110.075036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohashi-Ito K., Matsukawa M., Fukuda H. An atypical bHLH transcription factor regulates early xylem development downstream of auxin. Plant Cell Physiol. 2013;54:398–405. doi: 10.1093/pcp/pct013. [DOI] [PubMed] [Google Scholar]
- Ohashi-Ito K., Saegusa M., Iwamoto K., Oda Y., Katayama H., Kojima M., Sakakibara H., Fukuda H. A bHLH complex activates vascular cell division via cytokinin action in root apical meristem. Curr. Biol. 2014;24:2053–2058. doi: 10.1016/j.cub.2014.07.050. [DOI] [PubMed] [Google Scholar]
- Okada K., Ueda J., Kornaki M.K., Bell C.J., Shimura Y. Requirement of the auxin polar transport system in early stages of Arabidopsis floral bud formation. Plant Cell. 1991;3:677–684. doi: 10.1105/tpc.3.7.677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palovaara J., Saiga S., Weijers D. Transcriptomics approaches in the early Arabidopsis embryo. Trends Plant Sci. 2013;18:514–521. doi: 10.1016/j.tplants.2013.04.011. [DOI] [PubMed] [Google Scholar]
- Parker G., Schofield R., Sundberg B., Turner S. Isolation of COV1, a gene involved in the regulation of vascular patterning in the stem of Arabidopsis. Development. 2003;130:2139–2148. doi: 10.1242/dev.00441. [DOI] [PubMed] [Google Scholar]
- Paul-Victor C., Rowe N. Effect of mechanical perturbation on the biomechanics, primary growth and secondary tissue development of inflorescence stems of Arabidopsis thaliana. Ann. Bot. 2011;107:209–218. doi: 10.1093/aob/mcq227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pesquet E., Zhang B., Gorzsás A., Puhakainen T., Serk H., Escamez S., Barbier O., Gerber L., Courtois-Moreau C., Alatalo E., Paulin L., Kangasjärvi J., Sundberg B., Goffner D., Tuominen H. Non-cell-autonomous postmortem lignification of tracheary elements in Zinnia elegans. Plant Cell. 2013;25:1314–1328. doi: 10.1105/tpc.113.110593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pineau C., Freydier A., Ranocha P., Jauneau A., Turner S., Lemonnier G., Renou J.P., Tarkowski P., Sandberg G., Jouanin L., Sundberg B., Boudet A.M., Goffner D., Pichon M. hca: an Arabidopsis mutant exhibiting unusual cambial activity and altered vascular patterning. Plant J. 2005;44:271–289. doi: 10.1111/j.1365-313X.2005.02526.x. [DOI] [PubMed] [Google Scholar]
- Prigge M.J., Otsuga D., Alonso J.M., Ecker J.R., Drews G.N., Clark S.E. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell. 2005;17:61–76. doi: 10.1105/tpc.104.026161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ragni L., Nieminen K., Pacheco-Villalobos D., Sibout R., Schwechheimer C., Hardtke C.S. Mobile gibberellin directly stimulates Arabidopsis hypocotyl xylem expansion. Plant Cell. 2011;23:1322–1336. doi: 10.1105/tpc.111.084020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ragni L., Hardtke C.S. Small but thick enough - the Arabidopsis hypocotyl as a model to study secondary growth. Physiol. Plant. 2014;151:164–171. doi: 10.1111/ppl.12118. [DOI] [PubMed] [Google Scholar]
- Sanchez P., Nehlin L., Greb T. From thin to thick: major transitions during stem development. Trends Plant Sci. 2012;17:113–121. doi: 10.1016/j.tplants.2011.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sankar M., Nieminen K., Ragni L., Xenarios I., Hardtke C.S. Automated quantitative histology reveals vascular morphodynamics during Arabidopsis hypocotyl secondary growth. Elife. 2014;3:e01567. doi: 10.7554/eLife.01567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkar A.K., Luijten M., Miyashima S., Lenhard M., Hashimoto T., Nakajima K., Scheres B., Heidstra R., Laux T. Conserved factors regulate signalling in Arabidopsis thaiiana shoot and root stem cell organizers. Nature. 2007;446:811–814. doi: 10.1038/nature05703. [DOI] [PubMed] [Google Scholar]
- Scarpella E., Meijer A.H. Pattern formation in the vascular system of monocot and dicot plant species. New Phytol. 2004;164:209–242. doi: 10.1111/j.1469-8137.2004.01191.x. [DOI] [PubMed] [Google Scholar]
- Scheres B., Wolkenfelt H., Willemsen V., Terlouw M., Lawson E., Dean C., Weisbeek P. Embryonic origin of the Arabidopsis primary root and root meristem initials. Development. 1994;120:2475–2487. [Google Scholar]
- Scheres B., Di Laurenzio L., Willemsen V., Hauser M.T., Janmaat K., Weisbeek P., Benfey P.N. Mutations affecting the radial organisation of the Arabidopsis root display specific defects throughout the embryonic axis. Development. 1995;121:53–62. [Google Scholar]
- Schlereth A., Möller B., Liu W., Kientz M., Flipse J., Rademacher E.H., Schmid M., Jürgens G., Weijers D. MONOPTEROS controls embryonic root initiation by regulating a mobile transcription factor. Nature. 2010;464:913–916. doi: 10.1038/nature08836. [DOI] [PubMed] [Google Scholar]
- Schoof H., Lenhard M., Haecker A., Mayer K.F., Jürgens G., Laux T. The stem cell population of Arabidopsis shoot me riste ms in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell. 2000;100:635–644. doi: 10.1016/s0092-8674(00)80700-x. [DOI] [PubMed] [Google Scholar]
- Schweingruber F.H., Börner A., Schulze E.D. Atlas of Stem Anatomy in Herbs. Shrubs and Trees; Springer: 2011. p. 495. [Google Scholar]
- Sehr E.M., Agusti J., Lehner R., Farmer E.E., Schwarz M., Greb T. Analysis of secondary growth in the Arabidopsis shoot reveals a positive role of jasmonate signalling in cambium formation. Plant J. 2010;63:811–822. doi: 10.1111/j.1365-313X.2010.04283.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheard L.B., Tan X., Mao H., Withers J., Ben-Nissan G., Hinds T.R., Kobayashi Y., Hsu F.F., Sharon M., Browse J., He S.Y., Rizo J., Howe G.A., Zheng N. Jasmonate perception by inositolphosphate-potentiated COI1-JAZ co-receptor. Nature. 2010;468:400–405. doi: 10.1038/nature09430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirakawa M., Ueda H., Koumoto Y., Fuji K., Nishiyama C., Kohchi T., Hara-Nishimura I., Shimada T. CONTINUOUS VASCULAR RING (COV1) is a trans-Golgi network-localized membrane protein required for Golgi morphology and vacuolar protein sorting. Plant Cell Physiol. 2014;55:764–772. doi: 10.1093/pcp/pct195. [DOI] [PubMed] [Google Scholar]
- Sibout R., Plantegenet S., Hardtke C.S. Flowering as a condition for xylem expansion in Arabidopsis hypocotyl and root. Curr. Biol. 2008;18:458–463. doi: 10.1016/j.cub.2008.02.070. [DOI] [PubMed] [Google Scholar]
- Smith H.M.S., Hake S. The interaction of two homeobox genes, BREVIPEDICELLUS and PENNYWISE, regulates internode patterning in the Arabidopsis inflorescence. Plant Cell. 2003;15:1717–1727. doi: 10.1105/tpc.012856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparks E., Wachsman G., Benfey P.N. Spatiotemporal signalling in plant development. Nat. Rev. Genet. 2013;14:631–644. doi: 10.1038/nrg3541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suer S., Agusti J., Sanchez P., Schwarz M., Greb T. WOX4 imparts auxin responsiveness to cambium cells in Arabidopsis. Plant Cell. 2011;23:3247–3259. doi: 10.1105/tpc.111.087874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabata R., Sumida K., Yoshii T., Ohyama K., Shinohara H., Matsubayashi Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science. 2014;346:343–346. doi: 10.1126/science.1257800. [DOI] [PubMed] [Google Scholar]
- Talbert P.B., Adler H.T., Parks D.W., Comai L. The REVOLUTA gene is necessary for apical meristem development and for limiting cell divisions in the leaves and stems of Arabidopsis thaiiana. Development. 1995;121:2723–2735. doi: 10.1242/dev.121.9.2723. [DOI] [PubMed] [Google Scholar]
- Taylor-Teeples M., Lin L., de Lucas M., Turco G., Toal T.W., Gaudinier A., Young N.F., Trabucco G.M., Veling M.T., Lamothe R., Handakumbura P.P., Xiong G., Wang C., Corwin J., Tsoukalas A., Zhang L., Ware D., Pauly M., Kliebenstein D.J., Dehesh K., Tagkopoulos I., Breton G., Pruneda-Paz J.L., Ahnert S.E., Kay S.A., Hazen S.P., Brady S.M. An Arabidopsis gene regulatory network for secondary cell wall synthesis. Nature. 2015;517:571–575. doi: 10.1038/nature14099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Telewski F.W., Jaffe M.J. Thigmomorphogenesis: Field and laboratory studies of Abies fraseri in response to wind or mechanical perturbation. Physiol. Plant. 1986;66:211–218. doi: 10.1111/j.1399-3054.1986.tb02411.x. [DOI] [PubMed] [Google Scholar]
- Torii K.U., Mitsukawa N., Oosumi T., Matsuura Y., Yokoyama R., Whittier R.F., Komeda Y. The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant Cell. 1996;8:735–746. doi: 10.1105/tpc.8.4.735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchida N., Lee J.S., Horst R.J., Lai H.H., Kajita R., Kakimoto T., Tasaka M., Torii K.U. Regulation of inflorescence architecture by intertissue layer ligand-receptor communication between endodermis and phloem. Proc. Natl. Acad. Sci. USA. 2012;109:6337–6342. doi: 10.1073/pnas.1117537109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchida N., Tasaka M. Regulation of plant vascular stem cells by endodermis-derived EPFL-family peptide hormones and phloem-expressed ERECTA-family receptor kinases. J. Exp. Bot. 2013;64:5335–5343. doi: 10.1093/jxb/ert196. [DOI] [PubMed] [Google Scholar]
- van den Berg C., Willemsen V., Hendriks G., Weisbeek P., Scheres B. Short-range control of cell differentiation in the Arabidopsis root meristem. Nature. 1997;390:287–289. doi: 10.1038/36856. [DOI] [PubMed] [Google Scholar]
- Vatén A., Dettmer J., Wu S., Stierhof Y.D., Miyashima S., Yadav S.R., Roberts C.J., Campilho A., Bulone V., Lichtenberger R., Lehesranta S., Mähönen A.P., Kim J.Y., Jokitalo E., Sauer N., Scheres B., Nakajima K., Carlsbecker A., Gallagher K.L., Helariutta Y. Callose biosynthesis regulates symplastic trafficking during root development. Dev. Cell. 2011;21:1144–1155. doi: 10.1016/j.devcel.2011.10.006. [DOI] [PubMed] [Google Scholar]
- Vlad D., Kierzkowski D., Rast M.I., Vuolo F., Dello Ioio R., Galinha C., Gan X., Hajheidari M., Hay A., Smith R.S., Huijser P., Bailey C.D., Tsiantis M. Leaf shape evolution through duplication, regulatory diversification, and loss of a homeobox gene. Science. 2014;343:780–783. doi: 10.1126/science.1248384. [DOI] [PubMed] [Google Scholar]
- Wang J., Kucukoglu M., Zhang L., Chen P., Decker D., Nilsson O., Jones B., Sandberg G., Zheng B. The Arabidopsis LRR-RLK, PXC1, is a regulator of secondary wall formation correlated with the TDIF-PXY/TDR-WOX4 signaling pathway. BMC Plant. Biol. 2013;13:94. doi: 10.1186/1471-2229-13-94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitford R., Fernandez A., De Groodt R., Ortega E., Hilson P. Plant CLE peptides from two distinct functional classes synergistically induce division of vascular cells. Proc. Natl. Acad. Sci. USA. 2008;105:18625–18630. doi: 10.1073/pnas.0809395105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiang D., Venglat P., Tibiche C., Yang H., Risseeuw E., Cao Y., Babic V., Cloutier M., Keller W., Wang E., Selvaraj G., Datla R. Genome-wide analysis reveals gene expression and metabolic network dynamics during embryo development in Arabidopsis. Plant Physiol. 2011;156:346–356. doi: 10.1104/pp.110.171702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yaginuma H., Hirakawa Y., Kondo Y., Ohashi-Ito K., Fukuda H. A novel function of TDIF-related peptides: promotion of axillary bud formation. Plant Cell Physiol. 2011;52:1354–1364. doi: 10.1093/pcp/pcr081. [DOI] [PubMed] [Google Scholar]
- Yamada H., Suzuki T., Terada K., Takei K., Ishikawa K., Miwa K., Yamashino T., Mizuno T. The Arabidopsis AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin signals across the membrane. Plant Cell Physiol. 2001;42:1017–1023. doi: 10.1093/pcp/pce127. [DOI] [PubMed] [Google Scholar]
- Yokoyama A., Yamashino T., Amano Y., Tajima Y., Imamura A., Sakakibara H., Mizuno T. Type-B ARR transcription factors, ARR10 and ARR12, are implicated in cytokinin-mediated regulation of protoxylem differentiation in roots of Arabidopsis thaliana. Plant Cell Physiol. 2007;48:84–96. doi: 10.1093/pcp/pcl040. [DOI] [PubMed] [Google Scholar]
- Zhang J., Elo A., Helariutta Y. Arabidopsis as a model for wood formation. Curr. Opin. Biotechnol. 2011;22:293–299. doi: 10.1016/j.copbio.2010.11.008. [DOI] [PubMed] [Google Scholar]
- Zhong R., Taylor J.J., Ye Z.H. Transformation of the collateral vascular bundles into amphivasal vascular bundles in an Arabidopsis mutant. Plant Physiol. 1999;120:53–64. doi: 10.1104/pp.120.1.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Ye Z.H. Alteration of auxin polar transport in the Arabidopsis ifl1 mutants. Plant Physiol. 2001;126:549–563. doi: 10.1104/pp.126.2.549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Ye Z.H. amphivasal vascular bundle 1, a gain-of-function mutation of the IFL1/REV gene, is associated with alterations in the polarity of leaves, stems and carpels. Plant Cell Physiol. 2004;45:369–385. doi: 10.1093/pcp/pch051. [DOI] [PubMed] [Google Scholar]
- Zhong R., Richardson E.A., Ye Z.H. The MYB46 transcription factor is a direct target of SND1 and regulates secondary wall biosynthesis in Arabidopsis. Plant Cell. 2007;19:2776–2792. doi: 10.1105/tpc.107.053678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Lee C., Ye Z.H. Global analysis of direct targets of secondary wall NAC master switches in Arabidopsis. Mol. Plant. 2010;3:1087–1103. doi: 10.1093/mp/ssq062. [DOI] [PubMed] [Google Scholar]
- Zhong R., Ye Z.H. MYB46 and MYB83 bind to the SMRE sites and directly activate a suite of transcription factors and secondary wall biosynthetic genes. Plant Cell Physiol. 2012;53:368–380. doi: 10.1093/pcp/pcr185. [DOI] [PubMed] [Google Scholar]
- Zhou G.K., Kubo M., Zhong R., Demura T., Ye Z.H. Over-expression of miR165 affects apical me riste m formation, organ polarity establishment and vascular development in Arabidopsis. Plant Cell Physiol. 2007;48:391–404. doi: 10.1093/pcp/pcm008. [DOI] [PubMed] [Google Scholar]
- Zhou J., Wang X., Lee J.Y., Lee J.Y. Cell-to-cell movement of two interacting AT-hook factors in Arabidopsis root vascular tissue patterning. Plant Cell. 2013;25:187–201. doi: 10.1105/tpc.112.102210. [DOI] [PMC free article] [PubMed] [Google Scholar]