

Abstract
Glycogen synthase kinase 3β (GSK3β) and phosphorylated GSK3β at Ser9 (pS9GSK3β) are crucial in cellular proliferation and metabolism. GSK3β and pS9GSK3β are deregulated in many diseases including tumors. Data on altered expression of GSK3β and pS9GSK3β are mainly limited to tumor tissues, thus the expression of GSK3β and pS9GSK3β in normal human tissue has been largely unknown. Thus, we examined the immunohistochemical localization of GSK3β and pS9GSK3β in human fetal and adult tissues, and also compared the expression pattern of GSK3β and pS9GSK3β with that of the CK7 and CK20. We found GSK3β expression in neurons of brain, myenteric plexus in gastrointestinal tract, squamous epithelium of skin, and mammary gland. The expression of pS9GSK3β was restricted to the epithelial cells of breast and pancreaticobiliary duct, distal nephron of kidney, gastrointestinal tract, fallopian tube, epididymis, secretory cell of prostatic gland, and umbrella cell of urinary tract. The staining pattern of pS9GSK3β and CK7 was overlapped in most organs except for gastrointestinal tract where CK7 was negative and CK20 was positive. Our results show that the expression of GSK3β may be associated with differentiation of ectodermal derived tissues and pS9GSK3β with that of epithelial cells of endodermal derived tissues in human. In addition, the expression of pS9GSK3β in the selective epithelial cells may indicate its association with secretory or barrier function of specific cells and may serve as another immunohistochemical marker for epithelial cells.
Keywords: GSK3β, pS9GSK3β, keratin, normal tissues, immunohistochemical study
Introduction
Glycogen synthase kinase 3 (GSK3) is a serine (Ser)/threonine (Thr) kinase involved in multiple cellular processes, including proliferation, differentiation, and cell cycle regulation [1]. GSK3 exists as α and β isoforms, and GSK3β has lower molecular weight (47 kDa) than GSK3α (51 kDa) due to lack of glycine-rich N-terminal domain [1]. Their activity is inhibited by Ser/Thr phosphorylation (Ser21 in GSK3α and Ser9 and Thr390 in GSK3β) and activated by tyrosine phosphorylation (Tyr279 in GSK3α and Tyr216 in GSK3β) [1]. Of the two isoforms, GSK3β has been known to be associated with the development of variable diseases including cancer [2-4]. Main regulators of GSK3β activity are the phosphoinositide 3-kinase (PI3K)/Akt pathway and mitogen activated protein kinase-activated protein kinase 1 (MAPKAP-K1) of Ras/Raf/MEK/MAPK pathway [5]. Insulin-induced PI3K/Akt activation mediates Ser9 phosphorylation of GSK3β (pS9GSK3β), and pS9GSK3β leads to the dephosphorylation of glycogen synthase, resulting in increased glycogen synthesis [5]. pS9GSK3β promotes protein and lipid synthesis via activating the mTOR/S6K1 signaling pathway, whereas GSK3β inhibits cell growth by downregulating cyclin D1 [6]. GSK3β plays an inhibitory role in the Wnt/β-catenin pathway, which is associated with embryo development, epithelial repair, and tumor cell survival [1,2,7]. When Wnt signal is absent, GSK3β forms complex with axin and adenomatous polyposis coli (APC) and promotes the proteasomal degradation of β-catenin. In contrast, when Wnt binds to frizzled (Frz), GSK3β is inhibited by the activation of dishevelled (Dsh) and β-catenin accumulates in the cytoplasm, subsequently shifting to the nucleus. Nuclear β-catenin assembles with T cell factor/lymphoid enhancer factor (TCF/LEF), leading to transcription of specific target genes [8].
GSK3β has paradoxical role either as a tumor suppressor or as a tumor promoter [3]. GSK3β is a promoter of glioblastoma multiforme (GBM) by protecting the tumor cells from apoptosis via the inactivation of p53- and/or Rb-mediated pathways [3]. In GBM, breast and colon cancer patients, high level expression of GSK3β has been reported [2,4,9]. On the contrary, GSK3β function as a tumor suppressor in squamous cell carcinoma (SCC) of skin [10]. The overexpression of pS9GSK3β has been observed in adenocarcinoma of lung [11] and pancreas [12].
Although the expression level of GSK3β and pS9GSK3β in human tumors has been studied widely, their expression pattern in normal human tissue has been only sporadically reported and received a little attention. The comparison of distribution of GSK3β and pS9GSK3β in normal human tissues would provide better understanding of physiologic and functional role of these proteins. Therefore, we performed an immunohistochemical analysis of GSK3β and pS9GSK3β in normal human adult and fetal tissues, and also compared their expression with that of the intermediate filament protein cytokeratin (CK), CK7 and CK20.
Materials and methods
Tissue samples and arrays
The list of human tissues was obtained from tissue archives within the Department of Pathology at Eulji General Hospital, Eulji University School of Medicine. The tissues were collected with informed consent prior to each operation and the study was performed with the approval of the Institutional review board of Eulji General Hospital. Slides of normal adult tissues from surgical specimens and fetal tissues from autopsy were reviewed. Two fetuses were at 21 weeks and 38 weeks of gestation, respectively.
Representative areas of normal tissues were taken from the paraffin blocks and tissue microarray (TMA) was constructed as previously described [13]. In case of cores of TMA insufficient for representing the tissue, the sections of normal tissue samples were separately prepared.
Immunohistochemistry
Immunohistochemical staining was performed using DAKO Autostainer (DakoCytomation, Carpinteria, CA, USA). Four micron-thick tissue sections were obtained from TMA blocks and transferred onto poly-L-lysine coated slides. After deparaffinization and rehydration, antigen retrieval was performed using citrate buffer (pH 6.0) at 121°C for 10 minutes. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 5 minutes, and the sections were incubated with antibodies against GSK3β (BD Biosciences, Lexington, KY, 1:20), pS9GSK3β (Abcam, Cambridge, UK, 1:250), CK7 (Dako, Carpinteria, CA, 1:100) and CK20 (Dako, 1:50). Color was developed with diaminobenzidine, and the slides were counterstained with hematoxylin. The tissue section of GBM was used as a positive control for GSK3β and that of pancreatic adenocarcinoma was used as a positive control for pS9GSK3β. Cases omitted primary antibodies were served as negative control. The cytoplasmic and/or membranous expression of GSK3β, pS9GSK3β, CK7 and CK20 was approved as positive staining. The staining intensity with the number of positive cells was scored as: -, undetectable; ±, < 5% positive cells; 1+, mild intensity in most cells; 2+, moderate intensity in most cells and 3+, strong intensity in most cells.
Results
We found tissue-specific distribution of GSK3β and pS9GSK3β in normal human tissues and corresponding expression of pS9GSK3β and CK7 in epithelia of many organs. Tables 1 and 2 summarize the patterns of GSK3β, p pS9GSK3β, CK7, and CK20 expression in normal adult and fetal tissues, respectively.
Table 1.
Expression of GSK3β, pS9GSK3β, CK7 and CK20 in normal human adult tissues
| Organ | Cell type | GSK3β | pS9GSK3β | CK7 | CK20 |
|---|---|---|---|---|---|
| Brain | Neuron | +++ | - | - | - |
| Neuroglia | + | - | - | - | |
| Skin | Epidermis | ++ | ++ | - | - |
| Sweat gland | + | +++ | +++ | - | |
| Esophagus | Squamous epithelium | + | - | - | - |
| Stomach | Foveolar epithelium | - | ± | ± | ++ |
| Glands | - | ± | ± | - | |
| Ganglion (myenteric plexus) | +++ | - | - | - | |
| Nerve fiber (myenteric plexus) | ++ | - | - | - | |
| Bowel | Enterocyte | - | ++ | - | ++ |
| Liver | Hepatocyte | - | - | - | - |
| Bile duct | - | +++ | +++ | - | |
| Pancreas | Islet | - | - | - | - |
| Acinar | - | ± | - | - | |
| Centroacinar | - | +++ | +++ | - | |
| Duct | - | +++ | +++ | - | |
| Kidney | Bowman’s capsule | - | ± | ± | - |
| Proximal tubule | - | ± | ± | - | |
| Distal tubule | - | +++ | +++ | - | |
| Collecting duct | - | +++ | +++ | - | |
| Bladder | Urothelium (umbrella cell) | - | +++ | +++ | ++ |
| (basal to intermediate cell) | - | + | +++ | - | |
| Prostate | Gland (luminal cell) | - | +++ | ± | - |
| (basal cell) | - | - | ± | - | |
| Testis | Germ cell (seminiferous tubule) | - | - | - | - |
| Rete testis | - | ++ | +++ | - | |
| Epididymis | Pseudostratified epithelium | - | ++ | ± | - |
| Ovary | Surface epithelium | - | ++ | ++ | - |
| Follicular cell (primordial follicle) | - | ± | - | - | |
| Oocyte (primordial follicle) | - | - | ± | - | |
| Fallopian tube | Ciliated cell | - | - | - | - |
| Non-ciliated cell | - | ++ | +++ | - | |
| Uterus | Endometrial gland (pro*) | - | ++ | ± | - |
| Endometrial gland (sec**) | - | ± | ± | - | |
| Endocervix | - | - | ++ | - | |
| Exocervix | - | - | - | - | |
| Breast | Epithelial cell | + | +++ | +++ | - |
| Myoepithelial cell | - | - | - | - | |
| Lung | Bronchial epithelium | - | ± | ++ | - |
| Pneumocyte | - | - | ± | - | |
| Mesothelial cell | - | +++ | ++ | - | |
| Lymph node | Germinal center | + | - | - | - |
-, undetectable; ±, < 5% positive cells; +, mild intensity in most cells; ++, moderate intensity in most cells; +++, strong intensity in most cells,
proliferative phase;
secretory phase.
Table 2.
Expression of GSK3β, pS9GSK3β, CK7 and CK20 in normal human fetal tissues
| Organ | Cell type | GSK3β | pS9GSK3β | CK7 | CK20 |
|---|---|---|---|---|---|
| Stomach | Epithelium | - | + | ± | +++ |
| Bowel | Epithelium | - | +++ | - | ++ |
| Liver | Hepatocyte | - | - | - | - |
| Bile duct | - | +++ | +++ | - | |
| Pancreas | Islet | - | - | - | - |
| Acinar | - | ± | - | - | |
| Centroacinar | - | +++ | +++ | - | |
| Duct | - | +++ | +++ | - | |
| Kidney | Bowman’s capsule | - | - | - | - |
| Proximal tubule | - | - | - | - | |
| Distal tubule | - | ++ | ± | - | |
| Collecting duct | - | +++ | +++ | - | |
| bladder | Urothelium (umbrella cell) | - | +++ | +++ | +++ |
| (basal to intermediate cell) | - | - | +++ | - | |
| Lung | Bronchial epithelium | - | ± | ± | - |
| Pneumocyte | - | ++ | ++ | - |
-, undetectable; ±, < 5% positive cells; +, mild intensity in most cells; ++, moderate intensity in most cells; +++, strong intensity in most cells.
Expression of GSK3β in normal human tissues
GSK3β expression was found in brain, myenteric plexus in gastrointestinal tract, skin, mammary gland, and lymphoid tissues. Neurons in brain showed strong GSK3β immunoreactivity, while glial fibrils were weakly stained for GSK3β (Figure 1A). GSK3β expression was strong for ganglion cells of myenteric plexus and moderate for nerve sheath cells (Figure 1B). GSK3β was moderately expressed in squamous epithelial cells of skin (Figure 1C) and weakly stained in squamous mucosa of the esophagus, but not detected in uterine cervix. Sweat glands of skin, mammary gland, and germinal center cells of lymph node showed weak GSK3β staining.
Figure 1.
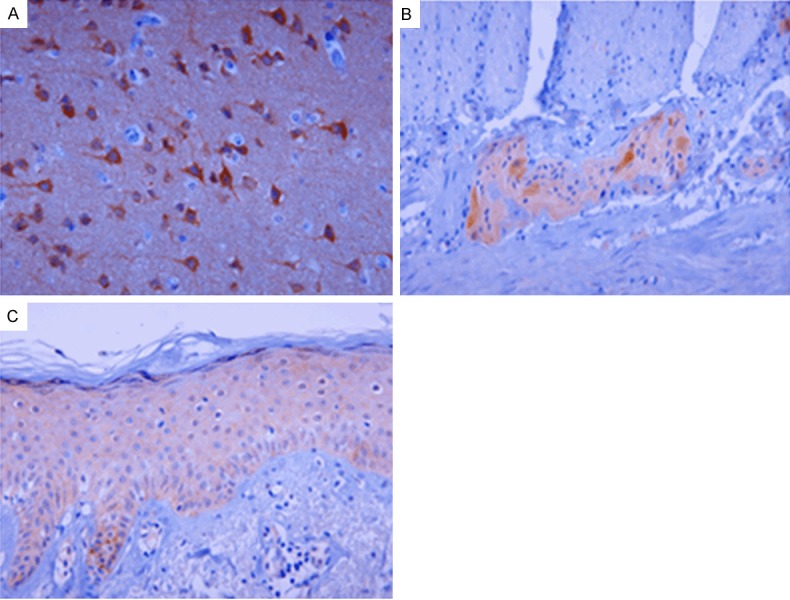
Immunoreactions of GSK3β in normal adult human tissues. GSK3β shows strong cytoplasmic staining in neurons and weak staining in glial fibrils in the brain (A). Cytoplasmic GSK3β staining is strong in ganglion cells and moderate in nerve sheath cells of myenteric plexus (B). GSK3β shows moderate perimembranous staining in squamous cells of the skin (C). Original magnifications: × 400.
Expression of pS9GSK3β in normal human tissues
The expression of pS9GSK3β was mainly restricted to the epithelial cells, which lined ductal structures within organs such as breast, prostate, pancreas, liver, kidney, bladder, fallopian tube, and epididymis. The luminal epithelial cells of the ducts and acini of the breast (Figure 2A) and prostate (Figure 2B) displayed prominent pS9GSK3β expression, whereas basal/myoepithelial layer cells were unreactive. In the pancreas (Figure 2C), the immunoreaction of pS9GSK3β was intense in ductal and centroacinar cells and occasionally weakly seen in acinar cells, but negative in Langerhans islets. In the liver, pS9GSK3β expression was only observed in bile duct cells. In the kidney (Figure 2D), pS9GSK3β expression was found strongly in distal tubules and collecting ducts, and occasionally in Bowman’s capsule and proximal tubules. In the bladder, pS9GSK3β expression was strong in urothelial superficial cells and weakly seen in basal to intermediate cells. Strong pS9GSK3β staining in umbrella cells was accentuated in fetal bladder (Figure 2E).
Figure 2.
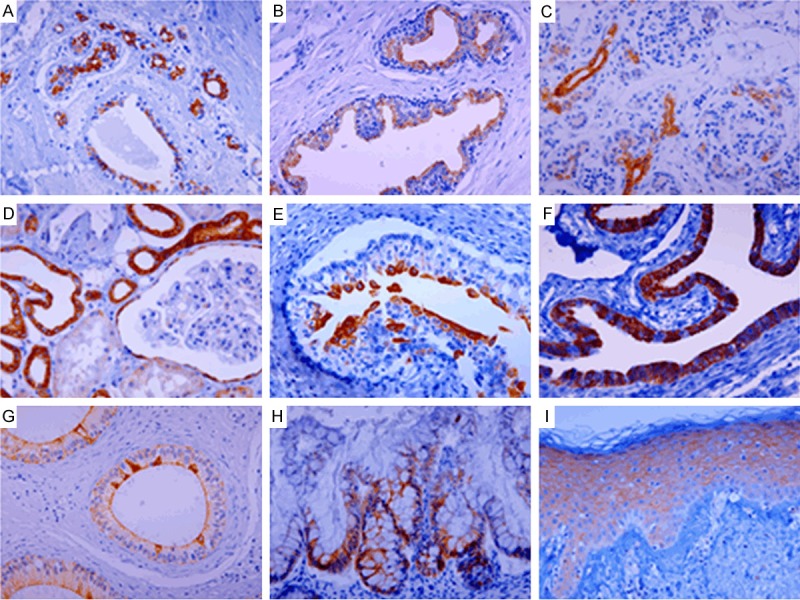
Immunoreactions of pS9GSK3β in normal adult (A, B, D, F, G and I) and fetal (C, E and H) human tissues. pS9GSK3β shows strong cytoplasmic and membranous staining in the luminal epithelial cells of the breast (A) and prostate (B). In the pancreas, pS9GSK3β shows strong membranous staining in ductal and centroacinar cells, occasional cytoplasmic staining in acinar cells, and negativity in islet (C). In the kidney, strong cytoplasmic and membranous staining of pS9GSK3β is seen in distal tubule and collecting duct and pS9GSK3β staining is occasionally seen in Bowman’s capsule and proximal tubule (D). In the bladder, membranous pS9GSK3β expression is strong in urothelial superficial cells and faintly seen in basal to intermediate cells (E). In fallopian tube, strong membranous pS9GSK3β expression is exclusively seen in secretory cells, but not in ciliated cells (F). In the epididymis, pS9GSK3β staining is diffusely seen in whole epithelial cells with membranous pattern and moderate intensity. Strong cytoplasmic pS9GSK3β staining is seen in scattered cells of luminal side (G). pS9GSK3β expression is mainly located in the crypts of large intestine with strong intensity (H). In the epidermis, pS9GSK3β is moderately stained in intercellular bridges of squamous cells (I). Original magnifications: × 400.
In the female reproductive tract, ovarian surface epithelial cells were moderately immunoreactive for pS9GSK3β. Follicular cells of some primordial follicles were positive for pS9GSK3β, however, oocytes and more mature form of follicles, and stromal cells were all negative for this protein In the fallopian tube (Figure 2F), secretory cells were strongly positive for pS9GSK3β, whereas ciliated cells were negative for pS9GSK3β. In the endometrium of the uterus, pS9GSK3β was constantly positive with moderate intensity in proliferative phase glands, but variable in secretory phase glands. The columnar cells of endocervix and squamous cells of exocervix were all negative for pS9GSK3β.
In the male reproductive tract, rete testis showed moderate pS9GSK3β staining, but seminiferous tubules showed negative staining. In the epididymis (Figure 2G), the epithelial cells showed diffusely moderate membranous pS9GSK3β staining with strong cytoplasmic staining of pS9GSK3β in scattered cells of the luminal side.
The expression pattern of pS9GSK3β was similar in most organs of adult and fetal tissues, but the difference was noted in the gastrointestinal and respiratory tracts. In fetal digestive tract, pS9GSK3β was strongly expressed in the crypts of small and large intestine (Figure 2H) and weakly in the foveolar pits of the stomach. In adult tissue, the intensity of pS9GSK3β was weaker than that in the fetal tissue with additional staining in the surface epithelium of gastrointestinal tract and body glands of stomach. In fetal lung, pS9GSK3β expression was occasionally seen in bronchial epithelium and gradually intensified in terminal alveolar unit, whereas, in adult lung, pS9GSK3β expression was faintly seen in certain bronchial epithelia and negative in pneumocytes. Pleural mesothelial cells were strongly positive for pS9GSK3β.
The expression of pS9GSK3β was different in the squamous epithelium from mucosa and skin. In the squamous mucosa such as esophagus and uterine cervix, pS9GSK3β was not detectable. However, in the epidermis (Figure 2I), pS9GSK3β was moderately stained in intercellular bridges of squamous cells. In addition, pS9GSK3β was strongly stained in secretory cells of sweat gland with cytoplasmic and membranous patterns. The lymphoid tissue, neural tissue and mesenchymal elements including fibroblasts, endothelial cells, and muscle cells were unreactive for pS9GSK3β.
Comparison of GSK3β, pS9GSK3β, CK7, and CK20 expression pattern
GSK3β, pS9GSK3β and CK7 showed common expression pattern only in epithelial cells of sweat glands and breast with stronger intensity in CK7 and pS9GSK3β than GSK3β. The expression pattern of pS9GSK3β was mostly corresponding to that of CK7 in epithelia of breast, pancreas, liver, kidney, fallopian tube, and stomach. In the prostate, pS9GSK3β expression was limited to the luminal secretory cells of glands, whereas CK7 expression was observed sporadically in the basal cells as well as luminal cells. In the bladder, the difference between pS9GSK3β and CK7 expression was that pS9GSK3β was strongly stained in the umbrella cells but CK7 was in the whole urothelial layer. In the epididymis, CK7 showed similar scattered staining pattern as seen in pS9GSK3β stain. The ovarian surface epithelia and mesothelial cells were positive for CK7, similar to pS9GSK3β. Interestingly, CK7 was negative in follicular cells of primordial follicles, but dot-like CK7 immunoreaction was noted in the cytoplasm of primary oocytes of the ovary.
CK20 expression was restricted in the gastrointestinal epithelia and umbrella cells of urinary bladder. In the gastrointestinal tract, the expression site of pS9GSK3β and CK20 was slightly different. The expression of pS9GSK3β seen in fetal gastric foveolar pits and intestinal crypts extended to the surface epithelia of the gut in the adult tissue, whereas CK20 was localized in the surface epithelia of gastric and intestinal mucosa in both fetal and adult tissues. Distinct CK20 expression in umbrella cell of bladder corresponded to that of pS9GSK3β, which was intensified in fetal tissue than in adult tissue.
Discussion
In this study, we observed tissue and cell type restricted expression of GSK3β and pS9GSK3β in normal human tissue. GSK3β was selectively expressed in neural tissue, skin, sweat gland and mammary gland, whereas pS9GSK3β was expressed in the epithelial lining of gastrointestinal tract, urinary tract and duct containing organs, such as pancreas and liver. This result is corresponding with the previous data showing that GSK3β and pS9GSK3β are associated with the development of ectodermal and endodermal germ layer, respectively [14-16]. GSK3β has been known to regulate cell-fate specification in vertebrates and invertebrates [17]. In Xenopus embryos [16], GSK3β mediates the initial steps of neural tissue specification and modulates anterior ectodermal development. In sea urchin embryo [15], inactive GSK3β expands endoderm and GSK3β overexpression provokes ectoderm territory, showing similar expression pattern to ours. In our result, pS9GSK3β was not detected in neural tissue as expected, however, other ectodermal derivatives, such as skin, sweat gland and mammary gland showed the expression of pS9GSK3β as well as GSK3β, suggesting the physiologic requirement of balanced expression of GSK3β and pS9GSK3β in these tissues. Previously, Ma et al [10] report colocalization of GSK3β and pS9GSK3β in the cytoplasm of human keratinocytes, which is different from our result showing cytoplasmic or occasional nuclear GSK3β expression and pS9GSK3β expression in intercellular bridges of keratinocytes.
In normal breast tissue, GSK3β is shown to be strongly stained in the cytoplasm of mammary gland epithelium [9], however, there is no study of pS9GSK3β expression in breast tissue. In dairy cow, GSK3β and pS9GSK3β regulate proliferation of mammary epithelial cells and milk synthesis via the mTOR/S6K1 signaling pathway [6]. The inhibition of GSK3β by lithium chloride, a known inhibitor of GSK3β, promotes GSK3β phosphorylation and increases the expression of mTOR, p-mTOR, S6K1, p-S6K1, CyclinD1, SREBP1, and β-casein, leading to cell growth and synthesis of protein and lipid [6]. In our study, the staining intensity of pS9GSK3β was higher than that of GSK3β in mammary epithelial cells, however, comparative expression level of these two proteins is supposed to be variable depending on the physiological condition and hormonal status of the host. The luminal epithelial localization of pS9GSK3β was also observed in secretory cells of sweat gland and prostate in our study. These combined findings may imply that the luminal localization of pS9GSK3β is associated with the secretory function of the cells of these organs.
We observed intense expression of pS9GSK3β and CK7 in centroacinar and ductal cells of the pancreas, which is corresponding to the result of Pham et al [12]. Pancreatic ductal cell differentiation is known to be mediated by PI3K/Akt pathway, while ductal cell proliferation is by the MEK-ERK1/2 pathway [18]. Taken together, PI3K/Akt mediated inactivation of GSK3β appears to be involved in pancreatic ductal cell differentiation.
In the bladder, pS9GSK3β staining was distinctively membranous with strong intensity in umbrella cells, corresponding with the staining pattern of CK20 in the urothelium. In mouse urothelium [19], CK20 expression appears in superficial cells on late embryonic days than CK7 expression. CK20 is known as differentiation marker of umbrella cells [20] and contributes to the elastic properties of cytokeratin network [19]. The umbrella cells form tight barrier to urine, toxin, and pathogen [21]. Sharp membranous staining of pS9GSK3β in umbrella cells may show its association with barrier function of umbrella cells. In addition to CK20, pS9GSK3β is considered to be a possible differentiation marker of umbrella cell.
In reproductive tract, pS9GSK3β showed cell type specific expression pattern. We found that pS9GSK3β and CK7 were only positive in secretory cells of fallopian tube, which are attracting an attention because of recent concept that clonal expansion of the secretory cells may cause ovarian serous carcinoma [22,23]. Previously, bcl2 and PAX8 are known as markers identifying tubal secretory cell [22,23] and we add p-GSK3β and CK7 to the tubal secretory cell marker. In the epididymal lining cells, there are four cell types: narrow, clear, principal, and basal cells [24]. Their delicate communication, together with spermatozoa, controls the acidic luminal environment for the maturation and viability of spermatozoa [24]. Narrow and clear cells expressing the proton-pumping ATPase and secreting protons, principal cells secreting HCO(3)(-), and basal cells secreting nitric oxide which stimulate proton secretion in clear cells work in a concerted manner for luminal acidification [24]. Our study showed strong cytoplasmic expression of pS9GSK3β in scattered cells of luminal side, which were considered as narrow or clear cells. Cell type restricted expression of pS9GSK3β was also observed in the kidney, where the staining was confined to the distal tubule and collecting duct. Distal tubule is involved in the reabsorption of sodium ions from the tubular fluid coupled with the secretion of hydrogen and potassium ions into the tubule. The expression of pS9GSK3β in specific epididymal cells and distal nephron in our study may show that pS9GSK3β is associated with proton secretory function and acid-base balance in these cells. Kjaersgaard et al [25] showed the same restricted distribution of GSK3β and pS9GSK3β immunoreaction in distal tubule and collecting duct in fetal and adult kidney. They also displayed GSK3β, pS9GSK3β, and PCNA immunolabeling in the epithelia of microcysts developed in patients chronically treated with lithium. They concluded that lithium enters into the aldosterone-sensitive distal nephron and causes inactivation of GSK-3β, proliferation, and microcyst formation [25]. The discrepancy between our and their studies’ results was that GSK3β staining was negligible in the kidney in our study. Further studies would be needed to define the pattern of GSK3β expression in the kidney by comparing various anti-GSK3β antibodies.
We found different expression pattern of pS9GSK3β in the gastrointestinal and respiratory tracts from fetal and adult tissues. In adult gut mucosa, surface epithelial cells were stained for pS9GSK3β, which was not detected in fetal surface cells of gastrointestinal tract. The intestinal epithelial monolayer provides barrier against various luminal insults and defect in this layer necessitates a rapid repair. In response to wounding, PI3K-dependent GSK3β (Ser9)-phosphorylation is involved in the intestinal epithelial restitution [7]. The expression of pS9GSK3β in the intestinal surface epithelia in our result may demonstrate the involvement of pS9GSK3β in maintenance of epithelial barrier, which is more important in adult tissue. Epithelial barrier function is maintained by GSK-3 regulating apical junctional complex transmembrane proteins such as occludin, claudin-1 and E-cadherin [26].
In the respiratory tract, pS9GSK3β was not detected in pneumocytes of adult lung, whereas moderately stained in those of fetal lung. In fetal mice [27], the activation of PI3K-Akt-mTOR pathway in the lung is associated with delayed maturation of the lung epithelial cells and reduced alveolar capillary density. Thus, downregulation of the PI3K-Akt-mTOR pathway is required for normal lung epithelial maturation [27]. Although the link of pS9GSK3β with PI3K-Akt-mTOR axis during the development of lung remains unclear, lower expression of pS9GSK3β in the adult than in the fetal lung tissue in our study suggests an association between downregulated pS9GSK3β and pneumocyte maturation. In addition, the expression of pS9GSK3β is highest in adenocarcinoma compared with other types of lung carcinomas [11]. Taken together, pS9GSK3β is suspected to be critical for the fetal alveolar structuring to the certain point in fetal development and tumor histogenesis in the lung.
The correlation between pS9GSK3β and CK is not known, however, Akt/mTOR signaling pathway, in which GSK3β is a downstream molecule, is shown to be closely linked with CK in keratinocyte repair [28]. In wounded keratinocyte, CK17 is induced and binds with the adaptor protein 14-3-3σ, leading to mTOR activity and cell growth [28]. Recently, 14-3-3σ is reported to regulate embryonic stem cell proliferation by binding, phosphorylating and sequestrating GSK3β in a PI3K/Akt-dependent manner and enhancing β-catenin translocation into nucleus and proliferative transcription [29]. In our study, CK7 and pSGSK3β were colocalized in epithelial cells of pancreas, kidney, breast, fallopian tube, and CK20 and pSGSK3β in umbrella cells of bladder. Simultaneous expression of CK and pSGSK3β in certain epithelial cells may be coincidental finding. However, in that CK is involved in cell growth and GSK3β is a downstream molecule of PI3K/Akt/mTOR signaling, further study clarifying the relation between CK and 14-3-3σ mediated pS9GSK3β will be of interest.
In summary, we showed GSK3β expression in ectodermal derived tissues such as, neural tissue, skin, sweat gland and mammary gland, and pS9GSK3β expression in the epithelial cells of endodermal derived tissues, such as gastrointestinal tract, pancreaticobiliary tract, and urogenital tract. The cytoplasmic and/or membranous expression of pS9GSK3β in the specific epithelial cells may implicate its association with secretory or barrier function depending on the tissue and cell type.
Our comprehensive study on normal adult and fetal tissues would give invaluable information for the future studies on different disease conditions, and pS9GSK3β may serve as another immunohistochemical marker for epithelial cells.
Disclosure of conflict of interest
None.
References
- 1.Doble BW, Woodgett JR. GSK-3: tricks of the trade for a multi-tasking kinase. J Cell Sci. 2003;116:1175–1186. doi: 10.1242/jcs.00384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Shakoori A, Ougolkov A, Yu ZW, Zhang B, Modarressi MH, Billadeau DD, Mai M, Takahashi Y, Minamoto T. Deregulated GSK3beta activity in colorectal cancer: its association with tumor cell survival and proliferation. Biochem Biophys Res Commun. 2005;334:1365–1373. doi: 10.1016/j.bbrc.2005.07.041. [DOI] [PubMed] [Google Scholar]
- 3.Mishra R. Glycogen synthase kinase 3 beta: can it be a target for oral cancer. Mol Cancer. 2010;9:144. doi: 10.1186/1476-4598-9-144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Miyashita K, Kawakami K, Nakada M, Mai W, Shakoori A, Fujisawa H, Hayashi Y, Hamada J, Minamoto T. Potential therapeutic effect of glycogen synthase kinase 3beta inhibition against human glioblastoma. Clin Cancer Res. 2009;15:887–897. doi: 10.1158/1078-0432.CCR-08-0760. [DOI] [PubMed] [Google Scholar]
- 5.Cross DA, Alessi DR, Cohen P, Andjelkovich M, Hemmings BA. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature. 1995;378:785–789. doi: 10.1038/378785a0. [DOI] [PubMed] [Google Scholar]
- 6.Zhang X, Zhao F, Si Y, Huang Y, Yu C, Luo C, Zhang N, Li Q, Gao X. GSK3beta regulates milk synthesis in and proliferation of dairy cow mammary epithelial cells via the mTOR/S6K1 signaling pathway. Molecules. 2014;19:9435–9452. doi: 10.3390/molecules19079435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Karrasch T, Spaeth T, Allard B, Jobin C. PI3K-dependent GSK3ss(Ser9)-phosphoryla tion is implicated in the intestinal epithelial cell wound-healing response. PLoS One. 2011;6:e26340. doi: 10.1371/journal.pone.0026340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Clevers H, van de Wetering M. TCF/LEF factor earn their wings. Trends Genet. 1997;13:485–489. doi: 10.1016/s0168-9525(97)01305-x. [DOI] [PubMed] [Google Scholar]
- 9.Mylona E, Vamvakaris I, Giannopoulou I, Theohari I, Papadimitriou C, Keramopoulos A, Nakopoulou L. An immunohistochemical evaluation of the proteins Wnt1 and glycogen synthase kinase (GSK)-3beta in invasive breast carcinomas. Histopathology. 2013;62:899–907. doi: 10.1111/his.12095. [DOI] [PubMed] [Google Scholar]
- 10.Ma C, Wang J, Gao Y, Gao TW, Chen G, Bower KA, Odetallah M, Ding M, Ke Z, Luo J. The role of glycogen synthase kinase 3beta in the transformation of epidermal cells. Cancer Res. 2007;67:7756–7764. doi: 10.1158/0008-5472.CAN-06-4665. [DOI] [PubMed] [Google Scholar]
- 11.Zheng H, Saito H, Masuda S, Yang X, Takano Y. Phosphorylated GSK3beta-ser9 and EGFR are good prognostic factors for lung carcinomas. Anticancer Res. 2007;27:3561–3569. [PubMed] [Google Scholar]
- 12.Pham NA, Schwock J, Iakovlev V, Pond G, Hedley DW, Tsao MS. Immunohistochemical analysis of changes in signaling pathway activation downstream of growth factor receptors in pancreatic duct cell carcinogenesis. BMC Cancer. 2008;8:43. doi: 10.1186/1471-2407-8-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lee H, Choi SK, Ro JY. Overexpression of DJ-1 and HSP90alpha, and loss of PTEN associated with invasive urothelial carcinoma of urinary bladder: Possible prognostic markers. Oncol Lett. 2012;3:507–512. doi: 10.3892/ol.2011.522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Diekmann U, Lenzen S, Naujok O. A reliable and efficient protocol for human pluripotent stem cell differentiation into the definitive endoderm based on dispersed single cells. Stem Cells Dev. 2015;24:190–204. doi: 10.1089/scd.2014.0143. [DOI] [PubMed] [Google Scholar]
- 15.Emily-Fenouil F, Ghiglione C, Lhomond G, Lepage T, Gache C. GSK3beta/shaggy mediates patterning along the animal-vegetal axis of the sea urchin embryo. Development. 1998;125:2489–2498. doi: 10.1242/dev.125.13.2489. [DOI] [PubMed] [Google Scholar]
- 16.Itoh K, Tang TL, Neel BG, Sokol SY. Specific modulation of ectodermal cell fates in Xenopus embryos by glycogen synthase kinase. Development. 1995;121:3979–3988. doi: 10.1242/dev.121.12.3979. [DOI] [PubMed] [Google Scholar]
- 17.Kim L, Kimmel AR. GSK3, a master switch regulating cell-fate specification and tumorigenesis. Curr Opin Genet Dev. 2000;10:508–514. doi: 10.1016/s0959-437x(00)00120-9. [DOI] [PubMed] [Google Scholar]
- 18.Uzan B, Figeac F, Portha B, Movassat J. Mechanisms of KGF mediated signaling in pancreatic duct cell proliferation and differentiation. PLoS One. 2009;4:e4734. doi: 10.1371/journal.pone.0004734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Erman A, Veranic P, Psenicnik M, Jezernik K. Superficial cell differentiation during embryonic and postnatal development of mouse urothelium. Tissue Cell. 2006;38:293–301. doi: 10.1016/j.tice.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 20.Moll R, Lowe A, Laufer J, Franke WW. Cytokeratin 20 in human carcinomas. A new histodiagnostic marker detected by monoclonal antibodies. Am J Pathol. 1992;140:427–447. [PMC free article] [PubMed] [Google Scholar]
- 21.Apodaca G. The uroepithelium: not just a passive barrier. Traffic. 2004;5:117–128. doi: 10.1046/j.1600-0854.2003.00156.x. [DOI] [PubMed] [Google Scholar]
- 22.Li J, Fadare O, Xiang L, Kong B, Zheng W. Ovarian serous carcinoma: recent concepts on its origin and carcinogenesis. J Hematol Oncol. 2012;5:8. doi: 10.1186/1756-8722-5-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li J, Ning Y, Abushahin N, Yuan Z, Wang Y, Wang Y, Yuan B, Cragun JM, Chambers SK, Hatch K, Kong B, Zheng W. Secretory cell expansion with aging: risk for pelvic serous carcinogenesis. Gynecol Oncol. 2013;131:555–560. doi: 10.1016/j.ygyno.2013.09.018. [DOI] [PubMed] [Google Scholar]
- 24.Shum WW, Ruan YC, Da Silva N, Breton S. Establishment of cell-cell cross talk in the epididymis: control of luminal acidification. J Androl. 2011;32:576–586. doi: 10.2164/jandrol.111.012971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kjaersgaard G, Madsen K, Marcussen N, Christensen S, Walter S, Jensen BL. Tissue injury after lithium treatment in human and rat postnatal kidney involves glycogen synthase kinase-3beta-positive epithelium. Am J Physiol Renal Physiol. 2012;302:F455–465. doi: 10.1152/ajprenal.00144.2011. [DOI] [PubMed] [Google Scholar]
- 26.Severson EA, Kwon M, Hilgarth RS, Parkos CA, Nusrat A. Glycogen Synthase Kinase 3 (GSK-3) influences epithelial barrier function by regulating occludin, claudin-1 and E-cadherin expression. Biochem Biophys Res Commun. 2010;397:592–597. doi: 10.1016/j.bbrc.2010.05.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ikeda H, Shiojima I, Oka T, Yoshida M, Maemura K, Walsh K, Igarashi T, Komuro I. Increased Akt-mTOR signaling in lung epithelium is associated with respiratory distress syndrome in mice. Mol Cell Biol. 2011;31:1054–1065. doi: 10.1128/MCB.00732-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kim S, Wong P, Coulombe PA. A keratin cytoskeletal protein regulates protein synthesis and epithelial cell growth. Nature. 2006;441:362–365. doi: 10.1038/nature04659. [DOI] [PubMed] [Google Scholar]
- 29.Chang TC, Liu CC, Hsing EW, Liang SM, Chi YH, Sung LY, Lin SP, Shen TL, Ko BS, Yen BL, Yet SF, Wu KK, Liou JY. 14-3-3sigma regulates beta- catenin-mediated mouse embryonic stem cell proliferation by sequestering GSK-3beta. PLoS One. 2012;7:e40193. doi: 10.1371/journal.pone.0040193. [DOI] [PMC free article] [PubMed] [Google Scholar]