

Abstract
Introduction: Increasing evidence suggested that microRNAs (miRNAs) play a critical role in tumorigenesis. Decreased expression of miRNA-139-5p has been observed in various types of cancers. However, the biological function of miRNA-139-5p in non-small cell lung cancer (NSCLC) is still largely unknown. Methods: Quantitative real-time PCR (qRT-PCR) was used to explore the expression level of miRNA-139-5p in NSCLC tissues and cell lines. Then, we investigated the role of miRNA-139-5p to determine its potential roles on lung cancer cell proliferation, migration and invasion in vitro. A luciferase reporter assay was performed to confirm the target gene of miRNA-139-5p and the results were validated in renal cancer cells. Results: miRNA-139-5p was significantly decreased in NSCLC tissues and cell lines. Over-expression of miRNA-139-5p could inhibit lung cancer cell proliferation, migration, and invasion in vitro. Furthermore, we identified insulin-like growth factor 1 receptor (IGF1R) as a target of miR-139-5p and miR-139-5p function as a tumor suppressor via targeting IGF1R in NSCLC. Conclusions: Our results indicated that miR-139-5p acts as a tumor suppressor in NSCLC partially via down-regulating IGF1R expression.
Keywords: Non-small cell lung cancer, miR-139-5p, insulin-like growth factor 1 receptor
Introduction
Lung cancer is the most common cause of cancer-associated deaths worldwide, with increasing incidence and mortality rates [1]. Among all forms of lung cancer, non-small cell lung cancer (NSCLC) accounts for nearly 85% of the cases [2]. Despite recent advances in clinical and experimental oncology, the prognosis of lung cancer is still unfavorable, with a 5-year overall survival rate of approximately 15% [3]. Thus, it is urgent to develop novel strategies and therapeutic targets for the treatment of human NSCLC.
MicroRNAs (miRNAs) are evolutionarily conserved, endogenous non-coding RNAs of about 22 nucleotides that post-transcriptionally regulate gene expression through binding to the 3’-untranslated region (3’-UTR) of target mRNAs in a sequence-specific manner [4]. miRNAs play crucial roles in various biological processes such as cell differentiation, proliferation, apoptosis, individual development, and metabolism [5]. Dysfunction of miRNAs occurred in many cancers and aberrantly altered expression of miRNAs was involved in tumorigenesis [6]. A variety of miRNAs have been found to dysregulated in NSCLC. For example, Liu et al showed that miRNA-196a promoted non-small cell lung cancer cell proliferation and invasion through targeting HOXA5 [7]. Mei et al demonstrated that miRNA-141 promoted the proliferation of non-small cell lung cancer cells by regulating expression of PHLPP1 and PHLPP2 [8]. Zhou et al suggested that miRNA-195 inhibited non-small cell lung cancer cell proliferation, migration and invasion by targeting MYB [9]. Ma et al found that miRNA-34a suppressed the proliferation and promoted the apoptosis of non-small cell lung cancer cells by targeting TGFβR2 [10]. These findings indicated that deregulation of miRNA expression may be associated with tumorigenesis of NSCLC.
Previous studies revealed that miR-139-5p was down-regulated in several types of human cancers. For example, Krishnan et al found that miR-139-5p was decreased in invasive breast cancer and could inhibit breast cancer cell invasion and migration [11]. Liu et al revealed that miR-139-5p was down-regulated expression in esophageal squamous cell carcinoma (ESCC) and associated with lymph node metastases. Furthermore, they suggested that miR-139-5p induced cell cycle arrest in the G0/G1 phase and to suppress the invasive capability of esophageal carcinoma cells by targeting the 3’UTR of oncogenic NR5A2 [12]. Song et al demonstrated that miR-139-5p suppressed migration and invasion of colorectal cancer by down-regulating AMFR and NOTCH1 expression [13]. However, the role of miR-139-5p in NSCLC cells remains unclear.
In the present study, we found that miR-139-5p was significantly down-regulated in NSCLC tissues and cell lines. Over-expression of miR-139-5p suppressed lung cancer cell proliferation, migration and invasion in vitro. Furthermore, insulin-like growth factor 1 receptor (IGF1R) was identified as a target of miR-139-5p in NSCLC cells and showed that miR-139-5p function as a tumor suppressor by silencing IGF1R expression. These results provided a potential therapeutic target for the treatment of NSCLC.
Materials and methods
Patients and specimens
A total of 28 paired NSCLC and adjacent non-tumor specimens were collected from the Nanjing Medical University Affiliated Wuxi Second Hospital. All tissue samples were flash-frozen in liquid nitrogen immediately after collection and stored at -80°C until use. The study protocol was approved by Nanjing Medical University Ethical Committee. Informed consent was obtained from all patients. Both tumor and non-tumor samples were confirmed by pathological examination. No patients received chemotherapy or radiotherapy prior to surgery.
Cell culture and cell transfection
Three NSCLC cell lines (A549, SK-MES-1, and H520) and a normal human bronchial epithelial cell line HBE were all purchased from the Institute of Biochemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China). All cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen) supplemented with 10% fetal bovine serum (FBS, Invitrogen), 100 μg/mL streptomycin, and 100 U/mL penicillin, in a tissue culture incubator under the atmosphere of 5% CO2 and 37°C.
The miR-139-5p mimics (miR-139-5p) and negative control miRNA mimics (miR-NC) were synthesized by RiboBio (China). Small interfering RNA against IGF1R (si-IGF1R) and negative control (si-NC) were designed by Genepharma (China). Transfection was carried out using Lipofectamine 2000 (Invitrogen) method. The final concentration was 200 nm for mimics and 100 nm for siRNA.
Cell proliferation assay
Cell proliferation was measured using the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide] assay. Transfected cells were seeded into 96-well plates with a density of 4000 cells/well, and cultured for different time. 10 μL of MTT was added into each well, and incubated at 37°C for 4 h. Then the supernatant was discarded, and 200 μL of DMSO was added to each well. Optical density (OD) was detected at the wavelength of 490 nm. Data were derived from three independent experiments.
Cell migration and invasion assays
Cell migration assay was performed using millicell chambers (Millipore). 24 h after transfection, 5×104 cells were seeded into serum-free medium on the upper chambers of an insert. Media containing 10% FBS were added to the lower chamber. The chamber was cultivated in 5% CO2 at 37°C for 24 h. Then, the cells in the upper chamber were removed with cotton wool, whereas the attached cells that had migrated or invaded into the lower section were stained with 0.1% crystal violet and counted in ten randomly selected fields under fluorescence microscope. The invasion assay was the same with migration assay except that matrigel (Sigma) was used in the transwell chambers and the cell suspension for the upper chambers were 1×105 cells.
Quantitative real-time PCR
Total RNA was extracted from tissues or cells using TRIzol Regent (Invitrogen) according to the manufacturer’s protocol. Reverse transcription was performed using MMLV RTase cDNA Synthesis Kit (TaKaRa). A cDNA library of miRNAs was constructed by QuantiMir cDNA Kit (TaKaRa). The level of mRNA or miRNA was measured by qRT-PCR using SYBR Green PCR master mixture (TaKaRa). The amplification conditions were 35 cycles of 12 s at 95°C and 1 min at 60°C. 18S RNA and U6 snRNA were used as the endogenous control for mRNA and miRNA expression. Fold change was calculated by relative quantification (2-ΔΔCt).
Western blot analysis
Cell lysates were prepared using a lysis buffer (Beyotime), separated by 10% SDS-PAGE and transferred to PVDF membranes (Millipore). After blocking, the membranes were incubated with primary anti-bodies overnight at 4°C, followed by further incubation with HRP-labeled secondary antibodies for 1 h at 37°C. The blots were developed using ECL system (Amersham Biosciences).
Luciferase reporter assay
To observe the binding of miR-139-5p to IGF1R mRNA, the 3’-UTR segment of IGF1R mRNA was amplified by PCR and inserted into the pGL3/luciferase vector (Promega). The mutant 3’-UTR of IGF1R mRNA was cloned using the wild type 3’-UTR as a template and inserted into pGL3/luciferase as described for the wild type 3’-UTR. Co-transfections of IGF1R 3’-UTR or mut IGF1R 3’-UTR plasmid with miR-139-5p mimics into the cells were accomplished by using Lipofectamine 2000 (Invitrogen). Luciferase activity was measured 48 h after transfection by the Dual-Luciferase Reporter Assay System (Promega). Data are presented as the mean value for triplicate experiments.
Statistical analysis
Statistical analyses were performed using SPSS version 18.0 for Windows (IBM). Data are presented as the mean ± SD from at least three independent experiments. Differences between samples were determined by one-way ANOVA and Student’s t-test, with P value less than 0.05 was considered to be statistically significant.
Results
miR-139-5p expression is down-regulated in human NSCLC tissues and cell lines
To assess the biological role of miR-139-5p in lung cancer carcinogenesis, the expression level of miR-139-5p was detected by qRT-PCR. As shown in Figure 1A, miR-139-5p was obviously down-regulated in NSCLC tissues compared to that in non-tumor tissues. Furthermore, miR-139-5p was also significantly reduced in NSCLC cell lines (A549, SK-MES-1, and H520) compared with the normal human bronchial epithelial cell line HBE, and it was the lowest in A549 cells (Figure 1B). These results indicated that miR-139-5p might be involved in human NSCLC progression.
Figure 1.
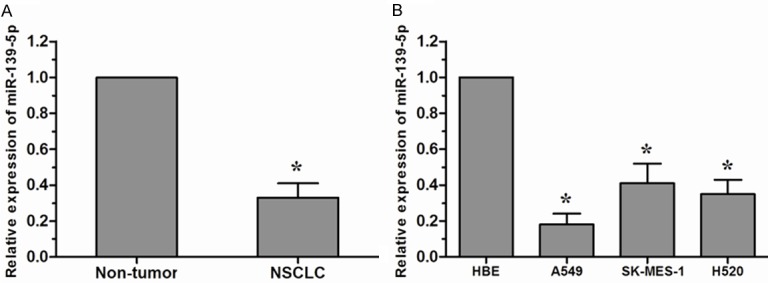
miR-139-5p was down-regulated in NSCLC tissues and cell lines. A. miR-139-5p was decreased in NSCLC tissues compared with the adjacent non-tumor tissues. B. miR-139-5p was down-regulated in NSCLC cell lines (A549, SK-MES-1, and H520) compared with the normal human bronchial epithelial cell line HBE. *P<0.05.
Effects of miR-139-5p on NSCLC cell proliferation, migration, and invasion
To assess the biological role of miR-139-5p in NSCLC, we transfected NSCLC cell line A549 cells with either miR-139-5p mimics (miR-139-5p) or negative control miRNA mimics (miR-NC). qRT-PCR assay was performed to detect the expression of miR-139-5p in A549 cells (Figure 2A). The effect of miR-139-5p on the proliferation of A549 cells was detected by MTT assay. The results revealed that proliferation of A549 cells was significantly decreased in miR-139-5p mimics transfected cells compared with miR-NC group (Figure 2B). Furthermore, we investigated whether miR-139-5p could also inhibit migration and invasion of NSCLC cells. We found that enforced expression of miR-139-5p dramatically inhibit tumor cell migration in A549 cells compared with the miR-NC group (Figure 2C). Similarly, transwell invasion assay demonstrated that miR-139-5p markedly decreased the invasive capacity of A549 cells (Figure 2D). Taken together, these data suggested that miR-139-5p act as a tumor suppressor that can inhibit the proliferation, migration, and invasion of NSCLC cells in vitro.
Figure 2.
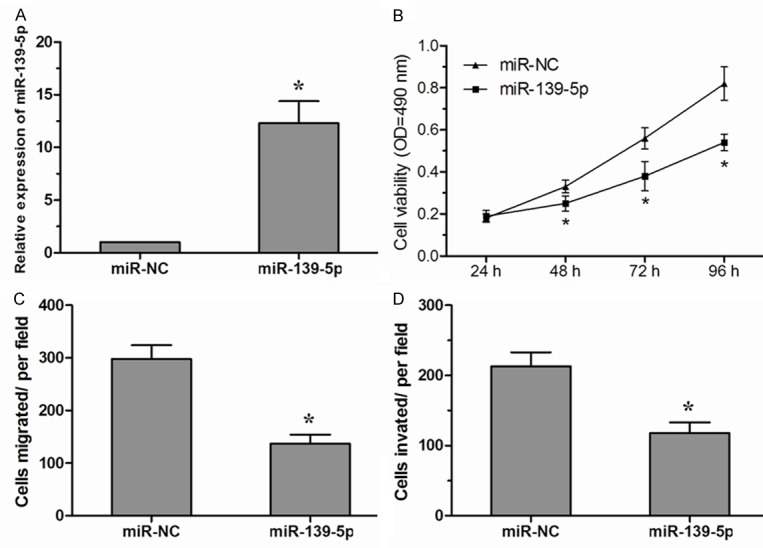
miR-139-5p inhibited NSCLC cell proliferation, migration and invasion. A. Expression of miR-139-5p in A549 cells transfected with miR-139-5p mimics or miR-NC. B. The vitality of A549 cells transfected with miR-139-5p mimics or miR-NC was detected using the MTT assay. C. In vitro transwell migration assay of A549 cells transfected with miR-139-5p mimics or miR-NC. D. In vitro transwell invasion assay of A549 cells transfected with miR-139-5p mimics or miR-NC. Data are presented as means ± SD from three independent experiments. *P<0.05.
IGF1R is a direct target of miR-139-5p
To identify the potential target genes of miR-139-5p, TargetScan and miRanda was used in combination. Our analysis revealed that IGF1R was a potential target of miR-139-5p based on putative target sequences at position 2486-2493 of the IGF1R 3’-UTR (Figure 3A). To confirm IGF1R as a direct target of miR-139-5p, we engineered luciferase reporter constructs containing the wild-type (WT) or mutant (Mut) 3’-UTR of the IGF1R gene. Luciferase reporter assay showed that miR-139-5p significantly decreased the luciferase activity of the IGF1R 3’UTR but not that of the mutant in A549 cells (Figure 3B). Western blot analyses showed that over-expression of miR-139-5p significantly decreased the expression of IGF1R in A549 cells (Figure 3C). Taken together, these results indicated that IGF1R is a direct target of miR-139-5p in NSCLC.
Figure 3.
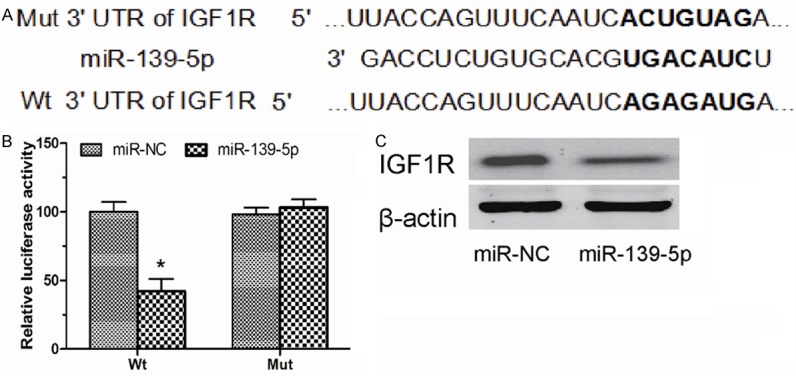
IGF1R was a target of miR-139-5p in NSCLC cells. A. A schematic illustration of Wt and Mut 3’-UTR of IGF1R. B. Luciferase activity assay shown that miR-139-5p suppressed Wt IGF1R 3’-UTR luciferase activity, while it had no effect on Mut IGF1R 3’-UTR luciferase activity compared to control in A549 cells. C. Expression of IGF1R was detected by Western blot in A549 cells transfected with miR-139-5p mimics or miR-NC. β-actin was used as a control. Data are presented as means ± SD from three independent experiments. *P<0.05.
Effect of IGF1R on NSCLC cells proliferation, migration and invasion
To determine whether IGF1R could also inhibit NSCLC cell growth and metastasis in vitro, we performed targeted knockdown of IGF1R expression using RNAi in A549 cells. The expression levels of IGF1R in A549 cells transfeced with si-IGF1R were significantly decreased compared with si-NC transfected cells (Figure 4A). MTT assay showed that A549 cells transfected with si-IGF1R displayed a significantly lower proliferation ability compared with cells transfected with si-NC (Figure 4B). Next, we performed transwell migration and invasion assays to investigate cell metastasis in vitro. Transwell assays revealed that inhibition of IGF1R suppressed NSCLC cell migration and invasion in vitro (Figure 4C, 4D). Taken together, these findings indicated that IGF1R is a functionally important target of miR-139-5p that is involved in the proliferation, migration and invasion of NSCLC cells.
Figure 4.
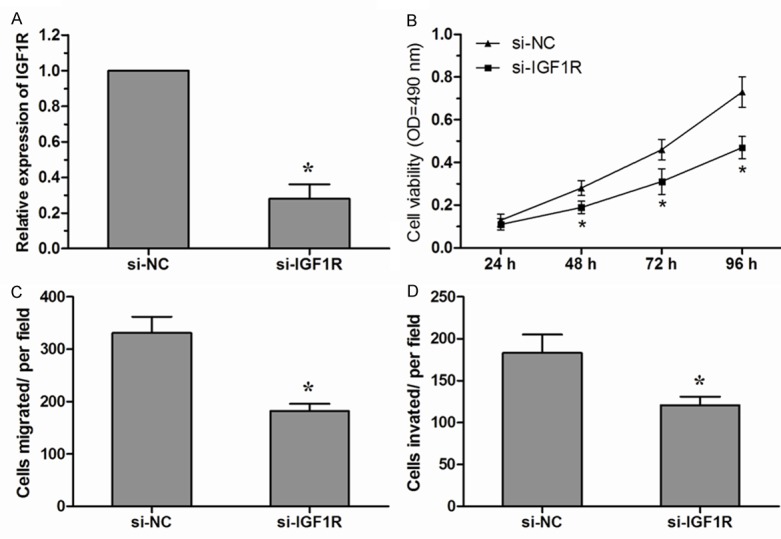
Decreased expression of IGF1R showed similar effect with miR-139-5p over-expression. (A) Knockdown of IGF1R by siRNA (si-IGF1R) was confirmed by qRT-PCR in A549 cells. (B) The vitality of A549 cells transfected with si-IGF1R or si-NC was detected using the MTT assay. (C, D) Transwell assays were used to detect migration (C) and invasion (D) of A549 cells transfected with si-IGF1R or si-NC. Data are presented as means ± SD from three independent experiments. *P<0.05.
Discussion
Understanding the molecular mechanisms of carcinogenesis and cancer progression is crucial for early diagnosis and effective therapy [14]. Aberrant expression of miRNA often occurs in human cancers and play critical roles in tumor progression [15]. Therefore, it is very important to explore the function of deregulated molecules in cancers. In the present study, we investigated the roles of miR-139-5p in tumor growth and metastasis of NSCLC cells in vitro. We found that miR-139-5p was dramatically down-regulated in NSCLC tissues and cell lines. In addition, enforced expression of miR-139-5p could suppress NSCLC cell proliferation, migration and invasion. These results suggested that miR-139-5p plays an important role in the development and progression of NSCLC.
We further investigated the molecular mechanisms whereby miR-139-5p inhibits the proliferation, migration and invasion of NSCLC cells. Insulin-like growth factor 1 receptor (IGF1R) is a tyrosine kinase receptor for IGF-1 and IGF-2 and is frequently increased in many types of cancers [16]. The IGF receptor family consists of three transmembrane proteins, and the IGF1R gene is located on chromosome 15q26, which encodes a single polypeptide of 1367 amino acids that is constitutively expressed in most cells [17]. IGF1R activation leads to autophosphorylation in the kinase domain, followed by recruitment of specific docking intermediates, such as IRS-1 and Shc proteins [18]. These molecules link the IGF1R to diverse signaling pathways, allowing the induction of growth, transformation, differentiation and protection against apoptosis [19]. Recently, amount of studies showed miRNAs could play an important role in the regulation of IGF1R. For example, Corcoran et al showed that miR-630 targets IGF1R to regulate response to HER-targeting drugs and overall cancer cell progression in HER2 over-expressing breast cancer [20]. Li et al found that miRNA-100/99a, deregulated in acute lymphoblastic leukaemia, suppress proliferation and promote apoptosis by regulating the FKBP51 and IGF1R/mTOR signaling pathways [21]. Zhang et al suggested that miRNA-503 acts as a tumor suppressor in glioblastoma for multiple antitumor effects by targeting IGF1R [22]. However, the role of IGF1R in NSCLC still largely unclear. In the present study, our results demonstrated that IGF1R act as a target of miR-139-5p and showed that miR-139-5p over-expression is correlated with IGF1R down-regulation leading to the inhibition of cell proliferation, migration, and invasion. Our data revealed that the tumor suppressor role of miR-139-5p is mediated by down-regulation of IGF1R expression.
In conclusion, we demonstrated that miR-139-5p is down-regulated in NSCLC. Over-expression of miR-139-5p could inhibit NSCLC cell proliferation, migration, and invasion by down-regulating IGF1R expression. These data suggested that miR-139-5p might act as a potential therapeutic target for NSCLC.
Acknowledgements
This study was supported by grants from National Natural Science Foundation of China (no. 81400161).
Disclosure of conflict of interest
None.
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Ettinger DS, Akerley W, Bepler G, Blum MG, Chang A, Cheney RT, Chirieac LR, D’Amico TA, Demmy TL, Ganti AK, Govindan R, Grannis FW Jr, Jahan T, Jahanzeb M, Johnson DH, Kessinger A, Komaki R, Kong FM, Kris MG, Krug LM, Le QT, Lennes IT, Martins R, O'Malley J, Osarogiagbon RU, Otterson GA, Patel JD, Pisters KM, Reckamp K, Riely GJ, Rohren E, Simon GR, Swanson SJ, Wood DE, Yang SC NCCN Non-Small Cell Lung Cancer Panel Members. Non-small cell lung cancer. J Natl Compr Cancer Netw. 2010;8:740–801. doi: 10.6004/jnccn.2010.0056. [DOI] [PubMed] [Google Scholar]
- 3.Molina JR, Yang P, Cassivi SD, Schild SE, Adjei AA. Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship. Mayo Clin Proc. 2008;83:584–594. doi: 10.4065/83.5.584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5:522–531. doi: 10.1038/nrg1379. [DOI] [PubMed] [Google Scholar]
- 5.Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2009;10:704–714. doi: 10.1038/nrg2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Liu XH, Lu KH, Wang KM, Sun M, Zhang EB, Yang JS, Yin DD, Liu ZI, Zhou J, Liu ZJ. MicroRNA-196a promotes non-small cell lung cancer cell proliferation and invasion through targeting HOXA5. BMC Cancer. 2012;12:348. doi: 10.1186/1471-2407-12-348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mei Z, He Y, Feng J, Shi J, Du Y, Qian L, Huang Q, Jie Z. MicroRNA-141 promotes the proliferation of non-small cell lung cancer cells by regulating expression of PHLPP1 and PHLPP2. FEBS Lett. 2014;588:3055–3061. doi: 10.1016/j.febslet.2014.06.020. [DOI] [PubMed] [Google Scholar]
- 9.Yongchun Z, Linwei T, Xicai W, Lianhua Y, Guangqiang Z, Ming Y, Guanjian L, Yujie L, Yunchao H. MicroRNA-195 inhibits non-small cell lung cancer cell proliferation, migration and invasion by targeting MYB. Cancer Lett. 2014;347:65–74. doi: 10.1016/j.canlet.2014.01.019. [DOI] [PubMed] [Google Scholar]
- 10.Ma ZL, Hou PP, Li YL, Wang DT, Yuan TW, Wei JL, Zhao BT, Lou JT, Zhao XT, Jin Y, Jin YX. MicroRNA-34a inhibits the proliferation and promotes the apoptosis of non-small cell lung cancer H1299 cell line by targeting TGFbetaR2. Tumour Biol. 2015;36:2481–90. doi: 10.1007/s13277-014-2861-5. [DOI] [PubMed] [Google Scholar]
- 11.Krishnan K, Steptoe AL, Martin HC, Pattabiraman DR, Nones K, Waddell N, Mariasegaram M, Simpson PT, Lakhani SR, Vlassov A. miR-139-5p is a regulator of metastatic pathways in breast cancer. RNA. 2013;19:1767–1780. doi: 10.1261/rna.042143.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu R, Yang M, Meng Y, Liao J, Sheng J, Pu Y, Yin L, Kim SJ. Tumor-suppressive function of miR-139-5p in esophageal squamous cell carcinoma. PLoS One. 2013;8:e77068. doi: 10.1371/journal.pone.0077068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Song M, Yin Y, Zhang J, Zhang B, Bian Z, Quan C, Zhou L, Hu Y, Wang Q, Ni S, Fei B, Wang W, Du X, Hua D, Huang Z. MiR-139-5p inhibits migration and invasion of colorectal cancer by downregulating AMFR and NOTCH1. Protein Cell. 2014;5:851–861. doi: 10.1007/s13238-014-0093-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nagpal JK, Das BR. Oral cancer: reviewing the present understanding of its molecular mechanism and exploring the future directions for its effective management. Oral Oncol. 2003;39:213–221. doi: 10.1016/s1368-8375(02)00162-8. [DOI] [PubMed] [Google Scholar]
- 15.Zhang B, Pan X, Cobb GP, Anderson TA. microRNAs as oncogenes and tumor suppressors. Dev Biol. 2007;302:1–12. doi: 10.1016/j.ydbio.2006.08.028. [DOI] [PubMed] [Google Scholar]
- 16.Larsson O, Girnita A, Girnita L. Role of insulinlike growth factor 1 receptor signalling in cancer. Br J Cancer. 2005;92:2097–2101. doi: 10.1038/sj.bjc.6602627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hartog H, Wesseling J, Boezen HM, van der Graaf WT. The insulin-like growth factor 1 receptor in cancer: old focus, new future. Eur J Cancer. 2007;43:1895–1904. doi: 10.1016/j.ejca.2007.05.021. [DOI] [PubMed] [Google Scholar]
- 18.Riedemann J, Macaulay VM. IGF1R signalling and its inhibition. Endocr Relat Cancer. 2006;13(Suppl 1):S33–43. doi: 10.1677/erc.1.01280. [DOI] [PubMed] [Google Scholar]
- 19.Sachdev D, Yee D. The IGF system and breast cancer. Endocr Relat Cancer. 2001;8:197–209. doi: 10.1677/erc.0.0080197. [DOI] [PubMed] [Google Scholar]
- 20.Corcoran C, Rani S, Breslin S, Gogarty M, Ghobrial IM, Crown J, O’Driscoll L. miR-630 targets IGF1R to regulate response to HER-targeting drugs and overall cancer cell progression in HER2 over-expressing breast cancer. Mol Cancer. 2014;13:71. doi: 10.1186/1476-4598-13-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li XJ, Luo XQ, Han BW, Duan FT, Wei PP, Chen YQ. MicroRNA-100 99a, deregulated in acute lymphoblastic leukaemia, suppress proliferation and promote apoptosis by regulating the FKBP51 and IGF1R/mTOR signalling pathways. Br J Cancer. 2013;109:2189–2198. doi: 10.1038/bjc.2013.562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang Y, Chen X, Lian H, Liu J, Zhou B, Han S, Peng B, Yin J, Liu W, He X. MicroRNA-503 acts as a tumor suppressor in glioblastoma for multiple antitumor effects by targeting IGF-1R. Oncol Rep. 2014;31:1445–1452. doi: 10.3892/or.2013.2951. [DOI] [PMC free article] [PubMed] [Google Scholar]