

Abstract
The Hyporheic Zone is among the most important interstitial freshwater habitats, but the relationship between biotic and abiotic factors in this zone remains under-explored. Enterobacteria were expected to be present, but no specific studies had ever confirmed this prediction. The aim of this study was, therefore, to evaluate the total coliforms, Escherichia coli and Salmonella spp. in hyporheic water and to determine the relationship of the physical, chemical and environmental factors at different depths in a rainforest stream. To this end, thirty-six water samples were collected at three depths in sites located in the first, second and third orders in diverse substrates. The total coliforms, Escherichia coli and Salmonella sp. were evaluated in terms of their CFU/ml. In the interstitial samples, coliforms were detected in 100% of the samples. The total coliform counts had higher values at intermediate depths, while E. coli and Salmonella spp. instead had higher values at intermediate and large depths, often reaching or exceeding the values of the surface samples. Our results revealed that Salmonella spp. and the coliforms have different microhabitat preferences. Salmonella spp. and coliform species prefer deposition areas, such as lateral sides of pools, curves and bars, but they have a tendency to distribute into different depths, likely due to temperature differences. Salmonella spp. prefer compact substrata, with fewer fluids passing through and with upwelling areas with lower oxygen inflow. The coliform species showed the opposite preference. Our results suggest that bacterial variation is related to environmental factors and physical-chemical parameters within the HZ and may play a key role in the microbial diversity and distribution in these ecosystems.
Introduction
Many microorganisms are known to be relevant contaminants of the surface water of aquatic environments across diverse geographic areas [1,2,3,4,5]. As a means of assessing the level of fecal pollution in environmental waters, the use of enteric indicators, such as fecal coliforms (FC) or Escherichia coli, is widely accepted [6]. The validity of using such organisms as indicators of water pollution depends on their fecal specificity and inability to multiply outside their primary host environment, the gastrointestinal tracts of humans and warm-blooded animals [7]. Coliforms, Escherichia coli and Salmonella spp. reside in the gastrointestinal tracts of humans and animals and are used as indicator organisms to assess the microbiological safety of drinking and recreational waters [6]. There is some evidence that standard fecal indicators may originate from non-enteric sources, may survive significantly longer in tropical waters than in temperate ones, and may even become part of the aquatic microbial community [8]. E. coli thrives in the intestinal tract of humans and other warm-blooded animals and can contaminate water sources via untreated wastewater release [9,10]. The majority of E. coli strains are commensal; however, some lineages may have acquired specific virulence attributes that allow them to cause a wide spectrum of clinical manifestations, including diarrhea, urinary tract infections, meningitis, and septicemia [9,10]. It has been suggested that a full half of the total E. coli population resides in the external environment [11]. In this natural habitat, they can survive approximately 1 day in water, 1.5 days in sediment, and 3 days in soil [12]. This implies, in general, that the E. coli populations found in the secondary habitats are maintained by the constant arrival of microorganisms from hosts [11,13]. Some authors have suggested that E. coli and enterococci may multiply in warm, subtropical waters [12,14,15,16], and high concentrations of E. coli have been found in tropical surface waters in the absence of known human fecal sources [17,18]. Salmonella spp. are essentially vertebrate parasites, and in addition to humans, their natural hosts include all mammalian species, birds, reptilians and amphibians [19,20,21,22]. Consequently, in addition to polluted superficial water and groundwater, they may be present in various types of natural waters, such as rivers, lakes and estuaries [23]. Compared to other bacteria, Salmonella has higher survival rates in aquatic environments, ensuring its passage to the next host [24].
Other studies have demonstrated the extended persistence of indicator bacteria in the sediments of environmental waters [25,26] and in bromeliad water [27]. Anderson et al. [13] used laboratory microcosm experiments to show that the persistence tended to be greater in sediments than in fresh water columns and that there are differential survival rates for certain E. coli and Enterobacter strains in the two habitats. According to Winfield and Groisman [12], Salmonella spp. can survive in a septic tank for 10 to 15 days, can adhere to the substrata of a river, and may survive in soil for up to one year.
The Hyporheic Zone (HZ) is one of the most important interstitial freshwater habitats. It is a sub-benthic habitat composed of the spaces between the riverbed particles, thus representing a natural interface between stream water and subterranean water [28,29]. The exchange of water, nutrients, and biota between surface water and groundwater in the hyporheic zone has been demonstrated in various past publications [30,31,32,33,34,35]. This unique environment allows biological and chemical microzones to occur, facilitating diverse microbiological processes in a small volume and affecting the bacterial population dynamics.
In this zone, the microbiota consists of bacteria, archaea, protozoa and fungi [31]. These form thin films, called biofilms, on the granules. The biofilms are perhaps the most important component of the interstitial environment [36] and are estimated to spread over surfaces of up to 1 m2 in 10 grams of sediment with particle diameter of 0.1 mm [37].
The HZ is becoming increasingly recognized for its role as a nutrient contributor, providing a storage area for allochthonous and autochthonous organic matter. Its concentrations of nitrates, phosphates, silicates, organic carbon and heavy metals are higher than on the surface, and it is a habitat in which bacteria process the dissolved organic carbon more efficiently [38].
From the ecological point of view, the HZ and its biological components are strongly and persistently influenced by anthropogenic disturbances such as the discharge of industrial or municipal waste [39] and mining and deforestation [40]. The temperature, pH, inorganic nutrients, dissolved organic carbon and dissolved oxygen all have been found to affect the diversity, activity and metabolism of the microbial community in different HZ environments [33]. Recent studies on diversity have detected microorganisms such as fungi, heterotrophic aerobic bacteria, aerobic and anaerobic ammonium-oxidizing bacteria in the HZ [30,34,35]. Little information is available regarding the value of microbial indicators for fecal pollution in tropical regions [41]. Contamination of well waters by Salmonella spp. has been documented [42]. Salmonella spp. were detected in the HZ by Braioni et al. [43]. However, a significant knowledge gap exists in terms of the identities and roles of enteric bacteria in neotropical HZs. The aims of this study are: (1) to illustrate the results of the first survey of enteric bacteria in a neotropical stream, (2) to determine possible correlations between microbiological, physicochemicaland environmental parameters of the interstitial HZ, and (3) to define variables that influence this microbiota to lead future research in the neotropical region.
Study Area, Sampling Characterization and Analysis
Study Area
Brazilian legislation does not require permission for sampling water, but our sampling permission reference is:Ministerio do Meio Ambiente-ICMbio-SISBIO n.32349-2 23/04/2013.
The Tijuca National Park is located entirely within the urban area of Rio de Janeiro, Brazil, between S22° 55’-S23° 00’ and W43° 11’-W43° 19’and contains an area of approximately 32 km2. Its vegetation is characteristic of the Atlantic Forest biome [44]. The climate is humid and subtropical, with an average annual temperature between 20°C and 25°C and an annual rainfall of more than 1,500 mm. The distribution of rainfall is characterized by two seasons: the rainy season, between November and February (with more than 250 mm/month of rain), and the dry season, from June to September (with less than 100 mm/month of rain). At other times of the year, the rainfall is within these limits. The geological substratum is predominantly comprised of granite [45].
Samples were collected in October 2012 from three stream reaches (P I: 22°57'35.68"S, 43°16'33.54"W, 530 m altitude; P II: 22°57'13.08"S, 43°16'55.45"W, 475 m altitude; P III: Lat. 22°56'56.15"S, Long 43°17'9.84"O, 355 m altitude), located in the first, second and third orders, respectively (sensu Strahler [46]) in the Tijuca River (Fig 1).
Fig 1. Map of the study area.

Map of the study area showing the sampling sites in the Tijuca River (Tijuca National Park, Rio de Janeiro, Rio de Janeiro State, Brazil).
To describe the sampling sites a visual-based habitat assessment system was used [47]. The habitat was defined valuing the structure of the surrounding physical habitat that influences the condition of the resident aquatic community as the variety and quality of the substrate, channel morphology, bank structure, and riparian vegetation.
The sampling sites used were quite different. In the first order site (P I), boulders covered 80% of the riverbed and the sand filled the cavities between the rocks; the transverse section of the riverbed was concave. The second order site (P II) was less steep, with sand representing 60% of the riverbed. The rest was comprised of cobble and coarse gravel, with regular riffles and pools; in this site, the transverse section of riverbed was flat. In the third order site (P III), the riverbed was 60% boulders, 30% pebbles, and 10% coarse sand. Here, the riffle-pool sequences were less regular and were separated by long areas of fine sediment deposition; the transverse section of the riverbed was concave. In this site, the stream received no treated effluents from the park administration and visitation center.
The Alfonso Viseu water-collecting checkpoint for CEDAE (the water company of Rio de Janeiro city) supplied street of the Alto da Tijuca neighborhood of Rio de Janeiro and was located three hundred meters downstream of the second order site.
Sample collection
At each site, polyvinyl chloride (PVC) mini-piezometers of 3/4" (2.6 cm) in diameter with holes of 0.5 cm in diameter distributed along a 5 cm perforated band were positioned in July 2012 at three depths: 10 cm (I); 25 cm (II) and 45 cm (III) (Fig 2).
Fig 2. Mini-piezometers.

Mini-piezometers and the position of the perforated bands at the different sampling depths.
No water was found in the sampling site located at a depth of 45 cm in the third order. For each depth, five mini-piezometers were positioned in four habitat typologies: up riffle (a), down riffle (b), lateral side of pools (c) areas of sand accumulation (bars, curves) (d1 and d2), for a total of 40 mini-piezometers.
Hyporheic and surface water samples of 0.5 liters each were collected in October 2012 using a diaphragm pump. However, four mini-piezometers were occluded, so a total of thirty-six samples were collected. All samples were stored and transported in a cool box kept below 4°C.
Environmental parameters: hydraulics and limnology
In the field, the Vertical Hydraulic Gradient (VHG), Hydraulic Conductivity (KH), water suction time, oxygen, temperature and mini-piezometer depth were determined as described in Malard et al. [37].
The VHG describes the direction and intensity of the water exchange between the HZ and the surface or the subterranean zone. KH is a measure of the ability of a porous material to allow fluids to pass through it. Oxygen and temperature were measured in the mini-piezometer using an oximeter.
In the laboratory, several physicochemical parameters were assessed. These included the Total Hardness, Calcium Hardness, Magnesium Hardness, Total Alkalinity, Hydroxide Alkalinity, Carbonate Alkalinity, Bicarbonate Alkalinity in mg/L CaCO3, mg/L Ca, mg/L Mg, pH, SO4 -, Chloride (mg/L Cl-), Conductivity (μS/cm), mg/L OH-, mg/L CO-, mg/L HCO-, mg/L Total Dissolved Solids (TDS), mg/L salinity, mg/L N-Nitrogen, mg/L N-Nitrite, and Sulfate (mg/L SO4 -). The alkalinity was determined via titration with indicators, while the total hardness and calcium hardness were measured by using titration with EDTA. The chloride parameters were determined with the Mohr titration method, the conductivity and TDS were determined by the electrometric method using a conductivity meter, and the Nitrogen ammonia was determined via the Nessler method. The sulfate levels were measured by the turbidimetric method, the Nitrogen nitrite levels by the diazotization method, and the pH by the potentiometric method, using a pH meter as recommended for the APHA [48], FEEMA [49] and FUNASA [50] methods. All analyses were completed in duplicate, and the results are expressed as means.
Bacteriological Analysis
Hyporheic and surface water samples were collected in sterile bottles and transported under refrigeration for immediate laboratory processing. Water samples (5 mL) were diluted (10-1 to 10-2) in phosphate water (45 mL, solution A 0.25 M KH2PO4, solution B 0.4 M MgCl2.6H2O, pH 7.5) and filtered (10 mL) on a 0.45 μm cellulose acetate membrane (Millipore) [51]. The surface plate technique was used for the simultaneous detection of the total coliforms, E. coli and Salmonella spp. Duplicate plates were used for each dilution step. The membrane containing the retained bacterial cells was inoculated onto Chromocult coliform agar (CCA) for 18–24 h at 37°C [41]. After the bacterial growth period, colonies were assigned to bacterial species using their phenotypic traits, including morphological and physiological characteristics. Colonies between a salmon and red color were produced by salmon-galactoside cleavage by b-D-galactosidase, and these were classified the coliforms. In contrast, the dark blue to violet colonies resulted from salmon-galactoside and X-glucuronide cleavage by b-D-galactosidase and b-D-glucuronidase, and such colonies were classified as E. coli. Color-less colonies were instead classified as Salmonella spp. The results from the quantification of bacterial contamination were expressed as colony-forming units (CFU) per milliliter of water [52]. All tests were performed in duplicate.
Statistical Analysis
For the statistical analysis, the data were log(x+1) transformed. To avoid collinearity between the variables, a Pearson’s correlation was calculated, excluding correlated variables pairs with r>0.6 or r<-0.6 [53]. To compare the physicochemical data between the sites, the micro-habitats and the depths, a Principal Component Analysis (PCA) was completed with a correlation matrix. ANOVA analysis was used to compare the bacteriological characteristics between the sites, micro-habitats and depths. Canonical Correspondence Analysis (CCA) elucidated the relationships between the biological assemblages and their environment. The ordinal variables, such as the river order and the micro-habitat, were treated quantitatively, with the coding 0, 1, 2, 3 and so on [54]. Tests were performed using GraphPad Prism 5.0 and Past packages.
Results
The total coliform counts were detected in all samples. E. coli and Salmonella spp. were each detected in the surface of one site: E. coli was detected at a concentration of 0.1 CFU/ml in the third order site, and Salmonella spp. was detected at a concentration of 0.5 CFU/ml in the first order site. In the hyporheic samples, the number of positive samples varied (Table 1).
Table 1. Bacteriological analysis and percentages of positive results.
| Depth | Totals Coliforms % | Escherichia coli % | Salmonella % |
|---|---|---|---|
| surface | 100 | 33 | 33 |
| I | 100 | 17 | 92 |
| II | 100 | 29 | 86 |
| III | 100 | 40 | 80 |
The maximum total coliform concentration (9.5 CFU) was found at a depth of 45 cm in the P III site on the wall of a pool. The highest values, between 2.5 and 3.5, for sites P I and P II were found at depths of 10 and 25 cm (Fig 3).
Fig 3. Total coliform.
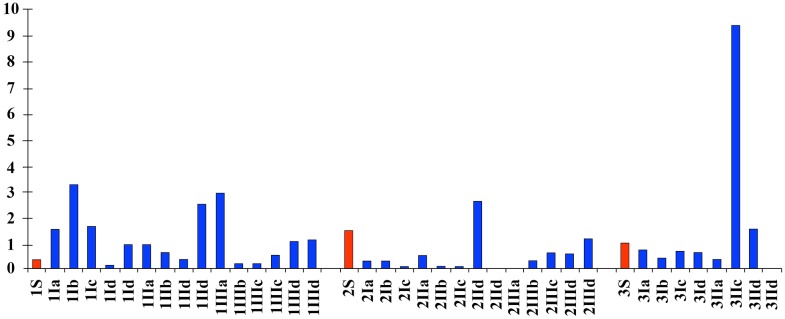
The total coliform levels (CFU /ml) at different sites, depths and habitat typologies. Surface samples are shown in red. Arabic numbers represent the sampling site. Roman numbers represent the sampling depth: I: 10 cm, II: -25 cm, III: -45 cm. Letters: a—up riffle, b—down riffles, c—lateral side of pools, d—areas of sand accumulation (bars, curves).
The maximum E. coli concentration (0.15 CFU) was found at the P I site at a depth of 45 cm on depth in a lateral side on pool. The highest value of sites P I, P II and P III at 10 and 25 cm depths was 0.1 CFU (Fig 4).
Fig 4. Eschericia coli.
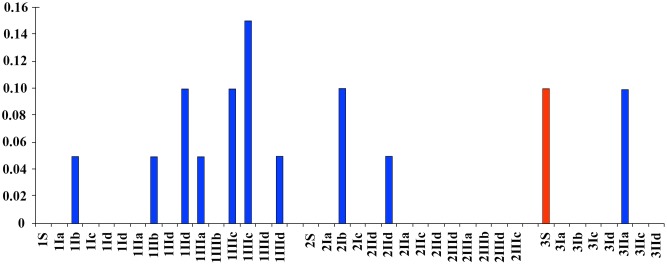
The concentrations of E. coli (CFU/ml) found at different sites, depths and habitat typologies. The surface samples are shown in red. Arabic numbers represent the sampling site. Roman numbers represent the sampling depth; the sampling depths were I—10 cm, II—25 cm, III -45 cm. The letters indicate the following: a—up riffle, b—down riffle, c—lateral side on pools, d—sand accumulation areas (bars, curves).
The maximum concentration of Salmonella spp. (67.2 CFU) was found at site P III at a depth of 25 cm on lateral side on pools a pool. High values of 3.3 and 2.5 were found in the P I and P III sites at 45 and 25 cm depths lateral side on pools and in areas of sand accumulation (Fig 5).
Fig 5. Salmonella spp.

Amount of Salmonella spp. (CFU/ml) found at different sites, depths and habitat typologies. In red, surface samples. Arabic numbers represent the sampling site. Roman numbers represent the sampling depth: I, -10 cm; II, -25 cm; III, -45 cm. The letters represent: a—up riffle; b—down riffles; c—lateral side of pools; d—areas of sand accumulation (bars, curves).
The results of the physical and chemical analysis of surface and hyporheic samples are shown as mean values and standard deviations (Table 2). The most variable parameters were the conductivity and oxygen, while the least variable parameter was the pH; all decreased with increasing depth. From the physiochemical standpoint, the P II site is the most heterogeneous. In this sampling site, the calcium hardness and sulfate parameters were high and variable. In contrast, at PI and PIII, the temperature was lower and the oxygen and conductivity were higher, both varying with the minipiezometers.
Table 2. Physicochemical parameters.
| Site | P I | P II | P III | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| depth | SURF | I | II | II | SURF | I | II | III | SURF | I | II | ||||||||
| mean | SD | mean | SD | mean | SD | mean | SD | mean | SD | mean | SD | mean | SD | mean | SD | ||||
| HardCa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 11.20 | 12.08 | 7.68 | 11.05 | 28.80 | 26.51 | 0.00 | 0.00 | 0.00 | 2.40 | 4.80 |
| pH | 5.79 | 5.92 | 0.08 | 5.65 | 0.20 | 5.61 | 0.18 | 5.69 | 5.17 | 0.36 | 5.41 | 0.31 | 4.92 | 0.11 | 6.17 | 5.79 | 0.24 | 5.77 | 0.03 |
| Sulfate | 4.02 | 4.65 | 1.20 | 6.36 | 0.98 | 7.01 | 4.50 | 2.93 | 27.26 | 31.14 | 17.63 | 17.92 | 53.52 | 43.59 | 4.39 | 8.20 | 7.27 | 4.16 | 0.89 |
| Chlor | 11.54 | 11.54 | 2.10 | 13.64 | 2.57 | 12.77 | 2.85 | 11.54 | 11.54 | 0.00 | 11.12 | 1.76 | 11.54 | 1.48 | 15.74 | 13.64 | 2.97 | 12.07 | 1.05 |
| Cond | 49.60 | 51.84 | 1.10 | 54.35 | 2.10 | 52.37 | 0.97 | 48.70 | 78.73 | 37.78 | 67.50 | 24.30 | 115.54 | 51.74 | 53.70 | 60.72 | 9.52 | 55.40 | 1.13 |
| Tot Alk | 14.72 | 9.20 | 1.84 | 10.12 | 3.52 | 8.59 | 3.00 | 11.04 | 7.37 | 0.02 | 7.74 | 2.01 | 8.10 | 3.08 | 11.04 | 9.57 | 2.94 | 9.21 | 2.11 |
| N-Nitrog | 0.00 | 0.00 | 0.00 | 0.02 | 0.03 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.01 | 0.03 | 0.01 | 0.02 | 0.00 | 0.01 | 0.02 | 0.00 | 0.00 |
| Nitrite | 0.00 | 0.01 | 0.00 | 0.01 | 0.01 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.01 | 0.09 | 0.16 | 0.00 | 0.01 | 0.01 | 0.00 | 0.00 |
| Ox% | 52.50 | 27.02 | 4.97 | 31.00 | 8.82 | 20.58 | 9.09 | 42.70 | 38.30 | 14.13 | 31.26 | 13.81 | 34.20 | 9.52 | 27.50 | 17.64 | 7.87 | 18.93 | 10.25 |
| Temp | 20.20 | 21.52 | 0.88 | 20.53 | 0.52 | 20.82 | 1.31 | 20.30 | 19.17 | 1.05 | 19.16 | 0.61 | 19.36 | 0.68 | 21.20 | 21.30 | 0.58 | 21.35 | 0.37 |
The physicochemical parameters of the surface and HZ, shown as the mean and standard deviation.
The P I, P II and P III, sampling sites were at I—10 cm, II—25 cm, and III—45 cm. SD, standard deviation; HardCa, calcium hardness; Chlor, chloride; Cond, conductivity; N-Nitrog, N-nitrogen; Temp, temperature.
A two-way ANOVA demonstrated significant physicochemical differences (P<0.0001, F 9, 49) the test was followed by a post tests two-way ANOVA using the Bonferroni method that show significant differences between P I and P II, as determined by the total hardness (P<0.01), calcium hardness (P<0.01), conductivity (P<0.001), TDS (P<0.001) and sulfate (P<0.001).
The CCA analysis with bacteriological, physicochemical and environmental data (axis 1 eigenvalue 0.18679, % variance 57.86) discriminates Salmonella spp. from E. coli and the total coliforms (axis 2 0.039992, % variance 12.90) and discriminates the total coliforms from Salmonella spp. and E. coli. All three bacterial types were significantly influenced by the site, especially Salmonella spp. On axis 1, Salmonella spp. was found to be correlated with the oxygen level (-), river order (site) (+), nitrite (+), calcium hardness (-), temperature (+), sulfate (-) and VHG (-). In contrast, the total coliforms and E. coli levels were correlated with the same factor but in the opposite way. On axis 2, the total coliforms and Salmonella spp. are correlated with VHG (+), pH (+), KH (+), suction time (-), Conductivity (-), Sulfate (-) and the micro-habitat (-). The E. coli levels were correlated with the same factors, but in the opposite way (Fig 6).
Fig 6. CCA analysis.

CCA analysis with physicochemical, bacteriological and environmental data. The first order samples are shown in green, the second order samples are shown in red, and the third order samples are shown in blue.
The same analyses were performed when excluding data from mini-piezometer 3IIc. The results showed no substantial changes.
The PCA analysis with the physicochemical data shows differences in P II relative to the other sampling sites (axis 1 eigenvalue 5.10935, % variance 73.21; axis 2 eigenvalue 2.76331, % variance 15.88) (Fig 7), especially in its calcium hardness (0.4202), conductivity (0.4161), STD (0.416), sulfate content (0.4086) and pH (-.3263). Of note, sulfates and Ca hardness could be detected only in this site.
Fig 7. PCA with physicochemical data.

The first order samples are shown in green, the second order samples are shown in red, and the third order samples are shown in blue.
The relationship between Salmonella spp., the total coliform count, E. coli, and the total hardness and sulfate concentration is shown in a histogram (Fig 8).
Fig 8. Salmonella spp., total coliforms, Escherichia coli, total hardness and sulfates.

Histogram showing the Salmonella spp., total coliforms, Escherichia coli, total hardness and sulfates logarithmic scale for all measured sampling sites and depths.
Discussion
Since 1914, coliform counts have been used as a bioindicator of the microbiological quality of treated and untreated drinking water and as a standard indicator of recent fecal contamination in most countries.
Historically, many countries have been using fecal indicator bacteria as a monitoring tool to predict the presence of microorganims originating fromin fecal contamination. E. coli is still the bacteria that best satisfies the fecal indicator microorganism criteria since it is a species that is specific to the intestines of humans and other warm-blooded animals but is not necessarily pathogenic. Total coliforms include bacterial species of fecal origin, as well as other bacterial groups that commonly occur in the soil. Moreover, the persistence in water, the relationship between fecal indicator bacteria and the occurrence of diseases may not be accurate in all the locations of sampling and environmental settings [9,10,55,56]. Additionally, indicators may be severely underrepresented when injured by disinfectants used in water treatment processes.
The persistence and survival of these bacteria in underground media have been shown to be significantly influenced by a complex array of physical, chemical, and biological factors, including the concentration of indigenous microorganisms, the growth and decay rates, the concentration of available nutrients and the consumption kinetics [13, 57].
Hendricks and Morrison [58] showed that enteric bacteria could multiply in both polluted and unpolluted river water at temperatures as low as 10°C. Hendricks [59] and Burton et al. [60] reported an increased recovery rate of streambed sediment in relation to surface waters due to a combination of sedimentation and sorption, which provides protection from bacteriophages and microbial toxicants. The adhesion of Salmonella spp. cells to soil particles is correlated with cell surface hydrophobicity [61]. Bacterial association with soil particles provides bacteria with high concentrations of nutrients due to the release of organic molecules from attached algal cells. The association also gives the bacteria protection against predation, by providing shelter against protozoan grazing [25]. The long-term survival of Salmonella in water, manure, soil and sediment has been well documented [12], and a high quantity of coliforms have been recorded in unpolluted tropical waters [13,18,62]. As in the mammalian host environment, nutrients in some tropical ecosystems are maintained at high concentrations. With consistently warm air, soil, and water, they can provide an ideal habitat for the survival, growth, and proliferation of E. coli [13,18,62].
The prevalence of Salmonella spp. in the water is highly influenced by conductivity, temperature, alkalinity and oxygen concentration [63,64]. Jimenez et al. [17], Carrillo et al. [18] and Burton et al. [60] have all reported a strong positive correlation with the water temperature and nutrient concentrations and E. coli cells found in superficial water located upstream of known fecal sources in tropical rain forest ecosystems. Accordingly, our data identify the same parameters as being important determinants but also point to other key factors, such as the structure and composition of meso and microhabitat.
In the P III sampling site located in the third order at a depth of 45 cm, no water was found. This may be due to the presence of a cascade followed by a small upstream dam of 1.5 m in height. Dams along small-order streams are known to alter the link between the headwater and the downstream areas, potentially increasing the hyporheic connectivity in the riparian zone over short distances [65]. However, dams can also simplify the alluvial system structure, reducing peak flows, preventing rejuvenation of alluvial aquifer structure and reducing the hyporheic flow [66].
The P I and P III sampling sites were quite similar and were distinct from the P II site of second Straler’s river order in sand/silt quantity, heterogeneity, concavity, width and steepness, resembling a higher Straler’s order in structure.
The HZ can be an important sink or source for the stream ecosystem and is an important site for the remineralization of organic matter [38]. The organic matter found in the hyporheic sediments originates from surface carried by the downward flow of stream water [67,68] and from decaying organic particles, dissolved molecules, free-living organisms and attached biofilms [69,70].
Anderson et al. [71] and White [72] reported that the magnitude of vertical hydraulic gradients varies along the longitudinal stream profile. The hyporheic exchange flow increases predictably with increases in the water surface concavity, heterogeneity and channel unit spacing in headwater and mid-order streams [71]. Downwelling occurs when the stream profile is convex, when permeability increases in the downstream direction, or when the bed depth increases in the downstream direction. Conversely, upwelling occurs when the stream profile is concave, when permeability decreases in the downstream direction, or when the bed depth decreases in the downstream direction. The downwelling and upwelling zones differ significantly in terms of the temperature, pH, redox potential, and the dissolved oxygen and nitrate levels, with significant positive correlations occurring among the latter three factors. This creates distinct patterns in the microbial activity [73,74].
Sulfur is one of the major constituents of ground water, commonly occurring as sulfate molecules. In anoxic conditions, biofilms can use calcium for respiration [38]. These facts justify the measurements of Ca hardness and sulfate levels in the P2 sampling site. Calcium can be derived from the dissolution of plagioclase and/or fluorite minerals from volcanic and metamorphic stones; sulfates are generally of organic origin in anoxic environments and arrive via the upwelling process.
This work show that viable enterobacteria are present and probably reproduce in the HZ. Our results demonstrate that enteric bacteria are heterogeneously distributed along the sampling sites assessed. Their spatial variation was related to environmental factors and physical-chemical parameters within the HZ and may play a key role in the microbial diversity and distribution within these ecosystems. These factors possibly influence the survival and viability of bacteria. Our results revealed that Salmonella spp. and coliforms have quite different microhabitat preferences. Salmonella spp. and coliforms both are frequently deposited on the lateral sides of pools and curves and bars, but the two tend to distribute into different depths, probably due to the temperature. However, Salmonella spp. prefers compact substrata with a lower fluid flow and upwelling areas with less oxygen inflow. In contrast, coliforms have the opposite preference. Statistical analyses suggest the existence of different habitat preferences between the total coliforms and E. coli, but our data do not permit us to discriminate. The recovery of enteric isolates from particular HZ microzones exhibiting extreme physiological conditions was unexpected. These results strongly support the hypothesis that environmental enteric isolates may exhibit an important adaptive advantage capable of surviving and proliferating in this ecosystem. The circulation and persistence of these microorganisms in the HZ may promote close interaction and horizontal genetic exchanges. Considering that E. coli and Salmonella isolates may comprise virulent strains, the recovery of these microorganisms from the HZ water samples have implications for human health and disease risk. Virulence genetic markers are commonly encoded on mobile genetic elements and promote the rapid evolution of bacteria and the emergence of variants within species, including, pathogenic lineages.
Our data revealed the presence of viable enterobacteria in the HZ of a neotropical stream in Rio de Janeiro, Brazil, whose distribution differed significantly across microzones. This is of special relevance considering that circulating bacteria may comprise virulent strains or reservoirs of resistance determinants. The presence of potentially pathogenic bacteria in HZ raises questions about the increased risk to ecosystems and human health. Future work is needed in order to investigate molecular markers to better clarify the diversity, pathogenic potential and the biology of these circulating microorganisms as well as the population dynamics in this aquatic ecosystem. Similar to our work, a prior study, using metagenomic and amplicon sequencing, showed how environmental parameters can alter river sediment microbial community structure and function [75].
Conclusions
The present study detected a wide occurrence of enterobacteria in HZ microzones of Tijuca National Park which spatial location leads to the hypothesis that these microorganisms may be sensitive to environmental parameters. The detection of E.coli and Salmonela in HZ does not guarantee that circulating microorganisms are pathogenic or resistant, thus requiring additional characterization target to specific virulence traits and antibiotic resistance markers. This may represent a safety risk, in view of the usage of this area’s water for human consumption and recreational activities.
Therefore, we must reevaluate the use of these bacteria as indicators of human-caused pollution in tropical regions. Further studies are needed to define the precise conditions and microhabitats necessary to determine the life cycle of these organisms in the HZ environment to prevent future health hazards.
Acknowledgments
Many thanks to Dr. Kury Adriano Museu Nacional/UFRJ for allowing the research in the Arachnology Laboratory. Thank you also to Gatti Mario Jorge Curator of Coleção Micológica de Trichocomaceae IOC/FIOCRUZ for technical support and to Wander Ribeiro Ferreira for the helpful suggestions.
Funding Statement
The research is a partial result of a postdoctoral activity. The research has as sources CAPES (Coordenação de Aperfeçoamento de Pessoal de nível Superior), who paid the postdoctoral salary (about $1,700), and the FAPERJ (Fondação do Emparo a Pesquisa do Estado do Rio de Janeiro), who paid for laboratory costs (about $416, including car gasoline, reagents and equipment). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Akter S, Islam M, Afreen KS, Azmuda N, Khan SI, Birkeland NK (2013) Prevalence and distribution of different diarrhoeagenic Escherichia coli virulotypes in major water bodies in Bangladesh. Epidemiol Infect 1–10. 10.1017/S0950268813002367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Harwood VJ, Staley C, Badgley BD, Borges K, Korajkic A (2013) Microbial source tracking markers for detection of fecal contamination in environmental waters: relationships between pathogens and human health outcomes. FEMS Microbiol Rev 1–40. 10.1111/1574-6976.12031 [DOI] [PubMed] [Google Scholar]
- 3. Lanata CF, Fischer-Walker CL, Olascoaga AC, Torres CX, Aryee MJ, Black RE (2013) Global causes of diarrheal disease mortality in children <5 years of age: a systematic review. Plos One 8(9): 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Maal-Bared R, Bartlett KH, Bowie WR, Hall ER (2013) Phenotypic antibiotic resistance of Escherichia coli and E. coli O157 isolated from water, sediment and biofilms in an agricultural watershed in British Columbia. Sci Total Environ 443: 315–323. 10.1016/j.scitotenv.2012.10.106 [DOI] [PubMed] [Google Scholar]
- 5. Rebello RCDL, Regua-Mangia AH (2014) Potential enterovirulence and antimicrobial resistance in Escherichia coli isolates from aquatic environments in Rio de Janeiro, Brazil. Sci Total Environ 490: 19–27. 10.1016/j.scitotenv.2014.04.040 [DOI] [PubMed] [Google Scholar]
- 6. Tallon P, Magajna B, Lofranco C, Leung KT (2005) Microbial Indicators of Fecal Contamination in Water: A Current Perspective. Water, Air, and Soil Poll 166(1–4): 139–166. [Google Scholar]
- 7. Committee on Indicators for Waterborne Pathogens, National Research Council (2004) Indicators for waterborne pathogens. National Academies Press, Washington, DC: 332p. [Google Scholar]
- 8. Toranzos GA, Mcfeters GA (1997) Detection of indicator microorganisms in environmental fresh waters In: Hurst CJ, Knudsen GR, McInerney MJ, Stetzenbach LD, Walter MV, editors. Manual of environmental microbiology. American Society for Microbiology, Washington, D.C: pp.184–194. [Google Scholar]
- 9. Torres AG, Arenas-Hernández MMP, Martínez-Laguna Y (2010) Overview of Escherichia coli In: Torres AG, editor. Pathogenic Escherichia coli in Latin America. Bentham Science e-books, San Diego, pp.1–7. [Google Scholar]
- 10. Williams ND, Torres AG, Lloyd SJ (2010) Evolution and epidemiology of diarrheagenic Escherichia coli In: Torres AG, editor. Pathogenic Escherichia coli in Latin America. Bentham Science e-books, San Diego, pp. 8–24. [Google Scholar]
- 11. Savageau MA (1983) Escherichia coli habitats, cell types, and molecular mechanisms of gene control. Am Nat 122:732–744. [Google Scholar]
- 12. Winfield MD, Groisman EA (2003) Role of nonhost environments in the lifestyle of Salmonella and Escherichia coli . Appl Environ Microbiol 69(7): 3687–3694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Anderson KL, Whitlock JE, Valerie J, Harwood VJ (2005) Persistence and Differential Survival of Fecal Indicator Bacteria in Subtropical Waters and Sediments. Appl Environ Microbiol 71(6): 3041–3048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Byappanahalli MN, Fujioka RS (1998) Evidence that tropical soil can support the growth of Escherichia coli. Water Sci Technol 38: 171–174. [Google Scholar]
- 15. Desmarais TR, Solo-Gabriele HM, Palmer CJ (2002) Influence of soil on fecal indicator organisms in a tidally influenced subtropical environment. Appl Environ Microbiol 68: 1165–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Solo-Gabriele HM, Wolfert MA, Desmarais TR, Palmer CJ (2000) Sources of Escherichia coli in a coastal subtropical environment. Appl Environ Microbiol 66: 230–237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Jimenez L, Muniz I, Toranzos GA, Hazen TC (1989) Survival and activity of Salmonella typhimurium and Escherichia coli in tropical freshwater. J Appl Bacteriol 67: 61–69. [DOI] [PubMed] [Google Scholar]
- 18. Carrillo M, Estrada E, Hazen TC (1985) Survival and enumeration of the fecal indicators Bifidobacterium adolescentis and Escherichia coli in a tropical rain forest watershed. Appl Environ Microbiol 50: 468–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Hoff GL, White FH (1977) Salmonella in Reptiles: Isolation from Free-Ranging Lizards (Reptilia, Lacertilia) in Florida. J Herpetol 11(2): 123–129. [Google Scholar]
- 20. Kawakami M, Ihara I, Ihara S, Suzuki A, Fukui K (1984) A group of bactericidal factors conserved by vertebrates for more than 300 million years. J Immunol 132(5): 2578–2581. [PubMed] [Google Scholar]
- 21. Lins ZC (1964) Ocorrência do gênero Salmonella em animais silvestres capturados na floresta amazônica. An Microbiol Rio de Janeiro 12: 19–29. [Google Scholar]
- 22. Simpson VR (2002) Wild Animals as Reservoirs of Infectious Diseases in the UK. Vet J 163(2): 128–146. [DOI] [PubMed] [Google Scholar]
- 23. Levantesi C, Bonadonna L, Briancesco R, Grohmann E, Toze S, Tandoi V (2012) Salmonella in surface and drinking water: Occurrence and water-mediated transmission. Food Res Internat 45(2): 587–602. [Google Scholar]
- 24. Harwood VJ, Whitlock J, Withington V (2000) Classification of antibiotic resistance patterns of indicator bacteria by discriminant analysis: use in predicting the source of fecal contamination in subtropical waters. Appl Environ Microbiol 66: 3698–3704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Fish JT, Pettibone GW (1995) Influence of freshwater sediment on the survival of Escherichia coli and Salmonella sp. as measured by three methods of enumeration. Lett Appl Microbiol 20: 277–281. [DOI] [PubMed] [Google Scholar]
- 26. Sherer BM, Miner JR, Moore JA, Buckhouse JD (1992) Indicator bacterial survival in stream sediments. J Environ Qual 21: 591–595. [Google Scholar]
- 27. Rivera SC, Hazen TC, Toranzos GA (1988) Isolation of Fecal Coliforms from Pristine Sites in a Tropical Rain Forest. Appl Environ Microbiol 54(2): 513–517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Bencala KE (2000) Hyporheic zone hydrological processes. Hydrol Process 14: 2797–2798. [Google Scholar]
- 29. Orghidan T (1959–2010). Ein neuer Lebensraum des unterirdischen Wasser: der hyporheische Biotop. Arch Hydrobiol 55: 392–414. [Google Scholar]
- 30. Bärlocher F, Nikolcheva LG, Wilson KP, Williams DD (2006) Fungi in the Hyporheic Zone of a Springbrook. Microbial Ecol 52: 708–715. [DOI] [PubMed] [Google Scholar]
- 31. Danielpol DL, Griebler C, Gunatilaka A, Hahn HJ et al. (2008) Incorporation of groundwater ecology in environmental policy In: Groundwater Science and Policy: An International Overview. Royal Society of Chemistry, 2008, 754 p. [Google Scholar]
- 32. Dole-Olivier MJ (1998) Surface water–groundwater exchanges in three dimensions on a backwater of the Rhône River. Freshwater Biol 40: 93–109. [Google Scholar]
- 33. Franken RJM, Storey RG, Williams DD (2001). Biological, chemical and physical characteristics of downwelling and upwelling zones in the hyporheic zone of a north-temperate stream. Hydrobiologia 444(1–3): 183–195. [Google Scholar]
- 34. Halda-Alija L, Hendricks SP, Johnston TC (2001) Spatial and Temporal Variation of Enterobacter Genotypes in Sediments and the Underlying Hyporheic Zone of an Agricultural Stream. Microb Ecol 42: 286–294. [DOI] [PubMed] [Google Scholar]
- 35. Wang Z, Qi Y, Wang J, Pei Y (2012) Characteristics of aerobic and anaerobic ammonium-oxidizing bacteria in the hyporheic zone of a contaminated river. World J Microbiol Biotechnol 28: 2801–2811. 10.1007/s11274-012-1090-6 [DOI] [PubMed] [Google Scholar]
- 36. Bretschko G, Leichtfried M (1988) Distribution of organic matter and fauna in a second order alpine gravel stream (Ritrodat-Lunz study area, Austria). Verh Intern Ver Limnol 23: 1333–1339. [Google Scholar]
- 37.Malard F, Dole-Olivier MJ, Mathieu J (2002) Sampling manual for the assessment of regional groundwater biodiversity. Malard F, Dole-Olivier MJ, Mathieu J, Stoch F (2002) PASCALIS D4 Deliverable for Workpackage 4: Sampling manual for the assessment of regional groundwater biodiversity. European Project Protocols for the Assessment and Conservation of Aquatic Life in the Subsurface (PASCALIS; No EVK-CT-2001-00121)
- 38. Storey RG, Fulthorpe RR, Williams DD (1999) Perspectives and predictions on the microbial ecology of the hyporheic zone. Freshwat Biol 41(1): 119–130. 10.1046/j.1365-2427.1999.00377.x [DOI] [Google Scholar]
- 39. Khebiza M, Boughrous AA, Gabbanini C, Messouli M, Messana G (2006) Impact of waste discharges on the water quality and interstitial community structure of two Mediterranean rivers. Ital J Zool 73(2): 153–166. [Google Scholar]
- 40. Richards C, Bacon KL (1994) Influence of fine sediment on macroinvertebrate colonization of surface and hyporheic stream substrates. Great Basin Nat 54: 106–113. [Google Scholar]
- 41. Byamukama D, Kansiime F, Mach RL, Farnleitner AH (2000) Determination of Escherichia coli Contamination with Chromocult Coliform Agar Showed a High Level of Discrimination Efficiency for Differing Fecal Pollution Levels in Tropical Waters of Kampala, Uganda. Appl Environ Microbiol 66(2): 864–868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. McFeters GA, Bissonnette GK, Jezeski JJ, Thomson CA, Stuart DG (1974) Comparative survival of indicator bacteria and enteric pathogens in well water. Appl Environ Microbiol 27(5): 823–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Braioni MG, Salmoiraghi G, Bracco F, Villani M, Braioni A, Girelli L (2002) Functional Evaluations in the Monitoring of the River Ecosystem Processes: The Adige River as a Case Study. Freshwat Sci 2: 660–683.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Martins MF (2011) Historical biogeography of the Brazilian Atlantic forest and the Carnaval–Moritz model of Pleistocene refugia: what do phylogeographical studies tell us? Biol J Linn Soc 104: 499–509. [Google Scholar]
- 45. Brasil. Ministerio das Minas e Energia; (1987) Projeto Radambrasil Levantamento de recursos naturais. IBGE, Brasilia: Vol 32 Pp. 775. [Google Scholar]
- 46. Strahler AN (1957) Quantitative Analysis of Watershed Geomorphology. Trans Am Geoph Un 38: 913–920. [Google Scholar]
- 47. Barbour MT, Gerritsen J, Snyder BD, Stribling JB (1999) Rapid Bioassessment Protocols for use in Streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish. Second Edition EPA/841-B-99-002. U.S. EPA, Office of Water, Washington, D.C: pp. 197 plus appendices. [Google Scholar]
- 48. APHA. American Public Health Association. (2005) Standard methods for the examination of water and wastewater. 20th ed APHA, Washington, DC: pp. 541. [Google Scholar]
- 49. FEEMA. Fundação Estadual de Engenharia do Meio Ambiente; (Rio de Janeiro: ). (1982) Métodos de análises físico-químicas da água Rio de Janeiro. Sumula n. 8. Vol. 3 pp. 86. [Google Scholar]
- 50. FUNASA. Fundação Nacional de Saúde. (2006) Manual prático de análise de água 2. ed rev. Brasília,. 146 p. [Google Scholar]
- 51. Eaton AD, Clescerl LS, Rice EW, Greenberg AE, Franson MA (2005) Standard Methods for the Examination of Water and Wastewater, 21st Edition American Water Works Association; New York: pp. 1368. [Google Scholar]
- 52. Brasil. Ministério da Saúde, Fundaçäo Nacional de Saúde (1999) Manual técnico: análise de água para consumo humano / Technical Manual—Analysis of Water for Human onsumption. Fundaçäo Nacional de Saúde, Brasília: pp. 211. [Google Scholar]
- 53. Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, et al. (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography, 36(1), 27–46. [Google Scholar]
- 54. Braak CJF, Verdonschot PFM (1995) Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat Sci 57(3): 255–289. 10.1007/BF00877430 [DOI] [Google Scholar]
- 55. Bridle H (2014) Waterborne Pathogens: Detection Methods and Applications, first ed AcademicPress,Amsterdam; [Google Scholar]
- 56. Savichtcheva O, Okabe S (2006) Alternative indicators of fecal pollution: relations with pathogens and conventional indicators, current methodologies for direct pathogen monitoring and future application perspectives.WaterRes.40, 2463–2476 [DOI] [PubMed] [Google Scholar]
- 57. Bermudez M, Hazen TC (1988) Phenotipic and genotypic camparison of Escherichia coli from pristine tropical waters. Appl Environ Microbiol 54: 979–983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Hendricks CW, Morrison SM (1967) Multiplication and growth of selected enteric bacteria in clear mountain stream water. Water Res 1: 567–576. [Google Scholar]
- 59. Hendricks CW (1971) Increased Recovery Rate of Salmonellae from Stream Bottom Sediments Versus Surface Waters. Appl Environ Microbiol 21(2): 379–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Burton GA Jr., Gunnison D, Lanza GR (1987) Survival of Pathogenic Bacteria in Various Freshwater Sediments. Appl Environ Microbiol 53(4): 633–638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Stenstrom TA (1989) Bacterial hydrophobicity, an overall parameter for the measurement of adhesion potential to soil particles. Appl Environ Microbiol 55: 142–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Perez-rosas N, Hazen TC (1989) In Situ Survival of Vibrio cholerae and Escherichia coli in a Tropical Rain Forest Watershed. Appl Environ Microbiol 55(2): 495–499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Ateba BH, Nougang ME, Noah EO, Tamatcho KB, Tawedi RE, et al. (2012) Occurrence of Salmonella spp in surface waters of Yaoundé, Cameroon. J Environ Sci Wat Res 1(10): 243–250. [Google Scholar]
- 64. Danyluk MD, Nozawa-Inoue M, Hristova KR, Scow KM, Lampinen B, Harris LJ (2008) Survival and growth of Salmonella Enteritidis PT 30 in almond orchard soils. Appl Environ Microbiol 104: 1391–1399. 10.1111/j.1365-2672.2007.03662.x [DOI] [PubMed] [Google Scholar]
- 65. Duke JR, White JD, Allen PM, Muttiah RS (2007) Riparian influence on hyporheic-zone formation downstream of a small dam in the Blackland Prairie region of Texas. Hydrol Process 21(2): 141–150. 10.1002/hyp.6228 [DOI] [Google Scholar]
- 66. Berman GC, Pole CH (2000) Pathways of Human Influence on Water Temperature Dynamics in Stream Channels. Environ Manage 1: 1–20. [Google Scholar]
- 67. White DS, Elzinga CH, Hendricks SP (1987) Temperature patterns within the hyporheic zone of a northern Michigan River. J North Am Benthol Soc 6: 85–91. [Google Scholar]
- 68. Pusch M., Schwoerbel J (1994) Community respiration in hyporheic sediments of a mountain stream (Steina, Black Forest). Arch Hydrobiol 130(1): 35–52. [Google Scholar]
- 69. Leichtfried M (1988) Bacterial substrate in gravel beds of a second order alpine stream (Project Ritrodat-Lunz, Austria). Verh Internat Verein Limnol 23: 1325–1332. [Google Scholar]
- 70. Leichtfried M (1991) POM in bed sediments of a gravel stream (Ritrodat-Lunz study área, Austria). Verh Internat Verein Limnol 24: 1921–1925. [Google Scholar]
- 71. Anderson JK, Wondzell SM, Gooseff MN, Haggerty R (2005) Patterns in stream longitudinal profiles and implications for hyporheic exchange flow at the H.J. Andrews Experimental Forest, Oregon, USA. Hydrol Process 19(15): 2931–2949. 10.1002/hyp.5791 [DOI] [Google Scholar]
- 72. White DS (1990) Biological relationships to convective flow patterns within stream beds. Hydrobiologia 196: 149–158. [Google Scholar]
- 73. Brunke M, Gonser T (1997) The ecological significance of exchange processes between rivers and groundwater. Freshwater Biol 37(1): 1–33. 9142434 [Google Scholar]
- 74. Franken RJM, Storey RG, Williams DD (2001) Biological, chemical and physical characteristics of downwelling and upwelling zones in the hyporheic zone of a north-temperate stream. Hydrobiologia 444(1–3): 183–195. [Google Scholar]
- 75. Gibbons SM., Jones E 3, Bearquiver A, Blackwolf F, Roundstone W, Scott N, Hooker J, Madsen R, Coleman ML, Gilbert JA. 2014. Human and Environmental Impacts on River Sediment Microbial Communities. PLoS One. 9(5): e97435 10.1371/journal.pone.0097435 [DOI] [PMC free article] [PubMed] [Google Scholar]