

Abstract
Endogenous neural stem cells in central canal of adult mammalian spinal cord exhibit stem cell properties following injury. In the present study, the endogenous neural stem cells were labeled with Dil to track the differentiation of cells after mild spinal cord injury (SCI). Compared with 1 and 14 days post mild injury, the number of endogenous neural stem cells significantly increased at the injured site of spinal cord on 3 and 7 days post-injury. Dil-labeled βIII-tublin and GFAP expressing cells could be detected on 7 days post-injury, which indicated that the endogenous neural stem cells in central canal of spinal cord differentiated into different type of neural cells, but there were more differentiated astrocytes than the neurons after injury. Furthermore, after injury the expression of inhibitory Notch1 and Hes1 mRNA began to increase at 6 hours and was evident at 12 and 24 hours, which maintained high levels up to 7 days post-injury. These results indicated that a mild SCI in rat is sufficient to induce endogenous neural stem cells proliferation and differentiation. However, the ability to differentiate into neurons is limited, which may be, at least in part, due to high expression of inhibitory Notch1 and Hes1 genes after injury.
Keywords: Endogenous neural stem cells, proliferation, differentiation, spinal cord injury
Introduction
Spinal cord injury (SCI) usually leads to lose functions distal to the injured lesion due to axonal damage, demyelination and loss of neurons [1,2]. Currently, the majority of developing treatments for SCI focus on the use of stem cells to regenerate injured different phenotype of neural cells. Neural stem cells (NSCs) are multi-potent cells that can give rise to neurons, oligodendrocytes and astrocytes. However, the exogenous NSCs involve many challenges such as the source of NSCs, oncogenicity and the integration of transplanted NSCs with the host [3-5].
It was reported that ependymal stem cells (EpSCs) in central canal of spinal cord are endogenous neural stem cells (NSCs) found in the adult tissue [6,7]. Proliferation of ependymal stem cells is common during embryonic and early postnatal periods of development in most species, but they turnover declines significantly after birth [8,9]. During this period, series of genes regulating the proliferation and differentiation of NSCs changed accordingly. Therefore activating endogenous NSCs through regulating special genes in adult spinal cord to repair the injury might be a promising way to treat SCI.
However, whether the endogenous NSCs can proliferate or differentiate into special neural cell phenotype after mild SCI and the changes of different genes expression are not clear yet. In the present study, the endogenous NSCs in central canal of spinal cord were labeled and the proliferation, differentiation and expression of related genes after injury were examined.
Materials and methods
Animals
Forty adult female Wistar rats weighing 220-250 g were applied in the study. All animal experiments were approved by the Third Military Medical University Committee on Ethics for the Care and Use of Laboratory Animals.
Labeling of ependymal cells of spinal cord
To label the endogenous NSCs of adult rat spinal cord, the rats were anesthetization with sodium pentobarbital (40 mg/kg, i.p) and 10 ul of a 0.2% (w/v) solution of 1,1-dioctadecyl-6,6-di(4-sulfophenyl)-3,3,3,3-tetramethylindocarbocyanine (Dil, Sigma, USA) in dimethylsulfoxide was stereotactically injected into the right lateral ventricle 24 h prior to injury through a burr hole. The injecting location was 0.9 mm posterior and 2.5 mm lateral to bregma and 3.5 mm below the dura mater.
Mild spinal cord injury model
Twenty-four hours after Dil injection, the rats were re-anesthetized as described above. A 2-cm longitudinal skin incision was centered over the T10 spinous process along the midline. Para-spinal muscles and ligaments were laterally dissected and retracted, then followed by removal of bony elements of the posterior spine using a microrongeur. Without disrupting the dura mater, the tenth thoracic (T10) spinal segment was exposed by removing the dorsal part of the vertebra. The exposed spinal cord was next impacted by a 10 g weight dropped from a height of 10 mm. The impacted injury was induced by the weight-drop device presented by Wise Young (Hong Kong University). This type of injury leads to a mild lesion in the spinal cord. After injury, the muscles were sutured in layers and the skin was closed. The rats were returned to their cages with water and food.
Pathological staining and immunohistochemical examination of injured spinal cord tissues
Twenty rats were killed on 1, 3, 7, 14 days after mild injury. The experimental rats were deeply anesthetized with i.p. injection of sodium pentobarbital (40 mg/kg) and perfused with 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS). A segment of spinal cord tissue encompassing the injured site was fixed in 4% paraformaldehyde for at least 6 hours and then embedded in paraffin. After that the tissues were cut transversely into 5 um serial sections to stain with hematoxylin and eosin (HE) for general morphology examination.
For examining the Dil staining, tissues were cryoprotected through 30% sucrose in 0.1 M PBS for at least 24 h and a segment of the spinal cord 1.0 cm in length encompassing the injured site was removed and cryosectioned transversely into 20 um serial sections. The Dil signal was detected immediately with laser confocal microscrope.
For immunofluorescent staining, the following antibodies were applied on cryosectioned sections: rabbit anti-rat glial fibrillary acidic protein (GFAP; 1:400; Sigma, USA) detecting for astrocytes, mouse anti-rat βIII-tublin (1:800; Sigma, USA) for neurons. All staining procedures were performed in the dark. Sections were rehydrated in 0.1 M PBS and blocked with 10% (v/v) normal goat serum in 0.1 M PBS for 20 min. After that the sections were incubated with special primary antibodies overnight at 4°C and washed for 15 min. and then were incubated with fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse or goat anti-rabbit secondary antibodies (1:100; Chemicon, MA, USA) for 1 h at 37°C. At last the sections were counterstained with 300 nM of 4-6-diamidino-2-phenylindole dihydrochloride (DAPI, Sigma-Aldrich, USA) for 3 min. The positive signals were scanned with a Leica confocal microscope (SP-2, Leica, Germany). For negative controls, the sections were processed by the same immunofluorescent staining technique, but with omission of the primary antibodies. Immunohistochemical data shown in this manuscript are representative of three independent experiments from at least three animals in different time point.
Quantitative analysis of endogenous NSCs proliferation after SCI
The proliferation of endogenous NSCs in central canal of spinal cord was observed by examining DAPI stained sections. The number of endogenous NSC with DAPI-labeled nuclei was calculated by Leica SP-2 confocal microscope. The average number of endogenous NSCs on different time points was determined in five animals of each group. All the data were collected and evaluated blindly.
Semi-quantitative RT-PCR assay
For spinal tissue preparation, rats were sacrificed under anesthesia with sodium pentobarbital (45 mg/kg) at 6, 12, 24, 72 hours and 7 days after injury. A segment of 0.8 cm spinal cord in length encompassing the injury site was removed and immediately frozen in liquid nitrogen. Total RNA of the tissues was extracted using Trizol (Roche) at different time points after injury. RNA concentration and purity were determined by measuring the absorbance at 260 and 280 nm in a microtiter plate reader (Thermo, USA). RT was performed in a 20 µl reaction containing RNA 4 µl, OligodT (Takara) 1 µl, DEPC water 4 µl, at 65°C for 10 min and on ice for 5 min; moreover, added RNAase inhibitor 0.5 µl, 5× buffer 4 µl, 10 mM dNTP 2 µl, AMV (Takara) 1.5 µl and DEPC water 3 µl, at 42°C for 90 min in PCR machine. The 25 µl PCR reaction additionally contained the following components: 1× SYBR Green buffer, 50 pM of each primer, 0.2 mM dNTP mixture, and 2.5 U Tag plus DNA polymerase (Takara). The PCRs were conducted in a programmable thermocycler using an initial denaturing temperature of 94°C for 2 min, 35 cycles of 94°C for 1 min, 55°C for 1 min, 72°C for 1 min. The sequence of primers was as follows [10]:
Hes1: Forward primer: 5’-CTACCCCAGCCAGTGTCAAC-3’; Reverse primer: 5’-AAGCGGGTCACCTCGTTCAT-3’, 315 bp; Notch1: Forward primer: 5’-CTCACGCTGATGTCAATGCT-3’; Reverse primer: 5’-GTGGGAGACAGAGTGGGTGT-3’, 364 bp; GAPDH: Forward primer: 5’-ACCACAGTCCATGCCATCAC-3’; Reverse primer: 5’-TCCACCACCCTGRRGCTGTA-3’, 452 bp.
Statistical analysis
The data were analyzed with Origin75 software and presented as mean ± S.D. Statistical differences between groups were evaluated with one way analysis of variance. P value <0.05 was considered statistically significant.
Results
Pathological changes of injured spinal cord tissue and Dil labeled endogenous NSCs located in central canal after mild SCI
The mild SCI model was built by Allen’s weight-drop equipment. It was indicated that one day post-injury, the hemorrhagic focus could be found in the dorsal side of spinal cord, but the central canal of spinal cord was intact (Figure 1B). Three days after injury, the inflammatory cells were accumulated in injured lesion and the astrocytes around the lesion became active, which exhibited characteristic responses to trauma, such as the number of which increased and the body and dendrite of astrcocytes become thick and wide (Figure 1C). Seven days after injury, the cavity was formed in the injured lesion and astrocytes surrounding the cavity formed glial scars (Figure 1D).
Figure 1.
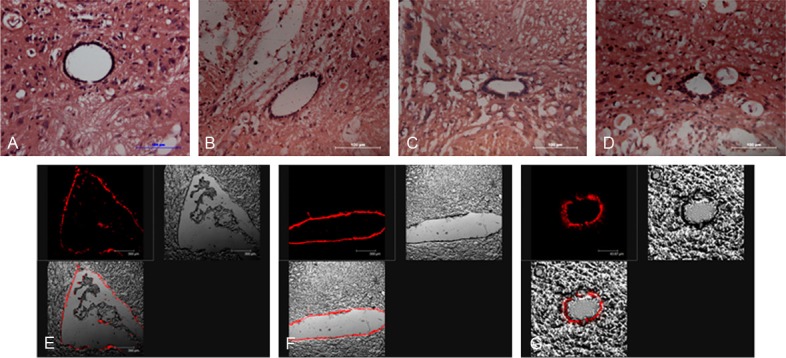
HE staining of mild spinal cord injury at different time post-injury and Dil labeling the endogenous neural stem cells in ependymal regions 24 after injection. A. The histological morphology of normal spinal cord with HE staining. B. The histological changes of spinal cord tissue at 1d post-injury with HE staining. C. The histological changes of spinal cord tissue at 3d post-injury. D. The histological changes of spinal cord tissue at 7d post-injury. E. Dil labeling the lining ependymal cells of ventricular. F. Dil labeling the lining ependymal cells of the fourth ventricle. G. Dil labeling the cells of central canal of spinal cord at T10 level. Scar bars = 100 um in A-D, E and F = 40 um.
Dil injected into the lateral ventricle was carried in the cerebrospinal fluid (CSF).The lining cells located in ventricular region of the lateral ventricles (Figure 1E), fourth ventricle (Figure 1F) were positively stained. After 24 hours, Dil through intra-ventricular injection labeled the endogenous NSCs lining the central canal of the spinal cord, as shown in Figure 1G. In vivo, the membranous labeling initially observed with Dil is largely replaced by a granular cytoplasmic labeling as the tracer becomes internalized by endocytosis.
Endogenous NSCs located in central canal appeared proliferation in response to mild SCI
The proliferative activity of endogenous NSCs of spinal cord after injury was examined via DAPI staining as shown in Figure 2, which demonstrated that a monolayer endogenous NSCs were lining the central canals 1 day after injury (Figure 2A). However, the endogenous NSCs exhibited remarkable proliferating ability 3 days post injury, which maintained to 7 days (Figure 2B and 2C). 14 days after injury, the number of endogenous NSCs lining the central canals again restored a monolayer condition (Figure 2D). As indicated in the bar graph of Figure 2, the number of proliferated endogenous NSCs on 3 and 7d post-injury was more than those on 1d and 14d after injury, which had significant difference in statistics.
Figure 2.
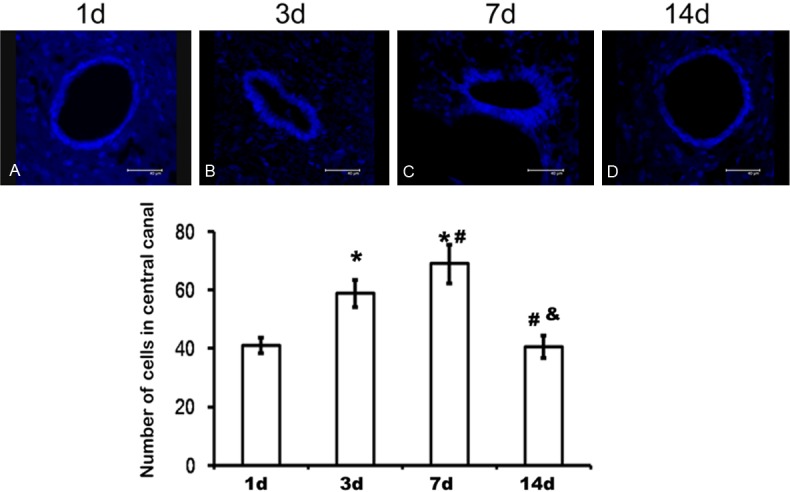
Proliferation of endogenous neural stem cells in central canal of T10 level after injury. A. DAPI labeling endogenous neural stem cells of spinal cord 1d after injury. B. DAPI labeling endogenous neural stem cells of spinal cord 3d after injury. C. DAPI labeling endogenous neural stem cells of spinal cord 7d after injury. D. DAPI labeling endogenous neural stem cells of spinal cord 14d after injury. *P<0.05 indicated statistical significance compared with 1d post-injury. #P<0.05 indicated statistical significance compared with 3d post-injury. &P<0.05 indicated statistical significance compared with 7d post-injury. Data represent mean ± SD of five independent samples in each group.
Endogenous NSCs in central canal of spinal cord differentiated into neurons and astrocytes after mild SCI
Interestingly, on 3 days post mild injury, clusters of Dil-labeled endogenous NSCs surrounding the region of the central canal were apparent at the level of the injury site (Figure 3A), indicating that a part of endogenous NSCs were migrating from the region of central canal. Antibodies against GFAP and βIII-tublin were used to identify astrocyte and neuron differentiated from Dil-labeled endogenous NSCs respectively. It was indicated that these neural cell markers are not expressed by endogenous NSCs lining the central canal in normal spinal cord. A few of Dil/βIII-tublin/DAPI- labeled cells were examined 7 days post-injury (as shown in Figure 3), which indicated that proliferated Dil-labeled endogenous NSCs can differentiate into neurons after mild spinal cord injury, but the number is limited. At the same time, Dil/GFAP/DAPI-labeled astrocytes were also detected post-injury. The differentiated astrocytes from Dil-labeled endogenous NSCs were apparent at 7 days following mild injury, which was more than those of differentiated neurons from Dil-labeled endogenous NSCs in spinal cord.
Figure 3.
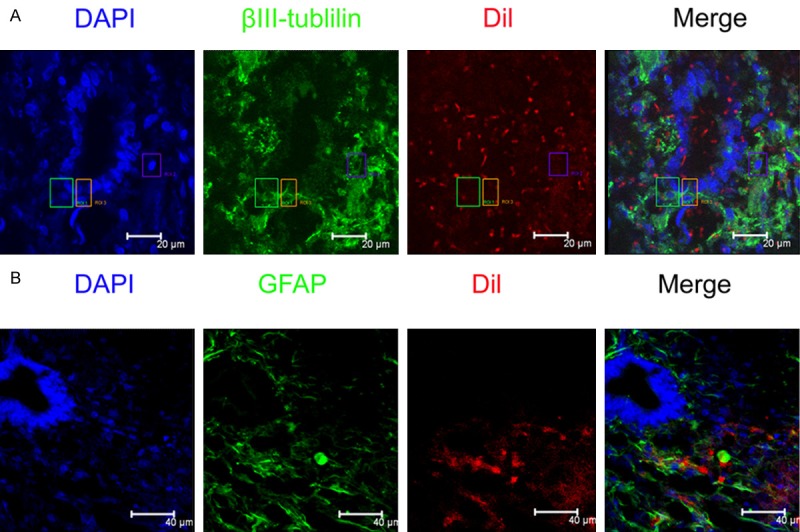
Identification of differentiated endogenous neural stem cells 7 days post-injury. A. Immunohistochemical staining of differentiated neurons from Dil-labeled endogenous neural stem cells. Red: Dil-labeled endogenous neural stem cells; Green: βIII-tublilin labeled differentiated neurons from endogenous neural stem cells; Blue: DAPI labeled nuclei. B. Immunohistochemical staining of differentiated astrocytes from Dil-labeled endogenous neural stem cells. Red: Dil-labeled endogenous neural stem cells; Green: GFAP labeled differentiated astocytes from endogenous neural stem cells; Blue: DAPI labeled neuclei.bar = 40 um.
Hes1 and Notch1 mRNA increased after spinal cord injury
RT-PCR examination of injured lesions in spinal cord indicated that inhibitory Hes1 and Notch1 mRNA began to increase at 6 hours, evident at 12 and 24 hours post-injury, which maintained the high levels up to 7 days post injury. As illustrated in Figure 4, there was significant difference compared to normal spinal cord tissues.
Figure 4.
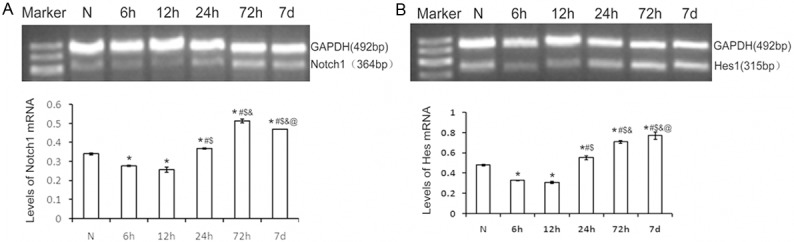
Expression of related genes after mild spinal cord injury. A. Expression of Notch1 gene at different times post-injury. B. Expression of Hes1 gene at different time post-injury.*P<0.05 indicated statistical significance compared with normal group. #P<0.05 indicated statistical significance compared with 6 h post-injury. $P<0.05 indicated statistical significance compared with 12 h post-injury. &P<0.05 indicated statistical significance compared with 24 h post-injury. @P<0.05 indicated statistical significance compared with 72 h post-injury. Data represent mean ± SE of three independent experiments.
Discussion
Although important advances have been made in treatment of SCI recently, it is not possible to restore neuronal functions entirely after SCI. In rodents and primates, SCI causes irreversible loss of function distal to the lesion as a result of axonal damage, demyelination and death of neural cells such as neurons, oligodendrocytes and astrocytes [2,3,11]. Replacing the lost cells and integrating newly transplanted or generated cells into the injured spinal cord lesions are key aims in designing potential therapies for patients that suffered from SCI. Because of the characteristics of self-renew and multiple differentiations potential, NSCs were regarded as a hopeful cell to enhance neuroprotection, stimulate neuro-plasticity, and repair neuronal loss in models of neural degenerative diseases or trauma, including SCI [9,10,19,20]. However, currently at least two major barriers need to be overcome before NSC grafts can be effectively used to treat CNS trauma: (1) the source of NSCs (2) the formation of glioma [9,10,20]. Therefore, activation the endogenous NSC in vivo of adult to repair the central nervous system injury will attract more and more attention of researchers.
Ependymal cells are remnants of the primitive neuro-epithelium from which NSCs were originated during development. In adult mammals, the ependymal cells in central canal exhibited remarkably limited proliferative activity in the normal spinal cord [8,12]. Moreover, the proliferation of ependymal cells in response to SCI has been reported. Subsequently, it was suggested that ependymal cells exhibit stem cell properties following injury, which were regard as endogenous NSCs remaining in adult spinal cord in mammals [13,14].
In order to track the endogenous NSCs and their progeny after SCI, the cell layer lining the central canal of spinal cord should be labeled with certain tracer. Dil is a lipophilic tracer that is incorporated irreversibly into the outer leaflets of the plasma membrane and diffuses laterally along the membrane [15]. In this experiment, Dil was injected into the lateral ventricle resulting in labeling of the ependymal cell layer specially. The lining of the central canal of spinal cord is composed primarily of endogenous NSCs, so we could track the migration of these cells and their progeny after SCI. At the same time the cells of Choroid plexus in the lateral ventricles could also have been labeled by Dil in the CSF.
In the present study, a mild SCI model with preservation of the integrity of central canal was established and which allowed us to examine the response of endogenous NSCs lining the central canal post-injury in greater detail. The proliferative activity of endogenous NSCs was assessed by comparing the DAPI labeling cells in central canal of injured spinal cord at different time after injury. It was indicated that the NSCs in central canal exhibited an active response after a mild weight-drop injury of spinal cord, where endogenous NSCs proliferated significantly 3 and 7 days post injury compared with 1d and 14d after injury. Furthermore, the proliferated endogenous NSCs could migrated from the central canal and differentiate into neurons through Dil/βIII-tublin immunocytochemical staining, but the number is fewer. However, Dil/GFAP positive cells were more than those of Dil/βIII-tublin positive cells after mild SCI, which indicated that the endogenous NSCs located in central canal had the tendency to differentiate into astrcoytes rather than neurons after spinal injury. But according to the immunohistochemical results, it was found that although the labeled endogenous NSCs migrated from the central canal, but they could arrive in white matter. In a minimal spinal cord injury model by Mothe et al [15], the proliferated ependymal cells can not differentiate into neurons and astrocytes, it may be that the injured degree is too minimal to evoke the repairing action of endogenous NSCs.
Previous studies have suggested that the behavior of NSCs was regulated by both intrinsic characteristics and extrinsic signals originating from the surrounding environment. It has been indicated that Notch, bHLH, Wnt and BMP signaling pathways were all involved in the regulation of NSCs whether in vitro or in vivo [16,17]. Among which Notch and bHLH genes determined the proliferation and differentiating directions of NSC respectively, while Wnt and BMP genes determine the subtype of neurons. Among inhibitory bHLH genes, such as Hes and Hiary, may induce NSCs differentiating into astrocytes, but active bHLH genes, such as Ngn and Olig genes, may induce NSCs differentiating into neurons. For example, transfected with Ngn2 and oligo2 had been shown to promote NSCs differentiating into motor neurons and oligodendrocytes respectively [18,19]. In this experiment, it was observed that the expression of Notch1 and Hes1 increased significantly after SCI and lasted to 7 days post-injury, which was of significant difference compared with the normal control by RT-PCR examination. It was reported that high expression of Notch1 may inhibited the differentiation of NSCS and Hes1 may promote NSCs differentiating into astrocytes. So the limited ability of endogenous NSCs differentiating into neurons may be due to the high expression of inhibitory Notch1 and Hes1 genes after mild spinal cord injury.
The intrinsic potential of endogenous NSCs to replace some of the cells in the spinal cord following injury opens up the opportunity for developing non-invasive therapies for patients with SCI [20,21]. It would be interesting to examine the function of proliferation-related genes to increase the proliferative capacity of endogenous NSCs and also to examine regulation-specific genes to evoke endogenous NSCs differentiating into specific neural cell types for endogenous repair after SCI [22,23].
Acknowledgements
This work was supported by grants of the National Natural Science Foundation of China (No. 81071599).
Disclosure of conflict of interest
None.
References
- 1.Fanie BH, Jonas F. Stem cells for spinal cord repair. Cell Stem Cell. 2008;3:16–24. doi: 10.1016/j.stem.2008.06.011. [DOI] [PubMed] [Google Scholar]
- 2.Liu Y, Wang L, Long ZY, Zeng L, Wu YM. Protoplasmic Astrocytes enhance the ability of neural stem cells to differentiate into neurons in vitro. PLoS One. 2012;7:e38243. doi: 10.1371/journal.pone.0038243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Moyse E, Segura S, Liard O, Mahaut S, Mechawar N. Microenvironmental determinants of adult neural stem cell proliferation and lineage commitment in the healthy and injured central nervous system. Curr Stem Cell Res Ther. 2008;3:163–184. doi: 10.2174/157488808785740334. [DOI] [PubMed] [Google Scholar]
- 4.Liu Y, Liu RR, Wang L, Zeng L, Long ZY, Wu YM. The effects of different phenotype astrocytes on neural stem cells differentiation in co-culture. Neurosci Lett. 2012;50:61–66. doi: 10.1016/j.neulet.2011.12.019. [DOI] [PubMed] [Google Scholar]
- 5.Kozlova EN. Differentiation and migration of astrocytes in the spinal cord following dorsal root injury in the adult rat. Eur J Neurosci. 2003;17:782–790. doi: 10.1046/j.1460-9568.2003.02518.x. [DOI] [PubMed] [Google Scholar]
- 6.Kojima A, Tator CH. Intrathecal administration of epidermal growth factor and fibroblast growth factor 2 promotes ependymal proliferation and functional recovery after spinal cord injury in adult rats. J Neurotrauma. 2002;19:223–238. doi: 10.1089/08977150252806974. [DOI] [PubMed] [Google Scholar]
- 7.Horner PJ, Power AE, Kempermann G, Kuhn HG, Palmer TD, Winkler J, Thal LJ, Gage FH. Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J Neurosci. 2000;20:2218–2228. doi: 10.1523/JNEUROSCI.20-06-02218.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Johansson CB, Momma S, Clarke DL, Risling M, Lendahl U, Frisen J. Identification of a neural stem cell in the adult mammalian central nervous system. Cell. 1999;96:25–34. doi: 10.1016/s0092-8674(00)80956-3. [DOI] [PubMed] [Google Scholar]
- 9.Kojima A, Tator CH. Epidermal growth factor and fibroblast growth factor 2 cause proliferation of ependymal precursor cells in the adult rat spinal cord in vivo. J Neuropathol Exp Neurol. 2000;59:687–697. doi: 10.1093/jnen/59.8.687. [DOI] [PubMed] [Google Scholar]
- 10.Paul M, Patrik B. Harnessing endogenous stem cells to treat neurodegenerative disorders of the basal ganglia. Parkinsonism and Related Disorders. 2004;10:259–264. doi: 10.1016/j.parkreldis.2004.02.013. [DOI] [PubMed] [Google Scholar]
- 11.Zhang RL, Zhang ZG, Chopp M. Ischemic stroke and neurogenesis in the subventricular zone. Neuropharmacology. 2008;55:345–352. doi: 10.1016/j.neuropharm.2008.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Foret A, Quertainmont R, Botman O, Bouhy D, Amabili P, Brook G, Schoenen J, Franzen R. Stem cells in the adult rat spinal cord: plasticity after injury and treadmill training exercise. J Neurochem. 2010;12:762–772. doi: 10.1111/j.1471-4159.2009.06500.x. [DOI] [PubMed] [Google Scholar]
- 13.Reimer MM, Kuscha V, Wyatt C, Sensen I, Erank FR, Knüwer M, Becker T, Becker CG. Sonic hedgehog is a polarized signal for motorneuron regeneration in adult zebrafish. J Neurosci. 2009;29:15073–15082. doi: 10.1523/JNEUROSCI.4748-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ronaghi M, Erceg S, Moreno-Manzano V, Stojkovic M. Challenges of stem cell therapy for spinal cord injury: human embryonic stem cells, endogenous neural stem cells, or induced pluripotent stem cells? Stem Cells. 2010;28:93–99. doi: 10.1002/stem.253. [DOI] [PubMed] [Google Scholar]
- 15.Mothe AJ, Tator CH. Proliferation, migration, and differentiation of endogenous ependymal region stem/progenitor cells following minimal spinal cord injury in the adult rat. Neuroscience. 2005;131:177–187. doi: 10.1016/j.neuroscience.2004.10.011. [DOI] [PubMed] [Google Scholar]
- 16.Zhang N, Wimmer J, Qian SJ, Chen WS. Stem Cells: current approach and future prospects in spinal cord injury repair. Anat Rec. 2010;293:519–530. doi: 10.1002/ar.21025. [DOI] [PubMed] [Google Scholar]
- 17.Moreno-Manzano V, Rodriguez-Jimenez FJ, Garcia-Rosello M, Laínez S, Erceg S, Calv MT, Ronaghi M, Lloret M, Planells-Cases R, Sánchez- Puelles JM. Activated spinal cord ependymal stem cells rescue neurological function. Stem Cells. 2009;27:733–743. doi: 10.1002/stem.24. [DOI] [PubMed] [Google Scholar]
- 18.Barreiro-Iglesias A. Targeting ependymal stem cells in vivo as a non-invasive therapy for spinal cord injury. Dis Model Mech. 2010;3:667–668. doi: 10.1242/dmm.006643. [DOI] [PubMed] [Google Scholar]
- 19.Peljto M, Dasen JS, Mazzoni EO, Jessell TM, Wichterle H. Functional Diversity of ESC-Derived motor neuron subtypes revealed through intraspinal transplantation. Cell Stem Cell. 2010;7:355–366. doi: 10.1016/j.stem.2010.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cheng XX, Wang YP, He Q, Qiu MS, Whittemore SR, Cao QL. Bone Morphogenetic Protein Signaling and Olig1/2 Interact to Regulate the Differentiation and Maturation of Adult Oligodendrocyte Precursor Cells. Stem Cell. 2007;25:3204–3214. doi: 10.1634/stemcells.2007-0284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Solecki DJ, Liu XL, Tomoda T, Fang Y, Hatten ME. Activated Notch2 signaling inhibits differentiation of cerebellar granule neuron precursors by maintaining proliferation. Neuron. 2001;31:557–568. doi: 10.1016/s0896-6273(01)00395-6. [DOI] [PubMed] [Google Scholar]
- 22.Doetsch F. A niche for adult neural stem cells. Curr Opin Genet Dev. 2003;13:543–550. doi: 10.1016/j.gde.2003.08.012. [DOI] [PubMed] [Google Scholar]
- 23.Oliver TG, Grasfeder LL, Carroll AL, Kaiser C, Gillingham CL, Lin SM, Wickramasinghe R, Scott MP, Wechsler-Reya RJ. Transcriptional profiling of the sonic hedgehog response: a critical role for N-myc in proliferation of neuronal precursors. PNAS. 2003;100:7331–7336. doi: 10.1073/pnas.0832317100. [DOI] [PMC free article] [PubMed] [Google Scholar]