

Abstract
Haemolytic uraemic syndrome (HUS) is characterized by haemolytic anaemia, thrombocytopenia and acute renal failure. We studied the activation state of classical and alternative pathways of complement during the acute phase of Shiga toxin-associated HUS by performing a prospective study of 18 patients and 17 age-matched healthy controls to evaluate C3, C3c, C4, C4d, Bb and SC5b-9 levels. SC5b-9 levels were increased significantly in all patients at admission compared to healthy and end-stage renal disease controls, but were significantly higher in patients presenting with oliguria compared to those with preserved diuresis. C3 and C4 levels were elevated significantly at admission in the non-oliguric group when compared to controls. No significant differences were found for C4d values, whereas factor Bb was elevated in all patients and significantly higher in oliguric patients when compared to both controls and non-oliguric individuals. A positive and significant association was detected when Bb formation was plotted as a function of plasma SC5b-9 at admission. Bb levels declined rapidly during the first week, with values not significantly different from controls by days 3 and 5 for non-oligurics and oligurics, respectively. Our data demonstrate the activation of the alternative pathway of complement during the acute phase of Stx-associated HUS. This finding suggests that complement activation may represent an important trigger for the cell damage that occurs during the syndrome.
Keywords: acute renal injury, alternative pathway, complement, HUS
Introduction
Shiga toxin-associated haemolytic uraemic syndrome (Stx-HUS) is the first and second most frequent cause of acute renal injury and end-stage renal disease (ESRD) in Argentine children, respectively 1. In Argentina, 12–14 cases for every 100 000 children under 5 years of age are reported every year 2. Thus, the cumulative incidence of the disease has recently reached 500 new cases per year 3.
Clinically, children first present with bloody diarrhoea and then progress within 3–7 days to acute kidney injury, haemolytic anaemia and thrombocytopenia. Outcome is only apparently good: 3–5% of all affected children die during the acute phase and a similar percentage never recover renal function after the acute episode or transiently recover but develop ESRD. Approximately 30% have residual proteinuria and may develop evidence of chronic kidney disease in the long term. The remaining 60% of children with apparent full recovery remain at risk of developing ESRD between 15 and 25 years after the acute phase, possibly by progressive renal function deterioration due to maladaptive mechanisms initiated and sustained by the initial injury during acute disease 4. Severe central nervous system involvement and, more importantly, the duration of the acute oliguric phase, are good predictors of the likelihood of chronicity and mortality.
There are, however, cases of HUS not caused by infection by Shiga toxin-producing bacteria 5. Atypical HUS (aHUS) is the term used to classify any HUS of non-infectious nature. Although sharing most features at presentation and during the acute phase of the disease, the aetiology of aHUS centres on various genetic or acquired disorders of complement regulation and of the Von Willebrand protease ADAMTS13 (a disintegrin and metalloproteinase with a thrombospondin type 1 motif, member 13), among other causation factors 6. Mutations in different complement regulatory genes or the presence of antibodies against members of the complement regulatory system have been described in aHUS-affected patients, thus implicating the complement system in disease development 7,8.
The classical lectin and alternative pathways of complement are activated by different stimuli in various diseases and conditions. The different pathways converge to a common step, leading to the generation of the membrane terminal attack complex (MAC). MAC exists in two forms: the membrane-bound form, which can be found on cells and tissues, and the fluid phase SC5b-9 complex, which can be detected in body fluids 9.
While initial studies in patients with Stx-HUS showed normal levels of factors C3 and C4 10, others reported perturbations in serum levels of different complement components 11–15. More recent evidence on the role of complement in Stx-HUS arises from clinical studies showing activation of the alternative pathway 16, elevations of C3a 17 and SC5b-9 18. In line with those results, Orth et al. have demonstrated a direct activation of the complement cascade by purified Stx2 as well as a delayed co-factor activity of factor H on the cell surface when bound to Stx2 19. Furthermore, an improving effect of a monoclonal antibody against complement component 5 (C5) on a small group of severely affected Stx-HUS patients is also indicative of complement activation during disease 20. However, no data are currently available regarding the sequential contribution of the classical, lectin and alternative pathways of complement in Stx-HUS patients. In view of the established role of complement activation in aHUS development and on the contradictory data available for the typical form of the disease, a comprehensive exploration of complement status during Stx-induced HUS is of high relevance. We thus aimed here to study the activation state of both classical/lectin and alternative pathways of complement during the acute phase of endemic HUS. We performed a prospective study where we evaluated C3, C3c, C4, C4d, Bb, and SC5b-9 levels in order to determine the contribution of the classical/lectin and alternative pathways in the generation of MAC and the relationship between complement activation with clinical and laboratory parameters in patients with acute Stx-HUS. The determination of the relative contribution of the different pathways to overall complement activation may provide a clue about the possible initiating events of the disease and lead also to improvements in its clinical management.
Materials and methods
Subjects
Eighteen consecutive patients diagnosed with Stx-HUS and treated at the Pediatric Department of the Hospital Italiano de Buenos Aires between November 2006 and August 2007 were included in the present study. The clinical diagnosis of Stx-HUS was based on acute kidney injury, microangiopathic anaemia and thrombocytopenia following a prodromal phase of bloody diarrhoea. During the acute phase of Stx-HUS, all children were treated with intermittent peritoneal dialysis and/or haemodialysis and blood transfusion when required. Plasma therapy was used in patients with severe neurological signs (seizures, coma, nystagmus, impaired consciousness). Microbiological confirmation of Stx-producing Escherichia coli infection was required to enter the study and was performed by polymerase chain reaction (PCR) determination of Shiga toxin in stool cultures. Samples for establishing reference values were taken from 17 age-matched healthy volunteers. The inclusion criteria for healthy volunteers were no history of or no active renal disease or haemolytic anaemia, no active infectious disease, no medications and standard diet.
For comparison, 29 children with ESRD not caused by HUS or C3 glomerulonephritis under regular haemodialysis were also included in the study. The inclusion criterion for this group was no active disease.
Children were included after written informed consent was obtained from their parents, following recommendations of the World Medical Association. The study protocol, which adhered to the Declaration of Helsinki, was approved by the hospital's ethics committee.
Biological samples and collection
Blood was collected daily at 6 a.m. during the patients' hospital stay. For patients receiving dialysis and/or plasma therapy, blood was always drawn prior to the procedure. For healthy controls, a single sample was derived from the blood obtained after an extraction indicated during the course of a medical examination of subjects meeting the inclusion criteria. In both cases, blood was collected by venipuncture into polypropylene tubes containing ethylenediamine tetraacetic acid (EDTA). After centrifugation at 4°C for 10 min at 1500 g, the obtained plasma was removed carefully, aliquoted and frozen at −75°C until used.
Bb, SC5b-9 and C4d assays
Plasma levels of complement activated factors Bb, SC5b-9 (soluble C5b-9 bound) and C4d were determined by enzyme-linked immunosorbent assay using MicroVue Bb Plus, SC5b-9 and C4d enzyme immunoassay (EIA) kits (Quidel Corporation, San Diego, CA, USA), respectively, and according to the manufacturer's instructions. Briefly, plasma specimens in appropriate dilutions were added to EIA wells coated with monoclonal antibody to the respective complement product and incubated for 30 min. After washing, a second antibody conjugated with horseradish peroxidase was added. This was followed by addition of a chromogenic substrate; the reaction was stopped after 30 min and the absorbance read at 450 or 405 nm in a Synergy HT multiplate reader (BioTek Instruments, Inc., Winooski, VT, USA). For each analyte, concentrations in the test samples were read from a standard plot between absorbance and concentration of a set of standards.
C3 and C4 determination
Serum C3 and C4 were measured by kinetic immunonephelometry using polyclonal goat anti-human sera in a Beckman Coulter Image 800, according to standard laboratory techniques based on the manufacturer's instructions.
C3c determination
Serum C3c levels were determined by immunofixation using a specific antibody (Dako, Glostrup, Denmark), as described previously 21. Obtained serum was separated by agarose electrophoresis using the Hydrasys (Sebia, Norcross, GA, USA) semi-automated method. Separated proteins were incubated with the anti-C3c antibody and the resulting C3 and C3c bands visualized by staining the gel with acid violet. The results were expressed as percentage of total C3 for each patient.
Standard clinical laboratory determinations
The following laboratory determinations were performed throughout the hospital stay as part of the routine control: haemogram, platelet count, plasma urea, plasma creatinine, lactate dehydrogenase (LDH), ionogram and acid-base status. Determinations were performed at the Hospital Clinical Laboratory according to standardized laboratory practice.
Statistics
Unless stated otherwise, data are expressed as the mean ± standard error of the mean (s.e.m.). Significant differences were determined by one- or two-way analyses of variance (anova) followed by Bonferroni or one-tailed Dunnett's t-test as post-hoc tests, or by independent-samples t-test. Differences were deemed significant when P was less than 0·05.
Results
The subjects included in this study were all diagnosed with the typical form of HUS and presented with a diarrhoeal prodrome. Median age was 25 months, with a slight difference for girls over boys (10/8). Of the 18 patients studied, five presented with conserved diuresis while the other 13 showed oligoanuria at admission or developed the condition thereafter. Thus, according to the degree of renal impairment upon presentation, patients were categorized as oliguric or non-oliguric.
The main demographic features of the subjects under study, as well as salient disease course data, are presented in Table1. Expectedly, oliguric patients required longer hospitalization, dialysis (peritoneal dialysis or haemodialysis) and platelet or packed red blood cell (RBC) transfusions. The six patients who developed neurological signs (two seizures and six impaired consciousness) required haemodialysis, but evolved to complete recovery. None of the patients included in this study developed ESRD during follow-up. However, six patients developed proteinuria, including three with estimated filtration rates of 80, 61 and 50 ml/min/1·73 m2, after 3, 6 and 12 months, respectively.
Table 1.
Demographic features and outcome
| Non-oliguric | Oliguric | |
|---|---|---|
| Patients (male/female) | 5 (4/1) | 13 (4/9) |
| Age, months (range) | 23 (8–57) | 27 (7–92) |
| Hospital stay, days (range) | 7 (5–8) | 15 (10–44) |
| Neurological signs | 0 | 6 |
| Dialysis (HD/PD) | 0/0 | 6/7 |
| Dialysis, days (range) | 0 | 7 (1–100) |
| Transfusions | ||
| Red blood cells | 5 | 13 |
| Platelets | 0 | 7 |
| Plasma therapy* | 0 | 7 |
| Survival | 100% | 100% |
Patient age, hospital stay and days under dialysis are expressed as median months and days, respectively, and the range of observations is given. The variables neurological signs, dialysis and transfusions are shown as number of patients in the respective group. HD = haemodialysis; PD = peritoneal dialysis.
Plasma therapy = plasma infusion five patients and plasmapheresis two patients.
SC5b-9 levels are a measure of the amount of the cytolytic membrane attack complex generated as a result of activation of the common pathway by either the classical, lectin or alternative pathways and is thus a sensitive indicator of overall complement activation. We therefore measured the amount of SC5b-9 complex in plasma, and a profile of the first week after hospital admission is shown in Fig. 1. As observed, SC5b-9 levels at admission were elevated significantly over those of healthy individuals (295·6 ± 23·5 ng/ml), with oliguric patients exhibiting higher SC5b-9 values than non-oliguric patients. Plasma SC5b-9 levels declined rapidly during the first week, with values not significantly different from controls by day 3 in the case of non-oliguric patients, and by day 6 for the more severely compromised individuals. A two-way anova of the data revealed highly significant differences (P < 0·001) for both variable day and condition (oliguric or non-oliguric), but not for the interaction of those variables.
Figure 1.

Daily profile of SC5b-9 activation in haemolytic uraemic syndrome (HUS)-affected children. SC5b-9 activation was determined by enzyme-linked immunosorbent assay (ELISA) in the plasma of HUS-affected children. Obtained values for the different patients were pooled according to the day of evolution from hospitalization. Results are expressed as mean SC5b-9 (ng/ml) ± standard error of the mean (s.e.m.). Filled dots: oliguric group; open dots: non-oliguric group. Control range is indicated by solid (mean) and dotted (s.e.m.) lines for comparison. (a) P < 0·01 versus control; (b) P < 0·05 versus control by independent-samples t-test; (c) P < 0·01 versus day 1 by analysis of variance.
In view of these results, we determined the levels of C3 and C4 factors in serum by nephelometry. C3 was elevated significantly at admission in the non-oliguric group of patients, whereas subjects with impaired diuresis showed values comparable to those of healthy individuals (Fig. 2a). Similar results were registered for C4, with non-oliguric patients exhibiting significantly elevated C4 levels over controls, although within the accepted reference interval of 16–38 mg/dl (Fig. 2b).
Figure 2.
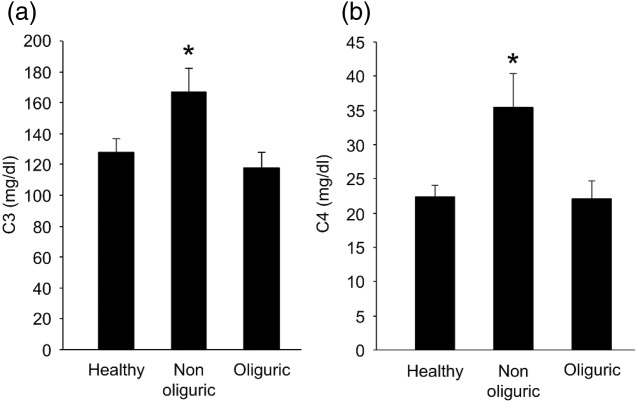
Complement components 3 and 4 activation. C3 (a) and C4 (b) levels were determined in serum by nephelometry. Results represent the mean ± standard error of the mean (s.e.m.) of healthy controls and of non-oliguric and oliguric patients and are expressed in mg/dl. *P < 0·05 by independent-samples t-test.
The activation of the classical pathway of complement was next studied by determining C4d levels. As shown in Fig. 3, neither group of patients showed significant differences in C4d values, thus ruling out the activation of the classical pathway.
Figure 3.

Daily profile of C4d levels in haemolytic uraemic syndrome (HUS)-affected children. C4d levels were determined by enzyme-linked immunosorbent assay (ELISA) in the plasma of HUS-affected children. Obtained values for the different patients were pooled according to the day of evolution from hospitalization. Results are expressed as mean C4d (μg/ml) ± standard error of the mean (s.e.m.). Filled dots: oliguric group; open dots: non-oliguric group. Control range is indicated by solid (mean) and dotted (s.e.m.) lines for comparison.
The status of the alternate pathway of complement was then tested by measuring Bb levels. As observed for the daily profile of Bb activation shown in Fig. 4, all patients were admitted with values significantly higher than those of controls (0·98 ± 0·2 μg/ml), and those who presented with oligoanuria showed increased levels compared to those with preserved diuresis (P < 0.05). A two-way anova of the data resulted in highly significant differences (P < 0·001) for both variables day and condition, but not for the interaction of those variables.
Figure 4.

Daily profile of Bb activation in haemolytic uraemic syndrome (HUS)-affected children. Bb activation was determined by enzyme-linked immunosorbent assay (ELISA) in the plasma of HUS-affected children. Obtained values for the different patients were pooled according to the day of evolution from hospitalization. Results are expressed as mean Bb (μg/ml) ± standard error of the mean (s.e.m.). Filled dots: oliguric group; open dots: non-oliguric group. Control range is indicated by solid (mean) and dotted (s.e.m.) lines for comparison. (a) P < 0·01 versus control; (b) P < 0·05 versus control by independent-samples t-test; (c) P < 0·05; (d) P < 0·01 versus day 1 by analysis of variance.
A positive and significant association was detected when Bb formation was plotted as a function of the amount of SC5b-9 detected in the plasma of patients at admission day (Fig. 5). Bb values also correlated with blood urea nitrogen (BUN) (r = 0·464; P < 0·01), lactate dehydrogenase (LDH) activity (r = 0·799; P < 0·01) and thrombocyte count (r = −0·370; P < 0·01) values during the disease course.
Figure 5.

Correlation between SC5b-9 and Bb activation in haemolytic uraemic syndrome (HUS)-affected children. Bb values were plotted against the respective SC5b-9 levels at admission for each patient. The lines represent the results of linear regression and values of the Pearson's correlation coefficient.
Finally, SC5b-9 and Bb values at hospitalization were compared with those obtained from a group of patients with ESRD not caused by HUS or C3 glomerulonephritis and undergoing regular haemodialysis. As observed in Fig. 6a,b, the ESRD group (n = 29) showed significantly elevated levels of both SC5b-9 and Bb when compared to healthy controls, while the oliguric group of HUS-affected patients exhibited a highly significant increase in both variables over control and ESRD groups.
Figure 6.

SC5b-9 and Bb activation. SC5b-9 (a) and Bb (b) activation was determined by enzyme-linked immunosorbent assay (ELISA) in the plasma of both non-oliguric and oliguric groups of haemolytic uraemic syndrome (HUS)-affected children, as well as in healthy controls (healthy) and in end-stage renal disease patients (ESRD). Results are expressed as mean SC5b-9 (ng/ml) or Bb (μg/ml) ± standard error of the mean (s.e.m.). (a), P < 0·05; (b), P < 0·01 versus healthy; (c), P < 0·01 versus oliguric, by independent-samples t-test.
It is noteworthy that no significant differences were found in the analysed complement parameters for the six patients with severe neurological signs when compared to the rest of the oliguric group.
Discussion
The results of the present work demonstrate that all individuals suffering from Stx-induced HUS show activation of the complement system via the alternative pathway during the acute phase of the disease.
Our study included 18 patients diagnosed clinically and microbiologically with Stx-associated HUS in whom we performed daily determinations of various laboratory parameters, as well as a comprehensive complement system exploration. First, we have determined the levels of circulating SC5b-9, a measure of generated MAC, a sensitive indicator of the overall complement activation. We found that all included individuals, irrespective of the condition at hospital admission, presented levels significantly elevated over our established reference values. However, the severity of the disease influenced this parameter, as individuals categorized in the oliguric group showed higher values than those with preserved diuresis (non-oliguric group) throughout the evaluation period (Fig. 1). A two-way anova of the data revealed highly significant differences for both variables day and condition (oliguric or non-oliguric), but not for the interaction of those variables, thus indicating that in all the individuals SC5b-9 is elevated initially regardless of the condition and that formation of membrane attack complex declines during the evaluated period in a similar fashion.
Complement activation can occur by the classical, lectin or alternative pathway, or by a combination of those. C4 and C3 levels can be used to make a distinction between the classical/lectin and alternative pathway. A recent study has confirmed C3 reduction in severe cases of Stx-HUS 20. Low levels of C4 have also been observed occasionally in patients with Stx-HUS, indicating activation of the classical and/or lectin pathways leading to C4 consumption 18. However, in our study, the non-oliguric group of patients showed statistically significant elevations of both C3 and C4 levels over healthy controls and oliguric patients. C3 and C4 levels are commonly expected to decrease due to consumption of the factor during complement activation. Our finding was thus unexpected, particularly with regard to C3. However, C3 and C4 are acute phase proteins, which are synthesized by stimulation of proinflammatory cytokines 22. It may then be possible that the levels of C3 may increase in this set of patients due to the important inflammatory component characteristic of HUS. Also, although its synthesis occurs mainly in the liver, several other cell types, including vascular cells, produce C3 22. The vascular component of HUS, along with the massive erythrocyte and thrombocyte destruction that occurs during the acute phase of the syndrome, may also explain altered levels of C3 and C4. Several conditions and diseases are known to associate with elevated C3 levels including myocardial infarction 23 and hypertension 24. The question still remains as to why only patients with preserved diuresis show elevations of C3 and C4. One possible interpretation is that the consumption rate of C3 and C4 is greater in the more severe form of the disease, and patients with that presentation would tend to show normal or even reduced levels of the factors. Also, considering the relatively wide ranges of C3 and C4 concentrations, a 10–20% cleavage would leave enough C3 to be well within the normal range. Hence, measuring only the cleavage products of these proteins, one can estimate the extent of an ongoing activation process in the early stage of acute Stx-HUS. We thus set out to study the breakdown products of C3 and C4 (C3c and C4d, respectively), and found only a mild increment trend in C3c levels for both groups of patients over controls (data not shown) but similar values for C4d in all investigated groups. We interpret these results as a negative indication for the participation of the classical and lectin pathways of complement in Stx-induced HUS. Activation of the alternative pathway of complement was confirmed in our study by means of measuring the levels of fragment Bb. As observed for SC5b-9, all individuals showed significantly elevated levels over controls at admission that declined rapidly during the first week of hospitalization, and attained values not significantly different from controls by days 4 and 6 in the non-oliguric and oliguric groups, respectively.
Thus far, our data suggest that the alternative pathway is the only contributing factor to complement activation in patients with acute Stx-HUS. This observation is based on the fact that C4d values, used as a measure of the activation of the classical and lectin pathways, were always normal, whereas factor Bb levels, that reflect the activation of the alternative pathway, were elevated in both groups of patients. Our observation is also reinforced by the positive and significant association detected when Bb levels were plotted as a function of formed SC5b-9.
After the pathogenic recognition of Stx-induced-HUS, two recent clinical studies confirmed complement activation. In 17 children with Stx-HUS studied during the acute phase, plasma levels of Bb and Sc5b-9 were increased at admission but then normalized by day 28 after discharge 16. Similar results were obtained in a cohort of 12 children with elevated plasma levels of C3a and SC5b-9 at admission that returned to normal 2–9 months after recovery 18. Our results agree with those reported although it should be noted that, in those two investigations, complement components were only assayed in plasma at the time of admission and resolution. We also show here increased levels of breakdown products of two components of the alternative pathway (C3c and Bb) and normal levels of one component of the classical and lectin pathways (C4d) in our patients with acute Stx-HUS. More importantly, we have measured complement components on a daily basis and could, thus, observe the rapid return of Bb and SC5b-9 levels to normal values within the first week after admission, which in some cases is well before the resolution of the acute episode. Interestingly, few patients (three) undergoing haemodialysis exhibited elevated Sc5b-9 with normal Bb levels by the time all other individuals had normalized their values.
Complement activation may contribute to the pathophysiology of typical HUS by different mechanisms. First, purified Stx-2 has been shown to selectively bind complement factor H and also to activate complement in the fluid phase when co-incubated with normal human serum 19. This demonstrates that Stx activates complement by a direct effect and also by reducing the activity of the regulatory factor, all of which may result in complement hyperactivation with the consequent damage of host cells 19. Secondly, the role of lipopolysaccharide (LPS) in inducing complement activation during E. coli infection cannot be excluded because, in a mouse HUS model, co-injection of Stx and LPS is required for both complement activation and disease development 17. Also, LPS derived from enterohaemorrhagic E. coli binds to the membrane adhesion molecule P-selectin in platelets in HUS patients 25, and P-selectin is known to bind C3b with high affinity, which could then trigger the alternative pathway 26.
Complement activation could also be the consequence of the renal failure that occurs during HUS, and we found here activation in a group of patients suffering ESRD not caused by HUS or C3 glomerulonephritis. The fact that ESRD patients exhibited a significant activation when compared to healthy controls may point to a non-specific complement activation due to renal failure irrespective of its cause. However, patients included in this study showed a significant activation of the alternative pathway of complement and the oliguric group exhibited Bb and SC5b-9 levels well above those of the ESRD group. Notably, complement status normalizes in HUS patients within days coinciding with a favorable resolution of the acute phase of the disease. Also, patients with a more severe disease presentation normalize complement status rapidly even though they remain hospitalized and requiring dialysis for longer periods of time.
We have reported recently on increased oxidative stress in the plasma of this same cohort of HUS-affected children 27. Interestingly, a number of reports have established that oxidative stress results in complement activation and deposition on the vascular endothelium 28–31 and it is now accepted that complement plays an important role in the inflammatory process following oxidative stress. The available data point to the role of the lectin pathway in ischaemia/reperfusion-induced complement activation mainly in the myocardium and gastrointestinal system, although it has been demonstrated that oxidative stress modulates factor H expression in retinal pigmented epithelial cells, a mechanism implicated in the development of age-related macular degeneration 32. Even more intriguing is the observation that oxidative stress mediates the symptoms of paroxysmal nocturnal haemoglobinuria 33, a rare disease linked to complement activation via the alternative pathway where the use of eculizumab is now established. In view of our results, it is of interest to explore the connection between oxidative stress and complement activation via the alternative pathway in typical HUS and the possible implications for disease development.
An association between complement activation and laboratory findings was observed in our Stx-HUS patients. Bb values correlated significantly with the classical clinical triad: haemolytic anaemia (in this case its surrogate LDH), thrombocytopenia and acute renal injury (BUN and serum creatinine). Moreover, Bb and Sc5b-9 levels were higher in severe Stx-HUS oliguric patients at admission and also during the first week of hospitalization in the oliguric when compared to the non-oliguric group of patients. Finally, almost 50% of oliguric patients with higher Bb and Sc5b-9 levels at admission developed chronic kidney disease (CKD) during follow-up. Therefore, the activity of the alternative pathway is a useful index of acute kidney injury and a prognostic marker of CKD.
Interestingly, very severe forms of HUS have been reported in children with factor H or membrane cofactor protein mutations after exposure to Stx-producing bacteria. We did not study complement regulatory proteins in our work; however, none of our patients and/or their family members had HUS before or after the actual episode.
Emerging evidence suggests that microbes (group A β-haemolytic streptococcus, Bordetella pertussis, Fusobacterium necrophorum, influenza A, parvovirus B19, HIV and varicella viruses) associated with HUS exhibit interaction with the complement system. However, other infectious agents produce complement activation but do not induce HUS. This may be indicative of an interplay between the toxin and complement activation for the development of HUS. Also, the genetic background of affected individuals may explain a differential susceptibility to the pathogen. Both possibilities are worth exploring.
In conclusion, our data indicate that, during the first days of the acute phase of Stx-HUS, disease progresses with activation of the alternative pathway of complement which is possibly a pathogenic effector leading to thrombocytopenia, haemolytic anemia and renal dysfunction. Also, our results indicate that the intensity of activation of the alternative pathway of complement should be a prognostic factor in the short- and long-term follow-up.
Acknowledgments
Financial support from Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 11420090100389) to C. F. M. is acknowledged.
Author contributions
J. R. F. and C. F. M. designed the study, analysed the data and wrote the paper; V. F. supervised the clinical aspects of the study and compiled the clinical and laboratory data; A. B. A. performed Bb, SC5b-9 and C4d determinations; P. S. and M. S. S. performed the clinical laboratory and nephelometric determinations, and A. G. perfomed C3 and C3c readings.
Disclosure
Authors declare no competing interests.
References
- Spizzirri FD, Rahman RC, Bibiloni N, Ruscasso JD, Amoreo OR. Childhood hemolytic uremic syndrome in Argentina: long-term follow-up and prognostic features. Pediatr Nephrol. 1997;11:156–60. doi: 10.1007/s004670050248. [DOI] [PubMed] [Google Scholar]
- Rivas M, Miliwebsky E, Chinen I, Deza N, Leotta GA. The epidemiology of hemolytic uremic syndrome in Argentina. Diagnosis of the etiologic agent, reservoirs and routes of transmission. Medicina (B Aires) 2006;66(Suppl. 3):27–32. [PubMed] [Google Scholar]
- Belardo M. Historical stages of hemolytic uremic syndrome in Argentina (1964–2009) Arch Argent Pediatr. 2012;110:416–20. doi: 10.5546/aap.2012.416. [DOI] [PubMed] [Google Scholar]
- Garg AX, Suri RS, Barrowman N, et al. Long-term renal prognosis of diarrhea-associated hemolytic uremic syndrome: a systematic review, meta-analysis, and meta-regression. JAMA. 2003;290:1360–70. doi: 10.1001/jama.290.10.1360. [DOI] [PubMed] [Google Scholar]
- Besbas N, Karpman D, Landau D, et al. A classification of hemolytic uremic syndrome and thrombotic thrombocytopenic purpura and related disorders. Kidney Int. 2006;70:423–31. doi: 10.1038/sj.ki.5001581. [DOI] [PubMed] [Google Scholar]
- Caprioli J, Noris M, Brioschi S, et al. Genetics of HUS: the impact of MCP, CFH, and IF mutations on clinical presentation, response to treatment, and outcome. Blood. 2006;108:1267–79. doi: 10.1182/blood-2005-10-007252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warwicker P, Goodship TH, Donne RLetal. Genetic studies into inherited and sporadic hemolytic uremic syndrome. Kidney Int. 1998;53:836–44. doi: 10.1111/j.1523-1755.1998.00824.x. [DOI] [PubMed] [Google Scholar]
- Noris M, Mescia F, Remuzzi G. STEC-HUS, atypical HUS and TTP are all diseases of complement activation. Nat Rev Nephrol. 2012;8:622–33. doi: 10.1038/nrneph.2012.195. [DOI] [PubMed] [Google Scholar]
- Dunkelberger JR, Song WC. Complement and its role in innate and adaptive immune responses. Cell Res. 2010;20:34–50. doi: 10.1038/cr.2009.139. [DOI] [PubMed] [Google Scholar]
- Gianantonio CA, Vitacco M, Mendilaharzu F, Gallo G. The hemolytic–uremic syndrome. Renal status of 76 patients at long-term follow-up. J Pediatr. 1968;72:757–65. doi: 10.1016/s0022-3476(68)80427-5. [DOI] [PubMed] [Google Scholar]
- Cameron JS, Vick R. Plasma-C3 in haemolytic uraemic syndrome and thrombotic thrombocytopenic purpura. Lancet. 1973;2:975. doi: 10.1016/s0140-6736(73)92645-7. [DOI] [PubMed] [Google Scholar]
- Kaplan BS, Thomson PD, MacNab GM. Serum-complement levels in haemolytic uraemic syndrome. Lancet. 1973;2:1505–6. doi: 10.1016/s0140-6736(73)92782-7. [DOI] [PubMed] [Google Scholar]
- Kim Y, Miller K, Michael AF. Breakdown of C3 and factor B in hemolytic–uremic syndrome. J Lab Clin Med. 1977;89:845–50. [PubMed] [Google Scholar]
- Monnens L, Molenaar J, Lambert PH, Proesmans W, van Munster P. The complement system in hemolytic–uremic syndrome. Clin Nephrol. 1980;4:168–71. [PubMed] [Google Scholar]
- Robson WL, Leung AK, Fick GH, McKenna AI. Hypocomplementemia and leukocytosis in diarrhea-associated hemolytic uremic syndrome. Nephron. 1992;3:296–99. doi: 10.1159/000187063. [DOI] [PubMed] [Google Scholar]
- Thurman JM, Marians R, Emlen W, et al. Alternative pathway of complement in children with diarrhea-associated hemolytic uremic syndrome. Clin J Am Soc Nephrol. 2009;4:1920–4. doi: 10.2215/CJN.02730409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morigi M, Galbusera M, Gastoldi S, et al. Alternative pathway activation of complement by Shiga toxin promotes exuberant C3a formation that triggers microvascular thrombosis. J Immunol. 2011;187:172–80. doi: 10.4049/jimmunol.1100491. [DOI] [PubMed] [Google Scholar]
- Ståhl AL, Sartz L, Karpman D. Complement activation on platelet–leukocyte complexes and microparticles in enterohemorrhagic Escherichia coli-induced hemolytic uremic syndrome. Blood. 2011;117:5503–13. doi: 10.1182/blood-2010-09-309161. [DOI] [PubMed] [Google Scholar]
- Orth D, Khan AB, Naim A, et al. Shiga toxin activates complement and binds factor H: evidence for an active role of complement in hemolytic uremic syndrome. J Immunol. 2009;182:6394–400. doi: 10.4049/jimmunol.0900151. [DOI] [PubMed] [Google Scholar]
- Lapeyraque AL, Malina M, Fremeaux-Bacchi V, et al. Eculizumab in severe Shiga-toxin-associated HUS. N Engl J Med. 2011;364:2561–3. doi: 10.1056/NEJMc1100859. [DOI] [PubMed] [Google Scholar]
- Koch FJ, Jenis EH, Valeski JE. Test for C3 nephritic factor activity by immunofixation electrophoresis. Am J Clin Pathol. 1981;76:63–67. doi: 10.1093/ajcp/76.1.63. [DOI] [PubMed] [Google Scholar]
- Moshage H. Cytokines and the hepatic acute phase response. J Pathol. 1997;181:257–66. doi: 10.1002/(SICI)1096-9896(199703)181:3<257::AID-PATH756>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- Muscari A, Bozzoli C, Puddu Getal. Association of serum C3 levels with the risk of myocardial infarction. Am J Med. 1995;98:357–64. doi: 10.1016/S0002-9343(99)80314-3. [DOI] [PubMed] [Google Scholar]
- Engstrom G, Hedblad B, Berglund G, Janzon L, Lindgarde F. Plasma levels of complement C3 is associated with development of hypertension: a longitudinal cohort study. J Hum Hypertens. 2007;21:276–82. doi: 10.1038/sj.jhh.1002129. [DOI] [PubMed] [Google Scholar]
- Ståhl AL, Svensson M, Mörgelin M, et al. Lipopolysaccharide from enterohemorrhagic Escherichia coli binds to platelets through TLR4 and CD62 and is detected on circulating platelets in patients with hemolytic uremic syndrome. Blood. 2006;108:167–76. doi: 10.1182/blood-2005-08-3219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Conde I, Crúz MA, Zhang H, López JA, Afshar-Kharghan V. Platelet activation leads to activation and propagation of the complement system. J Exp Med. 2005;201:871–9. doi: 10.1084/jem.20041497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferraris V, Acquier A, Ferraris JR, Vallejo G, Paz C, Mendez CF. Oxidative stress status during the acute phase of haemolytic uraemic syndrome. Nephrol Dial Transplant. 2011;26:858–64. doi: 10.1093/ndt/gfq511. [DOI] [PubMed] [Google Scholar]
- Collard CD, Lekowski R, Jordan JE, Agah A, Stahl GL. Complement activation following oxidative stress. Mol Immunol. 1999;36:941–8. doi: 10.1016/s0161-5890(99)00116-9. [DOI] [PubMed] [Google Scholar]
- Collard CD, Väkevä A, Morrissey MA, et al. Complement activation after oxidative stress: role of the lectin complement pathway. Am J Pathol. 2000;156:1549–56. doi: 10.1016/S0002-9440(10)65026-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collard CD, Montalto MC, Reenstra WR, Buras JA, Stahl GL. Endothelial oxidative stress activates the lectin complement pathway: role of cytokeratin 1. Am J Pathol. 2001;159:1045–54. doi: 10.1016/S0002-9440(10)61779-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hart ML, Walsh MC, Stahl GL. Initiation of complement activation following oxidative stress. In vitro and in vivo observations. Mol Immunol. 2004;41:165–71. doi: 10.1016/j.molimm.2004.03.013. [DOI] [PubMed] [Google Scholar]
- Wu Z, Lauer TW, Sick A, Hackett SF, Campochiaro PA. Oxidative stress modulates complement factor H expression in retinal pigmented epithelial cells by acetylation of FOXO3. J Biol Chem. 2007;282:22414–25. doi: 10.1074/jbc.M702321200. [DOI] [PubMed] [Google Scholar]
- Amer J, Zelig O, Fibach E. Oxidative status of red blood cells, neutrophils, and platelets in paroxysmal nocturnal hemoglobinuria. Exp Hematol. 2008;36:369–377. doi: 10.1016/j.exphem.2007.12.003. [DOI] [PubMed] [Google Scholar]