

Abstract
Circadian clocks regulate many aspects of plant physiology and development that contribute to essential agronomic traits. Circadian clocks contain transcriptional feedback loops that are thought to generate circadian timing. There is considerable similarity in the genes that comprise the transcriptional and translational feedback loops of the circadian clock in the plant Kingdom. Functional characterisation of circadian clock genes has been restricted to a few model species. Here we provide a functional characterisation of the Hordeum vulgare (barley) circadian clock genes Hv CIRCADIAN CLOCK ASSOCIATED 1 (HvCCA1) and Hv PHOTOPERIODH1, which are respectively most similar to Arabidopsis thaliana CIRCADIAN CLOCK ASSOCIATED 1 (AtCCA1) and PSEUDO RESPONSE REGULATOR 7 (AtPRR7). This provides insight into the circadian regulation of one of the major crop species of Northern Europe. Through a combination of physiological assays of circadian rhythms in barley and heterologous expression in wild type and mutant strains of A. thaliana we demonstrate that HvCCA1 has a conserved function to AtCCA1. We find that Hv PHOTOPERIOD H1 has AtPRR7-like functionality in A. thaliana and that the effects of the Hv photoperiod h1 mutation on photoperiodism and circadian rhythms are genetically separable.
Introduction
Circadian clocks are timing mechanisms that are an adaptation to the Earth’s rotation. These circadian clocks are entrained to the day/night cycle by sensing environmental cues such as light and temperature and act as master regulators to synchronise biological events to specific times of the day [1]. In plants, the circadian clock has been shown to control approximately one third of the genome and it regulates a wide range of key processes such as hypocotyl elongation, leaf movement, stomatal opening and flowering [1]. Furthermore, accurate and robust circadian function enhances yield, water use efficiency and overall plant performance [2,3].
While the molecular components of the clock are not conserved across taxonomic groups, most circadian oscillators comprise a number of coupled transcription/translation feedback loops [4]. In the model dicotyledonous plant Arabidopsis, the central clock is proposed to consist of several interlocking loops [5]. A loop of morning active genes is formed by the partially redundant MYB-transcription factors AtCCA1 (CIRCADIAN CLOCK ASSOCIATED 1) and AtLHY (LATE ELONGATED HYPOCOTYL) and two members of the pseudo-response regulators family, AtPRR7 (PSEUDO-RESPONSE REGULATOR 7), and AtPRR9 (PSEUDO-RESPONSE REGULATOR 9). AtCCA1/LHY activate AtPRR7/9 and in turn AtPRR7/9 repress AtCCA1/LHY. A loop of evening-active genes is formed by AtGI (GIGANTEA) and AtPPR1/TOC1 (PSEUDO-RESPONSE REGULATOR 1/TIMING OF CAB EXPRESSION) which both negatively regulate each other. The loops are coupled by the repressive effects of AtCCA1/LHY on AtTOC1 and AtTOC1 on AtCCA1/LHY [6,7]. Another loop of mutual repression exists between the morning loop and an “evening complex” of AtELF3, AtELF4 and LUX (EARLY FLOWERING 3, 4 and LUXARRHYTHMIO) through recruitment of the transcriptional repressor LUX by ELF3 to the PRR9 promoter [8].
Monocot circadian clocks appear to be comprised of a similar structure. For example, in Lemna gibba, a floating monocotyledon plant, orthologues for AtCCA1, AtLHY, AtGI, AtELF3 and several genes similar to the AtPRR family have been identified and their function in the Lemna circadian clock characterised [9]. In rice, another model monocot plant, several orthologues of central clock genes have also been identified, including OsGI, OsZTL (ZEITLUPE), OsCCA1/LHY and members of the OsPRR family [10]. These components are under circadian control with similar expression patterns reported for homologous clock genes in Arabidopsis.
Most monocots contain only a single orthologues of AtCCA1/LHY. For example, rice has a single orthologues of AtCCA1/LHY (OsLHY) which functions in a similar manner to the Arabidopsis, acting as a repressor of the rice AtTOC1 orthologues [11]. However, OsLHY does not homodimerize due to the loss of a key phosphorylation site, which demonstrates divergence between the rice and Arabidopsis clocks [11].
Barley contains orthologues of Arabidopsis circadian clock genes, with transcripts that are rhythmic suggesting that HvCCA1, HvGI and HvPRR1 have a functions similar to their Arabidopsis counterparts [12]. For some of the PRR homologues it has proven difficult to determine which barley gene is ortholgous to Arabidopsis members of the family and therefore the designation reflects the uncertainty. HvPRR37, HvPRR73, describe two genes both of which are ortholougues to either AtPRR3 or AtPRR7, and HvPRR59 and HvPRR95 are orthologus to either AtPRR5 or AtPRR9 [12]. Other circadian clock genes are thought to be important for agronomic traits such as HvLUX1 and HvELF3 underlying the EARLY MATURITY 10 [13] and 8 [14] loci respectively.
HvPRR37 is also known as Hv PHOTOPERIOD H 1 (HvPpd-H1), which controls sensitivity to photoperiod. The Ppd-H1 region contains a single PRR gene originally thought to be most similar to AtPRR7 [15]. Landraces from south-west Asia, southern Europe, and the Mediterranean basin have the Ppd-H1 allele that confers early flowering in long days of the spring. Whereas the photoperiodic-insensitive ppd-H1 allele is present in landraces from central and northern Europe (this is known as spring barley, because it is planted in the spring and flowers at the end of summer). The reduced response to photoperiod of ppd-H1 allows spring-sown plants to extend the period of vegetative growth and accumulate additional biomass that supports higher yields [15]. The late-flowering ppd-H1 allele is recessive, suggesting that reduced response results from a mutation that impairs gene function [15]. The modern variants of barley used to map Ppd-H1 were Igri which contains the photoperiod-sensitive allele (Ppd-H1) and Triumph which is late flowering (ppd-H1). The central role of HvPpd-H1 (and TaPpd-D1a) in flowering is well characterised, but it is not known whether HvPpd-H1 is a circadian clock gene in barley, or whether it is functionally equivalent to AtPRR7 or AtPRR3 because natural variation at the HvPpd1-H1 has no effect on circadian rhythms of circadian clock gene transcripts but does affect the timing of expression of genes associated with flowering time [12, 14].
Barley represents a challenging system for studying circadian behaviour and it does not have the wide resource tools for molecular genetics developed for the model Arabidopsis, therefore we have used a heterologous approach to examine barley clock gene function by misexpression in Arabidopsis. We provide evidence that the barley circadian clock genes have conserved functions with their counterparts in Arabidopsis. We demonstrate that Hv CCA1 has AtCCA1-like activity and HvPpd-H1 has AtPRR7-like functionality in Arabidopsis. We suggest that the effects of HvPpd-H1 on photoperiodism and circadian rhythms are separable. Our data suggests that flowering time in crops can be manipulated by appropriate breeding strategies without compromising circadian clock function.
Methods
Cloning of HvCCA1 into Arabidopsis
The entire HvCCA1 transcript was amplified using KOD TAQ polymerase (Novagen). HvCCA1 was cloned into GATEWAY pCR8/GW/TOPO entry vector using pCR8/GW/TOPO TA cloning kit (Invitrogen) following the manufacturer’s instructions. HvCCA1 was then subcloned into the binary plazmid PMDC32 using the Gateway LR recombination reaction (Invitrogen). The binary plasmid PMDC32 confers hygromycin resistance in plants and contains a double 35S cauliflower mosaic virus promoter for constitutive expression of the inserted gene [16]. The CaMV35S:HvCCA1 was introduced into GV3101 Agrobacterium tumefaciens and subsequently into Arabidopsis thaliana Ws-2 via the floral dip method [17]. Out of 21 primary transformants, nine were identified as homozygous lines carrying an insert at a single locus. HvCCA1 overexpression was confirmed by RT-PCR (S1 Fig). Homozygous lines were used for all experiments.
RNA extraction and transcript analysis
For circadian experiments seedlings were grown at 22°C in 12 h:12 h L:D for 7 days before being transferred to continuous light for 54 h. Samples were taken every 6 h until 72 h. Plant tissue was disrupted in the TissueLyser (QIAGEN) and total RNA was extracted using RNeasy plant kit (Qiagen). cDNA was synthesised from 0.5 μg of template RNA using the QuantiTect Reverse Transcription kit (Qiagen). For quantitative PCR, transcript abundance was measured using Power SYBR Green PCR master mix (Applied Biosystems, USA) in the Applied Biosystems 7500 Fast Real-Time PCR System. Transcript abundance of AtCCA1, AtLHY and AtTOC1 was normalised to UBIQUITIN10 (UBQ). The primers for q-PCR reactions are presented in S1 Table. All reactions were carried out in triplicate. Normalization and transcript abundance calculations were automated and performed using the Applied Biosystems 7500 Fast Real-Time PCR System software.
Non-homologous Complementation of Arabidopsis PRR Mutants
Arabidopsis PRR7 T-DNA insertion mutant (prr7-11) was complemented with HvPpd-H1 and Hvppd-H1 to test for similarity of function between AtPRR7 and HvPpd-H1. pENTRPpdH1 (Igri) cDNA and of pENTRppdH1 (Triumph) were generated, each containing a fragment comprising the coding region of the HvPpdH1 or HvppdH1 cDNA from Cultivar “Igri” or “Triumph” respectively, cloned directionally into a pENTR-D plasmid (Invitrogen, UK). The genomic AtPRR7 (AT5G02810) promoter (p) sequence was used to drive the HvPpd-H1 cDNA sequences extending from -871 to +1 bases where the first nucleotide of the inferred ATG initiation codon. The promoter was amplified (S1 Table) from pAtPRR7:LUC, a luciferase binary vector supplied by C. Robertson McClung, originally derived from pZPomegaLUCplus [18] by replacement of the gentamicin resistance cassette with the BASTA resistance gene from p35SBarn [19]. The resulting vector, pZPBAR, was then made Gateway compatible (Invitrogen, Carlsbad, CA) by inserting the PCR-amplified attR-flanked destination cassette from pK7WG2D [20] at the BamHI and HindIII sites upstream of LUC to create pZPBAR-DONR. The AtPRR7 promoter was inserted into pZPBAR-DONR. The binary vector used was pBGW, supplied by the Functional Genomics unit of the Department of Plant Systems Biology (VIB-Ghent University). The 894 bp pAtPRR7 product and pENTR-D Topo Ppd-H1 and pENTR-D Topo ppd-H1 recipient plasmids were digested with SacII restriction enzyme. The digested pENTR-D Topo PpdH1 and pENTR-D Topo ppdH1 samples were dephosphorylated by adding Shrimp Alkaline Phosphatase (SAP) buffer (Fermentas, UK),. The pAtPRR7 insert DNA and pENTR-D Topo PpdH1 / ppdH1 vector backbones were mixed in a ratio of 3:1 (insert:backbone) with ligase (New England BioLabs, UK) and incubated at 4°C overnight. pENTR-D Topo PpdH1 or pENTR-D Topo ppdH1 with the AtPRR7 promoter were introduced into chemically competent DH5α E. coli cells (Invitrogen, UK). Colony PCR was used to identify bacteria carrying the pENTR-D Topo PpdH1 or pENTR-D Topo ppdH1 with the AtPRR7 promoter in the correct orientation. Plasmid DNA was isolated using Promega miniprep Wizard kits (Promega, UK). The insertion and orientation of AtPRR7 into the pENTR-D Topo backbones was verified again by restriction digestion alongside the parent plasmids using SacI and SacII. Plasmids carrying the AtPRR7 promoter in the correct insertion were fully sequenced through the AtPRR7 promoter and the Igri or Triumph coding regions. A. tumefasciens strain GV3101 was transformed with pBGW-PRR7:PpdH1 or pBGW-PRR7:ppdH1 and then used to transform Arabidopsis by floral dip [17]. Positive transformants were selected by spray with Finale glufosinate ammonium (0.31% v/v; Bayer, Germany). For physiological assays the F2 transgenic plants were used.
To ensure functionality of the promoter construct tissue was collected from 12-day-old seedlings grown in 12 h light–12 h dark cycles (100 μmol m−2 s−1) six h after dawn. Total RNA was extracted from frozen tissue of three biological replicates of at least five pooled plants each, using the RNeasy Plant Mini Kit (QIAGEN) and RNase-Free DNase on-column treatment (QIAGEN). cDNA was synthesized from 500 ng RNA with the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific) using oligo(dT) primers. The gene-specific products were amplified using the Rotor-Gene SYBR Green PCR Kit on a Rotor-Gene 6000 Real-Time PCR machine (QIAGEN). The primer sequences were: PP2a-F, 5′-TAACGTGGCCAAAATGATGC-3′; PP2a-R, 5′-GTTCTCCACAACCGCTTGGT-3′ and Hvppd-H1-F, 5′-GATGGATTCAAAGGCAAGGA-3′; Hvppd-H1-R, 5′-GAACAATTGGCTCCTCCAAA-3′ [12]. Relative transcript levels were determined by incorporating PCR efficiencies [21].
Circadian rhythms of leaf movement, delayed fluorescence and analysis of hypocotyl elongation and flowering time in Arabidopsis
For leaf movement, delayed fluorescence and hypocotyl elongation experiments Arabidopsis seed was surface-sterilized in 70% ethanol for 1 min, followed by 50% (w/v) bleach for 10 min and 3 rinses with distilled water. Seeds were resuspended in 0.15% (w/v) agar and stratified at 4°C for 3 days prior to sowing on Murashige and Skoog medium 1.5% (w/v) agar plates. For leaf movement, ten day old seedlings entrained to 12h light 12 h dark cycles were moved to Sanyo MLR350 plant growth chambers and imaged under constant light at 22°C [22].
Delayed chlorophyll fluorescence was imaged in a Sanyo MIR-553 cooled incubator (Sanyo Gallenkamp, UK) using an ORCA-II-BT 1024 16-bit camera (Hamamatsu Photonics, Japan) cooled to -80°C [20]. Images from leaf movement and delayed fluorescence were processed using Metamorph 6.0 image-analysis software (Molecular Devices, UK). Period estimates and relative amplitude errors (RAE) were calculated in BRASS (available from http://www.amillar.org/downloads.html) by running fast Fourier transformed non-linear least-square analysis [23]. RAE is a measure of rhythm robustness that ranges from 0 (a perfect fit to the wave) to 1 (no fit).
Hypocotyl length was measured in ten day old seedlings grown at 22°C in 12h light 12 h dark of 100 μmol m-2 s-1. Seedlings were harvested and laid flat on agar plates for imaging. Images of Arabidopsis seedlings were analysed in Metamorph 6.0. Student’s t-test was used to determine statistical significance between the controls and mutants.
Flowering time assays were performed on Arabidopsis seedlings grown in a controlled environment under 16h light 8 h dark at 22°C. The flowering time was expressed as a number of days and rosette leaves when a bolt of 0.5 cm long was observed.
Infra-Red Gas Exchange Analyser (IRGA) Assay of Circadian Rhythms in Arabidopsis and barley
Circadian rhythms of gas exchange in Arabidopsis plants was measured as described in [24]. For measurement of circadian gas exchange in individual barley plants an IRGA system based on the portable LI-COR LI-6400XT (LI-COR, USA) was developed. Humid CO2 free air was obtained by pumping atmospheric air through two 4.5 cm x 30 cm cylinders in series filled with soda lime (Fisher Scientific, UK) and a flask of water. Relative humidity was limited to a maximum of 85% through the control of the IRGA. CO2 (BOC, UK) was provided at between 180 and 220 PSIG and a flow of 500 μmol s-1. The [CO2] in the chamber was controlled at 380 PPM. A 6 cm2 area of mature barley (4–8 weeks old) leaf was clamped in the cuvette with a clear glass screen and therefore received illumination from the cabinet equivalent to that of the rest of the plant. Gaseous exchange was recorded every 10 min.
Results
Over-expression of HvCCA1 causes arrhythmia of the Arabidopsis circadian clock
Over-expression of AtCCA1 in Arabidopsis abolishes circadian rhythms in multiple outputs of the circadian clock [25]. Here we investigated whether HvCCA1 is functionally orthologous to AtCCA1 by assaying circadian regulation of leaf movement, delayed chlorophyll fluorescence, transcript abundance and gas exchange in Arabidopsis overexpressing HvCCA1 driven by 2x 35S cauliflower mosaic virus promoters (HvCCA1-ox; (S1 Fig). In continuous light (LL), Ws-2 and Col-0 wildtypes had robust circadian rhythms of delayed fluorescence (Ws-2 = 23.4 h; Col-0 = 23.6 h; Table 1, Fig 1) and leaf movement (Ws-2 = 24.1 h; Col-0 = 23.6 h; Table 1, Fig 1). By contrast, in AtCCA1-ox [25] and HvCCA1-ox lines circadian rhythms of both delayed fluorescence and leaf movement were severely disrupted (Fig 1B, 1C and 1D) and there was a lower percentage of rhythmic plants (defined as RAE > 0.5; Table 1). Similarly, rhythms of stomatal conductance and photosynthesis were abolished by HvCCA1-ox and AtCCA1-ox (Table 2). Of seven HvCCA1-ox plants, only one plant had detectable rhythms of both CO2 assimilation (34.5 h, 0.4 RAE) and transpiration (24.4 h, 0.3 RAE). One other plant had rhythmic CO2 assimilation in LL (22.9 h, 0.5 RAE). In contrast, leaf gas exchange of the Ws-2 wildtype background line had robust circadian rhythms. Of eight Ws-2 plants, five had circadian rhythms of CO2 assimilation (23.7 ± 0.3 h, 0.3 ± 0.1 RAE) and four Ws-2 plants, had detectable circadian rhythms of transpiration (23.2 ± 0.3 h, 0.3 ± 0.1 RAE).
Table 1. Summary of mean circadian period estimates, standard error (SE) and percent rhythmic (%Rh) for leaf movement (LM) and delayed fluorescence (DF).
| LM data | DF Data | |||||||
|---|---|---|---|---|---|---|---|---|
| Line | Period (h) | SE | n | %Rh | Period (h) | SE | n | %Rh |
| Col | 23.6 | 0.1 | 30 | 93 | 23.6 | 0.1 | 8 | 100 |
| CCA1-ox | 26.6 | 1.0 | 30 | 70 | 24.9 | 0.9 | 8 | 38 |
| WS | 24.1 | 0.1 | 30 | 100 | 23.4 | 0.1 | 8 | 100 |
| 8–3 | 20.0 | 0.6 | 30 | 83 | 25.7 | 0.6 | 8 | 75 |
| 14–2 | 20.4 | 1.1 | 30 | 80 | N/A | N/A | N/A | N/A |
| 16–4 | 24.2 | 0.1 | 30 | 87 | 27.8 | 1.6 | 8 | 75 |
| 17–7 | 24.1 | 0.2 | 30 | 87 | 21.2 | 1.1 | 8 | 38 |
| 18–1 | 24.2 | 0.3 | 30 | 77 | 25.9 | 1.7 | 8 | 75 |
| 19–4 | 24.8 | 0.1 | 30 | 90 | 24.7 | 1.3 | 8 | 75 |
N/A–no data available. Col indicates Col-0 and WS indicates Ws-2 ecotypes.
Fig 1. Over expression of HvCCA1 causes circadian arrhythmia in Arabidopsis.

Normalised delayed chlorophyll fluorescence (a) and period estimates vs R.A.E (b) for Ws-2, Col-0, AtCCA1-ox (AtCCA1-ox 038) and two independent HvCCA1-ox transgenic lines (n = 8). Period estimates vs R.A.E for leaf movement in LL or individual leaves Col-0 and AtCCA1-ox (AtCCA1-ox 038) (c) and Ws-2 and two independent transgenic lines of HvCAA1-ox (d). n = 30. All experiments were independently repeated at least twice. Ws-2 (closed squares), Col-0 (closed triangles) AtCCA1-ox (open triangles) and two independent HvCCA1-ox transgenic lines (8–3 and 18–1) (open squares and diamonds).
Table 2. The effect of HvCCA1-ox on circadian rhythms of photosynthesis, transpiration rate and stomatal conductance in Arabidopsis.
| Photosynthesis | Transpiration rate | Stomatal conductance | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Line | Period (h) | SE | n | %Rh | Period (h) | SE | n | %Rh | Period (h) | SE | n | %Rh |
| Col | 23.5 | 0.4 | 5 | 100 | 24.6 | 0.5 | 5 | 60 | 24.7 | 2.0 | 5 | 80 |
| CCA1-ox | 26.4 | 8.5 | 5 | 45 | 26 | 2.2 | 5 | 60 | 29.2 | 0.4 | 5 | 60 |
| WS | 23.4 | 0.4 | 8 | 75 | 23.5 | 0.2 | 8 | 86 | 23.5 | 0.3 | 8 | 86 |
| 8–3 | 28.7 | 5.8 | 5 | 40 | 26.0 | 1.6 | 5 | 40 | - | - | 5 | 20 |
| 17.7 | - | - | 2 | 0 | - | - | 2 | 0 | - | - | 2 | 50 |
%Rh–percentage of rhythmic plants. “–” no rhythm could be assigned during data analysis. Col indicates Col-0 and WS indicates Ws-2 ecotypes.
Since analysis of circadian rhythms of delayed fluorescence, leaf movement, stomatal movements and photosynthesis all indicated that heterologous overexpression of barley HvCCA1 made Arabidopsis plants circadian arrhythmic, we next investigated the effects of HvCCA1-ox on the expression of the circadian oscillator genes AtCCA1, AtLHY and AtTOC1. Over-expression of HvCCA1 or AtCCA1 resulted in arrhythmic or reduced amplitude expression of the endogenous AtCCA1, AtTOC1 and AtLHY in LL (Fig 2).
Fig 2. Overexpression of HvCCA1 or AtCCA1 (AtCCA1-ox 038) in Arabidopsis abolishes rhythms of transcript abundance of (a) AtCCA1, (b) AtLHY and (c) AtTOC1.
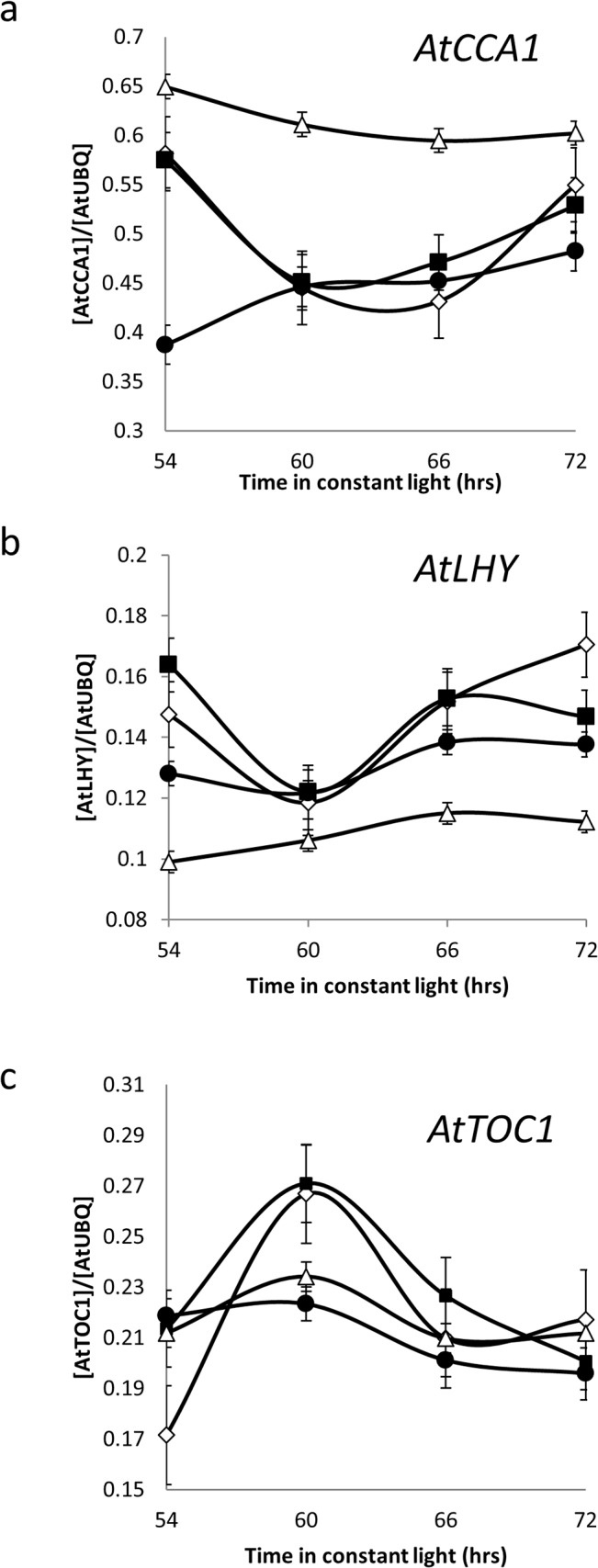
Ws-2 (white diamonds), HvCCA1-ox (line 8–3 (filled circles), Col-0 (black squares) and AtCCA1-ox 038 (white triangle). Transcript abundance was normalised to UBIQUITIN10.
Overexpression of HvCCA1 causes morphological and developmental defects in Arabidopsis
Rhythmic regulation by AtCCA1 has profound effects on Arabidopsis development, such as the elongation of the hypocotyl, and the transition from vegetative to reproductive growth. We next investigated whether HvCCA1 regulates development similar to AtCCA1. Over expression of both AtCCA1 and HvCCA1 resulted in longer hypocotyls, with those of HvCCA1-ox lines being significantly longer (3.5 to 5 mm) than the Col-0 and Ws-2 controls (1.5 mm and 2 mm; p<0.001; Fig 3A).
Fig 3. Over expression of HvCCA1 in Arabidopsis causes elongated hypocotyls and delayed flowering.

(a) Hypocotyl length was measured in WS and Col-0 background (white bars) or AtCCA1-ox (AtCCA1-ox 038) or HvCCA1-ox (indicated by the independent transgenic line numbers). Stars indicate significant difference in the hypocotyl experiments. (b) Bolting time in days (black bars) and numbers of leaves at bolting (white) in Ws-2 (WS) and Col-0 background or AtCCA1-ox (CCA1-ox) or HvCCA1-ox (indicated by the independent transgenic line numbers) when grown in 16h light, 8 h dark.
When grown under 16 h light 8 h dark, AtCCA1-ox delayed bolting and increased the number of leaves at bolt and this was phenocopied by over expression of HvCCA1. HvCCA1-ox lines flowered at least 20 days later than the wild type Ws-2 (p<0.001; Fig 3B) consistent with the CCA1-ox-038 line, which bolted approximately 30 days later than the control Col-0. The number of rosette leaves at bolting was also significantly higher in HvCCA1-ox plants compared to Ws-2 (p<0.001; Fig 3B).
Analysis of the function of HvPpd-H1 in barley
We next investigated whether HvPpd-H1 is similar in function to the Arabidopsis morning loop gene AtPRR7, with which HvPpD-H1 shares most similarity. We analysed the function of the photoperiod responsive HvPpd-H1 and photoperiod unresponsive Hvppd-H1 variants to determine whether this locus can function in circadian regulation and whether natural mutations that alter photoperiodic sensitivity can affect circadian function. Circadian rhythms of stomatal conductance and CO2 assimilation were measured in Ppd-H1 and ppd-H1 H. vulgare plants to determine if the natural mutant form ppd-H1 had a consequence for circadian rhythms in barley. No significant difference between Ppd-H1 and ppd-H1 in the period of circadian rhythms of CO2 assimilation and stomatal conductance was detected (Fig 4). Measurement of gas exchange in barley is a low throughput assay in which only one individual at a time can be recorded. We obtained 15 measurements in white light representing >75 days of continuous measurement. To ensure that the assay, despite its low-throughput nature had sufficient sensitivity to report changes in circadian period, we also investigated the effects of red light on circadian period. As expected the circadian period in equal intensity illumination was shorter under red than white light and this was most clear when assaying rhythms of stomatal conductance (Fig 4B). This demonstrated that whilst measurements of circadian rhythms of assimilation where not robust, the assay of stomatal conductance had sufficient sensitivity to detect period alteration when present (Fig 4A and 4B). The effect of both Ppd-H1 and ppd-H1 and also was monitored in red light, because it has previously been demonstrated that in Arabidopsis circadian period phenotypes in prr7-11 were more pronounced in red, rather than white light [26]. However, even in red light there was no significant difference between the period of the rhythms in Ppd-H1 and ppd-H1 (Fig 4).
Fig 4. Estimates of period of circadian rhythms of (a) photosynthesis and (b) stomatal conductance in HvPpd-H1 (filled circles) and Hvppd-H1 (open circles) in constant white light of 100 μmol m-2 s-1 (black outlined symbols) or red light (red symbols).

Each data point is derived from one individual seedling. Triangles represent the mean values.
HvPpd-H1 complements AtPRR7-11
The lack of a detectable effect of Hvppd-H1 on circadian rhythms in barley might suggest that the mutation which regulates photoperiodic sensitivity does not affect the function of the circadian clock. Additionally, this finding raises the possibility that HvPpd-H1 is not a circadian clock gene in barley, despite the sequence similarity to AtPPR7. We proceeded to use heterologous expression of barley genes in Arabidopsis to test whether HvPpd-H1 can function as a circadian clock gene. Arabidopsis Col-0 and the Atprr7-11 loss of function mutant were transformed with HvPpd-H1 and Hvppd-H1 under the control of the AtPRR7 promoter resulting in elevated expression of the transgenes (S2 Fig). We reasoned that complementation of the long circadian period phenotype of prr7-11 by heterologous expression of HvPpd-H1 would provide evidence for HvPpd-H1 acting in the circadian clock of barley.
The mean period of leaf movement rhythms of prr7-11 plants was significantly longer than that of the wildtype (prr7-11 26.6 ± 0.3 h compared to Col-0 24.9 ± 0.2 h P = 0.0046). The prr7-11 mutation was complemented by one of two lines of pAtPRR7::Ppd-H1 and both lines of pAtPRR7::ppd-H1, with the complemented lines not being significantly different from wild type, except in the case of line prr7-11-pAtPRR7:Ppd-H1-c (Table 3). Since three of the four lines complemented with natural variants of HvPpd-H1 were restored to wild type circadian period, we conclude that HvPpd-H1 in barley is likely to have a role orthologous to AtPRR7 in Arabidopsis and further supports the finding of the gas exchange analyses that Hvppd-H1 mutation is without effect on circadian rhythms.
Table 3. Summary of circadian period estimates for leaf movement in Col-0, prr7-11 and prr7-11 transformed with either pPRR7::Ppd-H1 or pPRR7::ppd-H1.
| Line | Period (h) | SEM | RAE | n | %Rh |
|---|---|---|---|---|---|
| Col-0 | 24.9 | 0.2 | 0.2 | 32 | 75 |
| prr7-11* | 26.6 | 0.3 | 0.2 | 29 | 59 |
| prr7-11 pAtPRR7:Ppd-H1-c | 25.8 | 0.3 | 0.2 | 35 | 69 |
| prr7-11 pAtPRR7:Ppd-H1-d * | 25.0 | 0.3 | 0.2 | 25 | 64 |
| prr7-11 pAtPRR7:ppd-H1-e * | 25.3 | 0.3 | 0.2 | 21 | 86 |
| prr7-11 pAtPRR7:ppd-H1-f * | 25.6 | 0.3 | 0.2 | 29 | 59 |
* indicates significant difference at 5% level compared to background.
The background for prr7-11 is Col-0. The background for the complemented lines is prr7-11. SEM = standard error of the mean. %Rh = Percentage of rhythmic seedlings.
Discussion
We provide experimental evidence that HvCCA1 and HvPpd-H1 have functionality similar to AtCCA1 and AtPRR7 in Arabidopsis. Heterologous expression in Arabidopsis of HvCCA1-ox results in circadian arrhythmia and morphological changes consistent with HvCCA1 being functionally equivalent to AtCCA1. Our data show that constitutive expression of HvCCA1 in Arabidopsis causes arrhythmia in leaf movement, delayed fluorescence and several circadian clock genes. It also causes significant hypocotyl elongation and late flowering in HvCCA1 transformants. These results are consistent with AtCCA1-ox Arabidopsis data and provide a strong support that CCA1 and its function are highly conserved throughout the plant kingdom. Expression of either HvPpd-H1 or Hvppd-H1 under control of the endogenous AtPRR7 promoter partially restores the wildtype period of circadian leaf movement rhythms in an AtPRR7 tDNA insertion mutant (prr7-11) suggesting that the function of Hv-Ppd1 is as a circadian clock gene. A similar experiment in rice provided evidence that the OsPRR37 and AtPRR7 genes are partially interchangeable [27]. Together these findings show that the clock-associated functions of CCA1 and PRR7 are conserved between Arabidopsis, rice and barley.
Since both the photoperiodic-sensitive and–insensitive alleles of Hv-Ppd1 were able to complement the prr7-11 phenotype, this suggested that the causative SNP affects photoperiodism independent of circadian rhythms. To test this directly, circadian rhythms of gas exchange from a Triumph (ppd-H1) H. vulgare line were compared to those from the same line into which the Igri Ppd-H1 allele had been introgressed and it was found that the circadian period was not significantly altered between barley plants carrying the Hv-Ppd1 and Hv-ppd1 alleles. Whilst mutation in HvPpd-H1 did not produce detectable effects on circadian function either in barley or through complementation tests in Arabidopsis Atprr-11, the Hvppd-H1 allele does abolish circadian rhythms of photoperiodic-response and vernalisation-response genes HvCO1, HvCO2 and Vrn-H1 [12]. These findings coupled with our demonstration that both HvPpd1-H1 and Hvppd-H1 can complement AtPrr7-11 suggest that HvPpd-H1 has at least two regulatory roles, one required for proper functioning of the circadian system, and one required for generation of rhythmic outputs, and it is the latter that is compromised in Hvppd-H1.
The analysis of circadian rhythms in barley and the other grain crops is of critical importance, since domestication has required breeders and early farmers to select for varieties with altered photoperiodic sensitivity and adaptations for cold tolerance. These are both traits that are regulated by the circadian clock [1, 28]. We have described the use of gas exchange and heterologous expression in Arabidopsis circadian mutants as new tools for the dissection of the barley circadian signalling network and determined that HvPpd-H1 is functionally ortholgous to AtPRR7.
Supporting Information
Seedlings were entrained for one week in 12 light/12hr dark cycles at 22°C then transferred to constant light. Samples were taken 3 h after dawn. Lines used were Ws-2, Col-0, AtCCA1-ox 038 and HvCCA1-ox (8–3 and 18–1). Transcript abundance was normalised to Ubiquitin10.
(PNG)
{kind=link}
(PNG)
{kind=link}
Forward (F) and reverse orientation (R) primers.
(PDF)
Acknowledgments
ZR is grateful to the National Institute of Agricultural Botany for the award of a Scholarship, We acknowledge funding from a Marie Curie Early Stage Training project MEST-CT-2005- 020526 for JK and the BBSRC-DTP for funding SC. AARW and MCM are grateful to the BBSRC for the award of BBSRC Grant BB/M006212/1, which supported aspects of the study.
Data Availability
Relevant data are available at: https://www.repository.cam.ac.uk/handle/1810/247449.
Funding Statement
ZR is grateful to the National Institute of Agricultural Botany for the award of a Scholarship. The authors acknowledge funding from a Marie Curie Early Stage Training project MEST-CT-2005-020526 for JK, and the Biology and the Biological Sciences Research Council Doctoral Training Programme for SC. AARW and MCM are grateful to the BBSRC for the award of BBSRC Grant BB/M006212/1, which supported aspects of the study. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Harmer SL (2009) The circadian system in higher plants. Ann. Rev. Plant Biol. 60: 357–377. [DOI] [PubMed] [Google Scholar]
- 2. Dodd AN, Salathia N, Hall A, Kévei E, Tóth R, Nagy F, et al. (2005) Plant circadian clocks improve growth, competitive advantage and survival. Science 309: 630–633. [DOI] [PubMed] [Google Scholar]
- 3. Green RM, Tingay S, Wang ZY, Tobin EM (2002) Circadian rhythms confer a higher level of fitness to Arabidopsis plants. Plant Phys. 129: 576–584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Johnson CH, Kyriacou CP (2007) Clock evolution and adaptation: Whence and whither? Ann. Plant Rev. 21: 237–260. [Google Scholar]
- 5. Locke JC, Kozma-Bognár L, Gould PD, Fehér B, Kevei E, Nagy F, et al. (2006) Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana. Mol. Sys Biol., 2(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Gendron JM, Pruneda-Paz JL, Doherty CJ, Gross AM, Kang SE, Kay SA (2012) Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proceedings of the National Academy of Sciences, 109, 3167–3172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Huang W, Perez-Garcia P, Pokhilko A, Millar AJ, Antoshechkin I, Riechmann JL, et al. (2012) Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science, 336: 75–79. 10.1126/science.1219075 [DOI] [PubMed] [Google Scholar]
- 8. Herrero E, Kolmos E, Bujdoso N, Yuan Y, Wang M, Berns M, et al. (2012) EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell, 24: 428–443. 10.1105/tpc.111.093807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Serikawa M, Miwa K, Kondo T, Oyama T (2008) Functional conservation of clock-related genes in flowering plants: Overexpression and RNA interference analyses of the circadian rhythm in the monocotyledon Lemna gibba. Plant Phys. 146: 1952–1963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Matsushika A, Makino S, Kojima M, Mizuno T (2000) Circadian waves of expression of the PRR1/TOC1 family of Pseudo-response regulators of A.thaliana: insight into the plant circadian clock. Plant Cell Physiol. 41: 1002–1012. [DOI] [PubMed] [Google Scholar]
- 11. Ogiso E, Takahashi Y, Sasaki T, Yano M, Izawa T (2010) The role of casein kinase II in flowering time regulation has diversified during evolution. Plant Phys. 152: 808–820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Campoli C, Shtaya M, Davis SJ, von Korff M (2012) Expression conservation within the circadian clock of a monocot: natural variation at barley Ppd-H1 affects circadian expression of flowering time genes, but not clock orthologs. BMC Plant Biol. 12: 97 10.1186/1471-2229-12-97 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Campoli C, Pankin A, Drosse B, Casao CM, Davis SJ, Korff M (2013) HvLUX1 is a candidate gene underlying the early maturity 10 locus in barley: phylogeny, diversity, and interactions with the circadian clock and photoperiodic pathways. New Phytologist 199: 1045–1059. 10.1111/nph.12346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Faure S, Turner AS, Gruszka D, Christodoulou V, Davis SJ, Korff M, et al. (2012) Mutation at the circadian clock gene EARLY MATURITY 8 adapts domesticated barley (Hordeum vulgare) to short growing seasons. Proc. Natl. Acad. Sci. (USA) 109: 8328–8333. 10.1073/pnas.1120496109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Turner A, Beales J, Faure S, Dunford RP, Laurie DA (2005) The Pseudo-Response Regulator Ppd-H1 Provides Adaptation to Photoperiod in Barley. Science 310: 1031–1034. [DOI] [PubMed] [Google Scholar]
- 16. Curtis MD, Grossniklaus U (2003) A gateway cloning vector set for high-throughput functional analysis of genes in planta . Plant Physiology 133: 462–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium‐mediated transformation of Arabidopsis thaliana . Plant J 16: 735–743. [DOI] [PubMed] [Google Scholar]
- 18. Schultz TF, Kiyosue T, Yanovsky M, Wada M, Kay SA (2001) A Role for LKP2 in the Circadian Clock of Arabidopsis. Plant Cell 13: 2659–2670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. LeClerc S, Bartel B (2001) A library of Arabidopsis 35S-cDNA lines for identifying novel mutants. Plant Mol. Biol. 46: 695–703. [DOI] [PubMed] [Google Scholar]
- 20. Karimi M, Inze D, Depicker A (2002) GATEWAY vectors for Agrobacterium mediated plant transformation. Trends Plant Sci. 7: 193–195. [DOI] [PubMed] [Google Scholar]
- 21. Ramakers C, Ruijter JM, Lekanne Deperz RH, Moorman AFM (2003) Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neuroscience Letters 339: 62–66. [DOI] [PubMed] [Google Scholar]
- 22. Gould PD, Diaz P, Hogben C, Kusakina J, Salem R, Hartwell J, et al. (2009) Delayed fluorescence as a universal tool for the measurement of circadian rhythms in higher plants. Plant J. 58: 893–901. 10.1111/j.1365-313X.2009.03819.x [DOI] [PubMed] [Google Scholar]
- 23. Plautz JD, Straume M, Stanewsky R, Jamison CF, Brandes C, Dowse HB, et al. (1997) Quantitative analysis of Drosophila period gene transcription in living animals. J. Biol. Rhyth. 12: 204–217. [DOI] [PubMed] [Google Scholar]
- 24. Dodd AN, Parkinson K, Webb AAR (2004) Independent circadian regulation of assimilation and stomatal conductance in the ztl-1 mutant of Arabidopsis. New Phyt. 162: 63–70. [Google Scholar]
- 25. Wang ZY, Tobin EM (1998) Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 93: 1207–1217. [DOI] [PubMed] [Google Scholar]
- 26. Farré EM, Harmer SL, Harmon FG, Yanovsky MJ, Kay SA (2005) Overlapping and distinct roles of PRR7 and PRR9 in the Arabidopsis circadian clock. Curr. Biol. 15: 47–54. [DOI] [PubMed] [Google Scholar]
- 27. Murakami M, Tago Y, Yamashino T, Mizuno T (2007) Characterization of the rice circadian clock-associated pseudo-response regulators in Arabidopsis thaliana. Bioscience, Biotechnology, and Biochemistry. 71: 1107–1110. [DOI] [PubMed] [Google Scholar]
- 28. Dodd AN, Kyed Jakobsen M, Baker AJ, Telzerow A, Hou S-W, Laplaze L, et al. (2006) Time of day modulation of Ca2+ signals in Arabidopsis. Plant J. 48: 962–973. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Seedlings were entrained for one week in 12 light/12hr dark cycles at 22°C then transferred to constant light. Samples were taken 3 h after dawn. Lines used were Ws-2, Col-0, AtCCA1-ox 038 and HvCCA1-ox (8–3 and 18–1). Transcript abundance was normalised to Ubiquitin10.
(PNG)
(PNG)
Forward (F) and reverse orientation (R) primers.
(PDF)
Data Availability Statement
Relevant data are available at: https://www.repository.cam.ac.uk/handle/1810/247449.