

Abstract
Chronic pain is one of the most prevalent health problems in our modern world, with millions of people debilitated by conditions such as back pain, headache and arthritis. To address this growing problem, many people are turning to mind–body therapies, including meditation, yoga and cognitive behavioural therapy. This article will review the neural mechanisms underlying the modulation of pain by cognitive and emotional states — important components of mind–body therapies. It will also examine the accumulating evidence that chronic pain itself alters brain circuitry, including that involved in endogenous pain control, suggesting that controlling pain becomes increasingly difficult as pain becomes chronic.
Pain is a complex sensory and emotional experience that can vary widely between people and even within an individual depending on the context and meaning of the pain and the psychological state of the person. Cognitive and emotional factors have a surprisingly important influence on pain perception. A negative expectation can completely reverse the analgesic effect of a clinical dose of the opioid agonist remifentanil1, whereas the expectation of pain relief is an important component of placebo analgesia2. Clinical and experimental studies show that even a simple psychological manipulation, such as distraction, can have a powerful effect on our perception of pain3. Our emotional state also has an enormous influence on pain; a negative emotional state increases pain, whereas a positive state lowers pain4. Not surprisingly, complex emotional states such as empathy, which incorporate emotional and cognitive factors, alter the way an individual feels pain5. Brain imaging studies have allowed us to examine the neural basis of psychological modulation of pain (BOX 1). These studies reveal that activity in afferent pain pathways is altered by the attentional state, positive and negative emotions, empathy and the administration of a placebo6. Additionally, imaging studies show that psychological factors activate intrinsic modulatory systems in the brain, including those involved in opioid-related pain relief. Furthermore, multiple descending pain modulatory systems are implicated in pain relief, with attentional states and emotions activating different systems in the brain.
Box 1. Real-time functional MRI and pain control.
Real-time functional MRI (rtfMRI) provides direct feedback on the activation of a specific brain area, allowing a person to learn how to control the activity in this brain region. RtfMRI feedback has recently been used to successfully train healthy controls to modulate the activation of their own anterior cingulate cortex (ACC) in order to alter their pain experience169. Participants were given several cognitive strategies that they could use to manipulate activity in the ACC on the basis of previous studies that showed activation of this region. Participants eventually learned to control the activity in the ACC, and the more control they developed, the more they were able to decrease the intensity of the experimental pain stimulus. Importantly, using the same paradigm, patients with chronic pain were able to decrease the level of their ongoing pain by controlling the activation of their ACC. Thus, rtfMRI could potentially be used as a tool to help patients directly activate pain modulatory systems and better control their pain. In the above study, subjects had to try to modify activity in a specific brain region to modify pain. However, as we know that multiple brain regions are involved in the cognitive and emotional control of pain, training subjects to control the activity in multiple brain regions associated with a particular task170 and applying this skill to pain modulation, might maximize the cognitive control of pain.
In parallel with our increased understanding of the circuitry underlying the psychological modulation of pain, recent evidence shows that chronic pain can lead to anatomical and functional alterations in this same circuitry, resulting not only in pain but also in altered cognition and affect. In Eastern cultures, the power of the mind to control one’s physical state with the use of practices such as meditation and yoga has been recognized for centuries. Today, people in the Western world have begun to adopt these techniques to reduce stress and control pain, and many pain sufferers now recognize the potential therapeutic effects of psychological procedures. In line with patient reports, new experimental studies show that interventions such as meditation not only decrease pain but also have powerful protective effects on brain grey matter and connectivity within pain modulatory circuits7–12. The wealth of new studies showing alterations in the brains of patients with chronic pain can now be integrated with our increased understanding of the brain circuitry involved in the psychological modulation of pain, allowing us to hypothesize a negative-feedback loop between impaired pain modulatory circuitry and pain processing, leading not only to increased chronic pain but also to cognitive and emotional deficits that are comorbid with the pain (FIG. 1).
Figure 1. Feedback loops between pain, emotions and cognition.
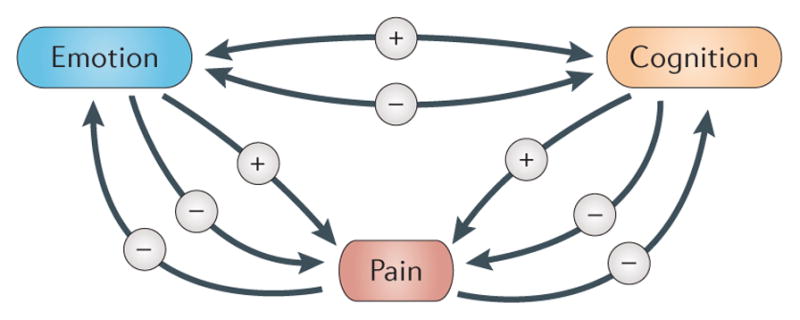
Pain can have a negative effect on emotions and on cognitive function. Conversely, a negative emotional state can lead to increased pain, whereas a positive state can reduce pain. Similarly, cognitive states such as attention and memory can either increase or decrease pain. Of course, emotions and cognition can also reciprocally interact. The minus sign refers to a negative effect and the plus sign refers to a positive effect.
Brain regions involved in the experience of pain
Multiple pathways in the CNS are involved in pain processing. Human brain imaging studies have revealed consistent cortical and subcortical networks that are activated by pain, including sensory, limbic and associative regions. The brain areas most commonly activated by noxious stimuli in human brain imaging studies are the primary somatosensory cortex (S1), secondary somatosensory cortex (S2), anterior cingulate cortex (ACC), insula, prefrontal cortex (PFC), thalamus and cerebellum13 (FIG. 2). Neural activation in these areas is consistent with anatomical and electrophysiological studies that show possible afferent nociceptive connectivity to these regions13–20. Other regions have also been shown to have nociceptive input through anatomical and electrophysiological studies, and these findings have been confirmed by brain imaging studies. These regions, which include the nucleus accumbens and amygdala21,22, probably receive nociceptive input through spinoparabrachial–amygdala projections23, as well as the periaqueductal grey (PAG)24, which receives nociceptive input through spinoreticular pathways25.
Figure 2. Afferent pain pathways include multiple brain regions.

Afferent nociceptive information enters the brain from the spinal cord. Afferent spinal pathways include the spinothalamic, spinoparabrachio–amygdaloid and spinoreticulo–thalamic pathways. Nociceptive information from the thalamus is projected to the insula, anterior cingulate cortex (ACC), primary somatosensory cortex (S1) and secondary somatosensory cortex (S2), whereas information from the amygdala (AMY) is projected to the basal ganglia (BG). See the main text for references. PAG, periaqueductal grey; PB, parabrachial nucleus; PFC, prefrontal cortex.
As would be expected, the multiple pain-related brain pathways are important for different aspects of the pain experience. Pain involves sensations with qualities such as stinging, burning or aching and has identifiable locations and durations. However, what makes pain ‘pain’ is usually the affective component of the experience — that is, how unpleasant it is. It is the unpleasantness that motivates the individual to engage in a behaviour, whether it is to flee, fight or freeze. In some cases, a painful sensation can be experienced as pleasurable, such as a deep massage, and in that case it creates a positive motivational state. Brain imaging and lesion studies implicate different brain regions in these different aspects of the pain experience. The somatosensory cortices (S1 and S2) encode information about sensory features, such as the location and duration of pain26–30. Alternatively, the ACC and insula, which have long been considered components of the limbic (emotional) part of the brain31,32, are more important for encoding the emotional and motivational aspects of pain. Patients with lesions of these regions show altered emotional responses to pain33–36, and imaging and stimulation studies show a relationship between emotional and motivational aspects of pain perception and neural activation in the ACC and insula37–40. Nevertheless, accumulating evidence implicates the insula in both pain sensation and pain affect, with the anterior portion being particularly important for the subjective experience of pain41,42.
Some studies show that a noxious insult to the body is not necessary for the experience of pain or for the activation of pain pathways. A number of studies have found that simply observing another individual in pain activates some pain-related brain regions (see REF. 43 for a meta-analysis), with such activation being stronger when one observes a loved one in pain rather than a stranger44. The activation of these regions in the absence of a pain stimulus could induce a state of ‘priming’ in the brain, in which a noxious stimulus given after emotionally priming a subject elicits an enhanced pain experience. This is exactly what was found in a study in which the authors created a state of positive or negative empathy towards someone experiencing pain5. When subjects were positively empathizing with the individual in pain, their own pain experience was enhanced. Similar empathetically enhanced pain responses have been reported in rodents. In another study, the authors produced intriguing evidence that mice are influenced by the pain state of a cage-mate45. When mice see cage-mates in pain, they show increased pain behaviours themselves — a phenomenon the authors call ‘emotional contagion’45.
Empathy is a complex cognitive and emotional state that could involve alterations in attention, emotions and expectations. Nevertheless, simpler cognitive manipulations, such as imagining or anticipating a painful experience, can also lead to activation of pain pathways. One study found that imagining a painful hand movement in an amputated arm activated pain pathways, involving the S1, insula and thalamus46. Similarly, a number of studies have shown that when an individual is anticipating the onset of pain, pain-related brain regions that are important for pain sensation, pain affect and pain modulation become activated, including the somatosensory cortices, ACC, insula, PFC and PAG47–53.
Together, these findings indicate that brain regions receive afferent pain input via multiple routes and that, at least in higher brain regions, there is no ‘labelled line’ between the peripheral nociceptive input and the final cortical representation of that input. This idea is consistent with evidence of multiple descending modulatory circuits, which either enhance or reduce the afferent signal at multiple regions throughout the neuraxis, from the cortex down to the spinal cord.
Attention and emotion influence pain differently
For centuries there have been anecdotal accounts of individuals apparently experiencing little or no pain in situations that by all accounts should be excruciating. Beecher54 described wounded soldiers who felt little pain from grievous injuries in the battlefield but reacted strongly to minor painful procedures when they returned home. However, Western medicine has traditionally ignored the role of the mind in pain control and has focused on pharmacological treatments of pain. Nevertheless, there is now a growing emphasis on mind–body techniques for controlling pain, with many patients with chronic pain turning to cognitive behavioural therapy, yoga, meditation, hypnosis and relaxation procedures to reduce their pain. These techniques are complex but most have both a cognitive component, such as attentional focus, and an emotional component. Growing evidence shows that these therapies can reduce both chronic and acute pain (for reviews, see REFS 55–57).
Both attentional and emotional factors are known to modulate pain perception in the clinic and in the laboratory3,58,59 (BOX 2). However, the nature and mechanisms of this modulation differ60–62. Focusing on pain increases the perceived intensity of the sensation, whereas a negative emotional state increases the perceived unpleasantness of the pain without altering the intensity4,59–61 (FIG. 3). One study found that emotional valence influenced pain ratings and a spinal nociceptive reflex in the same direction but distraction reduced pain while increasing the reflex62. Thus, it appears that different systems may be involved in the modulation of pain by attention and emotions.
Box 2. Cognitive control of pain in rodents: environmental enrichment.
Does enhancing a rodent’s social life and presenting distracting stimuli alter pain in the same way as has been found in humans? Evidence from studies of environmental enrichment indicates that the answer is ‘yes’. Environmental enrichment in rodents enhances the complexity of the living conditions of experimental animals: for example, rats or mice are housed in groups in larger cages with toys, tunnels, running wheels, and so on. In healthy animals, environmental enrichment improves learning and memory, enhances neuro- and gliogenesis in the cortex and hippocampus and induces dendritic and synaptic changes (see REF. 171 for a review). Enrichment can be classified as social (such as more animals per group), physical (such as access to running wheels) or both.
The beneficial effects of environmental enrichment have been investigated in rodent pain models. In rats, environmental enrichment may increase the speed of healing after an inflammatory injury172,173 and reduces the spinal astroglial response to peripheral inflammation174. In mice, enrichment reduces mechanical hypersensitivity after a nerve injury175 but may actually increase chronic inflammatory responses176. The contradictions in response could partly be due to the stress of group housing in rats and mice — rodents establish complex social hierarchies, and dominance or defeat can be inherently stressful.
Recent work has focused on physical enrichment alone as a modulator of pain behaviours. In one study, the authors used a shock-avoidance paradigm to force rats to run on a treadmill after nerve injury and saw reductions in thermal and tactile hypersensitivity within 3 weeks after injury, which reversed when exercise was stopped177. The effects of exercise were opioid-dependent — opioid antagonists reversed or prevented the beneficial effects of exercise, and increased levels of endogenous opioids were seen in the rostroventral medulla and periaqueductal grey, which are both key areas in the descending modulation of pain. However, as the running was motivated by shock avoidance, analgesia could be stress-related, and the shock could have been more stressful for nerve-injured animals. Thus, voluntary exercise paradigms are needed to attribute the reductions in pain specifically to exercise. Unfortunately, as in people, there is a wide variability in voluntary running behaviour in rats178. In our laboratory, we found that some rats liked to use their running wheel for a quiet nap.

Figure 3. Attentional and emotional factors modulate pain perception via different pathways.

a | Manipulating the attentional state primarily alters the perceived intensity of the pain sensation without significantly altering the perceived unpleasantness of the pain (top graph). By contrast, altering the mood state alters the perceived unpleasantness of the pain without altering the intensity of the sensation (bottom graph). b | Attention and emotion alter pain via different descending modulatory systems. Emotions (and placebo analgesia) activate circuitry involving the anterior cingulate cortex (ACC), prefrontal cortex (PFC) and periaqueductal grey (PAG) (shown in green), whereas attention activates circuitry involving projections from the superior parietal lobe (SPL) to the primary somatosensory cortex (S1) and insula (shown in blue). Grey regions show parts of the ascending pain pathways depicted in FIG. 2. AMY, amygdala; BG, basal ganglia; PB, parabrachial nucleus; RVM, rostroventral medulla; S2, secondary somatosensory cortex. * indicates p < 0.05, and ** indicates p < 0.01. Data used in part a are from REF. 4.
Human brain imaging studies examining the effects of distraction on pain processing find that when a person focuses on pain, the pain-evoked activity in several cortical areas, including the S1, insula and ACC, is stronger than when a person is distracted from the pain4,63–70. However, some of these studies used distracting stimuli that also alter arousal or emotional state64–68, so that the modulatory effects could be due to either attentional or emotional factors. The studies that varied the direction of attention while controlling for emotional state found that pain-evoked activity was modulated by attentional direction only in the insula and S1 (REFS 4,63,69,70), which is consistent with the role of these regions in pain sensation.
Neuroimaging studies evaluating the effects of emotional states on pain processing also find that negative emotional states produced by looking at emotional faces, listening to unpleasant music or smelling unpleasant odours alter pain-evoked cortical activation in a number of brain regions but most consistently in the ACC4,69,71–73. The modulation of activity in the ACC is in agreement with evidence that this region is particularly important for pain unpleasantness.
Different modulatory systems are involved in the attentional and emotional control of pain
The finding that attention and emotion have dissimilar effects on pain perception and alter pain-evoked activation of different cortical regions suggests that separate modulatory systems may underlie attentional and emotional influences on pain. The most commonly studied pain modulatory pathways involve projections from the midbrain PAG to brainstem nuclei, including the rostroventral medulla (RVM) and the locus coeruleus, to the dorsal horn of the spinal cord (for reviews, see REFS 74,75). These pathways involve endogenous opioids, noradrenaline and serotonin, and have both inhibitory and excitatory actions on spinal cord afferent projection neurons. Outputs from forebrain regions, including the ACC, PFC and amygdala reach the PAG, suggesting that these descending systems could be activated by psychological factors. Opioids are particularly powerful analgesics, and their analgesic action works at multiple sites within these descending pathways, including the ACC, PAG, RVM and dorsal horn of the spinal cord39,75.
Several studies that involved distraction from pain but used distraction procedures that probably increased arousal and/or negative emotions showed activation of this fronto–PAG–brainstem circuit67,76–78. However, when emotion and arousal were dissociated from attention4, it was the increased negative emotion that was associated with activation of the ACC–fronto–PAG circuitry. Furthermore, this activation correlated with negative mood-related increases in pain-evoked activity in the ACC (FIG. 3). By contrast, when subjects intentionally directed their attention to or away from a painful stimulus, attention-related alterations in pain-evoked activity in the insula correlated with activity in the superior parietal cortex (Brodmann area 7 (BA7)). This region is part of Corbetta and Shulman’s proposed ‘top-down orienting of attention’ system, which for visual attention involves the frontal eye fields and the superior parietal cortex79. Parts of BA7 project to the S1, S2 and insula80–82, providing a direct corticocortical pathway for the attentional modulation of pain, and descending pathways from the insula to the amygdala provide a possible descending component to the attentional modulation of pain14 (FIG. 3). Note that in addition to a top-down attention system, there is a bottom-up, stimulus-driven attentional circuit that is specialized in the detection of salient or unexpected stimuli79, involving the temporoparietal cortex and inferior frontal cortex (which is largely lateralized to the right hemisphere). This circuit could be differentially engaged when pain is presented in different emotional contexts.
Anticipation of pain relief activates descending pathways
Anticipation of the relief of pain is a primary contributor to placebo analgesia2. Brain imaging studies of expectation-related placebo analgesia find that immediately preceding the presentation of a noxious stimulus, when an individual receives a placebo ‘analgesic’ and is expecting a reduction in pain, there is activation of a ACC–fronto–PAG pathway83,84. Interestingly, this involves similar brain regions as those that are activated when a positive emotional state reduces pain. Moreover, placebo treatment alters endogenous opioid activity in parts of this pathway, including the PAG, ACC and PFC85. Further evidence that this ACC–fronto–PAG pathway is involved in placebo analgesia comes from findings showing that naloxone (a μ-opioid antagonist) modulates placebo-induced responses in the PAG and RVM and abolishes the coupling between rostral ACC and PAG activity83. There is also behavioural evidence that naloxone reduces expectation-related placebo analgesia in humans86 and conditioned placebo-analgesia in rodents87,88. Thus, it appears that descending modulatory pathways involved in placebo analgesia may overlap with those involved in the emotional modulation of pain. Conversely, circuitry involved in the attentional modulation of pain, including the superior parietal cortex and the insula, does not appear to be activated during expectation-related placebo analgesia. This independence of placebo analgesia from the attentional modulation of pain was further confirmed in a study showing additive reductions in pain caused by placebo and distraction89, indicating that placebo analgesia does not depend on the active redirection of attention. The finding that emotional factors, anticipation and attention alter pain processing is important in the therapeutic context. A patient receiving pharmacotherapy may have an enhanced or diminished response depending on his expectations, emotional state or focus of attention.
Effects of chronic pain on pain modulation
Patients with chronic pain often report that stimuli that should be innocuous are in fact painful for them. This can range from normal joint movement causing pain in patients with arthritis, to the touch of clothes against the skin causing burning pain in patients with nerve injuries. In laboratory studies, patients with various chronic pain syndromes, including arthritis, back pain, fibromyalgia, irritable bowel syndrome (IBS) and vulvar vestibulitis, show higher pain ratings and enhanced pain-evoked neural responses when experimental pain stimuli are presented than controls90–95. Even stimuli that are not painful in healthy people but are perceived to be painful by patients produce a pain-related activation pattern in the brain91,92,94.
The increased activation of pain pathways may arise from peripheral and/or central sensitization in ascending nociceptive pathways, but there is evidence that some of the amplification may arise from abnormalities in descending modulatory systems75,96. Perceptual evidence for abnormalities in descending pain modulation in patients with chronic pain comes from studies of conditioned pain modulation. In these paradigms, a painful test stimulus is evaluated in the absence and presence of a second painful (conditioning) stimulus applied to a remote body region. In a normally functioning nociceptive system, the amount of pain experienced at the primary testing site will be reduced when the second noxious stimulus is presented (classically termed counter-irritation or ‘pain inhibits pain’). Studies in anaesthetized rodents show a spino–brainstem–spinal modulatory loop that reduces afferent nociceptive activation, which is termed diffuse noxious inhibitory control97. However, in conscious humans, supraspinal mechanisms also contribute to conditioned pain modulation98. More than 30 studies have now examined conditioned pain modulation in patients with chronic pain, with varying paradigms and mixed results. Nevertheless, a recent meta-analysis concluded that conditioned pain modulation is impaired in populations with chronic pain99. This impairment in a system that normally reduces pain could contribute to the enhanced pain perception observed in patients with chronic pain. Functional brain imaging studies in patients with chronic pain also support the idea that endogenous pain modulatory systems may be dysfunctional in these patients. In response to experimental pain stimuli, patients with chronic pain disorders, including fibromyalgia, IBS and back pain, show abnormal activation patterns in brain regions involved in pain regulation, including the rostral ACC and frontal cortex100–103.
Structural changes in pain modulatory systems in chronic pain
There is now a wealth of evidence suggesting that patients with chronic pain may have anatomical alterations within regions involved in cognitive and emotional modulation of pain, such as the dorsolateral and medial PFC, the ACC and the insula. For example, there is less grey matter in the brains of patients with chronic back pain than in the brains of age-matched control subjects, especially in the dorsolateral PFC104–106. Grey matter loss has also been reported in patients suffering from other chronic pain disorders, such as fibromyalgia, headache, IBS, complex regional pain syndrome (CRPS) and osteoarthritis (for a review, see REF. 107). Although grey matter reductions are found in multiple brain regions, the most common regions to manifest such reductions are the insula, ACC and PFC. In addition to grey matter changes, alterations in white matter are beginning to be revealed using diffusion-weighted imaging techniques such as diffusion tensor imaging, in which fractional anisotropy — a measure of directionality of water movement — is measured as a surrogate for white matter integrity. These studies suggest that various types of chronic pain can lead to disruptions in white matter tracts108–114. Although conventional MRI cannot determine the histopathology underlying grey and white matter changes, several lines of evidence suggest the possibility that excessive nociceptive input may impair grey matter structure and function, including possible neuronal loss related to excitotoxicity115. Studies in rats and mice suggest that a chronic pain state can cause a supraspinal neuroinflammatory response116,117 in addition to changes in dendritic and synaptic structure and function in regions involved in pain processing118,119. Together with the anatomical data, this suggests that patients with chronic pain may undergo degenerative processes or at least functional alterations within brain areas involved in cognitively modulated analgesia.
The neurochemical basis of cognitive and emotional modulation of pain
Studies are now beginning to show that patients with chronic pain may have altered neurochemistry in brain systems that are involved in psychologically modulated analgesia. Findings from studies using in vivo proton magnetic resonance spectrometry show increases in glutamate and/or decreases of the neuronal marker N-acetyl aspartate in frontal cortices of patients with chronic back pain and fibromyalgia120–124; such findings support the idea that reduced grey matter in patients with chronic pain may be related to possible excitotoxicity. One study using transcranial magnetic stimulation found lower intracortical facilitation and shorter intracortical inhibition in patients with fibromyalgia than in controls125, suggesting that there are deficits in intracortical modulation involving both GABAergic and glutamatergic mechanisms. Other studies showed changes in opioidergic and dopaminergic systems in patients with chronic pain and rodent models of chronic pain. Several molecular imaging studies using positron emission tomography showed cerebral decreases in opioid receptor binding in patients with neuropathic pain, rheumatoid arthritis and fibromyalgia126–129. Decreases in the levels of dopamine in the forebrain of patients with fibromyalgia have also been reported130. Opioidergic dysfunction was also seen in the cortex and amygdala of mice with chronic inflammation or nerve injury, which was related to increases in anxiety-like behaviours131,132. Taken together, these studies suggest that changes in neurotransmitter systems may mean that patients with chronic pain have decreased receptor availability or heightened endogenous release of these neurotransmitters. In either case it seems that neurochemicals that are important for placebo analgesia are not acting in the same way as they do in healthy people.
Together, the evidence of altered neuroanatomical and neurochemical functioning in brain regions involved in the cognitive modulation of pain leads to the prediction that patients with chronic pain should have reduced or altered cognitive modulation of pain compared with healthy individuals (FIG. 4).
Figure 4. Consistently identified changes in the brains of patients with chronic pain.

The three cortical regions that consistently show decreases in grey matter are the anterior cingulate cortex (ACC), prefrontal cortex (PFC) and insula. Studies have also identified changes in white matter integrity in these regions; such changes are manifested by decreased fractional anisotropy (FA), which suggests that there is a decrease in white matter health. Molecular imaging studies show decreases in opioid receptor binding in patients with chronic pain in all three regions. Studies using in vivo proton magnetic resonance spectrometry show chronic pain-related decreases of the neuronal marker N-acetyl aspartate (NAA) in the frontal cortex and the insula. Finally, rodent studies show increased neuroinflammation in the ACC and PFC. Black arrows show the descending pathways from FIG. 3. Grey arrows show afferent pain pathways.
Is there evidence that disruption of endogenous pain modulatory systems alters cognitive and emotional modulation of pain in patients with chronic pain?
There is now accumulating evidence that the impairments observed in patients with chronic pain go beyond the pain itself. These patients also show changes across cognitive and emotional domains, and similar changes are found in preclinical models of chronic pain133 (BOX 3). For example, patients with fibromyalgia are less able to retain new information when rehearsal is prevented by a distraction than healthy controls134–136. Patients with back pain, fibromyalgia or CRPS have all been shown to have deficits in emotional decision-making when they were tested with the Iowa gambling task137–139. Preclinical studies confirm deficits in emotional decision-making tasks in rodent chronic pain models140. Deficits in these tasks are also observed in patients with disorders such as traumatic brain injury and multiple sclerosis who have lesions in brain regions implicated in descending pain modulation, including the medial PFC and rostral ACC141–145. Furthermore, the brain regions that are altered in patients with chronic pain are among those that are activated during the tasks in which patients perform poorly146,147. Thus, the anatomical changes observed in patients with chronic pain may well be an underlying factor in their altered performance in cognitive tasks.
Box 3. Possible mechanisms for human pain-related brain changes.
Rodents may not be able to tell us how they are feeling, but they do behave in ways that can inform us of their mental states, with the added advantage of giving us insight into the neurobiological changes that are induced by pain in a way that is just not possible in humans.
Similar to findings in humans, behavioural studies in rats have confirmed that acute pain interrupts attention179 and that distracting attention from that pain can reduce pain behaviours180. These interruptions in cognition can be traced to the circuitry involved — for example, arthritis pain in rats impaired decision-making in a ‘rat gambling task’ owing to feedforward inhibition of the medial prefrontal cortex from the basolateral amygdala181.
Cognitive deficits can also be seen in animal models of chronic pain. Inducing an inflammatory or neuropathic state in a rat can cause problems with attention168, decision-making182 and learning and memory183. Importantly, the neural bases of these can be investigated. Grey matter decreases can be shown via MRI167, and the cellular and molecular changes can be documented. For example, neuropathy has been shown to increase basal dendritic branching and spine density in medial prefrontal cortical areas, and the degree of hypersensitivity is related to the NMDA/AMPA ratio119. Other studies have shown increased neuroinflammation in frontal cortices after injury116,117 as well as shifts in opioidergic function131,132. Together, these studies suggest potential mechanisms to explain the changes in brain function seen in patients with chronic pain and the alterations seen in pain-related brain regions.
If the chronic pain-related anatomical alterations in brain regions involved in cognition, emotions and pain modulation truly have a functional consequence, then one would predict that patients with chronic pain should have altered psychological modulation of pain, including possible alterations in placebo responsiveness. Alternatively, the patients could have normal psychological modulation of pain, but they may be using different brain circuitry to achieve the modulation. Few studies have tested this concept, but there is some evidence pointing to both decreased psychological modulation of pain and the involvement of novel pathways in such modulation. For example, in response to relaxation, patients with IBS demonstrate reduced modulation of rectal distension-induced pain and reduced activation of the insula148. Conversely, these patients demonstrate more pronounced stress-induced modulation of neural activation in multiple brain sites involved in pain modulation, including the insula and ventrolateral PFC148. Whereas some studies find similar attentional149 and affective150 modulation of pain between patients with fibromyalgia and healthy controls, other studies have observed deficits in attentional pain modulation in patients with chronic pain151 and abnormal affective modulation of somatosensory brain processing in patients with fibromyalgia152. In terms of placebo analgesia, a study conducted in patients with IBS (a type of chronic visceral pain) found a placebo-related perceptual pain reduction but observed, contrary to findings in healthy subjects, that the opioid receptor antagonist naloxone did not reduce the placebo response, suggesting that any placebo response shown in patients with IBS was not opioid-mediated153. Although accumulating evidence suggests that patients with chronic pain may have altered psychological modulation of pain and altered modulatory systems in the brain, more direct comparisons between patients with chronic pain and healthy subjects need to be made to fully understand how these modulatory systems may be affected by the continued presence of pain.
Can altered pain modulatory circuitry be reversed in patients with chronic pain?
Several studies reveal that when a painful condition, such as hip osteoarthritis, headache or back pain, is eliminated after an individual has suffered chronic pain for years, grey matter reductions can be reversed. In these studies, brain regions implicated in pain modulation, including the dorsolateral PFC and ACC, had reduced grey matter in patients, but after successful treatment for the pain, the grey matter reductions were reversed so that the affected brain regions normalized in size104,154–156. These findings suggest that grey matter decreases related to chronic pain are not necessarily caused by neuronal death, but instead by other changes in neuronal tissue, such as reduced dendritic or synaptic density and possible changes in non-neuronal tissue119,157,158. Nevertheless, the anatomical normalization of brain regions involved in the psychological modulation of pain corresponds with chronic pain reductions after surgical or pharmacological treatment.
If cognitive and emotional factors can intrinsically activate modulatory circuitry in brain regions showing anatomical changes due to chronic pain, can psychological treatments reverse brain changes associated with chronic pain? Although there is little research available to date, some evidence suggests that answer may be ‘yes’. Cognitive behavioural therapy has been shown to increase pain-evoked neural activation in the dorsolateral and ventrolateral PFC: that is, in regions involved in pain modulation159. Meditation, alternatively, reduces pain-evoked PFC activity160,161 but increases activity in the rostral ACC and the anterior insula: that is, in areas involved in the cognitive regulation of nociceptive processing162. Although there is no direct evidence that activation of the pain regulatory circuitry has long-term functional consequences, there are a few studies showing that people who meditate have thicker cortices in frontal regions, including the PFC, ACC and insula8,163–165. Studies have not yet addressed the impact of such mind–body therapies on the brains of patients with chronic pain, but current evidence suggests that they may have a neuroprotective effect.
Conclusions, implications and limitations
There are huge inter-individual as well as intra-individual differences in pain perception depending on the context and meaning of the pain. Evidence presented in this Review shows that much of this variation can be explained by the interplay between afferent nociceptive signals to the brain and descending modulatory systems that are activated endogenously by cognitive and emotional factors. The descending modulatory systems involve brain regions that are important not only for pain but also for cognitive and emotional functioning in general166. Thus, alterations in the anatomical integrity and functioning of brain regions involved in both pain control and cognitive and/or emotional functioning could explain why patients with long-term chronic pain develop cognitive deficits as well as anxiety disorders and depression. Whereas the temporal relationship between pain, cognitive deficits, anxiety and depression is difficult to determine in most patients with pain, longitudinal animal studies now suggest that emotional and cognitive alterations may sometimes begin long after the onset of pain. For example, in a rodent model of neuropathic pain, rats exhibited anxiety-like behaviour and attentional deficits months after the injury and the onset of hypersensitivity, which was temporally coincident with anatomical changes in the frontal cortex167,168. Thus, there is growing evidence that pain may be detrimental to the brain and that long-term pain itself can both decrease an individual’s ability to endogenously control the pain and lead to many of the comorbidities that plague individuals with chronic pain. The old adage ‘no pain, no gain’ should probably be discarded in favour of ‘no pain, healthy brain’.
Nevertheless, caution is required when interpreting the results of the studies cited in this Review, as many of them involve small sample sizes and do not always include comprehensive control measures. Furthermore, most of the brain imaging studies are cross-sectional, so causal relationships are difficult to ascertain. Thus, longitudinal studies involving larger groups, as well as more preclinical studies back-translating clinical observations, will be important to fully confirm the hypothesis that chronic pain can alter pain modulatory systems in the brain.
Acknowledgments
Preparation of this manuscript was supported by the Intramural Research Program of the US National Institutes of Health, National Center for Complementary and Alternative Medicine.
Glossary
- Descending pain modulatory systems
Networks in the brain involving pathways from the cerebral cortex down to the spinal cord that can lead to inhibition or excitation of afferent pain signals at multiple levels of the brain
- Fibromyalgia
A disorder in which there is widespread pain in all four quadrants of the body for a minimum duration of 3 months. Additionally, at least 11 of 18 specified points on the neck, shoulder, chest, hip, knee and elbow regions show tenderness to pressure
- Vulvar vestibulitis
A disorder characterized by sensitivity around the vaginal orifice, with pain provoked by contact or pressure
- Ascending nociceptive pathways
Fibres travelling to the brain from receptors in body tissues that respond to tissue-damaging or potentially tissue-damaging stimuli (nociceptors). They make synapses with second-order neurons in the dorsal horn of the spinal cord, which send projections to the brainstem, thalamus or other brain regions. From there, third- and fourth-order neurons send projections to the cerebral cortex
- Complex regional pain syndrome
(CRPS). A chronic pain condition that can affect any part of the body but most frequently affects an arm or a leg. After what is often a minor injury, such as a sprained ankle, there is an intense burning pain that is much stronger than would be expected for the type of injury. The pain gets worse rather than better with time and is often accompanied by trophic changes, such as altered skin temperature and texture, faster growth of nails and hair and even loss of bone density
- Iowa gambling task
A psychological task used to investigate emotional decision-making. It involves playing with four card decks in order to win money. Playing with two of the decks leads to more wins than losses, whereas playing with the other decks leads to more losses than wins. Healthy people quickly gravitate to the ‘good’ decks. Patients with various types of frontal lobe lesions do not learn to preferentially use the ‘good’ decks
Footnotes
Competing interests statement
The authors declare no competing financial interests.
FURTHER INFORMATION
Division of Intramural Research Program at NCCAM: http://nccam.nih.gov/research/intramural
ALL LINKS ARE ACTIVE IN THE ONLINE PDF
References
- 1.Bingel U, et al. The effect of treatment expectation on drug efficacy: imaging the analgesic benefit of the opioid remifentanil. Sci Transl Med. 2011;3:70ra14. doi: 10.1126/scitranslmed.3001244. [DOI] [PubMed] [Google Scholar]
- 2.Benedetti F, Mayberg HS, Wager TD, Stohler CS, Zubieta JK. Neurobiological mechanisms of the placebo effect. J Neurosci. 2005;25:10390–10402. doi: 10.1523/JNEUROSCI.3458-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Villemure C, Bushnell MC. Cognitive modulation of pain: how do attention and emotion influence pain processing? Pain. 2002;95:195–199. doi: 10.1016/S0304-3959(02)00007-6. [DOI] [PubMed] [Google Scholar]
- 4.Villemure C, Bushnell MC. Mood influences supraspinal pain processing separately from attention. J Neurosci. 2009;29:705–715. doi: 10.1523/JNEUROSCI.3822-08.2009. This is the first study to dissociate the circuitry involved in emotional and attentional modulation of pain. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Loggia ML, Mogil JS, Bushnell MC. Empathy hurts: compassion for another increases both sensory and affective components of pain perception. Pain. 2008;136:168–176. doi: 10.1016/j.pain.2007.07.017. [DOI] [PubMed] [Google Scholar]
- 6.Schweinhardt P, Bushnell MC. Pain imaging in health and disease — how far have we come? J Clin Invest. 2010;120:3788–3797. doi: 10.1172/JCI43498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hölzel BK, et al. Mindfulness practice leads to increases in regional brain gray matter density. Psychiatry Res. 2011;191:36–43. doi: 10.1016/j.pscychresns.2010.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Grant JA, Courtemanche J, Duerden EG, Duncan GH, Rainville P. Cortical thickness and pain sensitivity in zen meditators. Emotion. 2010;10:43–53. doi: 10.1037/a0018334. [DOI] [PubMed] [Google Scholar]
- 9.Pagnoni G, Cekic M. Age effects on gray matter volume and attentional performance in Zen meditation. Neurobiol Aging. 2007;28:1623–1627. doi: 10.1016/j.neurobiolaging.2007.06.008. [DOI] [PubMed] [Google Scholar]
- 10.Mackey AP, Whitaker KJ, Bunge SA. Experience-dependent plasticity in white matter microstructure: reasoning training alters structural connectivity. Front Neuroanat. 2012;6:32. doi: 10.3389/fnana.2012.00032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Luders E, et al. Bridging the hemispheres in meditation: thicker callosal regions and enhanced fractional anisotropy (FA) in long-term practitioners. Neuroimage. 2012;61:181–187. doi: 10.1016/j.neuroimage.2012.02.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lutz A, McFarlin DR, Perlman DM, Salomons TV, Davidson RJ. Altered anterior insula activation during anticipation and experience of painful stimuli in expert meditators. Neuroimage. 2013;64:538–546. doi: 10.1016/j.neuroimage.2012.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Apkarian AV, Bushnell MC, Treede RD, Zubieta JK. Human brain mechanisms of pain perception and regulation in health and disease. Eur J Pain. 2005;9:463–484. doi: 10.1016/j.ejpain.2004.11.001. [DOI] [PubMed] [Google Scholar]
- 14.Friedman DP, Murray EA, O’Neill JB, Mishkin M. Cortical connections of the somatosensory fields of the lateral sulcus of macaques: evidence for a corticolimbic pathway for touch. J Comp Neurol. 1986;252:323–347. doi: 10.1002/cne.902520304. [DOI] [PubMed] [Google Scholar]
- 15.Rausell E, Jones EG. Histochemical and immunocytochemical compartments of the thalamic VPM nucleus in monkeys and their relationship to the representational map. J Neurosci. 1991;11:210–225. doi: 10.1523/JNEUROSCI.11-01-00210.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Apkarian AV, Shi T. In: Pain Mechanisms and Management. Ayrapetyan SN, Apkarian AV, editors. IOS Press; 1998. pp. 212–220. [Google Scholar]
- 17.Craig AD, Dostrovsky JO. Thermoreceptive lamina I trigeminothalamic neurons project to the nucleus submedius in the cat. Exp Brain Res. 1991;85:470–474. doi: 10.1007/BF00229424. [DOI] [PubMed] [Google Scholar]
- 18.Dum RP, Levinthal DJ, Strick PL. The spinothalamic system targets motor and sensory areas in the cerebral cortex of monkeys. J Neurosci. 2009;29:14223–14235. doi: 10.1523/JNEUROSCI.3398-09.2009. This is the first paper to show all of the cortical targets of the spinothalamic system. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Saab CY, Willis WD. The cerebellum: organization, functions and its role in nociception. Brain Res Brain Res Rev. 2003;42:85–95. doi: 10.1016/s0165-0173(03)00151-6. [DOI] [PubMed] [Google Scholar]
- 20.Monconduit L, Villanueva L. The lateral ventromedial thalamic nucleus spreads nociceptive signals from the whole body surface to layer I of the frontal cortex. Eur J Neurosci. 2005;21:3395–3402. doi: 10.1111/j.1460-9568.2005.04160.x. [DOI] [PubMed] [Google Scholar]
- 21.Becerra L, Breiter HC, Wise R, Gonzalez RG, Borsook D. Reward circuitry activation by noxious thermal stimuli. Neuron. 2001;32:927–946. doi: 10.1016/s0896-6273(01)00533-5. [DOI] [PubMed] [Google Scholar]
- 22.Baliki MN, Geha PY, Fields HL, Apkarian AV. Predicting value of pain and analgesia: nucleus accumbens response to noxious stimuli changes in the presence of chronic pain. Neuron. 2010;66:149–160. doi: 10.1016/j.neuron.2010.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bernard JF, Bester H, Besson JM. Involvement of the spino-parabrachio -amygdaloid and -hypothalamic pathways in the autonomic and affective emotional aspects of pain. Prog Brain Res. 1996;107:243–255. doi: 10.1016/s0079-6123(08)61868-3. [DOI] [PubMed] [Google Scholar]
- 24.Dunckley P, et al. A comparison of visceral and somatic pain processing in the human brainstem using functional magnetic resonance imaging. J Neurosci. 2005;25:7333–7341. doi: 10.1523/JNEUROSCI.1100-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Basbaum AI, Fields HL. Endogenous pain control systems: brainstem spinal pathways and endorphin circuitry. Annu Rev Neurosci. 1984;7:309–338. doi: 10.1146/annurev.ne.07.030184.001521. [DOI] [PubMed] [Google Scholar]
- 26.Kenshalo DR, Jr, Isensee O. Responses of primate SI cortical neurons to noxious stimuli. J Neurophysiol. 1983;50:1479–1496. doi: 10.1152/jn.1983.50.6.1479. [DOI] [PubMed] [Google Scholar]
- 27.Kenshalo DR, Jr, Chudler EH, Anton F, Dubner R. SI nociceptive neurons participate in the encoding process by which monkeys perceive the intensity of noxious thermal stimulation. Brain Res. 1988;454:378–382. doi: 10.1016/0006-8993(88)90841-4. [DOI] [PubMed] [Google Scholar]
- 28.Chudler EH, Anton F, Dubner R, Kenshalo DR., Jr Responses of nociceptive SI neurons in monkeys and pain sensation in humans elicited by noxious thermal stimulation: effect of interstimulus interval. J Neurophysiol. 1990;63:559–569. doi: 10.1152/jn.1990.63.3.559. [DOI] [PubMed] [Google Scholar]
- 29.Ploner M, Freund HJ, Schnitzler A. Pain affect without pain sensation in a patient with a postcentral lesion. Pain. 1999;81:211–214. doi: 10.1016/s0304-3959(99)00012-3. [DOI] [PubMed] [Google Scholar]
- 30.Greenspan JD, Lee RR, Lenz FA. Pain sensitivity alterations as a function of lesion location in the parasylvian cortex. Pain. 1999;81:273–282. doi: 10.1016/S0304-3959(99)00021-4. [DOI] [PubMed] [Google Scholar]
- 31.Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain. 1937;60:389–443. [Google Scholar]
- 32.MacLean PD. Psychosomatic disease and the “visceral brain. Recent developments bearing on the Papez theory of emotion. Psychosom Med. 1949;11:338–353. doi: 10.1097/00006842-194911000-00003. [DOI] [PubMed] [Google Scholar]
- 33.Foltz EL, Lowell EW. Pain “relief” by frontal cingulumotomy. J Neurosurg. 1962;19:89–100. doi: 10.3171/jns.1962.19.2.0089. [DOI] [PubMed] [Google Scholar]
- 34.Foltz EL, White LE. The role or rostral cingulumotomy in “pain” relief. Int J Neurol. 1968;6:353–373. [PubMed] [Google Scholar]
- 35.Corkin S, Hebben N. Subjective estimates of chronic pain before and after psychosurgery or treatment in a pain unit. Pain. 1981;1:S150. [Google Scholar]
- 36.Berthier M, Starkstein S, Leiguarda R. Asymbolia for pain: a sensory-limbic disconnection syndrome. Ann Neurol. 1988;24:41–49. doi: 10.1002/ana.410240109. [DOI] [PubMed] [Google Scholar]
- 37.Rainville P, Duncan GH, Price DD, Carrier B, Bushnell MC. Pain affect encoded in human anterior cingulate but not somatosensory cortex. Science. 1997;277:968–971. doi: 10.1126/science.277.5328.968. This is the first study to demonstrate the separation of sensory and affective pain processing in the cerebral cortex. [DOI] [PubMed] [Google Scholar]
- 38.Tolle TR, et al. Region-specific encoding of sensory and affective components of pain in the human brain: a positron emission tomography correlation analysis. Ann Neurol. 1999;45:40–47. doi: 10.1002/1531-8249(199901)45:1<40::aid-art8>3.0.co;2-l. [DOI] [PubMed] [Google Scholar]
- 39.Zubieta JK, et al. Regional μ opioid receptor regulation of sensory and affective dimensions of pain. Science. 2001;293:311–315. doi: 10.1126/science.1060952. This study provides the first demonstration of the relevance of forebrain opioid receptors to pain modulation. [DOI] [PubMed] [Google Scholar]
- 40.Ostrowsky K, et al. Representation of pain and somatic sensation in the human insula: a study of responses to direct electrical cortical stimulation. Cereb Cortex. 2002;12:376–385. doi: 10.1093/cercor/12.4.376. [DOI] [PubMed] [Google Scholar]
- 41.Craig AD. Significance of the insula for the evolution of human awareness of feelings from the body. Ann NY Acad Sci. 2011;1225:72–82. doi: 10.1111/j.1749-6632.2011.05990.x. [DOI] [PubMed] [Google Scholar]
- 42.Baliki MN, Geha PY, Apkarian AV. Parsing pain perception between nociceptive representation and magnitude estimation. J Neurophysiol. 2009;101:875–887. doi: 10.1152/jn.91100.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lamm C, Decety J, Singer T. Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain. Neuroimage. 2011;54:2492–2502. doi: 10.1016/j.neuroimage.2010.10.014. [DOI] [PubMed] [Google Scholar]
- 44.Cheng Y, Chen C, Lin CP, Chou KH, Decety J. Love hurts: an fMRI study. Neuroimage. 2010;51:923–929. doi: 10.1016/j.neuroimage.2010.02.047. [DOI] [PubMed] [Google Scholar]
- 45.Langford DJ, et al. Social modulation of pain as evidence for empathy in mice. Science. 2006;312:1967–1970. doi: 10.1126/science.1128322. [DOI] [PubMed] [Google Scholar]
- 46.Rosen G, Willoch F, Bartenstein P, Berner N, Rosjo S. Neurophysiological processes underlying the phantom limb pain experience and the use of hypnosis in its clinical management: an intensive examination of two patients. Int J Clin Exp Hypn. 2001;49:38–55. doi: 10.1080/00207140108410378. [DOI] [PubMed] [Google Scholar]
- 47.Porro CA, et al. Does anticipation of pain affect cortical nociceptive systems? J Neurosci. 2002;22:3206–3214. doi: 10.1523/JNEUROSCI.22-08-03206.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jensen J, et al. Direct activation of the ventral striatum in anticipation of aversive stimuli. Neuron. 2003;40:1251–1257. doi: 10.1016/s0896-6273(03)00724-4. [DOI] [PubMed] [Google Scholar]
- 49.Hsieh JC, Stone-Elander S, Ingvar M. Anticipatory coping of pain expressed in the human anterior cingulate cortex: a positron emission tomography study. Neurosci Lett. 1999;262:61–64. doi: 10.1016/s0304-3940(99)00060-9. [DOI] [PubMed] [Google Scholar]
- 50.Ploghaus A, et al. Dissociating pain from its anticipation in the human brain. Science. 1999;284:1979–1981. doi: 10.1126/science.284.5422.1979. This is the first study to examine the effect of pain anticipation on pain processing. [DOI] [PubMed] [Google Scholar]
- 51.Sawamoto N, et al. Expectation of pain enhances responses to nonpainful somatosensory stimulation in the anterior cingulate cortex and parietal operculum/posterior insula: an event-related functional magnetic resonance imaging study. J Neurosci. 2000;20:7438–7445. doi: 10.1523/JNEUROSCI.20-19-07438.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lopez-Sola M, et al. Dynamic assessment of the right lateral frontal cortex response to painful stimulation. Neuroimage. 2010;50:1177–1187. doi: 10.1016/j.neuroimage.2010.01.031. [DOI] [PubMed] [Google Scholar]
- 53.Fairhurst M, Wiech K, Dunckley P, Tracey I. Anticipatory brainstem activity predicts neural processing of pain in humans. Pain. 2007;128:101–110. doi: 10.1016/j.pain.2006.09.001. [DOI] [PubMed] [Google Scholar]
- 54.Beecher HK. Pain in men wounded in battle. Ann Surg. 1946;123:96–105. [PMC free article] [PubMed] [Google Scholar]
- 55.Morley S. Efficacy and effectiveness of cognitive behaviour therapy for chronic pain: progress and some challenges. Pain. 2011;152:S99–S106. doi: 10.1016/j.pain.2010.10.042. [DOI] [PubMed] [Google Scholar]
- 56.Zeidan F, Grant JA, Brown CA, McHaffie JG, Coghill RC. Mindfulness meditation-related pain relief: evidence for unique brain mechanisms in the regulation of pain. Neurosci Lett. 2012;520:165–173. doi: 10.1016/j.neulet.2012.03.082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jensen KB, et al. The use of functional neuroimaging to evaluate psychological and other non-pharmacological treatments for clinical pain. Neurosci Lett. 2012;520:156–164. doi: 10.1016/j.neulet.2012.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Beydoun A, Morrow TJ, Shen JF, Casey KL. Variability of laser-evoked potentials: attention, arousal and lateralized differences. Electroencephalogr Clin Neurophysiol. 1993;88:173–181. doi: 10.1016/0168-5597(93)90002-7. [DOI] [PubMed] [Google Scholar]
- 59.Roy M, Peretz I, Rainville P. Emotional valence contributes to music-induced analgesia. Pain. 2008;134:140–147. doi: 10.1016/j.pain.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 60.Villemure C, Slotnick BM, Bushnell MC. Effects of odors on pain perception: deciphering the roles of emotion and attention. Pain. 2003;106:101–108. doi: 10.1016/s0304-3959(03)00297-5. [DOI] [PubMed] [Google Scholar]
- 61.Loggia ML, Mogil JS, Bushnell MC. Experimentally induced mood changes preferentially affect pain unpleasantness. J Pain. 2008;9:784–791. doi: 10.1016/j.jpain.2008.03.014. [DOI] [PubMed] [Google Scholar]
- 62.Roy M, Lebuis A, Peretz I, Rainville P. The modulation of pain by attention and emotion: a dissociation of perceptual and spinal nociceptive processes. Eur J Pain. 2011;15:641–610. doi: 10.1016/j.ejpain.2010.11.013. [DOI] [PubMed] [Google Scholar]
- 63.Bushnell MC, et al. Pain perception: is there a role for primary somatosensory cortex? Proc Natl Acad Sci USA. 1999;96:7705–7709. doi: 10.1073/pnas.96.14.7705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Longe SE, et al. Counter-stimulatory effects on pain perception and processing are significantly altered by attention: an fMRI study. Neuroreport. 2001;12:2021–2025. doi: 10.1097/00001756-200107030-00047. [DOI] [PubMed] [Google Scholar]
- 65.Bantick SJ, et al. Imaging how attention modulates pain in humans using functional MRI. Brain. 2002;125:310–319. doi: 10.1093/brain/awf022. [DOI] [PubMed] [Google Scholar]
- 66.Brooks JC, Nurmikko TJ, Bimson WE, Singh KD, Roberts N. fMRI of thermal pain: effects of stimulus laterality and attention. Neuroimage. 2002;15:293–301. doi: 10.1006/nimg.2001.0974. [DOI] [PubMed] [Google Scholar]
- 67.Valet M, et al. Distraction modulates connectivity of the cingulo–frontal cortex and the midbrain during pain — an fMRI analysis. Pain. 2004;109:399–408. doi: 10.1016/j.pain.2004.02.033. [DOI] [PubMed] [Google Scholar]
- 68.Wiech K, et al. Modulation of pain processing in hyperalgesia by cognitive demand. Neuroimage. 2005;27:59–69. doi: 10.1016/j.neuroimage.2005.03.044. [DOI] [PubMed] [Google Scholar]
- 69.Ploner M, Lee MC, Wiech K, Bingel U, Tracey I. Flexible cerebral connectivity patterns subserve contextual modulations of pain. Cereb Cortex. 2011;21:719–726. doi: 10.1093/cercor/bhq146. [DOI] [PubMed] [Google Scholar]
- 70.Dunckley P, et al. Attentional modulation of visceral and somatic pain. Neurogastroenterol Motil. 2007;19:569–577. doi: 10.1111/j.1365-2982.2007.00908.x. [DOI] [PubMed] [Google Scholar]
- 71.Phillips ML, et al. The effect of negative emotional context on neural and behavioural responses to oesophageal stimulation. Brain. 2003;126:669–684. doi: 10.1093/brain/awg065. [DOI] [PubMed] [Google Scholar]
- 72.Roy M, Piche M, Chen JI, Peretz I, Rainville P. Cerebral and spinal modulation of pain by emotions. Proc Natl Acad Sci USA. 2009;106:20900–20905. doi: 10.1073/pnas.0904706106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Berna C, et al. Induction of depressed mood disrupts emotion regulation neurocircuitry and enhances pain unpleasantness. Biol Psychiatry. 2010:1083–1090. doi: 10.1016/j.biopsych.2010.01.014. [DOI] [PubMed] [Google Scholar]
- 74.Basbaum AI, Fields HL. Endogenous pain control mechanisms: review and hypothesis. Ann Neurol. 1978;4:451–462. doi: 10.1002/ana.410040511. This article provides the first complete analysis of descending pain modulatory circuits. [DOI] [PubMed] [Google Scholar]
- 75.Ossipov MH, Dussor GO, Porreca F. Central modulation of pain. J Clin Invest. 2010;120:3779–3787. doi: 10.1172/JCI43766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Petrovic P, Petersson KM, Ghatan PH, Stone-Elander S, Ingvar M. Pain-related cerebral activation is altered by a distracting cognitive task. Pain. 2000;85:19–30. doi: 10.1016/s0304-3959(99)00232-8. [DOI] [PubMed] [Google Scholar]
- 77.Frankenstein UN, Richter W, McIntyre MC, Remy F. Distraction modulates anterior cingulate gyrus activations during the cold pressor test. Neuroimage. 2001;14:827–836. doi: 10.1006/nimg.2001.0883. [DOI] [PubMed] [Google Scholar]
- 78.Tracey I, et al. Imaging attentional modulation of pain in the periaqueductal gray in humans. J Neurosci. 2002;22:2748–2752. doi: 10.1523/JNEUROSCI.22-07-02748.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Corbetta M, Shulman GL. Control of goal-directed and stimulus-driven attention in the brain. Nature Rev Neurosci. 2002;3:201–215. doi: 10.1038/nrn755. [DOI] [PubMed] [Google Scholar]
- 80.Cavada C, Goldman-Rakic PS. Posterior parietal cortex in rhesus monkey: I. Parcellation of areas based on distinctive limbic and sensory corticocortical connections. J Comp Neurol. 1989;287:393–421. doi: 10.1002/cne.902870402. [DOI] [PubMed] [Google Scholar]
- 81.Cavada C, Goldman-Rakic PS. Posterior parietal cortex in rhesus monkey: II. Evidence for segregated corticocortical networks linking sensory and limbic areas with the frontal lobe. J Comp Neurol. 1989;287:422–445. doi: 10.1002/cne.902870403. [DOI] [PubMed] [Google Scholar]
- 82.Prevosto V, Graf W, Ugolini G. Proprioceptive pathways to posterior parietal areas MIP and LIPv from the dorsal column nuclei and the postcentral somatosensory cortex. Eur J Neurosci. 2011;33:444–460. doi: 10.1111/j.1460-9568.2010.07541.x. [DOI] [PubMed] [Google Scholar]
- 83.Eippert F, et al. Activation of the opioidergic descending pain control system underlies placebo analgesia. Neuron. 2009;63:533–543. doi: 10.1016/j.neuron.2009.07.014. [DOI] [PubMed] [Google Scholar]
- 84.Wager TD, et al. Placebo-induced changes in fMRI in the anticipation and experience of pain. Science. 2004;303:1162–1167. doi: 10.1126/science.1093065. This study identifies the neural circuitry underlying placebo analgesia. [DOI] [PubMed] [Google Scholar]
- 85.Wager TD, Scott DJ, Zubieta JK. Placebo effects on human μ-opioid activity during pain. Proc Natl Acad Sci USA. 2007;104:11056–11061. doi: 10.1073/pnas.0702413104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Amanzio M, Benedetti F. Neuropharmacological dissection of placebo analgesia: expectation-activated opioid systems versus conditioning-activated specific subsystems. J Neurosci. 1999;19:484–494. doi: 10.1523/JNEUROSCI.19-01-00484.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Zhang RR, Zhang WC, Wang JY, Guo JY. The opioid placebo analgesia is mediated exclusively through μ-opioid receptor in rat. Int J Neuropsychopharmacol. 2013;16:849–856. doi: 10.1017/S1461145712000673. [DOI] [PubMed] [Google Scholar]
- 88.Guo JY, Wang JY, Luo F. Dissection of placebo analgesia in mice: the conditions for activation of opioid and non-opioid systems. J Psychopharmacol. 2010;24:1561–1567. doi: 10.1177/0269881109104848. [DOI] [PubMed] [Google Scholar]
- 89.Buhle JT, Stevens BL, Friedman JJ, Wager TD. Distraction and placebo: two separate routes to pain control. Psychol Sci. 2012;23:246–253. doi: 10.1177/0956797611427919. [DOI] [PubMed] [Google Scholar]
- 90.Derbyshire SW, et al. Cerebral responses to noxious thermal stimulation in chronic low back pain patients and normal controls. Neuroimage. 2002;16:158–168. doi: 10.1006/nimg.2002.1066. [DOI] [PubMed] [Google Scholar]
- 91.Gracely RH, Petzke F, Wolf JM, Clauw DJ. Functional magnetic resonance imaging evidence of augmented pain processing in fibromyalgia. Arthritis Rheum. 2002;46:1333–1343. doi: 10.1002/art.10225. This paper demonstrates enhanced pain processing in patients with chronic pain. [DOI] [PubMed] [Google Scholar]
- 92.Lawal A, Kern M, Sidhu H, Hofmann C, Shaker R. Novel evidence for hypersensitivity of visceral sensory neural circuitry in irritable bowel syndrome patients. Gastroenterology. 2006;130:26–33. doi: 10.1053/j.gastro.2005.10.020. [DOI] [PubMed] [Google Scholar]
- 93.Naliboff BD, et al. Cerebral activation in patients with irritable bowel syndrome and control subjects during rectosigmoid stimulation. Psychosom Med. 2001;63:365–375. doi: 10.1097/00006842-200105000-00006. [DOI] [PubMed] [Google Scholar]
- 94.Pukall CF, et al. Neural correlates of painful genital touch in women with vulvar vestibulitis syndrome. Pain. 2005;115:118–127. doi: 10.1016/j.pain.2005.02.020. [DOI] [PubMed] [Google Scholar]
- 95.Gwilym SE, et al. Psychophysical and functional imaging evidence supporting the presence of central sensitization in a cohort of osteoarthritis patients. Arthritis Rheum. 2009;61:1226–1234. doi: 10.1002/art.24837. [DOI] [PubMed] [Google Scholar]
- 96.Porreca F, Ossipov MH, Gebhart GF. Chronic pain and medullary descending facilitation. Trends Neurosci. 2002;25:319–325. doi: 10.1016/s0166-2236(02)02157-4. [DOI] [PubMed] [Google Scholar]
- 97.Le Bars D. The whole body receptive field of dorsal horn multireceptive neurones. Brain Res Brain Res Rev. 2002;40:29–44. doi: 10.1016/s0165-0173(02)00186-8. [DOI] [PubMed] [Google Scholar]
- 98.Sprenger C, Bingel U, Buchel C. Treating pain with pain: supraspinal mechanisms of endogenous analgesia elicited by heterotopic noxious conditioning stimulation. Pain. 2011;152:428–439. doi: 10.1016/j.pain.2010.11.018. [DOI] [PubMed] [Google Scholar]
- 99.Lewis GN, Rice DA, McNair PJ. Conditioned pain modulation in populations with chronic pain: a systematic review and meta-analysis. J Pain. 2012;13:936–944. doi: 10.1016/j.jpain.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 100.Jensen KB, et al. Evidence of dysfunctional pain inhibition in Fibromyalgia reflected in rACC during provoked pain. Pain. 2009;144:95–100. doi: 10.1016/j.pain.2009.03.018. [DOI] [PubMed] [Google Scholar]
- 101.Burgmer M, et al. Fibromyalgia unique temporal brain activation during experimental pain: a controlled fMRI study. J Neural Transm. 2010;117:123–131. doi: 10.1007/s00702-009-0339-1. [DOI] [PubMed] [Google Scholar]
- 102.Berman SM, et al. Reduced brainstem inhibition during anticipated pelvic visceral pain correlates with enhanced brain response to the visceral stimulus in women with irritable bowel syndrome. J Neurosci. 2008;28:349–359. doi: 10.1523/JNEUROSCI.2500-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Baliki MN, et al. Chronic pain and the emotional brain: specific brain activity associated with spontaneous fluctuations of intensity of chronic back pain. J Neurosci. 2006;26:12165–12173. doi: 10.1523/JNEUROSCI.3576-06.2006. This study shows that chronic pain activates unique patterns of cortical activity. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Seminowicz DA, et al. Effective treatment of chronic low back pain in humans reverses abnormal brain anatomy and function. J Neurosci. 2011;31:7540–7550. doi: 10.1523/JNEUROSCI.5280-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Schmidt-Wilcke T, et al. Affective components and intensity of pain correlate with structural differences in gray matter in chronic back pain patients. Pain. 2006;125:89–97. doi: 10.1016/j.pain.2006.05.004. [DOI] [PubMed] [Google Scholar]
- 106.Apkarian AV, et al. Chronic back pain is associated with decreased prefrontal and thalamic gray matter density. J Neurosci. 2004;24:10410–10415. doi: 10.1523/JNEUROSCI.2541-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Davis KD, Moayedi M. Central mechanisms of pain revealed through functional and structural MRI. J Neuroimmune Pharmacol. 2012 Jul 24; doi: 10.1007/s11481-012-9386-8. [DOI] [PubMed] [Google Scholar]
- 108.Geha PY, et al. The brain in chronic CRPS pain: abnormal gray–white matter interactions in emotional and autonomic regions. Neuron. 2008;60:570–581. doi: 10.1016/j.neuron.2008.08.022. This study links chronic pain with both grey and white matter changes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Lutz J, et al. White and gray matter abnormalities in the brain of patients with fibromyalgia: a diffusion-tensor and volumetric imaging study. Arthritis Rheum. 2008;58:3960–3969. doi: 10.1002/art.24070. [DOI] [PubMed] [Google Scholar]
- 110.Sundgren PC, et al. Diffusion-weighted and diffusion tensor imaging in fibromyalgia patients: a prospective study of whole brain diffusivity, apparent diffusion coefficient, and fraction anisotropy in different regions of the brain and correlation with symptom severity. Acad Radiol. 2007;14:839–846. doi: 10.1016/j.acra.2007.03.015. [DOI] [PubMed] [Google Scholar]
- 111.Gerstner G, Ichesco E, Quintero A, Schmidt-Wilcke T. Changes in regional gray and white matter volume in patients with myofascial-type temporomandibular disorders: a voxel-based morphometry study. J Orofac Pain. 2011;25:99–106. [PubMed] [Google Scholar]
- 112.Granziera C, DaSilva AF, Snyder J, Tuch DS, Hadjikhani N. Anatomical alterations of the visual motion processing network in migraine with and without aura. PLoS Med. 2006;3:e402. doi: 10.1371/journal.pmed.0030402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Szabo N, et al. White matter microstructural alterations in migraine: a diffusion-weighted MRI study. Pain. 2012;153:651–656. doi: 10.1016/j.pain.2011.11.029. [DOI] [PubMed] [Google Scholar]
- 114.Moayedi M, et al. White matter brain and trigeminal nerve abnormalities in temporomandibular disorder. Pain. 2012;153:1467–1477. doi: 10.1016/j.pain.2012.04.003. [DOI] [PubMed] [Google Scholar]
- 115.McEwen BS. The neurobiology of stress: from serendipity to clinical relevance. Brain Res. 2000;886:172–189. doi: 10.1016/s0006-8993(00)02950-4. [DOI] [PubMed] [Google Scholar]
- 116.Apkarian AV, et al. Expression of IL-1β in supraspinal brain regions in rats with neuropathic pain. Neurosci Lett. 2006;407:176–181. doi: 10.1016/j.neulet.2006.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Norman GJ, et al. Stress and IL-1β contribute to the development of depressive-like behavior following peripheral nerve injury. Mol Psychiatry. 2010;15:404–414. doi: 10.1038/mp.2009.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Xu H, et al. Presynaptic and postsynaptic amplifications of neuropathic pain in the anterior cingulate cortex. J Neurosci. 2008;28:7445–7453. doi: 10.1523/JNEUROSCI.1812-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Metz AE, Yau HJ, Centeno MV, Apkarian AV, Martina M. Morphological and functional reorganization of rat medial prefrontal cortex in neuropathic pain. Proc Natl Acad Sci USA. 2009;106:2423–2428. doi: 10.1073/pnas.0809897106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Grachev ID, Fredrickson BE, Apkarian AV. Brain chemistry reflects dual states of pain and anxiety in chronic low back pain. J Neural Transm. 2002;109:1309–1334. doi: 10.1007/s00702-002-0722-7. [DOI] [PubMed] [Google Scholar]
- 121.Harris RE, et al. Dynamic levels of glutamate within the insula are associated with improvements in multiple pain domains in fibromyalgia. Arthritis Rheum. 2008;58:903–907. doi: 10.1002/art.23223. [DOI] [PubMed] [Google Scholar]
- 122.Grachev ID, Fredrickson BE, Apkarian AV. Abnormal brain chemistry in chronic back pain: an in vivo proton magnetic resonance spectroscopy study. Pain. 2000;89:7–18. doi: 10.1016/S0304-3959(00)00340-7. [DOI] [PubMed] [Google Scholar]
- 123.Harris RE, et al. Elevated insular glutamate in fibromyalgia is associated with experimental pain. Arthritis Rheum. 2009;60:3146–3152. doi: 10.1002/art.24849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Gussew A, Rzanny R, Gullmar D, Scholle HC, Reichenbach JR. 1H-MR spectroscopic detection of metabolic changes in pain processing brain regions in the presence of non-specific chronic low back pain. Neuroimage. 2011;54:1315–1323. doi: 10.1016/j.neuroimage.2010.09.039. [DOI] [PubMed] [Google Scholar]
- 125.Mhalla A, de Andrade DC, Baudic S, Perrot S, Bouhassira D. Alteration of cortical excitability in patients with fibromyalgia. Pain. 2010;149:495–500. doi: 10.1016/j.pain.2010.03.009. [DOI] [PubMed] [Google Scholar]
- 126.Harris RE, et al. Decreased central μ-opioid receptor availability in fibromyalgia. J Neurosci. 2007;27:10000–10006. doi: 10.1523/JNEUROSCI.2849-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Jones AKP, et al. Changes in central opioid receptor binding in relation to inflammation and pain in patients with rheumatoid arthritis. Br J Rheumatol. 1994;33:909–916. doi: 10.1093/rheumatology/33.10.909. [DOI] [PubMed] [Google Scholar]
- 128.Jones AK, Watabe H, Cunningham VJ, Jones T. Cerebral decreases in opioid receptor binding in patients with central neuropathic pain measured by [11C]diprenorphine binding and PET. Eur J Pain. 2004;8:479–485. doi: 10.1016/j.ejpain.2003.11.017. [DOI] [PubMed] [Google Scholar]
- 129.Maarrawi J, et al. Differential brain opioid receptor availability in central and peripheral neuropathic pain. Pain. 2007;127:183–194. doi: 10.1016/j.pain.2006.10.013. [DOI] [PubMed] [Google Scholar]
- 130.Wood PB, et al. Fibromyalgia patients show an abnormal dopamine response to pain. Eur J Neurosci. 2007;25:3576–3582. doi: 10.1111/j.1460-9568.2007.05623.x. [DOI] [PubMed] [Google Scholar]
- 131.Narita M, et al. Chronic pain induces anxiety with concomitant changes in opioidergic function in the amygdala. Neuropsychopharmacology. 2006;31:739–750. doi: 10.1038/sj.npp.1300858. [DOI] [PubMed] [Google Scholar]
- 132.Narita M, et al. Chronic pain-induced emotional dysfunction is associated with astrogliosis due to cortical δ-opioid receptor dysfunction. J Neurochem. 2006;97:1369–1378. doi: 10.1111/j.1471-4159.2006.03824.x. [DOI] [PubMed] [Google Scholar]
- 133.Moriarty O, McGuire BE, Finn DP. The effect of pain on cognitive function: a review of clinical and preclinical research. Prog Neurobiol. 2011;93:385–404. doi: 10.1016/j.pneurobio.2011.01.002. [DOI] [PubMed] [Google Scholar]
- 134.Leavitt F, Katz RS. Distraction as a key determinant of impaired memory in patients with fibromyalgia. J Rheumatol. 2006;33:127–132. [PubMed] [Google Scholar]
- 135.Dick BD, Verrier MJ, Harker KT, Rashiq S. Disruption of cognitive function in Fibromyalgia Syndrome. Pain. 2008;139:610–616. doi: 10.1016/j.pain.2008.06.017. [DOI] [PubMed] [Google Scholar]
- 136.Munguia-Izquierdo D, Legaz-Arrese A. Assessment of the effects of aquatic therapy on global symptomatology in patients with fibromyalgia syndrome: a randomized controlled trial. Arch Phys Med Rehabil. 2008;89:2250–2257. doi: 10.1016/j.apmr.2008.03.026. [DOI] [PubMed] [Google Scholar]
- 137.Verdejo-Garcia A, Lopez-Torrecillas F, Calandre EP, Delgado-Rodriguez A, Bechara A. Executive function and decision-making in women with fibromyalgia. Arch Clin Neuropsychol. 2009;24:113–122. doi: 10.1093/arclin/acp014. [DOI] [PubMed] [Google Scholar]
- 138.Walteros C, et al. Altered associative learning and emotional decision making in fibromyalgia. J Psychosom Res. 2011;70:294–301. doi: 10.1016/j.jpsychores.2010.07.013. [DOI] [PubMed] [Google Scholar]
- 139.Apkarian AV, et al. Chronic pain patients are impaired on an emotional decision-making task. Pain. 2004;108:129–136. doi: 10.1016/j.pain.2003.12.015. [DOI] [PubMed] [Google Scholar]
- 140.Pais-Vieira M, Mendes-Pinto MM, Lima D, Galhardo V. Cognitive impairment of prefrontal-dependent decision-making in rats after the onset of chronic pain. Neuroscience. 2009;161:671–679. doi: 10.1016/j.neuroscience.2009.04.011. [DOI] [PubMed] [Google Scholar]
- 141.Hattori N, et al. Differential SPECT activation patterns associated with PASAT performance may indicate frontocerebellar functional dissociation in chronic mild traumatic brain injury. J Nucl Med. 2009;50:1054–1061. doi: 10.2967/jnumed.108.060368. [DOI] [PubMed] [Google Scholar]
- 142.Yu HJ, et al. Multiple white matter tract abnormalities underlie cognitive impairment in RRMS. Neuroimage. 2012;59:3713–3722. doi: 10.1016/j.neuroimage.2011.10.053. [DOI] [PubMed] [Google Scholar]
- 143.Sigurdardottir S, Jerstad T, Andelic N, Roe C, Schanke AK. Olfactory dysfunction, gambling task performance and intracranial lesions after traumatic brain injury. Neuropsychology. 2010;24:504–513. doi: 10.1037/a0018934. [DOI] [PubMed] [Google Scholar]
- 144.van Noordt S, Good D. Mild head injury and sympathetic arousal: investigating relationships with decision-making and neuropsychological performance in university students. Brain Inj. 2011;25:707–716. doi: 10.3109/02699052.2011.580312. [DOI] [PubMed] [Google Scholar]
- 145.Roca M, et al. Cognitive deficits in multiple sclerosis correlate with changes in fronto-subcortical tracts. Mult Scler. 2008;14:364–369. doi: 10.1177/1352458507084270. [DOI] [PubMed] [Google Scholar]
- 146.Owen AM, McMillan KM, Laird AR, Bullmore E. N-back working memory paradigm: a meta-analysis of normative functional neuroimaging studies. Hum Brain Mapp. 2005;25:46–59. doi: 10.1002/hbm.20131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Bechara A, Damasio H, Damasio AR. Emotion, decision making and the orbitofrontal cortex. Cereb Cortex. 2000;10:295–307. doi: 10.1093/cercor/10.3.295. [DOI] [PubMed] [Google Scholar]
- 148.Elsenbruch S, et al. Patients with irritable bowel syndrome have altered emotional modulation of neural responses to visceral stimuli. Gastroenterology. 2010;139:1310–1319. doi: 10.1053/j.gastro.2010.06.054. [DOI] [PubMed] [Google Scholar]
- 149.Tiemann L, et al. Behavioral and neuronal investigations of hypervigilance in patients with fibromyalgia syndrome. PLoS ONE. 2012;7:e35068. doi: 10.1371/journal.pone.0035068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Arnold BS, et al. Affective pain modulation in fibromyalgia, somatoform pain disorder, back pain, and healthy controls. Eur J Pain. 2008;12:329–338. doi: 10.1016/j.ejpain.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 151.Snijders TJ, Ramsey NF, Koerselman F, van Gijn J. Attentional modulation fails to attenuate the subjective pain experience in chronic, unexplained pain. Eur J Pain. 2010;14:282.e1–282.e10. doi: 10.1016/j.ejpain.2009.05.019. [DOI] [PubMed] [Google Scholar]
- 152.Montoya P, Pauli P, Batra A, Wiedemann G. Altered processing of pain-related information in patients with fibromyalgia. Eur J Pain. 2005;9:293–303. doi: 10.1016/j.ejpain.2004.07.012. [DOI] [PubMed] [Google Scholar]
- 153.Vase L, Robinson ME, Verne GN, Price DD. Increased placebo analgesia over time in irritable bowel syndrome (IBS) patients is associated with desire and expectation but not endogenous opioid mechanisms. Pain. 2005;115:338–347. doi: 10.1016/j.pain.2005.03.014. [DOI] [PubMed] [Google Scholar]
- 154.Obermann M, et al. Gray matter changes related to chronic posttraumatic headache. Neurology. 2009;73:978–983. doi: 10.1212/WNL.0b013e3181b8791a. [DOI] [PubMed] [Google Scholar]
- 155.Rodriguez-Raecke R, Niemeier A, Ihle K, Ruether W, May A. Brain gray matter decrease in chronic pain is the consequence and not the cause of pain. J Neurosci. 2009;29:13746–13750. doi: 10.1523/JNEUROSCI.3687-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Gwilym SE, Filippini N, Douaud G, Carr AJ, Tracey I. Thalamic atrophy associated with painful osteoarthritis of the hip is reversible after arthroplasty: a longitudinal voxel-based morphometric study. Arthritis Rheum. 2010;62:2930–2940. doi: 10.1002/art.27585. [DOI] [PubMed] [Google Scholar]
- 157.Zhao MG, Toyoda H, Wang YK, Zhuo M. Enhanced synaptic long-term potentiation in the anterior cingulate cortex of adult wild mice as compared with that in laboratory mice. Mol Brain. 2009;2:11. doi: 10.1186/1756-6606-2-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Ikeda H, Tsuda M, Inoue K, Murase K. Long-term potentiation of neuronal excitation by neuron-glia interactions in the rat spinal dorsal horn. Eur J Neurosci. 2007;25:1297–1306. doi: 10.1111/j.1460-9568.2007.05386.x. [DOI] [PubMed] [Google Scholar]
- 159.Jensen KB, et al. Cognitive Behavioral Therapy increases pain-evoked activation of the prefrontal cortex in patients with fibromyalgia. Pain. 2012;153:1495–1503. doi: 10.1016/j.pain.2012.04.010. [DOI] [PubMed] [Google Scholar]
- 160.Grant JA, Courtemanche J, Rainville P. A non-elaborative mental stance and decoupling of executive and pain-related cortices predicts low pain sensitivity in Zen meditators. Pain. 2011;152:150–156. doi: 10.1016/j.pain.2010.10.006. [DOI] [PubMed] [Google Scholar]
- 161.Gard T, et al. Pain attenuation through mindfulness is associated with decreased cognitive control and increased sensory processing in the brain. Cereb Cortex. 2012;22:2692–2702. doi: 10.1093/cercor/bhr352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Zeidan F, et al. Brain mechanisms supporting the modulation of pain by mindfulness meditation. J Neurosci. 2011;31:5540–5548. doi: 10.1523/JNEUROSCI.5791-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Lazar SW, et al. Meditation experience is associated with increased cortical thickness. Neuroreport. 2005;16:1893–1897. doi: 10.1097/01.wnr.0000186598.66243.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Holzel BK, et al. Investigation of mindfulness meditation practitioners with voxel-based morphometry. Soc Cogn Affect Neurosci. 2008;3:55–61. doi: 10.1093/scan/nsm038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Luders E, Toga AW, Lepore N, Gaser C. The underlying anatomical correlates of long-term meditation: larger hippocampal and frontal volumes of gray matter. Neuroimage. 2009;45:672–678. doi: 10.1016/j.neuroimage.2008.12.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Pessoa L. On the relationship between emotion and cognition. Nature Rev Neurosci. 2008;9:148–158. doi: 10.1038/nrn2317. [DOI] [PubMed] [Google Scholar]
- 167.Seminowicz DA, et al. MRI structural brain changes associated with sensory and emotional function in a rat model of long-term neuropathic pain. Neuroimage. 2009;47:1007–1014. doi: 10.1016/j.neuroimage.2009.05.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Low LA, et al. Nerve injury causes long-term attentional deficits in rats. Neurosci Lett. 2012;529:103–107. doi: 10.1016/j.neulet.2012.09.027. [DOI] [PubMed] [Google Scholar]
- 169.deCharms RC, et al. Control over brain activation and pain learned by using real-time functional MRI. Proc Natl Acad Sci USA. 2005;102:18626–18631. doi: 10.1073/pnas.0505210102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Yoo SS, et al. Brain–computer interface using fMRI: spatial navigation by thoughts. Neuroreport. 2004;15:1591–1595. doi: 10.1097/01.wnr.0000133296.39160.fe. [DOI] [PubMed] [Google Scholar]
- 171.van Praag H, Kempermann G, Gage FH. Neural consequences of enviromental enrichment. Nature Rev Neurosci. 2000;1:191–198. doi: 10.1038/35044558. [DOI] [PubMed] [Google Scholar]
- 172.Gabriel AF, et al. Enriched environment and the recovery from inflammatory pain: social versus physical aspects and their interaction. Behav Brain Res. 2010;208:90–95. doi: 10.1016/j.bbr.2009.11.015. [DOI] [PubMed] [Google Scholar]
- 173.Gabriel AF, Marcus MAE, Honig WMM, Joosten EAJ. Preoperative housing in an enriched environment significantly reduces the duration of postoperative pain in a rat model of knee inflammation. Neurosci Lett. 2010;469:219–223. doi: 10.1016/j.neulet.2009.11.078. [DOI] [PubMed] [Google Scholar]
- 174.Gabriel AF, Marcus MA, Honig WM, Helgers N, Joosten EA. Environmental housing affects the duration of mechanical allodynia and the spinal astroglial activation in a rat model of chronic inflammatory pain. Brain Res. 2009;1276:83–90. doi: 10.1016/j.brainres.2009.04.039. [DOI] [PubMed] [Google Scholar]
- 175.Abramov U, Kurrikoff K, Matsui T, Vasar E. Environmental enrichment reduces mechanical hypersensitivity in neuropathic mice, but fails to abolish the phenotype of CCK2 receptor deficient mice. Neurosci Lett. 2009;467:230–233. doi: 10.1016/j.neulet.2009.10.043. [DOI] [PubMed] [Google Scholar]
- 176.Shum FW, et al. Alteration of cingulate long-term plasticity and behavioral sensitization to inflammation by environmental enrichment. Learn Mem. 2007;14:304–312. doi: 10.1101/lm.530607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Stagg NJ, et al. Regular exercise reverses sensory hypersensitivity in a rat neuropathic pain model: role of endogenous opioids. Anesthesiology. 2011;114:940–948. doi: 10.1097/ALN.0b013e318210f880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Bakos J, et al. Oxytocin levels in the posterior pituitary and in the heart are modified by voluntary wheel running. Regul Pept. 2007;139:96–101. doi: 10.1016/j.regpep.2006.10.011. [DOI] [PubMed] [Google Scholar]
- 179.Boyette-Davis JA, Thompson CD, Fuchs PN. Alterations in attentional mechanisms in response to acute inflammatory pain and morphine administration. Neuroscience. 2008;151:558–563. doi: 10.1016/j.neuroscience.2007.10.032. [DOI] [PubMed] [Google Scholar]
- 180.Ford GK, Moriarty O, McGuire BE, Finn DP. Investigating the effects of distracting stimuli on nociceptive behaviour and associated alterations in brain monoamines in rats. Eur J Pain. 2008;12:970–979. doi: 10.1016/j.ejpain.2008.01.002. [DOI] [PubMed] [Google Scholar]
- 181.Ji G, et al. Cognitive impairment in pain through amygdala-driven prefrontal cortical deactivation. J Neurosci. 2010;30:5451–5464. doi: 10.1523/JNEUROSCI.0225-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Leite-Almeida H, et al. The impact of age on emotional and cognitive behaviours triggered by experimental neuropathy in rats. Pain. 2009;144:57–65. doi: 10.1016/j.pain.2009.02.024. [DOI] [PubMed] [Google Scholar]
- 183.Hu Y, Yang J, Hu Y, Wang Y, Li W. Amitriptyline rather than lornoxicam ameliorates neuropathic pain-induced deficits in abilities of spatial learning and memory. Eur J Anaesthesiol. 2010;27:162–168. doi: 10.1097/EJA.0b013e328331a3d5. [DOI] [PubMed] [Google Scholar]