

Abstract
The infant microbiome plays an essential role in human health and its assembly is determined by maternal– offspring exchanges of microbiota. This process is affected by several practices, including Cesarean section (C-section), perinatal antibiotics, and formula feeding, that have been linked to increased risks of metabolic and immune diseases. Here we review recent knowledge about the impacts on infant microbiome assembly, discuss preventive and restorative strategies to ameliorate the effects of these impacts, and highlight where research is needed to advance this field and improve the health of future generations.
The infant microbiome and immune and metabolic health
The suite of genes provided by microorganisms, or the microbiota, living in and on the human body is known as the human microbiome [1]. As the microbiome interacts dynamically with its host and environment, its composition varies markedly over time and between individuals [2]. The bacterial genes comprising our microbiome outnumber human genes by more than 100-fold and have such a broad influence on physiological regulation that they have been recognized as another organ [3].
Our previously limited view of human–microbe interactions, strictly as pathogens causing infectious diseases, has undergone rapid and dramatic expansion over the past two decades. While we now appreciate the essential role of the microbiota as commensals and symbionts integral to immune [4] and metabolic [5] health, we are just beginning to understand how and when these microorganisms assemble and the early-life factors that disrupt their natural ecological succession. Appreciation of the determinants and progression of the initial microbiome assemblage, particularly that of the gut (which is intimately involved in regulating our health), will afford insights into how the microbiome can be manipulated to improve health.
The initial development and maturation of the neonatal microbiome is largely determined by maternal–offspring exchanges of microbiota. Disrupting the mother-to-newborn transmission of bacteria by C-section delivery may increase the risk of celiac disease [6,7], asthma [8–11], type 1 diabetes [12,13], and obesity [14–16] in the offspring. Initial epidemiological evidence also indicates that disrupting microbial exchange through the use of antibiotics in pregnancy may increase offspring risk of childhood obesity [17] and asthma [18]. One study found that children exposed to prenatal antibiotics in the second or third trimester had 84% higher risk of obesity compared with unexposed children [17]. In the same study, C-section was associated with 46% higher offspring risk of childhood obesity. After birth, breastfeeding introduces new microbial communities and stimulates the maturation of the neonatal gut microbiome [19,20]. The use of infant formula compared with breast milk has been found to impair the proper development of the neonatal immune system [21] and alter metabolism later in life [22]. While more research is needed to determine whether antibiotics, C-section delivery, and formula feeding are causally associated with autoimmune and metabolic diseases and, if so, the magnitude of these associations, the best available evidence suggests that these practices that compromise the microbial colonization of the newborn gut should be used prudently and followed by measures to restore the natural composition of the microbiome.
Here we review the natural colonization and assembly of the neonatal microbiome, with particular focus on the gut, and the impacts exerted by antibiotics, C-section delivery, and formula feeding. We then discuss potential strategies for prevention and restoration of these microbiome insults. Lastly, throughout the review we indicate where further research regarding the acquisition, development, perturbation, and restoration of the neonatal microbiome is needed.
The maternal microbiome during pregnancy
Pregnancy affects all body systems, including the maternal microbiome. Gestational changes in the vaginal [23,24] and intestinal [25] microbiome are of particular relevance because these body sites are responsible for vertical microbial transmission to the newborn during vaginal delivery.
The composition of the vaginal microbiota changes throughout the course of pregnancy. In a cross-sectional study of 24 healthy gravid women at 18–40 weeks of gestation, Aagaard et al. found that, compared with non-pregnant women, pregnant women had lower vaginal bacterial diversity, with dominance of lactobacilli, Clostridiales, Bacteroidales, and Actinomycetales [23]. Furthermore, specific Lactobacillus species (L. iners, L. crispatus, L. jensenii, and L. johnsonii) had higher prevalence during later gestational ages [23]. A longitudinal study using sequence-based techniques analyzed vaginal samples serially collected from 22 non-pregnant and 32 pregnant participants. This study confirmed an increasing relative abundance of Lactobacillus species (L. vaginalis, L. crispatus, L. jensenii, and L. gasseri) as a function of gestational age and found that the vaginal microbiota became less diverse [23,24].
Beyond the vaginal microbiome, there is evidence from one Finnish cohort that the maternal gut microbiome also changes during the course of pregnancy. Relying on self-collected specimens from the first and third trimesters of 91 healthy pregnant women, Koren et al. reported that, as women progress through pregnancy, the bacterial diversity decreases [25]. Overall, the authors found that greater gestational age was associated with the presence of more high-energy-yielding fecal microbiota, which are typically characteristic of microbial communities found in individuals with metabolic syndrome [25]. Specifically, the proportion of proinflammatory Proteobacteria, including species of the Enterobacteriaceae family and Streptococcus genus, decreased from the first trimester to the third trimester, while the proportion of anti-inflammatory Faecalibacterium prausnitzii increased. These changes were independent of pre-pregnancy body weight, gestational diabetes, diet, and antibiotic use, suggesting that they were due to normal pregnancy-related alterations to the maternal endocrine and immune systems. A caveat in this study, however, was the use of primers in the V1V2 region that discriminate against bifidobacteria [26]. More prospective studies of diverse populations are needed to confirm these findings and determine whether they are modified by demographic or lifestyle factors.
The implications of the maternal gut and vaginal micro-biota changes for the health of the mother and her offspring are unclear, but gestational changes in the vaginal and fecal microbiota are likely to be part of an adaptive response to protect and promote the health of the fetus and provide the newborn with a specific microbial inoculum at birth, before exposure to other environmental microbes. It is worth noting that the composition of the microbial communities in the maternal gut and vagina are not independent of each other. A previous study of women at 35–37 weeks of gestation showed that many bacteria, including Lactobacillus and Bifidobacterium species, are shared between the rectum and the vagina [27].
In the vagina, the increasing presence of lactobacilli through the course of gestation helps maintain a low pH, thereby limiting bacterial diversity and preventing bacteria from ascending to the uterus, where they can infect the amniotic fluid, placenta, and fetus [28,29]. In addition to the anti-infective effect of lactobacilli metabolic products, such as hydrogen peroxide and bacteriocins [30], some species (L. jensenii and L. crispatus) have surface-associated proteins that may prevent pathogens, including Neisseria gonorrhea, from infecting the mother [31]. These species have also been found to reduce the incidence of bacterial vaginosis (BV) [32]. Evidence from murine models suggests that maternal vaginal lactobacilli colonize and play a digestive role in the offspring's gut [33], but research on this topic is still needed in humans.
Pregnancy-related shifts in the maternal gut microbiota may also be adaptive for the health of the mother and newborn. Koren et al. observed that gestational-age-related increases in gut bacteria promote weight gain in mice [25], suggesting that these microbes have coevolved with their hosts to allow greater energy harvest during pregnancy to support the growth of the mother and fetus. Furthermore, the transfer of these maternal gut microbiota to the newborn during vaginal delivery may afford the neonate immediate access at birth to microbiota that allow maximal energy harvest during the incipient hours of life [25,34]. Consistent with the hypothesis that newborns pick up fecal bacteria during birth, mice developmental studies have shown greater diversity in the microbiome of newborn mice than could be attributed to vaginal transfer alone [33]. Furthermore, cultivation-based human studies have found specific Bifidobacterium species from the mother's prenatal feces in the feces of infants born vaginally but not by C-section [35]. Culture-independent studies are needed to catalogue the fecally transmitted microbiota and to examine when and how they colonize the infant gut and educate the developing immune and metabolic systems.
Is there prenatal maternal–fetal exchange of microbiota?
The intrauterine environment during healthy pregnancy has been presumed to be free of bacteria (see [36] for a review and critique of this perspective), although recent evidence of microbes present in the amniotic fluid [37–40], umbilical cord blood [41], fetal membranes [42], and placenta [43–49] of healthy term pregnancies after both vaginal and C-section delivery has challenged this belief. However, caution is warranted in the interpretation of results from studies using high-throughput sequencing on low biomass samples such as those from the placenta and other intrauterine environs, since the risk of contamination of bacteria-free samples is high and strict controls are needed to exclude contamination.
One study reported that DNA of the common gut bacteria Lactobacillus and Bifidobacterium spp. was found in all 29 of 29 placental biopsies collected at term after elective C-section without rupture of membranes or signs of maternal infection [48]. While it remains unknown whether there are live bacteria in the intrauterine environment or whether the bacterial DNA found is merely the phagocytic remnant of a local bacterial infection [50], there is circumstantial evidence suggesting that bacteria can be transmitted from the mother to the fetus. Pregnant mice administered Enterococcus faecium transferred a low level of this strain to the amniotic fluid and fetal intestine [41]. Moreover, a randomized controlled trial has shown that consumption of probiotics during pregnancy can alter the expression of Toll-like receptor (TLR) genes in the placenta and meconium of neonates [49]. However, before conclusions can be drawn about in utero exchange of bacteria during healthy pregnancies, empirical research is needed to determine whether the bacteria in the intrauterine environment of healthy pregnancies are alive, where they originate, and whether they prime fetal immune and metabolic development. Sterile sampling and sequencing techniques will be of paramount importance for these studies to exclude bias from contamination. Continued research in this realm has potential implications for probiotic prophylaxis of pregnancy complications and prevention of neonatal pathologies.
Birthing and early neonatal microbiome assembly
Regardless of intrauterine exposure, massive bacterial colonization of the newborn occurs at birth upon exposure to vaginal, fecal, and skin microbiota [19,51,52]. During vaginal delivery, facultative anaerobic species such as Escherichia coli, Staphylococcus, and Streptococcus colonize the infant gut and produce anaerobic environs in the first few days of life that allow strict anaerobes like Bacteroides and Bifidobacterium spp. to thrive [33].
After birth, maternal breast milk promotes the colonization and maturation of the infant gut microbiome. Breast milk contains bacteria [53–61] that have been shown to vary from colostrum to late lactation [62] and by gestational age [62], maternal health status [56], and delivery mode [56,62]. The breast-milk microbiota is dominated by a few genera (Staphylococcus, Streptococcus, Serratia, Pseudomonas, Corynebacterium, Ralstonia, Propionibacterium, Sphingomonas, and Bradyrhizobiaceae) [55]. Bifidobacterium and Lactobacillus spp. are also found in breast milk and transfer of these microbes to the neonatal gut has been demonstrated using culture-and strain-level discrimination [60,61], indicating that breastfeeding is a postnatal route of mother–infant microbial exchange. However, the origin of these microbes, and the complex dynamics of their transmission (e.g., vaginal/fecal to breast, infant mouth to breast, breast to infant mouth and gut) and their site-specific colonization in the infant remains to be determined.
Importantly, breast milk also contains prebiotic human milk oligosaccharides (HMOs) – sugar polymers that promote the growth of specific microbial communities, including Bifidobacterium spp. [63–67]. Bifidobacteria in the infant gut are important for inhibiting the growth of pathogenic organisms, modulating mucosal barrier function, and promoting immunological and inflammatory responses [68]. The synergy of the probiotic and prebiotic components of human breast milk provides breastfed infants with a stable and relatively uniform gut microbiome compared with formula-fed babies [63–65]. While consumption of infant formula containing probiotics has also been shown to promote the development of a neonatal gut microbiome similar to that of breastfed infants [69], maternal breast milk remains the ideal source of nutrition for infants.
The early microbiome appears to follow a progression from organisms that facilitate lactate utilization during strict lactation to anaerobic organisms involved in the utilization of solid foods [70] after solids are introduced into the diet. At approximately the end of the first year of life, the infant microbiome achieves a more complex structure, and it becomes similar to that of adults by age 3 years [19,71,72]. In addition to facilitating nutrient utilization, the ecological succession of the infant microbiome is thought to educate the naïve immune system and program the metabolic system [71,73]. Disruption of this normal assembly may have considerable downstream consequences for the development of autoimmune [4] and metabolic [5] pathologies.
Disruptors of the colonization and development of the neonatal microbiome
The greatest insults to the natural assembly of the neonatal microbiome appear to be C-section delivery, antibiotic use, and formula feeding. The microbiological differences between C-section-and vaginally delivered babiesare striking. Unlike vaginally born babies, those born by C-section (without membrane rupture) harbor no vaginal microbes (e.g., Lactobacillus, Prevotella, Sneathia spp.) at birth [51]. Instead, they are colonized by skin bacteria (e.g., Staphylococcus, Corynebacterium, Propionibacterium spp.) [51]. Postnatal colonization of the neonatal intestine by Bacteroides and Bifidobacterium spp. is also delayed in babies delivered by C-section [74–76] and these babies have higher levels of intestinal Clostridium difficile [77–80]. While it remains unknown how long the differences in microbial composition introduced by delivery mode persist in the infant, differences in specific microbial species have been observed between C-section- and vaginally delivered babies after 1 month [81], 2 years [82], and even 7 years [83] of life.
Multiple lines of evidence indicate that antibiotics before, during, and after birth also disrupt the natural microbiome assembly [84–90]. In mice, prenatal antibiotics decrease the diversity and structure of the microbiota [90]. In humans, intrapartum antibiotic use has been associated with decreased bacterial diversity of the neo-nate's first stool [84,85] and lower abundance of lactobacilli and bifidobacteria in the neonatal gut [85–89]. Similar associations have been observed after administration of antibiotics to the neonate directly after birth [88,89]. Studies are needed to determine the effect of prenatal antibiotics on human offspring microbiota and to examine the potential effects of perinatal antibiotics on infants' health.
Introduction of formula or solid foods early in the postnatal period also perturbs the colonization and proliferation of the neonatal intestinal microbiota [19,20] and may thereby reduce the benefits of exclusive human-milk feeding [21]. Specifically, formula feeding has been associated with increased bacterial diversity [91], increased prevalence of C. difficile [79], Bacteroides fragilis, and E. coli [79,92], and decreased prevalence of bifidobacteria [93]. Formula, even given in small amounts during breastfeeding, can alter the structure [94] and relative abundances [69] of the bacterial communities normally found in a breastfed infant gut.
In addition to the major microbial insults described here (C-section delivery, antibiotic use, and formula feeding), suboptimal assembly of the infant microbiome may occur in babies born vaginally to mothers who have a disrupted vaginal or intestinal microbial ecology. An analysis of the aforementioned Finnish cohort found that women who were overweight or obese before pregnancy had higher counts of fecal Bacteroides and Staphylococcus aureus than normal-weight women in the third trimester of pregnancy, and that weight gain during pregnancy was positively associated with Bacteroides and inversely associated with growth of bifidobacteria [95]. Follow-up of infants from this cohortat 1 and 6months revealed that fecal concentrations of Bacteroides species commonly associated with obesity [96] were higher among newborns born to overweight mothers [97]. Other observational studies have reported that mothers with diabetes [98] or eczema [99] may transfer the microbiome structure characteristic of that pathology to their children.
Much of the research in this arena has been conducted in developed societies. Prospective studies are needed to determine how perinatal antibiotics, C-section delivery, and formula feeding affect the development of the microbiome and health in less-developed societies where environmental exposures to microbes differ in important ways, as indicated by studies on delivery mode and the microbiome in Bangladeshi [100] and African [101] children. Research is also needed to determine whether and how pathological conditions during pregnancy modulate maternal–offspring microbiota exchange and the health of the newborn.
Prevention and microbiome restorative strategies
Since the evolution of the mammalian birthing process must be adaptive, its disruption may prevent natural development of the neonatal microbiome and increase neonates' long-term risk of metabolic and immune diseases. In light of this, elevated rates of C-section delivery (Figure 1), pre- and perinatal antibiotic use [102,103], and formula feeding (http://www.cdc.gov/breastfeeding/pdf/2014breastfeedingreportcard.pdf) underscore the importance of promoting vaginal delivery and more conservative administration of antibiotics and a renewed emphasis on initiating and sustaining breastfeeding. Health policies and clinical practice models (e.g., midwifery care) that prioritize vaginal childbirth, as well as a reevaluation of when C-sections are considered medically necessary, have been suggested as approaches to preventing medically unnecessary C-sections [104]. Developing safe strategies that limit [e.g., rapid PCR testing for the presence of group B Streptococcus (GBS) at the time of admission to the delivery unit instead of at 35–37 weeks of gestation] or alter (e.g., give antibiotics to the mother after cord clamping) the use of antibiotics during pregnancy are also needed [102], as is broader use of antimicrobial stewardship programs (http://www.whitehouse.gov/the-press-office/2014/09/18/executive-order-combating-antibiotic-resistant-bacteria) and evidenced-based policies such as the UNICEF/WHO Baby Friendly Hospital Initiative and counselors to increase breastfeeding [105]. A list of potential strategies to prevent C-sections, perinatal antibiotic use, and formula feeding is given in Table 1.
Figure 1.
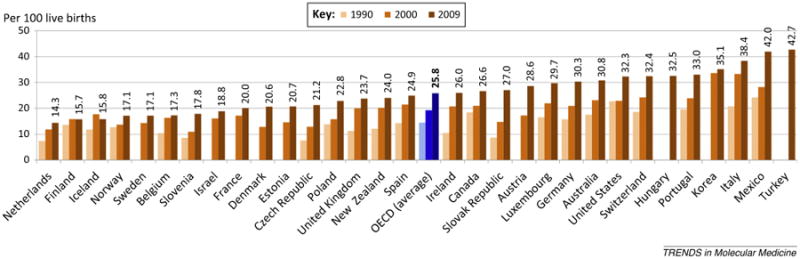
Cesarean section (C-section) rates in Organization for Economic Co-operation and Development (OECD) countries. C-sections per 100 live births for years 1990, 2000, and 2009 (as available). C-section rates have been rising dramatically, particularly in countries with already high rates, whereas Northern European states with effective health systems maintain relatively low C-section rates [OECD Health Data (2011) Health at a Glance 2011: OECD Indicators (http://dx.doi.org/10.1787/888932524906)].
Table 1. Perturbations to the assembly of the neonatal microbiome, prevention strategies, and approaches for microbiome restoration.
| Perturbation to microbiome assembly | Prevention strategies | Restoration approaches |
|---|---|---|
| C-section delivery | Support efforts to increase use of midwives Champion evidence-based labor management Optimize managing labor (reduce pain, increase maternal comfort) Educate women about the potential consequences of C-section delivery Change policies around physician incentives and malpractice insurance |
Inoculation of neonate with maternal vaginal flora immediately following C-section delivery Breastfeeding Pre- and probiotic supplementation of neonate |
| Gestational, perinatal, or postnatal antibiotics | Implement robust antimicrobial stewardship programs (http://www.whitehouse.gov/the-press-office/2014/09/18/executive-order-combating-antibiotic-resistant-bacteria) Develop safe strategies that limit use of antibiotics in women in labor (e.g., rapid PCR testing for group B Streptococcus at the time of admission to the delivery unit) During C-section delivery, give antibiotics after cord clamping to eliminate fetal exposure to antibiotics Use more prudency in antibiotic administration during pregnancy |
Breastfeeding Pre- and probiotic supplementation of mother during pregnancy and neonate after birth |
| Formula feeding | Adopt WHO/UNICEF Baby Friendly Hospital Initiative Develop other policies that incentivize breastfeeding Do not offer formula to newborns without request or medical indication Promote use of donor breast milk rather than formula when maternal milk is not an option |
Reintroduce breastfeeding Pre- and probiotic supplementation |
When prevention of insults to the developing neonatal microbiome is impossible, restoration of the neonate's exposure to a healthy maternal microbiota may provide the best chance of promoting robust microbiome development and thereby preventing long-term pathologies associated with a disrupted microbiome. Here we describe potential approaches to restoration when: (i) the maternal microbiome in pregnancy has been impacted by antibiotics or compromised maternal health; or (ii) natural microbial colonization of the neonate is disrupted by C-section delivery, antibiotic use, or formula feeding.
Maternal microbiota restoration during pregnancy
Babies born vaginally to mothers who receive antibiotics during pregnancy or labor [84–87] or to mothers with pregnancies complicated by immunological [99] or metabolic disorders such obesity [97] or diabetes [98] may acquire a marginalized inoculum of beneficial bacteria. Restorative actions could be applied either to the disrupted maternal microbiota during pregnancy, to promote the newborn's acquisition of a less-impacted microbiota, or directly to the infant after birth. The first approach would aim to alleviate the impact of disruptors on the maternal microbiota. This could be achieved through the administration of prebiotics and probiotics to the mother during pregnancy or labor, either as adjuncts to the antibiotic regimen or prophylactically to mothers with otherwise disrupted microbiota. A 2012 study by StojanoviĆ et al. examined the effect of intravaginal administration of Lactobacillus rhamnosus to 60 women once weekly for 12 weeks during the second and third trimesters of pregnancy [106]. The study found that the probiotic helped maintain a vaginal microbiome free of pathogenic microorganisms (including Candida albicans) and helped maintain a low vaginal pH. While these investigators examined the effects of probiotics on otherwise healthy pregnant women, such strategies could be employed for women with disrupted vaginal microbiota.
Probiotic supplementation of the mother during and after pregnancy has been shown to alter the infant's microbiome. Randomized trials [107–110] provide evidence that L. rhamnosus given during and after pregnancy can colonize the intestine of vaginally delivered, breastfed infants until 1–2 years of life and can increase the abundance of Bifidobacterium spp. in the infant gut. Administration of Lactobacillus reuteri to both mothers in late gestation and to infants throughout the first year of life decreased the infants' levels of IgE antibodies to food allergens at 2 years of age [111]. However, results from other human studies of probiotics administered in pregnancy are inconsistent [112]. Evidence is also mixed for the effectiveness of prebiotics during pregnancy. Initial enthusiasm regarding an experiment in mice that demonstrated the effect of gestational prebiotics on the intestinal bacterial composition of offspring [113] was tempered by null findings in humans [114]. While research has shown that probiotics during pregnancy can alter both the maternal and neonatal microbiota, there is a need for more randomized controlled clinical trials on pre- and probiotics that are consistent in terms of strains, dose, and timing; this research should be conducted free from commercial interests.
Neonatal microbiota restoration after C-section or after postnatal exposure to antibiotics or formula feeding
Birth via C-section alters the neonatal microbial profile by depriving the neonate of exposure to the maternal birth canal, including to the maternal vaginal and fecal micro-biota [19,51,52]. As noted above, a healthy maternal vaginal microbiome is rich in the lactobacilli typically found in a healthy pregnancy and free of potentially harmful bacteria (e.g., GBS) or viruses (e.g., HIV). This definition is, however, open to revision as research on what constitutes healthy-pregnancy-related changes of the vaginal microbiome continues, as does research on the composition of a healthy maternal gut microbiota.
For babies born via C-section without rupture of the amniotic membranes, restoration could be achieved by exposure of the neonate to maternal vaginal contents. Work is currently under way to determine the success of this restoration approach and preliminary results suggest that the fecal, skin, and oral microbiota of the exposed neonates more closely resemble those from vaginally born than from C-section-born babies (K. de Jesês-Laboy et al., unpublished). While the risks involved in this procedure should be similar to those during vaginal delivery, research is warranted to optimize the safety and mechanism of such exposure. Given the rapid development of the infant microbiome, early introduction of key founder populations may be key to facilitating a more natural microbial ecological succession and host immune and metabolic responses.
Direct manipulation of the infant microbiome, if possible, is an alternative, parsimonious way to restore the infant microbiota after C-section, antibiotics, or formula feeding. A vast literature shows that oral probiotics reduce the risk of death from necrotizing enterocolitis in preterm neonates [115–118]. A randomized controlled trial has reported that oral synbiotics (probiotics and prebiotics together) given to preterm babies alter the composition of their gut microbiota and decrease their risk of developing atopic diseases [119] and of fussing and crying [120].
While the literature on probiotics is promising, the beneficial effects are likely to be strain specific [121] and thus there is a need for future trials focusing on the best combinations of probiotic strains, the timing of administration, and whether these probiotics are more efficacious in conjunction with prebiotics (such as the oligosaccharides found in breast milk). These future trials would benefit from consistency in the comparison groups used (e.g., concurrent breastfeeding, using pasteurized formula).
Probably the best way to promote and maintain a healthy neonatal microbiome is through exclusive breastfeeding. Given that breast milk has the effect of seeding and selecting for particular populations of bacteria in the infant gut [122,123], it is to be expected that exclusive breastfeeding of previously formula-fed neonates would be restorative. For infants whose mothers cannot breastfeed, donor breast milk from breast-milk banks may be an alternative to formula; however, this milk is usually reserved for premature or ill infants and can be difficult to obtain for healthy babies. Milk banks in the USA pasteurize donor breast milk to destroy pathogenic microbes. This also kills beneficial bacteria, but at least does not affect HMOs or long-chain polyunsaturated fatty acids, and thus these prebiotic constituents are intact to facilitate the growth of Bifidobacterium spp. in the infant gut [124,125]. If breast milk is unavailable, formulas fortified with prebiotic and probiotic compounds may be restorative alternatives [69]. Moving forward, research aimed at understanding the complex microbial interactions in human milk will allow us to optimize pre- and probiotic formulas to mimic the bioactive nourishment provided by breastfeeding.
Concluding remarks
Vaginal delivery and breastfeeding are evolutionarily adaptive for mammals and therefore are paramount to human newborn development and health. Common perinatal interventions like C-section, antibiotic use, and formula feeding alter the infant microbiome and may be major factors shaping a new microbiome landscape in human history. While mechanistic questions remain (Box 1), epidemiological evidence suggests that these impacts on the early microbiome assembly are associated with metabolic and immune pathologies. Even if antibiotic use, C-section delivery, and formula feeding are only marginally associated with disease risk at the individual level, the widespread use of these practices in the USA and other countries may contribute to considerable disease burden at the population level [126]. Therefore, strategies to prevent perturbation of the healthy infant microbiome and restore it after alterations should be researched to help curb the epidemic trends of metabolic and immune diseases.
Box 1. Outstanding questions.
Are there live bacteria in the intrauterine environment of a healthy pregnancy?
Do these bacteria interact with the fetus, priming the immune and metabolic systems? If so, can this exchange be manipulated?
To what degree does amniotic membrane rupture allow in utero exposure to the maternal vaginal microbiome?
What are the key members of the maternal microbial communities that promote a healthy newborn microbiome?
What pathophysiological states during pregnancy lead to disrupted maternal–offspring exchange?
What bacterial strains are optimal for probiotic supplementation during pregnancy and in the neonate?
To what degree can breastfeeding restore microbes and their functions during development?
Can we restore the vaginal microbiota to C-section-delivered babies?
Does neonatal microbial restoration reduce the risk of microbiome-related diseases?
Acknowledgments
This work was partially supported by an anonymous grant (M.G.D-B.) and by the Emch Fund for Microbial Diversity (M.G.D-B.). N.T.M. received a fellowship from the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK 5T32DK091227-03) to help complete this work.
References
- 1.NIH HMP Working Group. The NIH Human Microbiome Project. Genome Res. 2009;19:2317–2323. doi: 10.1101/gr.096651.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Costello EK, et al. The application of ecological theory toward an understanding of the human microbiome. Science. 2012;336:1255–1262. doi: 10.1126/science.1224203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Baquero F, Nombela C. The microbiome as a human organ. Clin Microbiol Infect. 2012;18(Suppl. 4):2–4. doi: 10.1111/j.1469-0691.2012.03916.x. [DOI] [PubMed] [Google Scholar]
- 4.Fung I, et al. Do bugs control our fate? The influence of the microbiome on autoimmunity. Curr Allergy Asthma Rep. 2012;12:511–519. doi: 10.1007/s11882-012-0291-2. [DOI] [PubMed] [Google Scholar]
- 5.Cox LM, et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell. 2014;158:705–721. doi: 10.1016/j.cell.2014.05.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Decker E, et al. Cesarean delivery is associated with celiac disease but not inflammatory bowel disease in children. Pediatrics. 2010;125:e1433–e1440. doi: 10.1542/peds.2009-2260. [DOI] [PubMed] [Google Scholar]
- 7.Marild K, et al. Pregnancy outcome and risk of celiac disease in offspring: a nationwide case-control study. Gastroenterology. 2012;142:39–45. doi: 10.1053/j.gastro.2011.09.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kero J, et al. Mode of delivery and asthma – is there a connection? Pediatr Res. 2002;52:6–11. doi: 10.1203/00006450-200207000-00004. [DOI] [PubMed] [Google Scholar]
- 9.Roduit C, et al. Asthma at 8 years of age in children born by Caesarean section. Thorax. 2009;64:107–113. doi: 10.1136/thx.2008.100875. [DOI] [PubMed] [Google Scholar]
- 10.Couzin-Frankel J. Bacteria and asthma: untangling the links. Science. 2010;330:1168–1169. doi: 10.1126/science.330.6008.1168. [DOI] [PubMed] [Google Scholar]
- 11.Ege MJ, et al. Exposure to environmental microorganisms and childhood asthma. N Engl J Med. 2011;364:701–709. doi: 10.1056/NEJMoa1007302. [DOI] [PubMed] [Google Scholar]
- 12.Algert CS, et al. Perinatal risk factors for early onset of type 1 diabetes in a 2000-2005 birth cohort. Diabet Med. 2009;26:1193–1197. doi: 10.1111/j.1464-5491.2009.02878.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aumeunier A, et al. Systemic Toll-like receptor stimulation suppresses experimental allergic asthma and autoimmune diabetes in NOD mice. PLoS ONE. 2010;5:e11484. doi: 10.1371/journal.pone.0011484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huh SY, et al. Delivery by Caesarean section and risk of obesity in preschool age children: a prospective cohort study. Arch Dis Child. 2012;97:610–616. doi: 10.1136/archdischild-2011-301141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ajslev TA, et al. Childhood overweight after establishment of the gut microbiota: the role of delivery mode, pre-pregnancy weight and early administration of antibiotics. Int J Obes (Lond) 2011;35:522–529. doi: 10.1038/ijo.2011.27. [DOI] [PubMed] [Google Scholar]
- 16.Blustein J, et al. Association of caesarean delivery with child adiposity from age 6 weeks to 15 years. Int J Obes (Lond) 2013;37:900–906. doi: 10.1038/ijo.2013.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mueller NT, et al. Prenatal exposure to antibiotics, Cesarean section and risk of childhood obesity. Int J Obes (Lond) 2014 doi: 10.1038/ijo.2014.180. http://dx.doi.org/10.1038/ijo.2014.180 Published online November 11, 2014. [DOI] [PMC free article] [PubMed]
- 18.Metsälä J, et al. Prenatal and postnatal exposure to antibiotics and risk of asthma in childhood. Clin Exp Allergy. 2015;45:137–145. doi: 10.1111/cea.12356. [DOI] [PubMed] [Google Scholar]
- 19.Palmer C, et al. Development of the human infant intestinal microbiota. PLoS Biol. 2007;5:e177. doi: 10.1371/journal.pbio.0050177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Knol J, et al. Colon microflora in infants fed formula with galacto- and fructo-oligosaccharides: more like breast-fed infants. J Pedi atr Gastroenterol Nutr. 2005;40:36–42. doi: 10.1097/00005176-200501000-00007. [DOI] [PubMed] [Google Scholar]
- 21.Innis SM. Human milk: maternal dietary lipids and infant development. Proc Nutr Soc. 2007;66:397–404. doi: 10.1017/S0029665107005666. [DOI] [PubMed] [Google Scholar]
- 22.Gale C, et al. Effect of breastfeeding compared with formula feeding on infant body composition: a systematic review and meta-analysis. Am J Clin Nutr. 2012;95:656–669. doi: 10.3945/ajcn.111.027284. [DOI] [PubMed] [Google Scholar]
- 23.Aagaard K, et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS ONE. 2012;7:e36466. doi: 10.1371/journal.pone.0036466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Romero R, et al. The composition and stability of the vaginal microbiota of normal pregnant women is different from that of non-pregnant women. Microbiome. 2014;2:4. doi: 10.1186/2049-2618-2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Koren O, et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell. 2012;150:470–480. doi: 10.1016/j.cell.2012.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kuczynski J, et al. Experimental and analytical tools for studying the human microbiome. Nat Rev Genet. 2012;13:47–58. doi: 10.1038/nrg3129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.El Aila NA, et al. Identification and genotyping of bacteria from paired vaginal and rectal samples from pregnant women indicates similarity between vaginal and rectal microflora. BMC Infect Dis. 2009;9:167. doi: 10.1186/1471-2334-9-167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Goldenberg RL, et al. Intrauterine infection and preterm delivery. N Engl J Med. 2000;342:1500–1507. doi: 10.1056/NEJM200005183422007. [DOI] [PubMed] [Google Scholar]
- 29.van de Wijgert JH, et al. The vaginal microbiota: what have we learned after a decade of molecular characterization? PLoS ONE. 2014;9:e105998. doi: 10.1371/journal.pone.0105998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Corr SC, et al. Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc Natl Acad Sci U S A. 2007;104:7617–7621. doi: 10.1073/pnas.0700440104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Spurbeck RR, Arvidson CG. Lactobacillus jensenii surface-associated proteins inhibit Neisseria gonorrhoeae adherence to epithelial cells. Infect Immun. 2010;78:3103–3111. doi: 10.1128/IAI.01200-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rosenstein IJ, et al. Relationship between hydrogen peroxide-producing strains of lactobacilli and vaginosis-associated bacterial species in pregnant women. Eur J Clin Microbiol Infect Dis. 1997;16:517–522. doi: 10.1007/BF01708235. [DOI] [PubMed] [Google Scholar]
- 33.Pantoja-Feliciano IG, et al. Biphasic assembly of the murine intestinal microbiota during early development. ISME J. 2013;7:1112–1115. doi: 10.1038/ismej.2013.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Thum C, et al. Can nutritional modulation of maternal intestinal microbiota influence the development of the infant gastrointestinal tract? J Nutr. 2012;142:1921–1928. doi: 10.3945/jn.112.166231. [DOI] [PubMed] [Google Scholar]
- 35.Makino H, et al. Transmission of intestinal Bifidobacterium longum subsp. longum strains from mother to infant, determined by multilocus sequencing typing and amplified fragment length polymporphism. Appl Environ Microbiol. 2011;77:6788–6793. doi: 10.1128/AEM.05346-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Funkhouser LJ, Bordenstein SR. Mom knows best: the universality of maternal microbial transmission. PLoS Biol. 2013;11:e1001631. doi: 10.1371/journal.pbio.1001631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Oh KJ, et al. Detection of ureaplasmas by the polymerase chain reaction in the amniotic fluid of patients with cervical insufficiency. J Perinat Med. 2010;38:261–268. doi: 10.1515/JPM.2010.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bearfield C, et al. Possible association between amniotic fluid micro-organism infection and microflora in the mouth. BJOG. 2002;109:527–533. doi: 10.1111/j.1471-0528.2002.01349.x. [DOI] [PubMed] [Google Scholar]
- 39.Han YW, et al. Transmission of an uncultivated Bergeyella strain from the oral cavity to amniotic fluid in a case of preterm birth. J Clin Microbiol. 2006;44:1475–1483. doi: 10.1128/JCM.44.4.1475-1483.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang X, et al. Comparative microbial analysis of paired amniotic fluid and cord blood from pregnancies complicated by preterm birth and early-onset neonatal sepsis. PLoS ONE. 2013;8:e56131. doi: 10.1371/journal.pone.0056131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jimenez E, et al. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by Cesarean section. Curr Microbiol. 2005;51:270–274. doi: 10.1007/s00284-005-0020-3. [DOI] [PubMed] [Google Scholar]
- 42.Steel JH, et al. Bacteria and inflammatory cells in fetal membranes do not always cause preterm labor. Pediatr Res. 2005;57:404–411. doi: 10.1203/01.PDR.0000153869.96337.90. [DOI] [PubMed] [Google Scholar]
- 43.Aagaard K, et al. The placenta harbors a unique microbiome. Sci Transl Med. 2014;6:237ra265. doi: 10.1126/scitranslmed.3008599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Douvier S, et al. Chorioamnionitis with intact membranes caused by Capnocytophaga sputigena. Eur J Obstet Gynecol Reprod Biol. 1999;83:109–112. doi: 10.1016/s0301-2115(98)00240-1. [DOI] [PubMed] [Google Scholar]
- 45.Han YW, et al. Fusobacterium nucleatum induces premature and term stillbirths in pregnant mice: implication of oral bacteria in preterm birth. Infect Immun. 2004;72:2272–2279. doi: 10.1128/IAI.72.4.2272-2279.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fardini Y, et al. Transmission of diverse oral bacteria to murine placenta: evidence for the oral microbiome as a potential source of intrauterine infection. Infect Immun. 2010;78:1789–1796. doi: 10.1128/IAI.01395-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Stout MJ, et al. Identification of intracellular bacteria in the basal plate of the human placenta in term and preterm gestations. Am J Obstet Gynecol. 2013;208:226 e1–226.e7. doi: 10.1016/j.ajog.2013.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Satokari R, et al. Bifidobacterium and Lactobacillus DNA in the human placenta. Lett Appl Microbiol. 2009;48:8–12. doi: 10.1111/j.1472-765X.2008.02475.x. [DOI] [PubMed] [Google Scholar]
- 49.Rautava S, et al. Probiotics modulate host–microbe interaction in the placenta and fetal gut: a randomized, double-blind, placebo-controlled trial. Neonatology. 2012;102:178–184. doi: 10.1159/000339182. [DOI] [PubMed] [Google Scholar]
- 50.Kliman HJ. Comment on “The placenta harbors a unique microbiome”. Sci Transl Med. 2014;6:254le4. doi: 10.1126/scitranslmed.3009864. [DOI] [PubMed] [Google Scholar]
- 51.Dominguez-Bello MG, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A. 2010;107:11971–11975. doi: 10.1073/pnas.1002601107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Biasucci G, et al. Mode of delivery affects the bacterial community in the newborn gut. Early Hum Dev. 2010;86(Suppl. 1):13–15. doi: 10.1016/j.earlhumdev.2010.01.004. [DOI] [PubMed] [Google Scholar]
- 53.Delgado S, et al. Bacterial analysis of breast milk: a tool to differentiate Raynaud's phenomenon from infectious mastitis during lactation. Curr Microbiol. 2009;59:59–64. doi: 10.1007/s00284-009-9393-z. [DOI] [PubMed] [Google Scholar]
- 54.Collado MC, et al. Assessment of the bacterial diversity of breast milk of healthy women by quantitative real-time PCR. Lett Appl Microbiol. 2009;48:523–528. doi: 10.1111/j.1472-765X.2009.02567.x. [DOI] [PubMed] [Google Scholar]
- 55.Hunt KM, et al. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE. 2011;6:e21313. doi: 10.1371/journal.pone.0021313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cabrera-Rubio R, et al. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am J Clin Nutr. 2012;96:544–551. doi: 10.3945/ajcn.112.037382. [DOI] [PubMed] [Google Scholar]
- 57.Jeurink PV, et al. Human milk: a source of more life than we imagine. Benef Microbes. 2013;4:17–30. doi: 10.3920/BM2012.0040. [DOI] [PubMed] [Google Scholar]
- 58.Jost T, et al. Stability of the maternal gut microbiota during late pregnancy and early lactation. Curr Microbiol. 2014;68:419–427. doi: 10.1007/s00284-013-0491-6. [DOI] [PubMed] [Google Scholar]
- 59.Jost T, et al. Vertical mother-neonate transfer of maternal gut bacteria via breastfeeding. Environ Microbiol. 2014;16:2891–2904. doi: 10.1111/1462-2920.12238. [DOI] [PubMed] [Google Scholar]
- 60.Martin R, et al. Diversity of the Lactobacillus group in breast milk and vagina of healthy women and potential role in the colonization of the infant gut. J Appl Microbiol. 2007;103:2638–2644. doi: 10.1111/j.1365-2672.2007.03497.x. [DOI] [PubMed] [Google Scholar]
- 61.Solís G, et al. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe. 2010;16:307–310. doi: 10.1016/j.anaerobe.2010.02.004. [DOI] [PubMed] [Google Scholar]
- 62.Khodayar-Pardo P, et al. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J Perinatol. 2014;34:599–605. doi: 10.1038/jp.2014.47. [DOI] [PubMed] [Google Scholar]
- 63.Coppa GV, et al. The first prebiotics in humans: human milk oligosaccharides. J Clin Gastroenterol. 2004;38:S80–S83. doi: 10.1097/01.mcg.0000128926.14285.25. [DOI] [PubMed] [Google Scholar]
- 64.Barile D, Rastall RA. Human milk and related oligosaccharides as prebiotics. Curr Opin Biotechnol. 2013;24:214–219. doi: 10.1016/j.copbio.2013.01.008. [DOI] [PubMed] [Google Scholar]
- 65.Bode L. Human milk oligosaccharides: prebiotics and beyond. Nutr Rev. 2009;67(Suppl. 2):S183–S191. doi: 10.1111/j.1753-4887.2009.00239.x. [DOI] [PubMed] [Google Scholar]
- 66.Oozeer R, et al. Intestinal microbiology in early life: specific prebiotics can have similar functionalities as human-milk oligosaccharides. Am J Clin Nutr. 2013;98:561S–571S. doi: 10.3945/ajcn.112.038893. [DOI] [PubMed] [Google Scholar]
- 67.Gronlund MM, et al. Influence of mother's intestinal microbiota on gut colonization in the infant. Gut Microbes. 2011;2:227–233. doi: 10.4161/gmic.2.4.16799. [DOI] [PubMed] [Google Scholar]
- 68.Sudo N, et al. The requirement of intestinal bacterial flora for the development of an IgE production system fully susceptible to oral tolerance induction. J Immunol. 1997;159:1739–1745. [PubMed] [Google Scholar]
- 69.Guaraldi F, Salvatori G. Effect of breast and formula feeding on gut microbiota shaping in newborns. Front Cell Infect Microbiol. 2012;2:94. doi: 10.3389/fcimb.2012.00094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Koenig JE, et al. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci U S A. 2011;108(Suppl. 1):4578–4585. doi: 10.1073/pnas.1000081107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Murgas Torrazza R, Neu J. The developing intestinal microbiome and its relationship to health and disease in the neonate. J Perinatol. 2011;31(Suppl. 1):S29–S34. doi: 10.1038/jp.2010.172. [DOI] [PubMed] [Google Scholar]
- 72.Yatsunenko T, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486:222–227. doi: 10.1038/nature11053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Azad MB, Kozyrskyj AL. Perinatal programming of asthma: the role of gut microbiota. Clin Dev Immunol. 2012;2012:1–9. doi: 10.1155/2012/932072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kabeerdoss J, et al. Development of the gut microbiota in southern Indian infants from birth to 6 months: a molecular analysis. J Nutr Sci. 2013;2:e18. doi: 10.1017/jns.2013.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Tsuji H, et al. Molecular monitoring of the development of intestinal microbiota in Japanese infants. Benef Microbes. 2012;3:113–125. doi: 10.3920/BM2011.0038. [DOI] [PubMed] [Google Scholar]
- 76.Biasucci G, et al. Cesarean delivery may affect the early biodiversity of intestinal bacteria. J Nutr. 2008;138:1796S–1800S. doi: 10.1093/jn/138.9.1796S. [DOI] [PubMed] [Google Scholar]
- 77.Penders J, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. 2006;188:511–521. doi: 10.1542/peds.2005-2824. [DOI] [PubMed] [Google Scholar]
- 78.Pandey PK, et al. Comparative analysis of fecal microflora of healthy full-term Indian infants born with different methods of delivery (vaginal vs Cesarean); Acinetobacter sp. prevalence in vaginally born infants. J Biosci. 2012;37:989–998. doi: 10.1007/s12038-012-9268-5. [DOI] [PubMed] [Google Scholar]
- 79.Penders J, et al. Quantification of Bifidobacterium spp., Escherichia coli and Clostridium difficile in faecal samples of breast-fed and formula-fed infants by real-time PCR. FEMS Microbiol Lett. 2005;243:141–147. doi: 10.1016/j.femsle.2004.11.052. [DOI] [PubMed] [Google Scholar]
- 80.van Nimwegen FA, et al. Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy. J Allergy Clin Immunol. 2011;128:948–955. doi: 10.1016/j.jaci.2011.07.027. [DOI] [PubMed] [Google Scholar]
- 81.Huurre A, et al. Mode of delivery – effects on gut microbiota and humoral immunity. Neonatology. 2008;93:236–240. doi: 10.1159/000111102. [DOI] [PubMed] [Google Scholar]
- 82.Jakobsson HE, et al. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by Caesarean section. Gut. 2014;63:559–566. doi: 10.1136/gutjnl-2012-303249. [DOI] [PubMed] [Google Scholar]
- 83.Salminen S, et al. Influence of mode of delivery on gut microbiota composition in seven year old children. Gut. 2004;53:1388–1389. doi: 10.1136/gut.2004.041640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mshvildadze M, et al. Intestinal microbial ecology in premature infants assessed with non-culture-based techniques. J Pediatr. 2010;156:20–25. doi: 10.1016/j.jpeds.2009.06.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Jaureguy F, et al. Effects of intrapartum penicillin prophylaxis on intestinal bacterial colonization in infants. J Clin Microbiol. 2004;42:5184–5188. doi: 10.1128/JCM.42.11.5184-5188.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Keski-Nisulu L, et al. Maternal intrapartum antibiotics and decreased vertical transmission of Lactobacillus to neonates during birth. Acta Paediatr. 2013;102:480–485. doi: 10.1111/apa.12186. [DOI] [PubMed] [Google Scholar]
- 87.Aloisio I, et al. Influence of intrapartum antibiotic prophylaxis against group B Streptococcus on the early newborn gut composition and evaluation of the anti-Streptococcus activity of Bifidobacterium strains. Appl Microbiol Biotechnol. 2014;98:6051–6060. doi: 10.1007/s00253-014-5712-9. [DOI] [PubMed] [Google Scholar]
- 88.Fouhy F, et al. High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob Agents Chemother. 2012;56:5811–5820. doi: 10.1128/AAC.00789-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tanaka S, et al. Influence of antibiotic exposure in the early postnatal period on the development of intestinal microbiota. FEMS Immunol Med Microbiol. 2009;56:80–87. doi: 10.1111/j.1574-695X.2009.00553.x. [DOI] [PubMed] [Google Scholar]
- 90.Tormo-Badia N, et al. Antibiotic treatment of pregnant non-obese diabetic mice leads to altered gut microbiota and intestinal immunological changes in the offspring. Scand J Immunol. 2014;80:250–260. doi: 10.1111/sji.12205. [DOI] [PubMed] [Google Scholar]
- 91.Fallani M, et al. Intestinal microbiota of 6-week-old infants across Europe: geographic influence beyond delivery mode, breastfeeding, and antibiotics. J Pediatr Gastroenterol Nutr. 2010;51:77–84. doi: 10.1097/MPG.0b013e3181d1b11e. [DOI] [PubMed] [Google Scholar]
- 92.Cooke G, et al. Comparing the gut flora of Irish breastfed and formula-fed neonates aged between birth and six weeks. Microb Ecol Health Dis. 2005;17:163–168. [Google Scholar]
- 93.Bezirtzoglou E, et al. Microbiota profile in feces of breast- and formula-fed newborns by using florescence in situ hybridization (FISH) Anaerobe. 2011;17:478–482. doi: 10.1016/j.anaerobe.2011.03.009. [DOI] [PubMed] [Google Scholar]
- 94.Mackie RI, et al. Developmental microbial ecology of the neonatal gastrointestinal tract. Am J Clin Nutr. 1999;69:1035S–1045S. doi: 10.1093/ajcn/69.5.1035s. [DOI] [PubMed] [Google Scholar]
- 95.Collado MC, et al. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am J Clin Nutr. 2008;88:894–899. doi: 10.1093/ajcn/88.4.894. [DOI] [PubMed] [Google Scholar]
- 96.Cox LM, Blaser MJ. Pathways in microbe-induced obesity. Cell Metab. 2013;17:883–894. doi: 10.1016/j.cmet.2013.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Collado MC, et al. Effect of mother's weight on infant's microbiota acquisition, composition, and activity during early infancy: a prospective follow-up study initiated in early pregnancy. Am J Clin Nutr. 2010;92:1023–1030. doi: 10.3945/ajcn.2010.29877. [DOI] [PubMed] [Google Scholar]
- 98.Hu J, et al. Diversified microbiota of meconium is affected by maternal diabetes status. PLoS ONE. 2013;8:e78257. doi: 10.1371/journal.pone.0078257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Gosalbes MJ, et al. Meconium microbiota types dominated by lactic acid or enteric bacteria are differentially associated with maternal eczema and respiratory problems in infants. Clin Exp Allergy. 2013;43:198–211. doi: 10.1111/cea.12063. [DOI] [PubMed] [Google Scholar]
- 100.Huda MN, et al. Stool microbiota and vaccine responses of infants. Pediatrics. 2014;134:e362–e372. doi: 10.1542/peds.2013-3937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Grzeskowiak L, et al. Distinct gut microbiota in southeastern African and Northern European infants. J Pediatr Gastroenterol Nutr. 2012;54:812–816. doi: 10.1097/MPG.0b013e318249039c. [DOI] [PubMed] [Google Scholar]
- 102.Ledger WJ, Blaser MJ. Are we using too many antibiotics during pregnancy? BJOG. 2013;120:1450–1452. doi: 10.1111/1471-0528.12371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Broe A, et al. Increasing use of antibiotics in pregnancy during the period 2000-2010: prevalence, timing, category, and demographics. BJOG. 2014;121:988–996. doi: 10.1111/1471-0528.12806. [DOI] [PubMed] [Google Scholar]
- 104.Renfrew MJ, et al. Midwifery and quality care: findings from a new evidence-informed framework for maternal and newborn care. Lancet. 2014;384:1129–1145. doi: 10.1016/S0140-6736(14)60789-3. [DOI] [PubMed] [Google Scholar]
- 105.Spiby H, et al. A systematic review of education and evidence-based practice interventions with health professionals and breast feeding counsellors on duration of breast feeding. Midwifery. 2009;25:50–61. doi: 10.1016/j.midw.2007.01.006. [DOI] [PubMed] [Google Scholar]
- 106.Stojanovic N, et al. Normal vaginal flora, disorders and application of probiotics in pregnancy. Arch Gynecol Obstet. 2012;286:325–332. doi: 10.1007/s00404-012-2293-7. [DOI] [PubMed] [Google Scholar]
- 107.Tannock GW, et al. Plasmid profiling of members of the family Enterobacteriaceae, lactobacilli, and bifidobacteria to study the transmission of bacteria from mother to infant. J Clin Microbiol. 1990;28:1225–1228. doi: 10.1128/jcm.28.6.1225-1228.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Gueimonde M, et al. Effect of maternal consumption of Lactobacillus GG on transfer and establishment of fecal bifidobacterial microbiota in neonates. J Pediatr Gastroenterol Nutr. 2006;42:166–170. doi: 10.1097/01.mpg.0000189346.25172.fd. [DOI] [PubMed] [Google Scholar]
- 109.Schultz M, et al. Administration of oral probiotic bacteria to pregnant women causes temporary infantile colonization. J Pediatr Gastroenterol Nutr. 2004;38:293–297. doi: 10.1097/00005176-200403000-00012. [DOI] [PubMed] [Google Scholar]
- 110.Lahtinen SJ, et al. Prenatal probiotic administration can influence Bifidobacterium microbiota development in infants at high risk of allergy. J Allergy Clin Immunol. 2009;123:499–501. doi: 10.1016/j.jaci.2008.11.034. [DOI] [PubMed] [Google Scholar]
- 111.Abrahamsson TR, et al. Probiotics in prevention of IgE-associated eczema: a double-blind, randomized, placebo-controlled trial. J Allergy Clin Immunol. 2007;119:1174–1180. doi: 10.1016/j.jaci.2007.01.007. [DOI] [PubMed] [Google Scholar]
- 112.Ismail IH, et al. Prenatal administration of Lactobacillus rhamnosus has no effect on the diversity of the early infant gut microbiota. Pediatr Allergy Immunol. 2012;23:255–258. doi: 10.1111/j.1399-3038.2011.01239.x. [DOI] [PubMed] [Google Scholar]
- 113.Fujiwara R, et al. Maternal consumption of fructooligosaccharide diminishes the severity of skin inflammation in offspring of NC/Nga mice. Br J Nutr. 2010;103:530–538. doi: 10.1017/S000711450999198X. [DOI] [PubMed] [Google Scholar]
- 114.Shadid R, et al. Effects of galactooligosaccharide and longchain fructooligosaccharide supplementation during pregnancy on maternal and neonatal microbiota and immunity – a randomized, double-blind, placebo-controlled study. Am J Clin Nutr. 2007;86:1426–1437. doi: 10.1093/ajcn/86.5.1426. [DOI] [PubMed] [Google Scholar]
- 115.Deshpande G, et al. Updated meta-analysis of probiotics for preventing necrotizing enterocolitis in preterm neonates. Pediatrics. 2010;125:921–930. doi: 10.1542/peds.2009-1301. [DOI] [PubMed] [Google Scholar]
- 116.AlFaleh K, et al. Probiotics for prevention of necrotizing enterocolitis in preterm infants. Cochrane Database Syst Rev. 2011;3:CD005496. doi: 10.1002/14651858.CD005496.pub3. [DOI] [PubMed] [Google Scholar]
- 117.Guthmann F, et al. Probiotics for prevention of necrotising enterocolitis: an updated meta-analysis. Klin Padiatr. 2010;222:284–290. doi: 10.1055/s-0030-1254113. [DOI] [PubMed] [Google Scholar]
- 118.Deshpande G, et al. Probiotics for prevention of necrotising enterocolitis in preterm neonates with very low birthweight: a systematic review of randomised controlled trials. Lancet. 2007;369:1614–1620. doi: 10.1016/S0140-6736(07)60748-X. [DOI] [PubMed] [Google Scholar]
- 119.Luoto R, et al. Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: a randomized, placebo-controlled trial. J Allergy Clin Immunol. 2014;133:405–413. doi: 10.1016/j.jaci.2013.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Partty A, et al. Effects of early prebiotic and probiotic supplementation on development of gut microbiota and fussing and crying in preterm infants: a randomized, double-blind, placebo-controlled trial. J Pediatr. 2013;163:1272–1277. doi: 10.1016/j.jpeds.2013.05.035. [DOI] [PubMed] [Google Scholar]
- 121.Wallace TD, et al. Interactions of lactic acid bacteria with human intestinal epithelial cells: effects on cytokine production. J Food Prot. 2003;66:466–472. doi: 10.4315/0362-028x-66.3.466. [DOI] [PubMed] [Google Scholar]
- 122.Nakamura N, et al. Molecular ecological analysis of fecal bacterial populations from term infants fed formula supplemented with selected blends of prebiotics. Appl Environ Microbiol. 2009;75:1121–1128. doi: 10.1128/AEM.02359-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Shah N. Functional cultures and health benefits. Int Dairy J. 2007;17:1262–1277. [Google Scholar]
- 124.Bertino E, et al. Effects of Holder pasteurization on human milk oligosaccharides. Int J Immunopathol Pharmacol. 2008;21:381–385. doi: 10.1177/039463200802100216. [DOI] [PubMed] [Google Scholar]
- 125.Tully DB, et al. Donor milk: what's in it and what's not. J Hum Lact. 2001;17:152–155. doi: 10.1177/089033440101700212. [DOI] [PubMed] [Google Scholar]
- 126.Hyde MJ, Modi N. The long-term effects of birth by Caesarean section: the case for a randomised controlled trial. Early Hum Dev. 2012;88:943–949. doi: 10.1016/j.earlhumdev.2012.09.006. [DOI] [PubMed] [Google Scholar]