

Summary
C andida albicans biofilms are composed of highly adherent and densely arranged cells with properties distinct from those of free‐floating (planktonic) cells. These biofilms are a significant medical problem because they commonly form on implanted medical devices, are drug resistant and are difficult to remove. C . albicans biofilms are not static structures; rather they are dynamic and develop over time. Here we characterize gene expression in biofilms during their development, and by comparing them to multiple planktonic reference states, we identify patterns of gene expression relevant to biofilm formation. In particular, we document time‐dependent changes in genes involved in adhesion and metabolism, both of which are at the core of biofilm development. Additionally, we identify three new regulators of biofilm formation, Flo8, Gal4, and Rfx2, which play distinct roles during biofilm development over time. Flo8 is required for biofilm formation at all time points, and Gal4 and Rfx2 are needed for proper biofilm formation at intermediate time points.
Introduction
A biofilm is a community of cells arranged in a complex structure where individual cells have properties different from cells grown in suspension cultures (Hawser and Douglas, 1994; Baillie and Douglas, 1999; Chandra et al., 2001; Nobile and Mitchell, 2007; Bonhomme and d'Enfert, 2013). For example, cells in biofilms are more adherent and more drug resistant than free‐floating cells (Hawser and Douglas, 1994; Jabra‐Rizk et al., 2004; Nobile et al., 2006; 2008; López et al., 2010; Tobudic et al., 2012). Analogous to developing animal and plant tissues, biofilms undergo changes over time that require cell–cell communication, regulation of structure and cell differentiation (Baillie and Douglas, 1999; Chandra et al., 2001). In Candida albicans, a major fungal species of the human microbiome, the initial stage is adherence, in which yeast‐form cells attach to a surface and form a monolayer. Then, a subpopulation of yeast‐form cells undergoes a morphological transition into elongated hyphal cells. As the biofilm develops, a protein‐ and carbohydrate‐rich extracellular matrix forms, which encases the biofilm and plays a key role in drug resistance (Hawser et al., 1998; Nett et al., 2007, 2010a, 2010b; Zarnowski et al., 2014). The final stage is dispersal, where yeast‐form cells are released from mature biofilms and can seed new biofilms in distant locations (Uppuluri et al., 2010).
Candida albicans biofilms form on medical devices, including heart valves, catheters and prostheses (Kojic et al., 2004; Cauda, 2009). As biofilms are highly adherent and drug resistant, these infections typically require costly device removal and replacement. In some cases, biofilm infections act as reservoirs for systemic infections with high morbidity and mortality rates (Kojic et al., 2004; Raad et al., 2007; Cauda, 2009; Seddiki et al., 2013).
We examined regulation of biofilm formation over time, assessing biofilm samples immediately after adherence, at 8, 24 and 48 h. We measured gene expression levels at each time point and analyzed patterns that change throughout biofilm development. Because temperature, cell morphology and cell–cycle growth phase all affect gene expression patterns, we used four different ‘reference’ conditions to highlight changes that were specific to biofilms. Thus, we conducted four sets of gene expression microarray experiments, each with a different ‘reference’ strain grown under a particular set of conditions. These are (i) planktonic culture grown to log phase at 30°C (yeast‐form cells); (ii) planktonic culture grown to log phase at 37°C (predominantly filamentous cells); (iii) planktonic culture grown to stationary phase at 30°C (yeast‐form cells); and (iv) the unadhered, planktonic cells collected after 90 min at 37°C in our biofilm assay (predominantly filamentous cells). Previous studies have monitored gene expression in C. albicans over time in different ways (García‐Sánchez et al., 2004; Murillo et al., 2005; Yeater et al., 2007; Nett et al., 2012). García‐Sánchez et al. (2004) compared gene expression in biofilms grown in different conditions to that in planktonic cultures grown to stationary phase; Murillo et al. (2005) measured gene expression in young biofilms, up to 6 h, compared with planktonic cultures of the same age; Yeater et al. (2007) measured gene expression in 6, 12 and 48 h biofilms, compared with planktonic cultures of the same age; and Nett et al. (2012) measured gene expression in biofilms grown in a rat catheter central venous model grown for either 12 or 24 h, compared with log phase planktonic cultures. Our work measures gene expression during biofilm development over time in reference to log phase, stationary phase, filamentous and yeast form planktonic cultures, encompassing a comprehensive cohort of reference conditions, including several growth conditions that have not been previously investigated. We analyzed the gene expression data to identify biofilm‐specific sets of genes that are temporally regulated, with particular emphasis on expression patterns of transcription regulators, with the goal of identifying new transcription regulators important for biofilm development over time. In particular, we sought transcription regulators that are required for normal temporal biofilm development at one or multiple time points throughout the course of biofilm development, assessing biofilm samples immediately after adherence, at 8, 24 and 48 h.
Previous work showed that a network of six ‘master’ transcription regulators (Bcr1, Efg1, Ndt80, Rob1, Tec1 and Brg1) is required for normal biofilm formation both in vitro and in vivo in two different animal models designed to mimic human biofilm‐related candidiasis (Ramage et al., 2002a; Nobile and Mitchell, 2005; Fox and Nobile, 2012; Nobile et al., 2012). These regulators control about one thousand genes, many of which are differentially regulated in mature (48 h) biofilms compared with planktonic cultures. The master regulators were identified by screening a transcription factor deletion library of 165 mutants (Homann et al., 2009; Nobile et al., 2012) for biofilm formation at 48 h, and we used our gene expression data to prioritize construction of 27 additional regulator deletion mutant strains, creating a library of 192 mutants in total. In order to identify transcription regulators required for normal biofilm development, we screened this expanded library for biofilm formation at the same four time points used for transcriptional profiling: immediately after adherence, at 8, 24 and 48 h. We then validated our results using confocal scanning laser microscopy (CSLM) in vitro and SEM in vivo. In addition to the six previously identified master regulators (Nobile et al., 2012), the screen revealed three new transcription regulators that are required for normal biofilm development over time both in vitro and in vivo, Flo8, Rfx2 and Gal4. Chromatin immunoprecipitation experiments carried out with the three newly identified regulators show that each binds upstream of at least one previously identified master regulator gene. Moreover, the upstream regions of the three new genes are each bound by at least one master regulator. Thus, the newly identified regulators are well integrated into the previously described C. albicans transcriptional network.
Results
Multiple reference conditions reveal biofilm‐specific expression patterns
We measured gene expression during biofilm formation using microarrays that covered the complete genome of 6,111 genes (Dataset 1). We created four sets of arrays where biofilm gene expression at four time points was compared with one of four reference conditions: (i) planktonic culture grown to log phase at 30°C; (ii) planktonic culture grown to log phase at 37°C; (iii) planktonic culture grown to saturation at 30°C; and (iv) the unadhered, planktonic cells collected after 90 min at 37°C in our biofilm assay (Fig. S1, Dataset 1). Collectively, the reference conditions enabled us to filter out changes due to a wide variety of variables in order to highlight those changes related to biofilm formation per se. The use of multiple reference conditions allowed many conclusions to be drawn from the data. We first examined genes that were upregulated at specific time points irrespective of which planktonic reference condition was used. This allowed us to identify 81 genes that are biofilm‐specific and temporally regulated (Fig. 1). Eleven of these (ALS1, TRY6, RHR2, RHD1, ORF19.2669, HGT10, CWH8, ORF19.3483, SAP5, NAG3 and NAG 4) are upregulated early and include several genes previously identified as important for biofilm formation. TRY6 encodes a transcriptional regulator of adhesion (Finkel et al., 2012) and ALS1 encodes an adhesion protein involved in biofilm formation (Nobile et al., 2006). RHR2 encodes a glycerol biosynthetic enzyme and is required for normal biofilm formation (Desai et al., 2013). HGT10 and ORF19.3483 are involved in glycerol acquisition (Kunze et al., 2005; Kayingo et al., 2009), but play no known role in biofilms. NAG3 and NAG4 encode drug efflux pumps, which have not been studied in biofilms (Sengupta and Datta, 2003). Of the 70 genes upregulated at later time points, 24 are uncharacterized. Several other genes identified in our study are involved in amino acid metabolism and acquisition (ARG1, ARG3, ARG8, CPA1, CPA2, GLN1, AAP1, HNM3 and CAN1), which is consistent with previous measures of biofilm gene expression (García‐Sánchez et al., 2004; Yeater et al., 2007). Genes involved in fatty acid beta‐oxidation and lipid catabolism were also upregulated (ADR1 and CTA4).
Figure 1.
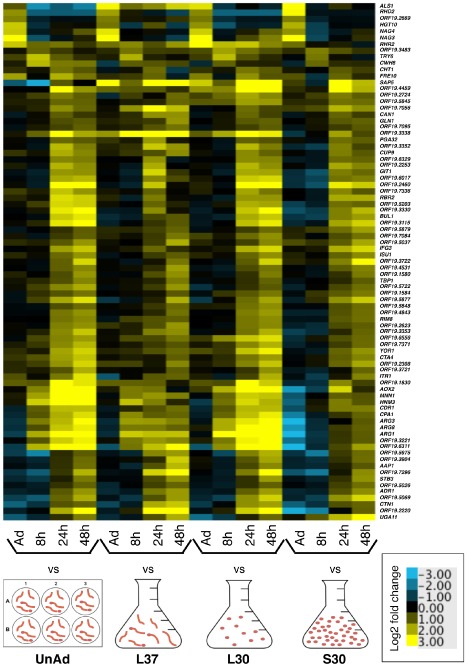
Eighty‐one genes are upregulated temporally in biofilms irrespective of which planktonic reference is used. Heat map of gene expression in C . albicans in biofilms at the indicated time points, compared with each of four reference conditions: unadhered cells (UnAd), log phase cells grown at 37°C (L37), log phase cells grown at 30°C (L30) and stationary phase cells grown at 30°C (S30). Shown are the median values of at least two biological replicates. On the y‐axis are 81 genes that are expressed early or late in biofilms compared with each reference condition. Upregulated genes are yellow; downregulated genes are blue. See also Fig. S1.
As the majority of the characterized genes temporally regulated in biofilms were involved in adhesion and metabolism, we chose to focus on these processes for further analysis. We also used the gene expression data to identify transcription regulators that are temporally regulated during biofilm formation to prioritize the construction of additional gene deletion mutant strains to test for biofilm formation. These results are described in detail below.
Adhesion proteins are differentially expressed over time
We compared gene expression in cells that were adhered after 1.5 h to those cells that were inoculated in the same medium but did not adhere. The adhered vs. unadhered gene expression dataset contained 251 genes upregulated more than twofold and 157 genes downregulated more than twofold (Dataset 1). Gene Ontology analysis (Inglis et al., 2012) revealed that more than half of the upregulated genes were significantly enriched for genes involved in DNA synthesis (131 genes, P = 1.25 × 10−39), transcription (128 genes, P = 9.22 × 10−36), RNA processing (110 genes, P = 3.82 × 10−61) or translation (109 genes, P = 9.06 × 10−82), indicating that physical adherence strongly upregulates DNA and protein synthesis, likely in preparation for the large shift in morphology and physiology that accompanies biofilm formation. Downregulated genes were largely of unknown function but were significantly enriched for genes involved in oxidation‐reduction (33 genes, P = 2.15 × 10−7), carbohydrate transport (10 genes, P = 5.11 × 10−6), and sulfur amino acid metabolism (nine genes, P = 2.63 × 10−5). We conclude that as C. albicans yeast form cells adhere to a solid surface, they undergo extensive changes in gene expression, many of which are in preparation for the later stages in biofilm formation.
Because adhesion is a key part of biofilm formation, we examined expression of known or predicted adhesion proteins over time (Fig. 2). Based on the transcript levels of adhesion genes in biofilms compared with the multiple reference states, we can distinguish two classes of adhesion genes that are upregulated in biofilms, largely irrespective of which planktonic reference they are compared to. (i) A group of ten adhesion genes (ALS1, ALS2, ALS3, ALS4, EAP1, MSB2, PGA6, SIM1, ORF19.2449 and ORF19.5126) was induced early in biofilm formation. Seven of these genes (PGA6, ORF19.5126, ALS4, ALS2, SIM1, EAP1 and ALS1) were upregulated compared with the unadhered planktonic reference, indicating that adhesion of cells to a solid surface was required for their expression. Three of these genes (MSB2, ORF19.2449 and ALS3) are also induced in the unadhered planktonic reference, indicating that their expression is mediated by a cue other than contact with a solid surface, perhaps temperature or serum. This conclusion is consistent with previous work showing that ALS3 expression is upregulated within an hour of temperature shift from 30°C to 37°C but drops again after 2 h (Kadosh and Johnson, 2005; Argimón et al., 2007). (ii) A group of 10 additional adhesion genes (FAV2, HYR1, IFF4, IFF6, PGA32, PGA55, ORF19.3988, ORF19.4906, ORF19.5813 and ORF19.7539.1) was induced at later times in biofilm formation. We suggest that the early adhesion proteins are involved primarily with attaching cells to the substrate and the late adhesion proteins mediate cell‐to‐cell contact.
Figure 2.
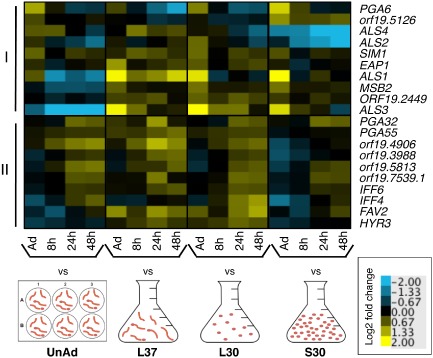
Two patterns of biofilm‐specific expression of adhesion genes are observed during biofilm development. Heat map of gene expression in C . albicans in biofilms at the indicated time points, compared with each of four reference conditions: unadhered cells (UnAd), log phase cells grown at 37°C (L37), log phase cells grown at 30°C (L30) and stationary phase cells grown at 30°C (S30). Shown are the median values of at least two biological replicates. On the y‐axis are known or predicted adhesion‐encoding genes differentially regulated at least twofold in at least one condition: I) genes upregulated early. II) genes upregulated late. Upregulated genes are yellow; downregulated genes are blue.
Biofilm metabolism is downregulated over time compared with stationary‐phase cells
It is known that metabolism is downregulated in mature biofilms (Yeater et al., 2007; Nobile et al., 2012); indeed, cells in biofilms are often likened to cells in stationary phase. To expose biofilm‐specific changes in gene expression, we chose stationary planktonic cells as one of the reference conditions for the microarray experiments. We examined the expression patterns of metabolic genes, specifically, those needed for glycolysis, the tricarboxylic acid cycle, the electron transport chain, fermentation, fatty acid metabolism, glycerol metabolism, galactose metabolism and ergosterol biosynthesis (Fig. 3). As might have been predicted, we found that many of these genes are downregulated over time during biofilm development; however, most are downregulated even more in mature biofilms than in stationary cultures. Compiled data from several studies indicates that glycolytic genes are expressed more highly in early biofilms and decrease over time (García‐Sánchez et al., 2004; Yeater et al., 2007; Nobile et al., 2012); this is consistent with our data. We also found that a few metabolism genes involved in glycerol degradation (GUT1 and GUT2) and genes involved in fermentation (ALD5, ALD6, ADH3 and ADH4) are upregulated over time in biofilms compared with stationary cultures, suggesting these processes are more important in mature biofilm cells than in stationary phase cells. Indeed, RHR2, which encodes a glycerol biosynthetic enzyme, is required for normal biofilm formation (Desai et al., 2013), suggesting that glycerol metabolism is important for biofilm cells. The importance of upregulating fermentation in biofilms is unknown, but we recently showed that the inside of a C. albicans biofilm is a hypoxic microenvironment (Fox et al., 2014), favorable for fermentation. We suggest that alcohols produced during fermentation may be used for energy or could be involved in cell signaling with neighboring yeast or other microbes; some alcohols have been shown to alter biofilm formation (Mukherjee et al., 2006; Martins et al., 2010; Rane et al., 2012; Chauhan et al., 2013). Overall, our analysis reveals that the regulation of metabolism genes in biofilms does not simply parallel that in stationary cultures.
Figure 3.
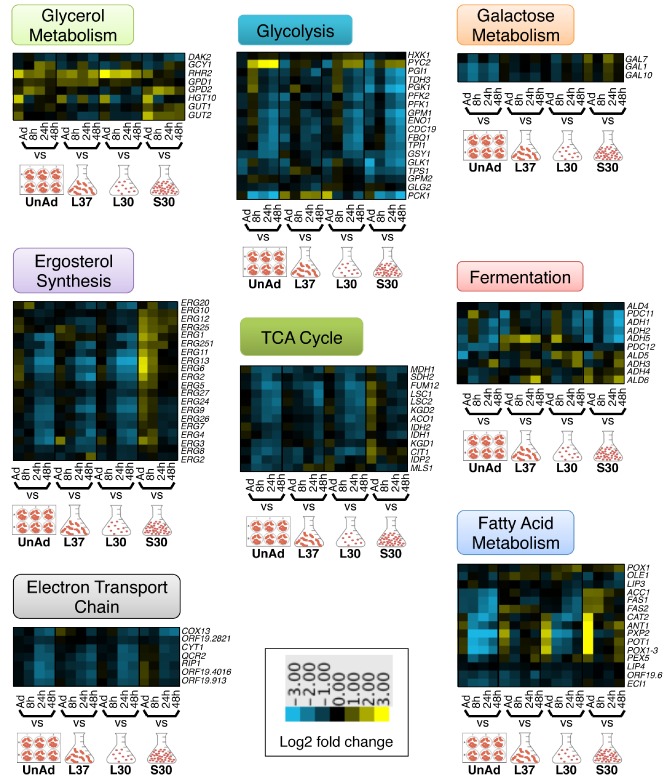
Metabolism is downregulated over time during biofilm development. Heat maps of gene expression in C . albicans in biofilms at the indicated time points, compared with each of four reference conditions: unadhered cells (UnAd), log phase cells grown at 37°C (L37), log phase cells grown at 30°C (L30) and stationary phase cells grown at 30°C (S30). Shown are the median values of at least two biological replicates. Each heat map shows expression of genes involved in the metabolic processes of glycolysis, the tricarboxylic acid cycle (TCA) cycle, the electron transport chain, fermentation, glycerol metabolism, galactose metabolism, fatty acid metabolism or ergosterol biosynthesis. Upregulated genes are yellow; downregulated genes are blue.
Newly identified transcription regulators play roles in biofilm development
We used the gene expression data to identify 11 previously untested transcription regulators whose transcript levels change temporally over biofilm development, and we added these to an existing transcription regulator deletion library (Homann et al., 2009; Fox et al., 2014) (see Materials and methods for details). We then blindly screened the expanded library of 192 mutant strains for biofilm formation in vitro at four time points: immediately after 1.5 h of adherence, and at 8, 24 and 48 h of biofilm growth (Figs 4, S2A). In addition to the six regulators identified previously (Nobile et al., 2012), we found seven other regulators, whose deletion resulted in a biofilm phenotype at at least one time point (Fig. 4), as measured by optical density of the cells remaining adhered to the bottom of the well after removal of the media (see Materials and methods). These seven were confirmed by testing independently constructed deletion strains (Fig. S2B). We also confirmed that none of the mutants had altered growth kinetics that could contribute to the observed phenotype (Fig. S3). One regulator deletion mutant (gal4Δ/Δ) formed biofilms with an increased optical density at 24 h and six regulator deletion mutants (rfx2Δ/Δ, rim101Δ/Δ, orf19.6874Δ/Δ, gzf3Δ/Δ, flo8Δ/Δ and rfg1Δ/Δ) formed biofilms with a decreased optical density at one or more time points (Fig. 4). ORF19.6874 was previously uncharacterized, and we named it Biofilm Partial Regulator 1 (BPR1).
Figure 4.
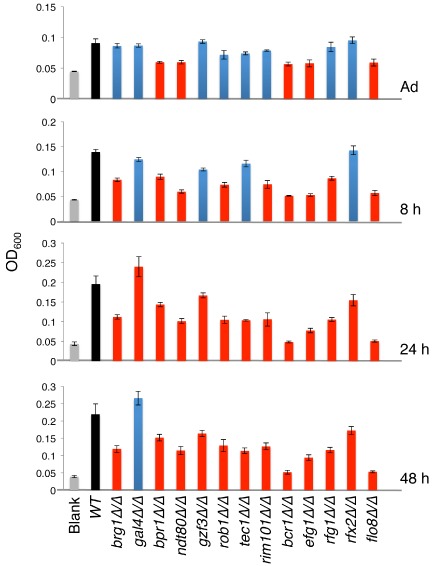
Thirteen transcription regulators are required for normal biofilm development in vitro in an optical density assay. The indicated wild type or mutant strains were adhered in a 96 well polystyrene plate for 90 min (labeled ‘Adhered’), unadhered cells were removed and adhered cells developed into biofilms for 8, 24 or 48 h. Biofilm formation was quantified by measuring OD600 of the cells adhered to the bottom of the well. Shown is the mean of at least six replicates, error bars are standard deviation. Gray bars are blank wells, black bars are wild type biofilms, blue bars are deletion strain biofilms not significantly different from wild type, red bars are deletion strain biofilms significantly different from wild type, P < 0.001 by Student's paired t‐test. See also Figs S2 and 3.
Of the seven additional regulator deletion mutants with optical density biofilm phenotypes, five of these formed biofilms morphologically distinct from those of wild type when examined by CSLM in vitro (Figs 5, S4); the phenotypes were rescued by adding back a wild‐type copy of each gene in the gene deletion background (Fig. S5). The flo8Δ/Δ mutant biofilm was severely deficient at all four time points, similar in severity to the biofilm phenotypes observed for the previously identified six master biofilm regulator mutants. At the 8 h time point, the rim101Δ/Δ mutant biofilm was thinner than the wild type and had fewer hyphae. The biofilm formed by rfg1Δ/Δ detached entirely from the surface and broke apart by 48 h. The gal4Δ/Δ mutant biofilm was thicker than wild type at the 24 h time point, in agreement with the optical density assay. The rfx2Δ/Δ mutant biofilm was thicker than wild type at 24 h, which is opposite of the optical density assay results. This discrepancy may be explained by the differences between the CSLM assay, which is performed on silicone squares in 12 well plates, and the OD assay, which is performed in 96 well plates. In the 96 well assay, it was observed that the rfx2Δ/Δ mutant strain formed a very thick pellicle at the liquid–air interface. This pellicle was not observed in the 12 well CSLM assay. Either the different substrate (silicone rather than polystyrene) or the larger wells may have allowed for additional cells that are more mobile and better able to incorporate into the biofilm to become a part of the biofilm in the larger‐well assay, resulting in an enhanced phenotype in the CSLM assay.
Figure 5.

Eleven transcription regulators are required for normal biofilm development in vitro by CSLM. The indicated wild type or mutant strains were adhered to silicone substrates for 90 min (labeled ‘Adhered’), unadhered cells were removed and adhered cells developed into biofilms for 8, 24 or 48 h. Biofilms were stained with conconavalin A – Alexa 594 and Syto 13 dyes, then imaged by CSLM. Images are maximum intensity projections of the top and side view. Representative images of at least three replicates are shown. Scale bars are 50 μm. See also Figs S4 and 5.
Because two of the mutant strains identified in the initial screen did not show clear differences from the reference strain by microscopy, we did not consider them further in this study. We next tested the five remaining regulator deletion strains with clear morphological phenotypes (flo8Δ/Δ, rim101Δ/Δ, gal4Δ/Δ, rfx2Δ/Δ and rfg1Δ/Δ) for biofilm formation in vivo, in a rat central venous catheter model (Andes et al., 2004). After the catheter was removed, SEM was used to examine biofilm morphology (Fig. 6). In order to minimize the number of animals used, we measured biofilm formation only at the time points at which a clear in vitro phenotype was observed, 8 h for rim101Δ/Δ, 24 h for gal4Δ/Δ, rfx2Δ/Δ and flo8Δ/Δ and 48 h for rfg1Δ/Δ. We found that three of these regulator mutant strains had phenotypes in vivo that matched those observed in vitro: gal4Δ/Δ and rfx2Δ/Δ were enhanced and flo8Δ/Δ was deficient at 24 h.
Figure 6.

Three new transcription regulators are required for normal biofilm development in vivo in a rat central venous catheter model. The indicated wild type or mutant strains were inoculated into rat intravenous catheters and developed into biofilms for 8, 24 or 48 h prior to visualization by SEM. The left column shows biofilms at 100×; the right column is at 1000×.
Three new transcriptional regulators fit in the biofilm regulatory network
As described above, three newly identified regulators, Gal4, Rfx2 and Flo8, are required for biofilm formation in vivo. To determine how they fit within the existing biofilm regulatory network, we tagged Gal4, Rfx2 and Flo8 and performed chromatin immunoprecipitation experiments followed by qPCR (ChIP‐qPCR), using probes to the intergenic regions upstream of each of the nine biofilm regulators (GAL4, RFX2, FLO8, BCR1, EFG1, NDT80, BRG1, TEC1 and ROB1) (Fig. 7). As shown in the figure, all three bind to the upstream regions of at least one of the previously identified master biofilm regulators. Moreover, the intergenic regions upstream of GAL4, RFX2 and FLO8 are each bound by at least one of the six previously identified master biofilm regulators (Nobile et al., 2012) – GAL4 and RFX2 are bound by Ndt80, and FLO8 is bound by Ndt80, Brg1 and Efg1, providing further evidence that these newly identified regulators are an integral part of the biofilm regulatory network.
Figure 7.
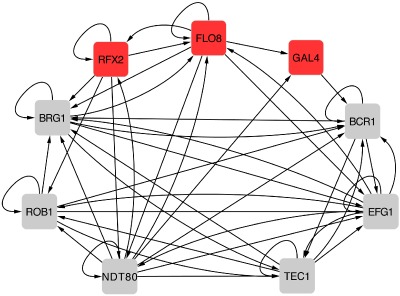
The expanded biofilm regulatory network model contains three newly identified regulators. Intergenic region binding data generated for Flo8, Rim101 and Gal4 was combined with binding data for Brg1, Bcr1, Rob1, Efg1, Ndt80 and Tec1 from Nobile et al. (2012) to determine which regulators bind upstream of the genes encoding each of the other regulators. An arrow indicates a binding event of the gene it points to. For example, Gal4 binds upstream of BCR1 and the region upstream of GAL4 is bound by Flo8 and Ndt80.
Discussion
Previous work identified six ‘master’ transcription regulators of biofilm formation in C. albicans (Ramage et al., 2002b; Nobile and Mitchell, 2005; Fox and Nobile, 2012; Nobile et al., 2012). In this paper, we describe three additional transcription regulators (Gal4, Rfx2 and Flo8) that are needed for proper biofilm formation both in vitro and in a rat in vivo model. These genes were identified by screening an expanded library of deletion mutants, based on transcriptional profiling described in this paper. Two of the newly identified genes (RFX2 and FLO8) were not represented in the libraries that had previously been screened; the third (GAL4) was revealed because we monitored multiple time points during biofilm formation. ChIP‐qPCR experiments showed that the new regulators are integrated into the complex biofilm transcription circuitry (see Fig. 7).
All three regulators were previously studied in contexts other than biofilm formation. Flo8 plays a role in hyphal growth and regulates a subset of genes also regulated by Efg1 (Cao et al., 2006), one of the biofilm master regulators. The flo8Δ/Δ mutant is defective in hyphal formation and is avirulent in a systemic tail vein mouse model (Cao et al., 2006). Our work also shows that the flo8Δ/Δ mutant is highly defective in biofilm formation in vivo in a rat central venous catheter model. The flo8Δ/Δ deletion phenotype was the most severe of the new regulators; the strain was defective in biofilm formation at all time points, and it closely resembled the biofilms formed by strains in which one of the six previously identified master regulators is deleted. Flo8 also plays a role in CO2 sensing during white‐to‐opaque morphological switching (Du et al., 2012), a finding that may provide a link between biofilm formation and white‐to‐opaque switching. FLO8 is expressed increasingly over time regardless of which planktonic reference condition is used (Dataset 1). Therefore, FLO8 expression appears to be biofilm specific, and upregulation is not dependent on whether the reference cells are filamentous or yeast form.
In the gal4Δ/Δ and rfx2Δ/Δ mutant strains, biofilm formation was enhanced rather than deficient, implying that these are negative regulators of biofilm formation. Consistent with this idea, both regulators are downregulated in biofilms relative to at least two of the reference conditions used in our expression array experiments (Dataset 1). Gal4 regulates glycolysis and carbohydrate metabolism, along with Tye7 (Askew et al., 2009), and our array data show that glycolysis and most metabolic pathways are downregulated over time during biofilm development. Tye7 also plays a role in biofilm formation; biofilms formed by the tye7Δ/Δ mutant contain more hyphae but are more fragile than wild‐type biofilms (Bonhomme et al., 2011). One possible explanation for the biofilm phenotypes exhibited by the gal4Δ/Δ and tye7Δ/Δ mutant strains is that the altered morphology is related to metabolism. As is proposed in Bonhomme et al. (2011) and Rossignol et al. (2009), glycolysis is an important part of survival during hypoxia and its misregulation could affect biofilm structure, which normally contains a hypoxic internal microenvironment (Fox et al., 2014). Rfx2 plays a role in hyphal formation; the rfx2Δ/Δ deletion mutant is hyperfilamentous, invasive and constitutively expresses several hyphal‐specific genes, including HWP1, ALS3, HYR1, ECE1 and CEK1 (Hao et al., 2009). Given these observations, it is perhaps not surprising that deletion of RFX2 results in enhanced biofilm formation.
Concurrent with the genetic approach, we examined gene expression in biofilms, measuring expression at four time points and comparing expression to four different planktonic reference conditions. García‐Sánchez et al. (2004) compared gene expression in mature biofilms with that of planktonic cultures and varied flow, aerobiosis and glucose concentration, whereas Murillo et al. (2005) compared gene expression in 6 h biofilms to that of planktonic culture. Yeater et al. (2007) measured gene expression in 6, 12 and 48 h biofilms, compared with planktonic cultures of the same incubation time. Nobile et al. (2012) measured gene expression in mature biofilms compared with equivalently aged planktonic cultures. In the only in vivo transcriptional profiling biofilm study to date, Nett et al. (2009) measured gene expression in biofilms grown in a rat central venous catheter model compared with in vitro planktonic cells. Our gene expression data adds a new set of comparisons to this body of C. albicans biofilm gene expression data. Our work is distinguished from previous studies in that we measured gene expression in newly adhered cells, 8, 24 and 48 h biofilms and compared these patterns with those of unadhered planktonic cells collected from the biofilm assay or to planktonic cells grown to log phase at 30°C, log phase at 37°C or stationary phase. Although there are many specific types of analyses that can be performed using our data (which is freely available), we discuss only two here. First, we found that there are two distinct expression classes of adhesion proteins involved in biofilm development: those that are enriched earlier and those that are enriched later during biofilm development. There is a third group of adhesion proteins whose expression does not differ between biofilm and planktonic cultures. These results indicate that the nature of adhesion in biofilms changes over time, consistent with the idea that biofilm cells must first adhere to a surface, then to one another, both in the yeast and filamentous forms. Moreover, at late times, at least some biofilm cells must detach from one another or from the surface to disperse to new locations. Second, we examined metabolism in biofilms over time and found that, regardless of which reference was used, metabolism is largely downregulated as a biofilm ages, consistent with the idea that a significant proportion of mature biofilm cells are in a metabolically quiescent state. This relative downregulation is observed even when mature biofilms are compared with stationary phase planktonic cultures; thus, cells in a mature biofilm are not metabolically equivalent to those in stationary phase.
Biofilms are a dynamic community of multiple cell types whose formation over time is orchestrated by a network of transcription regulators. In this paper, we have expanded the network of six master regulators to now include three additional regulators, Flo8, Gal4 and Rfx2, and have documented key gene expression changes that occur specifically in C. albicans cells that are forming biofilms.
Experimental procedures
Culture growth and maintenance
Candida albicans was streaked from a glycerol stock onto Yeast Peptone Dextrose (YPD) agar plates and grown at room temperature or 30°C. Suspension cultures were grown in YPD media at 30°C, with aeration.
Strain construction
Strains used are listed in Table S1 and include 165 strains published in Homann et al. (2009) as well as 19 strains published in Fox et al. (2014), and here we add eight additional homozygous gene deletion strains (some of which were previously published in Noble et al. (2010)), for a total of 192 strains in the updated transcription regulator knockout library. Gene deletion strains and complementation strains were constructed as described in Noble and Johnson (2005) and Fox et al. (2014), and Myc‐tagged strains were constructed as described in Nobile et al. (2012). Primers are listed in Table S2.
In vitro planktonic and biofilm culture conditions
Planktonic cultures and biofilms were grown in Spider medium (Liu, 1994) at 37°C at 200 rpm in an ELMI incubator (ELMI, Ltd. Riga, Latvia) for biofilms and 225 rpm in a New Brunswick Scientific incubator for planktonic cultures, as described in Nobile et al. (2012). Saturated overnight cultures grown at 30°C in YPD (2% Bacto Peptone, 2% dextrose, 1% yeast extract) were inoculated into Spider medium (OD600 = 0.02 in 50 ml for planktonic cultures or OD600 = 0.5 in 4 ml in a bovine serum‐treated 6 well plate for biofilm cultures). Stationary phase planktonic cultures were grown to an OD600 = 20 at 30°C and log phase planktonic cultures were grown to an OD600 = 0.8 at either 30°C or 37°C, shaking. Biofilm cultures were adhered for 90 min, washed with 4 ml PBS, then fresh 4 ml of Spider media was added, and biofilms were grown for the indicated times (8, 24 or 48 h). Adhered and unadhered cells were collected after the 90 min adherence step.
Biofilm formation assay by optical density at 600 nm
Biofilms were grown as described above but without bovine‐serum treating the plates, and in a volume of 200 μl, in 96 well polystyrene plates (BD Falcon 351172). The plates were sealed with breathable seals (Excel Scientific BS25) to prevent cross‐contamination between wells and plates were incubated at 350 rpm, 37°C. At least six replicates were performed for each mutant strain. At each time point, wells were washed with 200 μl PBS, all liquid was aspirated off, and optical density was read at 600 nm on a Tecan Infinite M200 plate reader (Tecan Systems).
Growth assay
The growth curves were performed using methods previously described in Wells et al. (2015). Briefly, cells from an overnight culture grown in YPD at 30°C were inoculated into 100 μl of YPD media at a starting OD600 = 0.01. The assay was performed in flat‐bottom 96 well plates (BD Falcon) with n = 3 for each strain and grown in a Tecan Infinite M1000 reader (Tecan Systems) at 30°C with 408 rpm orbital shaking. An optical density (OD595) measurement was taken every 15 min for 24 h.
Confocal scanning laser microscopy
Biofilms were imaged as described in Nobile et al. (2012). In brief, biofilms were formed on silicone squares for various time points. Biofilms were stained with 50 μg ml−1 of concanavalin A Alexa Fluor 594 conjugate (Molecular Probes C11253) for 1 h, 200 rpm, in the dark. Silicone squares were placed in a 6 well plate and covered with water for imaging by Confocal Scanning Laser Microscopy (CSLM) (Nikon Instruments Inc, Melville, New York, USA) under a 40X/0.08W Nikon water‐dipping objective lens (Nikon Instruments Inc, Melville, New York, USA). A 561 nm laser line was used, and images were acquired with Nikon EX‐C1 Version 3.80 software. Images were taken every 0.8 μm through the Z axis of the biofilm, and images were assembled into maximum intensity Z‐stack projections with Nikon NIS Elements Version 3.00 software (Nikon Instruments Inc, Melville, New York, USA).
In vivo rat catheter biofilm model
The rat central‐venous catheter infection model was used as described previously (Andes et al., 2004; Nobile et al., 2012). In brief, specific‐pathogen‐free male Sprague‐Dawley rats weighing 400 g (Harlan Sprague‐Dawley) were used. A heparinized (100 U ml−1) polyethylene catheter with 0.76 mm inner and 1.52 mm outer diameters was inserted into the external jugular vein and advanced to a site above the right atrium. The catheter was secured to the vein with the proximal end tunneled subcutaneously to the midscapular space and externalized through the skin. The catheters were inserted 24 h prior to infection to permit a conditioning period for deposition of host protein on the catheter surface. Infection was achieved by intraluminal instillation of 500 μl C. albicans cells (106 cells ml−1). After a 4 h dwelling period, the catheter volume was withdrawn and the catheter flushed with heparinized 0.15 M NaCl. Catheters were removed after 8, 24 or 48 h of C. albicans infection to assay biofilm development on the intraluminal surface by SEM. Catheter segments were washed with phosphate buffered saline (PBS), pH 7.2, fixed in 1% glutaraldehyde/4% formaldehyde, washed again with PBS for 5 min and placed in 1% osmium tetroxide for 30 min. Following PBS washing, the samples were dehydrated in a series of 10 min ethanol washes (30%, 50%, 70%, 85%, 95% and 100%), followed by critical point drying. Specimens were mounted on aluminum stubs, sputter coated with gold, and imaged using a Leo 1530 scanning electron microscope (Leo Electron Microscopy Ltd, Cambridge, Cambridgeshire, United Kingdom) in the high‐vacuum mode at 3 kV. Images were assembled using Adobe Photoshop Version 7.0.1 software (Adobe Systems Inc, San Jose, California, USA).
Gene expression microarrays
Planktonic and biofilm cells were grown as described above in flasks (for planktonic) or in bovine‐serum coated 6 well plates (for biofilms). Biofilm cells were harvested with a sterile transfer pipet, biofilm or planktonic cells were centrifuged at 3,000× g for 5 min, supernatant was removed and cell pellets were frozen in liquid nitrogen and stored at −80°C. Total RNA was extracted using the Ambion RiboPure‐Yeast RNA kit (AM1926) (Life Technologies, Grand Island, New York, USA) according to the manufacturer's protocol. Synthesis of cDNA and dye coupling was performed as described in Nobile et al. (2009). Gene expression microarrays from Agilent Technologies (AMID #020166) were used as described in Nobile et al. (2012). LOWESS normalization of microarray data was performed as previously described (Lohse and Johnson, 2010). At least two independent replicates were performed at each time point. Microarray data are reported in Dataset 1, and raw gene expression array data are stored at the Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo, accession #GSE61143).
Chromatin immunoprecipitation quantitative real‐time PCR (ChIP‐qPCR)
The Flo8, Gal4 and Rfx2 Myc‐tagged strains (Table S1) were grown under standard biofilm conditions for 24 h, and the biofilm cells were harvested for chromatin immunoprecipitation as described in Nobile et al. (2012). ChIP‐qPCR was performed using primers in the promoter regions of FLO8, GAL4, RFX2, BCR1, TEC1, EFG1, NDT80, BRG1 and ROB1 (Table S2).
Identification of transcription regulators with temporal expression patterns
We used an algorithm (see Supporting information) to search for gene expression trends in which the expression values were increasing or decreasing over time in at least two out of four of the datasets, using a starting list of 286 regulators. We also scanned the data by eye to include genes with expression values that did not fit the slope used in the algorithm but still showed patterns of increasing or decreasing expression. This analysis resulted in a list of 91 regulator candidates. We then applied the criteria that at least one gene expression value must represent at least a twofold difference from the reference (79 regulators), have not been previously tested and have no known biofilm phenotype (30 regulators), have not been annotated as essential (27 regulators) and must have their promoter bound by at least one of the six ‘master’ transcription regulators identified in Nobile et al. (2012). This resulted in 11 candidate transcription regulators (ORF19.7521, ORF19.4342, RFX2, WOR3, ZPR1, MDM34, ECM22, ORF19.7098, ADR1, ORF19.5953, and ORF19.7397), which were included in the target gene set for constructing deletion mutant strains.
Supporting information
Supporting information
Acknowledgements
We thank Julie Takagi and Jorge Mendoza for technical assistance. We are grateful for use of the Nikon Imaging Center at UCSF. This study was supported by National Institutes of Health (NIH) grant R01AI083311 (to A.D.J.), a UCSF Program for Breakthrough Biomedical Research (PBBR) award, funded in part by the Sandler Foundation, NIH grant R01AI073289 (to D.R.A.), and NIH grant R00AI100896 (to C.J.N.). E.P.F. was supported by NIH fellowship T32AI060537. J.E.N. was support by NIH grant K08AI108727 and the Burroughs Welcome Fund. The authors acknowledge use of instrumentation supported by the UW MRSEC (DMR‐1121288) and the UW NSEC (DMR‐0832760). A.D.J. and C.J.N. are co‐founders of BioSynesis Inc., a company developing inhibitors and diagnostics of C. albicans biofilm formation; however, there are no conflicts of interest to declare with regard to this paper.
References
- Andes, D. , Nett, J. , Oschel, P. , Albrecht, R. , Marchillo, K. , and Pitula, A. (2004) Development and characterization of an in vivo central venous catheter Candida albicans biofilm model. Infect Immun 72: 6023–6031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Argimón, S. , Wishart, J.A. , Leng, R. , Macaskill, S. , Mavor, A. , Alexandris, T. , et al (2007) Developmental regulation of an adhesin gene during cellular morphogenesis in the fungal pathogen Candida albicans . Eukaryot Cell 6: 682–692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Askew, C. , Sellam, A. , Epp, E. , Hogues, H. , Mullick, A. , Nantel, A. , and Whiteway, M. (2009) Transcriptional regulation of carbohydrate metabolism in the human pathogen Candida albicans . PLoS Pathog 5: e1000612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baillie, G.S. , and Douglas, L.J. (1999) Role of dimorphism in the development of Candida albicans biofilms. J Med Microbiol 48: 671–679. [DOI] [PubMed] [Google Scholar]
- Bonhomme, J. , and d'Enfert, C. (2013) Candida albicans biofilms: building a heterogeneous, drug‐tolerant environment. Curr Opin Microbiol 16: 398–403. [DOI] [PubMed] [Google Scholar]
- Bonhomme, J. , Chauvel, M. , Goyard, S. , Roux, P. , Rossignol, T. , and D'Enfert, C. (2011) Contribution of the glycolytic flux and hypoxia adaptation to efficient biofilm formation by Candida albicans . Mol Microbiol 80: 995–1013. [DOI] [PubMed] [Google Scholar]
- Cao, F. , Lane, S. , Raniga, P.P. , Lu, Y. , Zhou, Z. , Ramon, K. , et al (2006) The Flo8 transcription factor is essential for hyphal development and virulence in Candida albicans . Mol Biol Cell 17: 295–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cauda, R. (2009) Candidaemia in patients with an inserted medical device. Drugs 69 (Suppl. 1): 33–38. [DOI] [PubMed] [Google Scholar]
- Chandra, J. , Kuhn, D.M. , Mukherjee, P.K. , Hoyer, L.L. , McCormick, T. , and Ghannoum, M.A. (2001) Biofilm formation by the fungal pathogen Candida albicans: development, architecture, and drug resistance. J Bacteriol 183: 5385–5394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chauhan, N.M. , Shinde, R.B. , and Karuppayil, S.M. (2013) Effect of alcohols on filamentation, growth, viability and biofilm development in Candida albicans . Braz J Microbiol 44: 1315–1320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai, J.V. , Bruno, V.M. , Ganguly, S. , Stamper, R.J. , Mitchell, K.F. , Solis, N. , et al (2013) Regulatory role of glycerol in Candida albicans biofilm formation. MBio 4: 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du, H. , Guan, G. , Xie, J. , Cottier, F. , Sun, Y. , Jia, W. , et al (2012) The transcription factor Flo8 mediates CO2 sensing in the human fungal pathogen Candida albicans . Mol Biol Cell 23: 2692–2701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finkel, J.S. , Xu, W. , Huang, D. , Hill, E.M. , Desai, J.V. , Woolford, C.A. , et al (2012) Portrait of Candida albicans adherence regulators. PLoS Pathog 8: e1002525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox, E.P. , and Nobile, C.J. (2012) A sticky situation: untangling the transcriptional network controlling biofilm development in Candida albicans . Transcription 3: 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox, E.P. , Cowley, E.S. , Nobile, C.J. , Hartooni, N. , Newman, D.K. , and Johnson, A.D. (2014) Anaerobic bacteria grow within Candida albicans biofilms and induce biofilm formation in suspension cultures. Curr Biol 24: 2411–2416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- García‐Sánchez, S. , Aubert, S. , Iraqui, I. , Janbon, G. , Ghigo, J. , and D'Enfert, C. (2004) Candida albicans biofilms: a developmental state associated with specific and stable gene expression patterns. Eukaryot Cell 3: 536–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao, B. , Clancy, C.J. , Cheng, S. , Raman, S.B. , Iczkowski, K.A. , and Nguyen, M.H. (2009) Candida albicans RFX2 encodes a DNA binding protein involved in DNA damage responses, morphogenesis, and virulence. Eukaryot Cell 8: 627–639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawser, S.P. , and Douglas, L.J. (1994) Biofilm formation by Candida species on the surface of catheter materials in vitro. Infect Immun 62: 915–921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawser, S.P. , Baillie, G.S. , and Douglas, L.J. (1998) Production of extracellular matrix by Candida albicans biofilms. J Med Microbiol 47: 253–256. [DOI] [PubMed] [Google Scholar]
- Homann, O.R. , Dea, J. , Noble, S.M. , and Johnson, A.D. (2009) A phenotypic profile of the Candida albicans regulatory network. PLoS Genet 5: e1000783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inglis, D. , Arnaud, M. , Binkley, J. , Shah, P. , Skrzypek, M. , Wymore, F. , et al (2012) The Candida genome database incorporates multiple Candida species: multispecies search and analysis tools with curated gene and protein information for Candida albicans and Candida glabrata . Nucleic Acids Res 40: D667–D674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jabra‐Rizk, M.A. , Falkler, W.A. , and Meiller, T.F. (2004) Fungal biofilms and drug resistance. Emerg Infect Dis 10: 14–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kadosh, D. , and Johnson, A.D. (2005) Induction of the Candida albicans filamentous growth program by relief of transcriptional repression: a genome‐wide analysis. Mol Biol Cell 16: 2903–2912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kayingo, G. , Martins, A. , Andrie, R. , Neves, L. , Lucas, C. , and Wong, B. (2009) A permease encoded by STL1 is required for active glycerol uptake by Candida albicans . Microbiology 155: 1547–1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kojic, E.M. , Darouiche, R.O. , and Biofilms, C. (2004) Candida infections of medical devices. Society 17: 255–267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunze, D. , Melzer, I. , Bennett, D. , Sanglard, D. , MacCallum, D. , Nörskau, J. , et al (2005) Functional analysis of the phospholipase C gene CaPLC1 and two unusual phospholipase C genes, CaPLC2 and CaPLC3, of Candida albicans . Microbiology 151: 3381–3394. [DOI] [PubMed] [Google Scholar]
- Liu, H. , Köhler, J. , and Fink, G.R. (1994) Suppression of hyphal formation in Candida albicans by mutation of a STE12 homolog. Science 266: 1723–1726. [DOI] [PubMed] [Google Scholar]
- López, D. , Vlamakis, H. , and Kolter, R. (2010) Biofilms. Cold Spring Harb Perspect Biol 2: a000398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lohse, M.B. , and Johnson, A.D. (2010) Temporal anatomy of an epigenetic switch in cell programming: the white‐opaque transition of C. albicans . Mol Microbiol 78: 331–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martins, M. , Henriques, M. , Azeredo, J. , Rocha, S.M. , Coimbra, M.A. , and Oliveira, R. (2010) Candida species extracellular alcohols: production and effect in sessile cells. J Basic Microbiol 50: S89–S97. [DOI] [PubMed] [Google Scholar]
- Mukherjee, P.K. , Mohamed, S. , Chandra, J. , Kuhn, D. , Liu, S. , Antar, O.S. , et al (2006) Alcohol dehydrogenase restricts the ability of the pathogen Candida albicans to form a biofilm on catheter surfaces through an ethanol‐based mechanism. Infect Immun 74: 3804–3816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murillo, L.A. , Newport, G. , Lan, C. , Dungan, J. , Agabian, N.M. , and Habelitz, S. (2005) Genome‐wide transcription profiling of the early phase of biofilm formation by Candida albicans . Eukaryot Cell 4: 1562–1573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nett, J. , Lincoln, L. , Marchillo, K. , Massey, R. , Holoyda, K. , Hoff, B. , et al (2007) Putative role of beta‐1,3 glucans in Candida albicans biofilm resistance. Antimicrob Agents Chemother 51: 510–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nett, J.E. , Lepak, A.J. , Marchillo, K. , and Andes, D.R. (2009) Time course global gene expression analysis of an in vivo Candida biofilm. J Infect Dis 200: 307–313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nett, J.E. , Sanchez, H. , Cain, M.T. , and Andes, D.R. (2010a) Genetic basis of Candida biofilm resistance due to drug‐sequestering matrix glucan. J Infect Dis 202: 171–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nett, J.E. , Crawford, K. , Marchillo, K. , and Andes, D.R. (2010b) Role of Fks1p and matrix glucan in Candida albicans biofilm resistance to an echinocandin, pyrimidine, and polyene. Antimicrob Agents Chemother 54: 3505–3508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nett, J.E. , Marchillo, K. , and Andes, D.R. (2012) Host‐Fungus Interactions. Totowa, NJ: Humana Press. [Google Scholar]
- Nobile, C.J. , and Mitchell, A.P. (2005) Regulation of cell‐surface genes and biofilm formation by the C. albicans transcription factor Bcr1p. Curr Biol 15: 1150–1155. [DOI] [PubMed] [Google Scholar]
- Nobile, C.J. , and Mitchell, A.P. (2007) Microbial biofilms: e pluribus unum. Curr Biol 17: R349–R353. [DOI] [PubMed] [Google Scholar]
- Nobile, C.J. , Andes, D.R. , Nett, J.E. , Smith, F.J. , Yue, F. , Phan, Q.‐T. , et al (2006) Critical role of Bcr1‐dependent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoS Pathog 2: e63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobile, C.J. , Schneider, H.A. , Nett, J.E. , Sheppard, D.C. , Filler, S.G. , Andes, D.R. , and Mitchell, A.P. (2008) Complementary adhesin function in C. albicans biofilm formation. Curr Biol 18: 1017–1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobile, C.J. , Nett, J.E. , Hernday, A.D. , Homann, O.R. , Deneault, J.‐S. , Nantel, A. , et al (2009) Biofilm matrix regulation by Candida albicans Zap1. PLoS Biol 7: e1000133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobile, C.J. , Fox, E.P. , Nett, J.E. , Sorrells, T.R. , Mitrovich, Q.M. , Hernday, A.D. , et al (2012) A recently evolved transcriptional network controls biofilm development in Candida albicans . Cell 148: 126–138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noble, S.M. , and Johnson, A.D. (2005) Strains and strategies for large‐scale gene deletion studies of the diploid human fungal pathogen Candida albicans . Eukaryot Cell 4: 298–309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noble, S.M. , French, S. , Kohn, L.A. , Chen, V. , and Johnson, A.D. (2010) Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat Genet 42: 590–598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raad, I. , Hanna, H. , and Maki, D. (2007) Intravascular catheter‐related infections: advances in diagnosis, prevention, and management. Lancet Infect Dis 7: 645–657. [DOI] [PubMed] [Google Scholar]
- Ramage, G. , VandeWalle, K. , López‐Ribot, J.L. , and Wickes, B.L. (2002a) The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans . FEMS Microbiol Lett 214: 95–100. [DOI] [PubMed] [Google Scholar]
- Ramage, G. , Saville, S.P. , Wickes, B.L. , and López‐Ribot, J.L. (2002b) Inhibition of Candida albicans biofilm formation by farnesol, a quorum‐sensing molecule. Appl Environ Microbiol 68: 5459–5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rane, H.S. , Bernardo, S.M. , Walraven, C.J. , and Lee, S.A. (2012) In vitro analyses of ethanol activity against Candida albicans biofilms. Antimicrob Agents Chemother 56: 4487–4489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossignol, T. , Ding, C. , Guida, A. , D'Enfert, C. , Higgins, D.G. , and Butler, G. (2009) Correlation between biofilm formation and the hypoxic response in Candida parapsilosis . Eukaryot Cell 8: 550–559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seddiki, S.M.L. , Boucherit‐Otmani, Z. , Boucherit, K. , Badsi‐Amir, S. , Taleb, M. , and Kunkel, D. (2013) Assessment of the types of catheter infectivity caused by Candida species and their biofilm formation. First study in an intensive care unit in Algeria. Int J Gen Med 6: 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sengupta, M. , and Datta, A. (2003) Two membrane proteins located in the Nag regulon of Candida albicans confer multidrug resistance. Biochem Biophys Res Commun 301: 1099–1108. [DOI] [PubMed] [Google Scholar]
- Tobudic, S. , Kratzer, C. , Lassnigg, A. , and Presterl, E. (2012) Antifungal susceptibility of Candida albicans in biofilms. Mycoses 55: 199–204. [DOI] [PubMed] [Google Scholar]
- Uppuluri, P. , Chaturvedi, A.K. , Srinivasan, A. , Banerjee, M. , Ramasubramaniam, A.K. , Köhler, J.R. , et al (2010) Dispersion as an important step in the Candida albicans biofilm developmental cycle. PLoS Pathog 6: e1000828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wells, M.L. , Washington, O.L. , Hicks, S.N. , Nobile, C.J. , Hartooni, N. , Wilson, G.M. , et al (2015) Post‐transcriptional regulation of transcript abundance by a conserved member of the tristetraprolin family in Candida albicans . Mol Microbiol 95: 1036–1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeater, K.M. , Chandra, J. , Cheng, G. , Mukherjee, P.K. , Zhao, X. , Rodriguez‐Zas, S.L. , et al (2007) Temporal analysis of Candida albicans gene expression during biofilm development. Microbiology 153: 2373–2385. [DOI] [PubMed] [Google Scholar]
- Zarnowski, R. , Westler, W.M. , Lacmbouh, G.A. , Marita, J.M. , Bothe, J.R. , Bernhardt, J. , et al (2014) Novel entries in a fungal biofilm matrix encyclopedia. MBio 5: e01333–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information