

Abstract
The vagus nerve is composed primarily of nonmyelinated sensory neurons whose cell bodies are located in the nodose ganglion (NG). The vagus has widespread projections that supply most visceral organs, including the bladder. Because of its nonspinal route, the vagus nerve itself is not directly damaged from spinal cord injury (SCI). Because most viscera, including bladder, are dually innervated by spinal and vagal sensory neurons, an impact of SCI on the sensory component of vagal circuitry may contribute to post-SCI visceral pathologies. To determine whether SCI, in male Wistar rats, might impact neurochemical characteristics of NG neurons, immunohistochemical assessments were performed for P2X3 receptor expression, isolectin B4 (IB4) binding, and substance P expression, three known injury-responsive markers in sensory neuronal subpopulations. In addition to examining the overall population of NG neurons, those innervating the urinary bladder also were assessed separately. All three of the molecular markers were represented in the NG from noninjured animals, with the majority of the neurons binding IB4. In the chronically injured rats, there was a significant increase in the number of NG neurons expressing P2X3 and a significant decrease in the number binding IB4 compared with noninjured animals, a finding that held true also for the bladder-innervating population. Overall, these results indicate that vagal afferents, including those innervating the bladder, display neurochemical plasticity post-SCI that may have implications for visceral homeostatic mechanisms and nociceptive signaling.
Keywords: vagus nerve, nodose ganglion, spinal cord injury, bladder, immunohistochemical phenotype
spinal cord injury (SCI) results in deficits to sensorimotor systems and profoundly affects the functionality of the autonomic nervous system. Basic research focusing on improving pelvic-visceral outcomes following SCI is of great clinical importance, since complications such as bladder, bowel, and sexual dysfunction affect health and quality of life for this population (4, 47, 67). Despite the direct immediate impact of injury to the spinal-derived autonomic supply of the pelvic viscera, most of the body's visceral organs also are supplied by a nonspinal source through the vagus nerve. Since the vagus nerve does not travel directly through the spinal cord, its neurocircuitry is often considered intact following SCI. Nevertheless, there is some degree of indirect involvement of both vagal afferents and efferents. For example, following SCI, subsequent neuroplastic-responsive changes have been extensively described within the dorsal vagal complex controlling gastric function (79). Gastrointestinal (GI) alterations after human upper-thoracic SCI include conditions such as dysphagia (160), esophagitis (133), peptic ulcerations (62, 138), gastroparesis, and overall dysmotility (87, 123, 125, 159). Although the mechanisms of GI dysfunction in humans after SCI are not thoroughly understood, experimental studies in rats suggest that many of the delays in gastric emptying and transit may, in part, be attributed to vagally mediated pathways (60, 61, 79, 140). In fact, subdiaphragmatic vagotomy has been shown to prevent much of the SCI-induced GI sequelae (61).
The vagus nerve, with sensory cell bodies primarily located in the nodose ganglia (NG), provides innervation to the thoraco-abdominal structures. Despite the view that the vagus nerve does not innervate viscera caudal to the transverse colon, numerous experimental studies have demonstrated that it also provides sensory innervation to the majority of the pelvic viscera (2, 28, 38, 57, 77, 81, 84, 111, 149). Even though the vagus nerve has such widespread projections, SCI does not disconnect the anatomical relationship the nerve has with the tissue it innervates. However, SCI does lead to pathological changes and dysfunction of below-level target organs, such as the bladder (45, 46, 94), and can thereby influence neuronal phenotype (122, 145, 158, 167, 176). Furthermore, various classes of primary sensory neurons, including vagal afferents, have been shown to alter their phenotype and the expression of different receptors in response to nerve injury and tissue inflammation (11, 78, 103, 107, 155, 173).
In this experiment, P2X3 receptor and substance P (SP) immunoreactivity (ir), as well as isolectin B4 (IB4) binding were examined in NG neurons. These particular markers were selected based on their presence in the NG, involvement in the spinal and vagal circuitry, responsiveness in other sensory neurons to injury and/or inflammation, and the potential physiological role these markers may play in nociceptive signaling (11, 25, 30, 40, 42, 43, 83, 97, 103, 109, 146, 154, 163, 169, 172). In addition, anatomical evidence that the vagus nerve provides sensory innervation to the bladder in male rats (77) and the presence of these cellular markers in bladder tissue (6, 19, 100) add to the importance of understanding the relationship between target-organ tissue and its innervating neurons. It is, therefore, hypothesized that, following a spinal transection injury, which removes any potential sources of spinal input rostrally and isolates vagal afferent fibers, the expression profile of known injury-responsive factors P2X3, IB4, and SP would be altered in the NG in general and also for the subset of NG neurons innervating the bladder.
MATERIALS AND METHODS
Animals.
All experimental procedures were carried out according to National Institutes of Health guidelines and protocols reviewed and approved by the Institutional Animal Care and Use Committee at the University Of Louisville School of Medicine. All adult male Wistar rats (n = 16, Harlan Sprague-Dawley, Indianapolis, IN), ∼250 g in weight, were individually housed in an animal room with a 12:12-h light and dark cycle. They had ad libitum access to water and food (Laboratory Rodent Diet). Groups were either naive (n = 8) or spinal cord injured (n = 8). Each group had a subset (n = 4 each) that received retrograde neural tracer injected into the bladder to enable identification of single NG neurons, which innervated the bladder.
SCI.
One-half of the animals (n = 8) were anesthetized with a mixture of ketamine (80 mg/kg) and xylazine (10 mg/kg), injected intraperitoneally, for spinal transection. All surgeries were performed under aseptic conditions, and the body temperature was maintained within the range of 36–37°C via a warm water recirculator (Gaymar T/Pump, Gaymar Industries, Orchard Park, NY) throughout the surgery and recovery period. Following our laboratory's previously published protocol (86), a dorsal longitudinal incision was made to expose the T7 vertebra, and a laminectomy was performed to expose the underlying T8 spinal cord. The overlying dura was reflected laterally, and the spinal cord cut using a pair of surgical microdissecting scissors. Gentle suction with an air vacuum was used to carefully elevate the cut stump to verify the completion of the lesion. Gelfoam (Pharmacia & Upjohn, Kalamazoo, MI) soaked in topical hemostat solution (Henry Schein, Melville, NY) was placed in the lesion cavity. The incision was closed using 4-0 nylon suture for the muscle layers and fascia and surgical clips for the skin. Animals were given subcutaneous injections of ketoprofen (Ketofen, 2.5 mg/kg, Fort Dodge Animal Health, Fort Dodge, IA) for analgesia twice a day for 2 days, 0.5 ml of dual penicillin (Penicillin G coupled with Procaine, PenJect, The Butler, Columbus, OH) in a single dose perioperatively as a general prophylactic and 5 mg/kg gentamicin (GentaFuse, Butler Schein, Dublin, OH) once per day for 5 days to prevent bladder infections. After surgery, each animal was housed individually. The urinary bladder was emptied by manual crede every 8 h until the micturition reflex occurred automatically, typically 6–12 days after surgery (82). Animals survived for 6 wk, followed by euthanasia and tissue removal.
Retrograde tracer injection.
At 5 wk postinjury, four spinally transected rats and four age-matched naive control rats were anesthetized with a mixture of ketamine (80 mg/kg) and xylazine (10 mg/kg). They received a ventral/caudal midline peritoneal incision to expose the urinary bladder, which was subsequently manually voided by pressure. Using an established protocol (77, 124), the fluorescent tracer FAST DiI oil (1,1′-dilinoleyl-3,3,3′,3′-tetramethylindo-carbocyanine perchlorate, 5 mg dye dissolved in 0.1 ml methanol, Molecular Probes, Eugene, OR) was injected into the bladder wall with a dye-dedicated 33-gauge needle coupled to a Hamilton microsyringe (Fisher Scientific, Pittsburgh, PA). Note that the abbreviation DiI is used throughout the text and figure legends to refer to FAST DiI oil. Dye injections were made to the circumference of the trigone, body, and dome areas [10 μl volume per animal divided into 10 injections of 1 μl each (77)].
Animal body temperature was maintained within the range of 36–37°C during surgery via a warm water recirculator (Gaymar T/Pump, Gaymar Industries, Orchard Park, NY). After each injection, the needle was removed slowly, and any dye-leakage was removed by cotton-tipped applicators. After injections were completed, the exposed viscera were hydrated as necessary with 5% dextrose lactated Ringer, the abdominal musculature was sutured closed (Ethicon 4-0 nonabsorbable surgical suture), the skin closed with Michel clips (Fine Science Tools, Foster City, CA), and a topical antibiotic (Bacitracin, Actavis Mid Atlantic, Lincolnton, NC) applied. After surgery, animals were placed on a heating pad and core temperature monitored. Postoperative medication was provided as per spinal transection surgery. All animals were monitored daily to inspect the surgical incision and identify any changes in an animal's general condition.
Perfusion and tissue collection.
All 16 animals were deeply anesthetized with a ketamine (80 mg/kg body wt)-xylazine (10 mg/kg) mixture and transcardially exsanguinated with heparinized saline, followed by 4% paraformaldehyde perfusion. Each vagus nerve was identified adjacent to the carotid artery and gently separated from surrounding tissues and traced rostrally to the NG, which was excised. The bladder was dissected free from the prostate and extracted at the most distal aspect of the neck. Before weighing the bladder, any residual intravesicular urine was allowed to flow out. Superior cervical ganglia were identified on both sides at the bifurcation of the common carotid artery and removed to be used as control tissue. For the transected group of rats, following a dorsal spinal incision, removal of the spinal cord tissue 1 cm above and below the transection site was performed. All tissues were placed in individually labeled tubes of 4% paraformaldehyde for at least 48 h, followed by immersion in a cryoprotectant solution of 30% sucrose/phosphate buffer solution with 1% sodium azide for at least 24 h. Following removal from the cryoprotectant solution, NGs were embedded in OCT compound (Baxter Scientific) and 12- to 14-μm sections were cut on a cryostat (Leica CM 1850). During retrieval of the bladder, the abdominal cavity and surrounding viscera were inspected for tracer leakage.
Histology of the lesion epicenter.
The lesion cavity, coated in embedding media, was cut into 18-μm sagittal sections using a Leica CM 1850 cryostat and mounted onto gelatin-coated histological slides (Azer Scientific, Morgantown, PA). The slides were then stained with both Luxol fast blue and cresyl violet (Kluver-Barrera method) to observe myelin and Nissl substance, respectively. Spot Advanced software (Diagnostic Instruments, Sterling Heights, MI) and the Nikon E400 microscope were used to image the lesion cavity and verify the completeness of the spinal transection (86). The percentage of spared white matter from the transection lesion was calculated using Nikon Elements software. The boundary between spared tissue at the ventral portion of the cord and the lesion cavity was identified. The first anatomical region of interest (ROI) outlined was the portion of spared tissue at the ventral aspect of the spinal cord, and the second ROI outlined was the entire lesion cavity, encompassing both spared and nonspared tissue and extending from both the rostral and caudal cord stumps. Area was determined for each ROI, and the percentage of spared tissue was calculated by dividing these areas (95).
Hindlimb assessment for lesion completeness.
The Basso-Beattie-Bresnahan (BBB) scale (14), an open-field locomotor assessment, was used to evaluate hindlimb function as an assessment of postinjury spinal cord function. Each animal was placed in an open field and tested for 4 min by the same two scorers, who were presented with injured and noninjured animals in random order. The 21-point BBB scale was used to assess hindlimb coordination and rated parameters such as individual joint movements (0–7), weight support (8–13), and paw placement (14–21). Intact animals should demonstrate a locomotor score of 21. Animals that receive a complete T8 transection have been shown to exhibit BBB scores of 3 (extensive movement of 2 joints) on average (13, 130). To prevent any functional connections across the lesion site from potential spared tissue, gelfoam was placed between the two cut stumps. It has been demonstrated that as little as 4–5% sparing (primarily in the ventrolateral funiculi) was sufficient for attaining a BBB score of 7 following “complete” spinal transection (no gelfoam used across lesion) (53).
Immunofluorescence histochemistry.
Sections were thaw-mounted onto slides and allowed to air-dry. They were then encircled with hydrophobic resin (PAP Pen, Research Products). Slide-mounted sections were incubated at room temperature for 2 h in a solution of 2% Triton X-100 in phosphate-buffered saline (PBS). This pretreatment step improves the quality of P2X3-ir (116). The slides were rinsed in distilled water and then incubated for 30 min in a solution of 10% normal donkey serum (Jackson Immuno Research, West Grove, PA) in PBS with 0.3% Triton X-100 (MP Biomedicals, Solon, OH) to block nonspecific antibody binding. The immunohistochemical reagents and the labeling procedures are summarized in Table 1. Incubations in primary antisera were performed overnight (14–18 h) at 4°C. All steps were followed by multiple rinses with PBS. All fluorescent secondary antisera were diluted 1:100, and incubations were 2 h at room temperature. The tyramide signal-amplification reagent kit (Sigma-Aldrich, St. Louis, MO) (20–22) was used at 1:100, and incubations were for 4–5 min (124). Once all steps were completed, the slides were coverslipped with a glycerol-based photobleach-protective medium (Fluoromount-G, Southern Biotech).
Table 1.
Immunohistochemical reagents
| Target | Primary Detection | Dilution/Vendor/Catalog No. | Secondary Detection | Dilution/Vendor/Catalog No. |
|---|---|---|---|---|
| P2X3 | GP-anti-P2X3 | 1:1,000/Neuromics/GP10108 | Dky anti-GP-AF488 | 1:100/Jackson/706-545-148 |
| Biotinylated lectin from Bandeiraea simplicifolia isolectin B4 | HRP-SA | 1:500/Sigma-Aldrich/L2140 | Tyramide-AF350 | 1:100/Molecular Probes/T20937TSA kit no. 27 |
| SP | RBT-anti-SP | 1:1,000/Abcam/ab67006 | Dky anti-RBT-Cy3 | 1:100/Jackson/711-165-152 |
| NeuN | MS-anti-NeuN | 1:1,000/Chemicon/MAB377 | Dky anti-MS-Cy5 | 1:100/Jackson/715-175-151 |
SP, Substance P; GP, guinea pig; HRP, horseradish peroxidase; SA, strepavidin; RBT, rabbit; MS, mouse; Dky, donkey; AF, Alexa Fluor; TSA, tyramide signal amplification.
Cell quantification.
To view labeled sections, imaging was performed using the Nikon Eclipse TiE inverted microscope with NIS Elements software. Initially, images were captured using a ×10 lens (APO DIC N1 ×10/0.45 numeric aperture, Nikon) with consistent exposure times and computationally stitched together to visualize whole ganglion sections. Imaging of individual fluorophores was achieved with a mercury-arc light source and the following filter sets: for Cy3 (543/22 nm excitation, 593/40 nm emission, 562 nm dichroic, Semrock); for Alexa Fluor 488 (470/40 nm excitation, 525/50 nm emission, 495 nm dichroic, Nikon); for Alexa Fluor 350 (350/50 nm excitation, 460/50 nm emission, 400 nm dichroic, Nikon); for Cy5 (615/70 nm excitation, 700/74 nm emission, 660 nm dichroic, Nikon).
To obtain unbiased percentages of P2X3, SP, and IB4 positive neurons in the NG, the physical dissector method was applied (37, 113). Across the entire ganglion, assembled by automated stitching, counts of all singly labeled, multilabeled, as well as nonlabeled/other (collectively comprising total neuronal counts) NG neurons were made by a scorer blinded to treatment groups. Starting with a random section, neurons with a clearly visible nucleus and definable soma were counted only if they were not present in an adjacent “look up” serial section. As an added measure to avoid double counting single neurons, the counting sections were at least 60 μm apart (every 5th section). To differentiate background from foreground pixels, threshold values also were obtained based off the image histogram for each marker and held constant for each image quantified. Note that nonlabeled/other cells (NeuN+ only) were quantified and represented the total neuron population (59, 75, 105). Images of tissue without application of the primary antibody were taken and utilized as baseline controls (images not shown). With the use of Nikon Elements software, cell diameter between groups also was determined by estimating the average of two cross-sectional diameters (longest and shortest axis).
Positive neuron counts were expressed as a percentage of the total number of neurons (NeuN+ only) from within the entire stitched ganglion, as well as a percentage of all labeled neurons. An Olympus 3 Laser scanning confocal microscope with Fluoview 500 software (Mellville, NY) and a Nikon A1R MP+ confocal microscope with Elements software were used to collect high-resolution, serial optical sections of NG neurons.
Statistics.
Analyses were performed using SPSS version 19–20 (IBM, North Castle, NY). Levene's statistic was applied for homogeneity of variances, and data are expressed as means ± SD. A one-way ANOVA with Tukey's honestly significant difference post hoc t-tests was performed for the assessment of all histochemical markers and NG bladder labeling within groups. For the analysis of the histochemical markers between groups, data were normalized as percentages of total NG cells and analyzed via a two-way ANOVA with Tukey's honestly significant difference post hoc t-tests. For all other analyses, two-tailed Student's t-tests were performed assuming equal variance. Statistical significance was defined as P ≤ 0.05.
RESULTS
Immunohistochemical signature of NG neurons.
IB4 binding in NG neurons was localized to the plasma and axonal membranes as well as the Golgi complex. Immunoreactivity for P2X3 and SP was present in the cytoplasm, and P2X3-ir also could be found in the plasma and axonal membranes. All of the patterns of staining for these markers were consistent with previous studies in both NG and dorsal root ganglion (DRG) (8, 10, 83, 116, 151). When examining the total percentage of labeled NG neurons, all of the markers were well represented (Fig. 1A), with the majority of NG neurons binding IB4 (IB4, 65.5 ± 6.2% vs. SP, 31.1 ± 10.3%; IB4 vs. P2X3, 51.2 ± 7.9%). Overall, there were eight different histochemical signatures represented in the NG (Fig. 1B). The most prevalent combinations were neurons that were IB4+ only followed by the P2X3+ only and IB4+/P2X3+/SP− combinations (IB4+ only vs. P2X3+ only, 33.3 ± 7.1 vs. 15.0 ± 3.3%, P < 0.001; IB4+ only vs. IB4+/P2X3+ only, 33.3 ± 7.1 vs. 15.0 ± 5.0%, P < 0.001). When considering specific colocalization patterns in the NG, about one-third of all IB4 neurons contain P2X3 (30.9 ± 9.2%) and about one-half of all P2X3 neurons contain IB4 (49.1 ± 8.6%). A typical example of the quadruple immunohistochemical staining in the NG is provided in Fig. 2.
Fig. 1.

Immunohistochemical representation of P2X3, isolectin B4 (IB4), and substance P (SP) in nodose ganglion (NG) neurons. A: following staining of the immunohistochemical markers P2X3, SP, and IB4, the bar graph demonstrates that all three were well represented in the NG, with the majority being IB4+ (IB4 vs. P2X3, #P < 0.05; IB4 vs. SP, **P < 0.001; P2X3 vs. SP, *P < 0.01). Values are means ± SD; n = 3 rats and 6 ganglia (one-way ANOVA with Tukey post hoc t-tests). B: pie graph depicting all possible combinations of the molecular targets in the NG. Note that neurons that were IB4+ only were the most prevalent, and most NG neurons were labeled with at least one of the three cellular targets examined.
Fig. 2.

Quadruple immunohistochemical staining in the NG. A: NeuN. B: IB4. C: P2X3. D: SP. E: merged. A confocal image displays the typical staining within the NG. NeuN was used to label all neurons. Different histochemical combinations include neurons that were IB4+, P2X3+, but SP− (white arrows), and neurons that were IB4−, P2X3+, and SP− (yellow arrows). Scale bar indicates 25 μm (×20 objective).
Effect of SCI on NG neurons expressing P2X3, SP, and binding IB4.
Following a chronic transection injury at T8, BBB locomotor assessments at the 6-wk time point for all transected animals revealed an average score of 0.9 (±0.9). Only one animal had a BBB score greater than zero (score of 4.5:6 on the left hindlimb and 3 on the right hindlimb). Post hoc histological assessments of the lesion site revealed that this same animal in the spinal cord-transected group had a small percentage of area spared within the lesion site (16.1%, located at the most ventral extent of the epicenter). The data from this incomplete transection rat are provided, but as a separate group with an n of 1. Although statistical analyses could not be performed with n = 1, the immunohistochemical expression pattern in this SCI rat appeared to more closely resemble that of the naive group (P2X3+ only, 11.5 ± 5.2%; IB4+ only, 44.0 ± 2.2%). An example indicating a typical complete lesion is provided in Fig. 3. Note that it was also found that the average cell diameter for NG neurons was 29.6 ± 5.7 μm. This morphological feature was not affected by spinal transection injury (28.5 ± 6.04 μm).
Fig. 3.
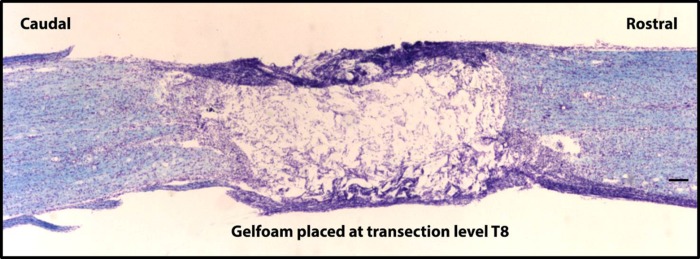
Spinal transection injury at T8. An 18-μm-thick sagittal section stained with the Kluver-Barrera method illustrates a complete spinal transection at T8. Gelfoam is placed in the lesion cavity to prevent contact from between rostral and caudal spinal cord tissue. The scale bar indicates 500 μm (×4 objective).
In rats with a complete transection (n = 3; 6 ganglia), there was a significant increase in the percentage of total NG neurons expressing P2X3 (P < 0.001), as well as a significant decrease in the percentage of total NG neurons binding IB4 (P < 0.05) relative to noninjured controls (Fig. 4). There were no significant differences in SP or NeuN expression between groups. P2X3 expression and IB4 binding in the NG are demonstrated in Fig. 5. Within the spinal cord-transected group, the percentage of neurons expressing P2X3 and the percentage of neurons binding IB4 were each significantly greater than the percentage of neurons expressing SP, which was unchanged from the noninjured group of animals (P2X3, 27.3 ± 4.8% vs. SP, 6.5 ± 2.8%, P < 0.01; IB4, 23.7 ± 6.5% vs. SP, 6.5 ± 2.8%, P < 0.01; SP, 6.5 ± 2.8% vs. SP-noninjured, 5.8 ± 3.8%). From the total population of NG neurons, SCI did not significantly impact the subset of NG neurons that coexpressed IB4 and P2X3 (naive, 15.0 ± 5.0 vs. transected, 13.5 ± 5.7, P > 0.05), nor did it impact the subset of all IB4 neurons that contained P2X3 (naive, 30.9 ± 9.2% vs. transected, 35.9 ± 11.2%, P > 0.05). However, following SCI, the subset of all P2X3 neurons that contained IB4 was significantly lower compared with noninjured animals (naive, 49.1 ± 8.6% vs. transected, 32.2 ± 10.5%, P < 0.01).
Fig. 4.

The effect of spinal cord injury (SCI) on number of NG neurons expressing the individual markers. Following SCI, there was an increase in P2X3-immunoreactivity (ir) in the transected group relative to intact/normal (SCI, 27.3 ± 4.8% vs. noninjured, 15.0 ± 3.3%, *P < 0.001) and a decrease in IB4 binding in the transected group relative to intact/normal (SCI, 23.7 ± 6.5% vs. noninjured, 33.3 ± 7.1%, #P < 0.05). No changes were apparent for SP. The “Other” category represents neurons that were NeuN+, but did not express or bind any of the markers. Values are means ± SD; n = 6 rats and 12 ganglia.
Fig. 5.

The effect of SCI on P2X3 and IB4 in the NG. An example displaying P2X3-ir (A) and IB4 binding (C) in the NG and following chronic spinal cord transection injury at T8 (B and D, respectively) is shown. Note the presence of increased P2X3-ir and decreased IB4 binding post-SCI. Images of sections from both SCI and noninjured animals were stained and captured with the same protocols and at the same time (×10 objective).
With respect to the total number of labeled neurons within their individual populations (i.e., all P2X3+ neurons and all IB4+ neurons), the distribution of the P2X3+/IB4−/SP− only subset represents 50.7% of all P2X3+ neurons, while the IB4+/P2X3−/SP− only subset represents 42.6% of all IB4+ neurons. Overall, these populations of neurons comprised about one-half of the total population of NG neurons examined.
Immunohistochemical profile of bladder noninjured and injured NG neurons.
The retrograde tracer DiI was injected into the bladder wall to determine whether the subsets of NG neurons affected by spinal cord transection include those that supply the bladder. Initially, we found that the percentage of NG neurons traced from the bladder in both groups in this study is similar to that in our previous study of spinally intact animals [22.2 ± 3.6 vs. 21.4 ± 4.0% (77)]. Assessment of the superior cervical ganglion, which is located adjacent to the NG, did not reveal evidence of DiI tracer, indicating the tracer did not spread nonspecifically (Fig. 6).
Fig. 6.

Superior cervical ganglion. There was no evidence of 1,1′-dilinoleyl-3,3,3′,3′-tetramethylindo-carbocyanine perchlorate (DiI) punctate labeling present in the superior cervical ganglion. The scale bar represents 20 μm (×20 objective).
With respect to the total population of P2X3+ NG neurons after chronic SCI (50.7 ± 8.2%), bladder-innervating neurons (DiI+/P2X3+/IB4−) represented 32.8 ± 1.1%, while with respect to the total population of neurons that were IB4+ after injury (42.6 ± 5.1%), bladder-innervating neurons (DiI+/P2X3−/IB4+) represented 21.5 ± 7.4%. Overall, in these two distinct subsets of NG neurons, more than one-half of the neurons are traced from the bladder (Fig. 7). Images of the DiI+/P2X3+ and DiI+/IB4+ subsets following transection are demonstrated in Fig. 8. Note that, in this study, the proportion of NG neurons traced from the bladder in spinally intact rats (23.7 ± 3.6%) did not differ significantly from the proportion traced from the bladder after chronic spinal transection injury (20.2 ± 3.0%). Transection injury did, however, result in a significant increase in bladder size (wet weight) compared with noninjured controls (0.267 ± 0.118 g SCI vs. 0.149 ± 0.35 g naive; P < 0.05), which can impact overall total bladder capacity and voiding efficiency, potentially leading to an alteration in bladder function.
Fig. 7.

Effect of SCI on bladder-traced NG neurons. The graph demonstrates that, out of the total percentage of either P2X3 or IB4 subsets after injury, more than one-half of the neurons were traced from bladder. Bladder innervating neurons in the P2X3+ subset represent 32.8 ± 1.1%, while, in the IB4+ subset, they represent 42.6 ± 5.1%. n = 3 rats and 6 ganglia.
Fig. 8.

P2X3-ir and IB4 binding in bladder-traced NG neurons after transection. A confocal image illustrates a DiI+ neuron in A that is also immunoreactive for P2X3 in B (white arrows). C: demonstration of the overlay. An image from the inverted Nikon microscope illustrates a DiI+ neuron in D that also binds IB4 in E (white arrowhead). F: demonstration of the overlay. One example of each is displayed. In both images, the scale bar indicates 25 μm (×20 objective).
DISCUSSION
Visceral organs, including those in the pelvic region, have a dual sensory innervation from spinal and nonspinal (i.e., the vagus nerve) sources (28, 38, 57, 77, 81, 84, 111). This study, using immunohistochemical techniques, examined the impact of SCI on the vagal component of that sensory innervation by assessing changes in the presence of P2X3, IB4, and SP in NG neurons. After spinal transection at T8, potential plasticity was evaluated in subsets of NG neurons that contain projections that bypass the spinal cord from visceral organs, including those projections that specifically supply the bladder. A major traumatic event to the nervous system, such as SCI, leads to dysfunction in multiple organ systems and ultimately influences the neurons that innervate these tissues. The findings of the present study indicate that vagal sensory cell bodies displayed an increase in P2X3 expression and a decrease in IB4 binding, which also held true for many neurons innervating the bladder. These results suggest that NG neurons, including the bladder subset, are sensitive to a spinal injury and are capable of responding by modifying their phenotype.
Immunohistochemical phenotype of NG neurons.
Overall, from the cellular markers examined in this study, the majority of NG neurons were IB4+. IB4 has been shown to label primarily a subpopulation of nonpeptidergic spinal sensory afferents that are thought to be functionally distinct from peptidergic neurons or neurons that are IB4 negative (135). Even though IB4+ neurons, in general, are widely expressed within the NG (83, 98, 129, 169, 172), the meaning of IB4 binding in vagal afferents and whether or not these neurons share common characteristics based on their IB4 binding is still unclear. Vagal afferents are largely known for their involvement in conveying information about the physiological state of the viscera to the brain as part of homeostatic regulation (32, 66, 85). In the GI tract, vagal afferent fibers are responsive to stretch and tension, as well as to locally released hormones following the ingestion of food (16, 18, 119, 156). Although they are typically not responders to visceral stimuli within the noxious range (112), previous data from our laboratory and that of others suggest otherwise. For example, while spinal afferents may be responsible for relaying mechanical nociceptive information, vagal afferents may play more of a role in conveying chemical nociceptive stimuli, thus contributing to disease-related conditions stemming from visceral hyperalgesia (44, 80, 86, 136). Therefore, besides the known role of vagal afferents in relaying homeostatic information to the brain, the population of vagal afferents that also are IB4+ may be involved in visceral nociceptive processing [based on the putative role of the majority of DRG IB4 binding neurons (117, 151)].
In the present study, it was shown that many vagal visceral afferents innervating the bladder also were IB4+. In other NG visceral afferents and in line with the results here, labeled vagal afferents from the stomach and duodenum demonstrated a substantial percentage of IB4 binding (172). Furthermore, numerous studies identify a low percentage of calcitonin gene related peptide (CGRP)-ir or peptide-containing NG neurons projecting to thoracoabdominal viscera (63, 175). Despite the fact that a large proportion of visceral NG neurons appear to be IB4+, the functional significance of these specific subsets also has yet to be determined, as evidence of particular markers for the coding of vagal afferent subtypes are limited (17). One exception may be calretinin (calcium binding protein), which appears to be expressed specifically by cervical esophageal vagal afferents (49).
Similar to other studies reporting P2X3 receptor expression in vagal sensory cell bodies using immunohistochemical techniques, P2X3 receptors were highly prevalent and distributed throughout the NG in the present study (11, 154). These findings suggest that large populations of vagal afferents are sensitive to ATP, and thus vagal pathways may be activated through purinergic signaling mechanisms. When considering individual subsets of neurons based on the cellular markers examined in this study, P2X3 receptor expression was present in about one-half of the IB4+ NG neurons, a coexpression subset previously reported by our group in the NG (83) and others in the DRG (25, 150, 152, 153, 164) and trigeminal ganglion (TG) (131). However, while the percentage of overlap of IB4+ neurons expressing P2X3 is relatively higher in the DRG [67.5% (25)], this coexpression pattern was similar in the TG [32% (131)] to our findings in the NG (31%). Furthermore, the percentages for all P2X3+ neurons containing IB4 are both greater in the DRG (98%) and TG (64%) compared with our findings in the NG (49%). The lower percentage of overlap between IB4 and P2X3 overall in the NG compared with other sensory ganglia may be attributed to differences in embryological origin (NG, epibranchial placode derived vs. DRG, neural crest derived vs. TG, both placode and neural crest), which appears to influence the neurochemical phenotype of visceral afferents. For instance, DRG afferents innervating different viscera, such as the stomach (63, 64, 128, 172), duodenum (172), and pancreas (54, 128), are primarily peptidergic, expressing transient receptor potential vanilloid 1, CGRP, or SP, whereas NG afferents innervating those same tissues display limited expression of these peptidergic markers. In addition, DRG afferents innervating bladder (168), colon (35), and gastroduodenal (172) tissue exhibit low IB4 binding, which is in contrast to the substantial degree of bladder-innervating IB4+ neurons reported here in the NG and the significantly greater percentage of IB4+ NG afferents innervating the stomach and duodenum reported previously (172).
In regards to the overall population of NG neurons, the percentage of SP+ only neurons we found was similar to an earlier report, around 30% (161). While it is noted that SP+ neurons are abundant in the NG (175), their distribution has been reported to be located near the rostral pole of the ganglion (72, 161, 175). We and others have previously found a homogenous distribution of visceral labeling throughout the NG (1, 77, 127, 172). Although we did not assess the existence of an organotypic distribution of labeling for the histological markers of interest within the NG in this study, the presence of many SP-ir neurons in the rostral region may be anticipated due to the fact that it is anatomically close to the jugular ganglion, containing neuropeptide-rich neurons that primarily project to rostrally located viscera, such as the esophagus and lungs (120, 143, 157), and is embryologically distinct (neural crest-derived) from the NG (placode-derived) (9). The proximity of neurons with similar neurochemical phenotypes may be important for performing like functions and even for sensory afferent integration (26). Furthermore, given the embryological distinction between the vagal ganglia, the phenotype of vagal afferents also may be influenced by whether or not the cell body is located in either the nodose or jugular ganglion (170).
Effect of SCI on P2X3-ir in NG neurons.
Following chronic SCI, two significant changes were observed in subsets of NG neurons. The first finding was a significant increase in the number of neurons expressing P2X3-ir in the spinal-transected group relative to noninjured controls. In the somatosensory system, alterations in P2X3 expression following nerve injury have been mixed. Both downregulation (25) and upregulation (50, 109) of the receptor have been documented in various peripheral nerve injury models, such as axotomy, ligation, and chronic constriction. In both studies, where there were increases in P2X3 expression, the injury model used resulted in some neurons that would be potentially “uninjured”. To assess these differences, activating transcription factor 3 (ATF3), a marker of peripheral nerve injury and absent from intact neurons (141), identified decreased P2X3 (mRNA) in ATF3-ir neurons, whereas the increased expression was evident in the intact subset of neurons (142). The significant increase in NG P2X3-ir found in the present study was consistent with the ATF3-ir findings (142), since the vagal afferents are likely not directly injured, given they by-pass the SCI, although this has not yet been directly tested.
Even though contact between the vagus nerve and its peripheral targets has not been severed, there is an overall effect of injury on the vagal afferent neurocircuitry (81, 86), and our discovery of the increased P2X3 expression seen in the NG here may help to improve our understanding of the indirect effect on the vagal system after injury. For instance, the increased P2X3-ir following SCI may be attributed to an inflammatory reaction of the system due to the nature of the injury itself. Acutely, SCI triggers an inflammatory response characterized by various resident [i.e., central nervous system (CNS) origin] (55, 90, 121) and blood-derived (134) cellular events, such as the synthesis of cytokines, chemokines, and the infiltration of leukocytes, neutrophils, and monocytes, which, over time, systemically may affect tissues outside the CNS, leading to organ dysfunction. Released inflammatory cells from the bloodstream can impact the functionality of different viscera due to the intimate relationship these organs have with the vascular system (12, 31, 65). In addition, both acute and chronic SCI induce significant changes in organs with spinal innervation from segments below the lesion level. Organs such as the bladder experience substantial stress and histopathology, which can lead to alterations in the integrity of the lining of the bladder (5), making the bladder more susceptible to chronic inflammation (76).
Structural changes after SCI also include bladder (detrusor muscle) hypertrophy, which triggers a release of neurotrophic factors, such as nerve growth factor (NGF), from the urothelial lining (56, 126, 147, 148). Increases in NGF following SCI (147, 158, 165) or inflammation (110, 132), as well as other excitatory neurotransmitters, such as ATP (137), play a major role in neuroepithelial interactions. For example, in a migraine headache model, retrograde transport of NGF from the periphery to the TG or exposure of trigeminal afferents to NGF led to an upregulation of P2X3 receptor protein in the cell bodies (41, 58). Given that the vagus nerve provides a substantial degree of innervation to the bladder (77), the fact that we found many colabeled DiI+/P2X3+ NG neurons after injury in this study, the presence of the high-affinity receptor for NGF (TrkA) (70, 96, 174) and low affinity (p75) receptor (144, 175), and that vagal afferents have the capability to transport NGF (70), the phenotypic changes with respect to P2X3-ir in the bladder-innervating NG neurons have the potential to be mediated through the actions of NGF. Importantly, in a manner distinct from the actions of NGF (41), CGRP-mediated insertion of P2X3 into the cell-surface membrane is an alternative mechanism, which has been demonstrated in sensitized TG neurons (52). However, since the majority of CGRP expressing neurons appear to reside in other cranial ganglia (petrosal, trigeminal, glossopharyngeal, and jugular) compared with the NG (68, 69, 71, 72), this molecular mechanism may indirectly affect NG neurons, perhaps acting at a distance through en passant synaptic contact (91).
The P2X3 receptor, predominantly expressed on sensory afferents (33, 97), including vagal fibers (88), also can be separately retrogradely transported from the periphery to the cell body via endosomes (34). This retrograde transport is thought to be important for maintaining neuronal activity and cell excitability through activation of transcription factors (34). In disease states, such as SCI, the extracellular milieu of ATP may be relatively high compared with healthy states, where excesses are rapidly hydrolyzed (29, 89, 108). Large amounts of ATP [likely released from damaged tissue (39)], can signal through P2X3 receptors and may show that P2X3 has a more extensive role in the NG besides normal visceral afferent transduction, perhaps contributing to nociceptive signaling following injury or tissue inflammation.
Effect of SCI on IB4 binding in NG neurons.
The second change following chronic transection injury was a decrease in NG IB4 binding relative to controls. This finding is similar to that of others in cases where decreases in the total number of IB4 binding DRG neurons on the contralateral side (uninjured side) also have been demonstrated following L5 spinal nerve transection (99). Importantly, the numbers remain reduced at the chronic time point (5 wk postinjury), suggesting the effect was not transient. Since many neurons in general in the NG were found to bind IB4, the significant post-SCI decrease in the coexpression subset of P2X3 neurons containing IB4 may be attributed to the overall decrease in this relatively large population of IB4 neurons.
An explanation for the decrease in IB4 binding may be attributed to a stress response to the system following transection. Since glial cell-line derived neurotrophic factor supports and aids in the regulation of IB4 neurons postnatally (104), perhaps some disruption to its availability or receptor complex, as well as alteration to the IB4 binding glycoconjugate, could explain the observed decrease (15, 118). However, in response to peripheral nerve injury, spared IB4 neurons also demonstrate the capability to sprout, forming perineuronal nets with both satellite and adjacent cells within the ganglion (99). It has been suggested that a mechanism behind this sprouting in response to nerve injury may involve inflammatory environmental changes that create a chemotactic gradient, attracting various chemokines (23). This communication between injured and noninjured “neighbors” within the ganglion may serve as a basis for cross-excitation and could eventually induce hyperalgesia or allodynia (3, 24). Even though the injury model used in this study does not directly injure vagal neurons, they could be considered “spared” neurons that also demonstrate a phenotypic switch in response to CNS damage and have the potential to drive visceral nociceptive signaling.
Effect of SCI on SP expression in NG neurons.
SP is one of the main neuropeptides released from a proportion of primary afferent terminal endings that express SP, in response to irritation or inflammation (7, 27, 48), and is present in NG neurons (175). No significant differences in SP-ir were present in this study between transected and noninjured groups. A previous report assessing changes in NG neurotransmitters found that SP-ir was unaffected by vagal axotomy (74). The lack of changes in the NG with respect to SP-ir following injury does not preclude any particular alterations at terminal endings, either peripherally in target organs or centrally (solitary nucleus). For instance, there is a high concentration of SP afferent terminals, primarily of vagal origin, present in the solitary nucleus (73, 102, 175). Alternatively, there may be molecular pathway alterations involved in the release of SP and translation at the cell body (114, 139). An acute SCI or direct tissue inflammation model (such as acetic acid instillation into the bladder) may provide more insight to vagal SP expression in the rat (10, 114).
Alterations to bladder NG neurons following SCI.
SCI did not result in differences compared with noninjured controls in the number of NG neurons labeled from bladder, confirming these vagal afferents remain intact after cord transection. Although vagal afferents exhibit a high degree of neurochemical and electrophysiological plasticity in response to trauma and inflammation (26, 106, 115, 171), it is likely that the observed neurochemical changes in this study are a result of interactions with the target organs that these vagal fibers innervate rather than direct neural damage. It should be noted, however, that other extrinsic sources of ATP can reach P2X3 receptors through release from sympathetic neurons, tumor cells, or from vascular endothelial cells associated with ischemia (30).
Plasticity related changes in bladder vagal afferents fall in line with evidence from the spinal system after SCI. Spinal sensory neurons innervating the bladder exhibit both morphological and physiological changes after SCI (93, 166). Given the important transduction role of P2X3 receptors in spinal bladder afferents (36) and the fact that many vagal neurons traced from the bladder expressed P2X3 suggest that the vagus nerve may participate in the sensory portion of micturition function. Our collective recent data indicating extensive vagal afferent innervation of mammalian urinary bladder (77) and SCI-induced changes in a transduction channel like P2X3 may have important clinical applications. These findings could contribute to whatever plasticity underlies reports of altered sensations stemming from the below-level viscera, such as sensations of bladder filling or fullness in clinically complete SCI patients above T10 (51, 92, 162).
A large proportion of the IB4 neurons were traced from the bladder, which is complementary to an earlier study showing that IB4 binds different types of visceral afferents in the NG (172). Although the distal urethra was not examined in the present study, a large proportion of spinal neurons that innervate this region of the lower urinary tract include IB4+ afferents (168).
Perspectives and Significance
Because of the chronic extent of multisystem functional impairment and disability, SCI presents a significant economic burden for the patient, family unit, and society overall with high direct and indirect costs estimated in the billions (101). Apart from paralysis, some of the major complications of SCI affecting quality of life, include deficits to urological function (4, 47, 67). The vagus nerve, with the majority of its cell bodies located in the NG, is an extraspinal pathway through which information from regions below the level of a spinal lesion can directly travel to the brain stem, bypassing the spinal cord entirely. Work from our laboratory previously identified an anatomical connection to the male rat urinary bladder through the vagus nerve (77). The present study examined the immunohistochemical phenotype of vagal sensory neurons overall, as well as in those that innervate the bladder. The results from this study demonstrate that vagal afferents are responsive to spinal injury, and further assessments of their functional nature may provide insight on how to take advantage of this route that by-passes the spinal cord to improve therapeutic interventions for SCI patients.
Conclusion.
The present study demonstrated neurochemical changes in the NG, a site remote from the injured spinal cord. Through target-organ neural interactions, vagal afferent fibers are influenced by their connections to the viscera. Therefore, overall changes in these organs following SCI could impact the neurochemical properties of vagal afferents innervating them. The increased expression of P2X3 and the decreased binding of IB4 in the sensory cell bodies of vagal afferents post-SCI indicates an indirect effect of injury on the vagal neurocircuitry. A majority of neurons in the P2X3 subset after spinal transection were DiI+, indicating many NG bladder afferents have the potential to respond to alterations in ATP, perhaps even playing a role in generating specific sensations associated with the bladder, such as fullness. In addition, the considerable proportion of bladder IB4+ NG neurons demonstrates that vagal afferents may participate in visceral nociceptive processing. Whether or not SCI induces changes in NG neuron function will require an examination of their electrophysiological properties in bladder populations.
GRANTS
This study was supported by National Institute of Neurological Disorders and Stroke Grant F31-NS-077750 (A. N. Herrity) and Intramural Research Incentive Grant from the University of Louisville (C. H. Hubscher). This project also utilized KSCIRC Neuroscience Core facilities that are supported by National Institute of General Medical Sciences/National Center for Research Resources P30 Grant 8P30-GM-103507.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: A.N.H., J.C.P., and C.H.H. conception and design of research; A.N.H. and K.K.R. performed experiments; A.N.H. and C.H.H. analyzed data; A.N.H., J.C.P., D.P.S., K.K.R., and C.H.H. interpreted results of experiments; A.N.H. and D.P.S. prepared figures; A.N.H. drafted manuscript; A.N.H., J.C.P., D.P.S., K.K.R., and C.H.H. edited and revised manuscript; A.N.H., J.C.P., D.P.S., K.K.R., and C.H.H. approved final version of manuscript.
ACKNOWLEDGMENTS
The authors thank Jim Armstrong, Jason Fell, Patricia Ward, Ann-Claude Rakotoniaina, Arkadiusz Slusarczyk, Jason Beare, Shelia Arnold, and Amanda Pocratsky for technical assistance. The authors also thank Drs. Scott Whittemore, Martha Bickford, and Theo Hagg for antibody donations, and Darlene Burke for statistical assistance.
REFERENCES
- 1.Altschuler SM, Bao XM, Bieger D, Hopkins DA, Miselis RR. Viscerotopic representation of the upper alimentary tract in the rat: sensory ganglia and nuclei of the solitary and spinal trigeminal tracts. J Comp Neurol 283: 248–268, 1989. [DOI] [PubMed] [Google Scholar]
- 2.Altschuler SM, Escardo J, Lynn RB, Miselis RR. The central organization of the vagus nerve innervating the colon of the rat. Gastroenterology 104: 502–509, 1993. [DOI] [PubMed] [Google Scholar]
- 3.Amir R, Devor M. Functional cross-excitation between afferent A- and C-neurons in dorsal root ganglia. Neuroscience 95: 189–195, 2000. [DOI] [PubMed] [Google Scholar]
- 4.Anderson KD. Targeting recovery: priorities of the spinal cord-injured population. J Neurotrauma 21: 1371–1383, 2004. [DOI] [PubMed] [Google Scholar]
- 5.Apodaca G, Kiss S, Ruiz W, Meyers S, Zeidel M, Birder L. Disruption of bladder epithelium barrier function after spinal cord injury. Am J Physiol Renal Physiol 284: F966–F976, 2003. [DOI] [PubMed] [Google Scholar]
- 6.Arms L, Vizzard MA. Neuropeptides in lower urinary tract function. Handb Exp Pharmacol 202: 395–423, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Avelino A, Cruz C, Nagy I, Cruz F. Vanilloid receptor 1 expression in the rat urinary tract. Neuroscience 109: 787–798, 2002. [DOI] [PubMed] [Google Scholar]
- 8.Averill S, McMahon SB, Clary DO, Reichardt LF, Priestley JV. Immunocytochemical localization of trkA receptors in chemically identified subgroups of adult rat sensory neurons. Eur J Neurosci 7: 1484–1494, 1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Baker CV, Bronner-Fraser M. Vertebrate cranial placodes. I. Embryonic induction. Dev Biol 232: 1–61, 2001. [DOI] [PubMed] [Google Scholar]
- 10.Banerjee B, Medda BK, Lazarova Z, Bansal N, Shaker R, Sengupta JN. Effect of reflux-induced inflammation on transient receptor potential vanilloid one (TRPV1) expression in primary sensory neurons innervating the oesophagus of rats. Neurogastroenterol Motil 19: 681–691, 2007. [DOI] [PubMed] [Google Scholar]
- 11.Banerjee B, Medda BK, Schmidt J, Zheng Y, Zhang Z, Shaker R, Sengupta JN. Altered expression of P2X3 in vagal and spinal afferents following esophagitis in rats. Histochem Cell Biol 132: 585–597, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bao F, Brown A, Dekaban GA, Omana V, Weaver LC. CD11d integrin blockade reduces the systemic inflammatory response syndrome after spinal cord injury. Exp Neurol 231: 272–283, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Basso DM, Beattie MS, Bresnahan JC. Graded histological and locomotor outcomes after spinal cord contusion using the NYU weight-drop device vs. transection. Exp Neurol 139: 244–256, 1996. [DOI] [PubMed] [Google Scholar]
- 14.Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotor rating scale for open field testing in rats. J Neurotrauma 12: 1–21, 1995. [DOI] [PubMed] [Google Scholar]
- 15.Bennett DL, Michael GJ, Ramachandran N, Munson JB, Averill S, Yan Q, McMahon SB, Priestley JV. A distinct subgroup of small DRG cells express GDNF receptor components and GDNF is protective for these neurons after nerve injury. J Neurosci 18: 3059–3072, 1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Berthoud HR, Lynn PA, Blackshaw LA. Vagal and spinal mechanosensors in the rat stomach and colon have multiple receptive fields. Am J Physiol Regul Integr Comp Physiol 280: R1371–R1381, 2001. [DOI] [PubMed] [Google Scholar]
- 17.Berthoud HR, Neuhuber WL. Functional and chemical anatomy of the afferent vagal system. Auton Neurosci 85: 1–17, 2000. [DOI] [PubMed] [Google Scholar]
- 18.Berthoud HR, Powley TL. Vagal afferent innervation of the rat fundic stomach: morphological characterization of the gastric tension receptor. J Comp Neurol 319: 261–276, 1992. [DOI] [PubMed] [Google Scholar]
- 19.Birder L, Andersson KE. Urothelial signaling. Physiol Rev 93: 653–680, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bobrow MN, Harris TD, Shaughnessy KJ, Litt GJ. Catalyzed reporter deposition, a novel method of signal amplification. Application to immunoassays. J Immunol Methods 125: 279–285, 1989. [DOI] [PubMed] [Google Scholar]
- 21.Bobrow MN, Litt GJ, Shaughnessy KJ, Mayer PC, Conlon J. The use of catalyzed reporter deposition as a means of signal amplification in a variety of formats. J Immunol Methods 150: 145–149, 1992. [DOI] [PubMed] [Google Scholar]
- 22.Bobrow MN, Shaughnessy KJ, Litt GJ. Catalyzed reporter deposition, a novel method of signal amplification. II. Application to membrane immunoassays. J Immunol Methods 137: 103–112, 1991. [DOI] [PubMed] [Google Scholar]
- 23.Bogen O, Dina OA, Gear RW, Levine JD. Dependence of monocyte chemoattractant protein 1 induced hyperalgesia on the isolectin B4-binding protein versican. Neuroscience 159: 780–786, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bogen O, Dreger M, Gillen C, Schroder W, Hucho F. Identification of versican as an isolectin B4-binding glycoprotein from mammalian spinal cord tissue. FEBS J 272: 1090–1102, 2005. [DOI] [PubMed] [Google Scholar]
- 25.Bradbury EJ, Burnstock G, McMahon SB. The expression of P2X3 purinoreceptors in sensory neurons: effects of axotomy and glial-derived neurotrophic factor. Mol Cell Neurosci 12: 256–268, 1998. [DOI] [PubMed] [Google Scholar]
- 26.Browning KN, Mendelowitz D. Musings on the wanderer: what's new in our understanding of vago-vagal reflexes? II. Integration of afferent signaling from the viscera by the nodose ganglia. Am J Physiol Gastrointest Liver Physiol 284: G8–G14, 2003. [DOI] [PubMed] [Google Scholar]
- 27.Bueno L, Fioramonti J. Visceral perception: inflammatory and non-inflammatory mediators. Gut 51, Suppl 1: i19–i23, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Burden HW, Leonard M, Smith CP, Lawrence IE Jr.. The sensory innervation of the ovary: a horseradish peroxidase study in the rat. Anat Rec 207: 623–627, 1983. [DOI] [PubMed] [Google Scholar]
- 29.Burnstock G. Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev 87: 659–797, 2007. [DOI] [PubMed] [Google Scholar]
- 30.Burnstock G. A unifying purinergic hypothesis for the initiation of pain. Lancet 347: 1604–1605, 1996. [DOI] [PubMed] [Google Scholar]
- 31.Campbell SJ, Perry VH, Pitossi FJ, Butchart AG, Chertoff M, Waters S, Dempster R, Anthony DC. Central nervous system injury triggers hepatic CC and CXC chemokine expression that is associated with leukocyte mobilization and recruitment to both the central nervous system and the liver. Am J Pathol 166: 1487–1497, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cervero F. Sensory innervation of the viscera: peripheral basis of visceral pain. Physiol Rev 74: 95–138, 1994. [DOI] [PubMed] [Google Scholar]
- 33.Chen CC, Akopian AN, Sivilotti L, Colquhoun D, Burnstock G, Wood JN. A P2X purinoceptor expressed by a subset of sensory neurons. Nature 377: 428–431, 1995. [DOI] [PubMed] [Google Scholar]
- 34.Chen XQ, Wang B, Wu C, Pan J, Yuan B, Su YY, Jiang XY, Zhang X, Bao L. Endosome-mediated retrograde axonal transport of P2X3 receptor signals in primary sensory neurons. Cell Res 22: 677–696, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Christianson JA, Traub RJ, Davis BM. Differences in spinal distribution and neurochemical phenotype of colonic afferents in mouse and rat. J Comp Neurol 494: 246–259, 2006. [DOI] [PubMed] [Google Scholar]
- 36.Cockayne DA, Hamilton SG, Zhu QM, Dunn PM, Zhong Y, Novakovic S, Malmberg AB, Cain G, Berson A, Kassotakis L, Hedley L, Lachnit WG, Burnstock G, McMahon SB, Ford AP. Urinary bladder hyporeflexia and reduced pain-related behaviour in P2X3-deficient mice. Nature 407: 1011–1015, 2000. [DOI] [PubMed] [Google Scholar]
- 37.Coggeshall RE, Lekan HA. Methods for determining numbers of cells and synapses: a case for more uniform standards of review. J Comp Neurol 364: 6–15, 1996. [DOI] [PubMed] [Google Scholar]
- 38.Collins JJ, Lin CE, Berthoud HR, Papka RE. Vagal afferents from the uterus and cervix provide direct connections to the brainstem. Cell Tissue Res 295: 43–54, 1999. [DOI] [PubMed] [Google Scholar]
- 39.Cook SP, McCleskey EW. Cell damage excites nociceptors through release of cytosolic ATP. Pain 95: 41–47, 2002. [DOI] [PubMed] [Google Scholar]
- 40.Cook SP, Vulchanova L, Hargreaves KM, Elde R, McCleskey EW. Distinct ATP receptors on pain-sensing and stretch-sensing neurons. Nature 387: 505–508, 1997. [DOI] [PubMed] [Google Scholar]
- 41.D'Arco M, Giniatullin R, Simonetti M, Fabbro A, Nair A, Nistri A, Fabbretti E. Neutralization of nerve growth factor induces plasticity of ATP-sensitive P2X3 receptors of nociceptive trigeminal ganglion neurons. J Neurosci 27: 8190–8201, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Dai Y, Fukuoka T, Wang H, Yamanaka H, Obata K, Tokunaga A, Noguchi K. Contribution of sensitized P2X receptors in inflamed tissue to the mechanical hypersensitivity revealed by phosphorylated ERK in DRG neurons. Pain 108: 258–266, 2004. [DOI] [PubMed] [Google Scholar]
- 43.Dang K, Bielfeldt K, Lamb K, Gebhart GF. Gastric ulcers evoke hyperexcitability and enhance P2X receptor function in rat gastric sensory neurons. J Neurophysiol 93: 3112–3119, 2005. [DOI] [PubMed] [Google Scholar]
- 44.Danzer M, Jocic M, Samberger C, Painsipp E, Bock E, Pabst MA, Crailsheim K, Schicho R, Lippe IT, Holzer P. Stomach-brain communication by vagal afferents in response to luminal acid backdiffusion, gastrin, and gastric acid secretion. Am J Physiol Gastrointest Liver Physiol 286: G403–G411, 2004. [DOI] [PubMed] [Google Scholar]
- 45.de Groat WC, Kawatani M, Hisamitsu T, Cheng CL, Ma CP, Thor K, Steers W, Roppolo JR. Mechanisms underlying the recovery of urinary bladder function following spinal cord injury. J Auton Nerv Syst 30, Suppl: S71–S77, 1990. [DOI] [PubMed] [Google Scholar]
- 46.de Groat WC, Kruse MN, Vizzard MA, Cheng CL, Araki I, Yoshimura N. Modification of urinary bladder function after spinal cord injury. Adv Neurol 72: 347–364, 1997. [PubMed] [Google Scholar]
- 47.Ditunno PL, Patrick M, Stineman M, Ditunno JF. Who wants to walk? Preferences for recovery after SCI: a longitudinal and cross-sectional study. Spinal Cord 46: 500–506, 2008. [DOI] [PubMed] [Google Scholar]
- 48.Domotor A, Peidl Z, Vincze A, Hunyady B, Szolcsanyi J, Kereskay L, Szekeres G, Mozsik G. Immunohistochemical distribution of vanilloid receptor, calcitonin-gene related peptide and substance P in gastrointestinal mucosa of patients with different gastrointestinal disorders. Inflammopharmacology 13: 161–177, 2005. [DOI] [PubMed] [Google Scholar]
- 49.Dutsch M, Eichhorn U, Worl J, Wank M, Berthoud HR, Neuhuber WL. Vagal and spinal afferent innervation of the rat esophagus: a combined retrograde tracing and immunocytochemical study with special emphasis on calcium-binding proteins. J Comp Neurol 398: 289–307, 1998. [PubMed] [Google Scholar]
- 50.Eriksson J, Bongenhielm U, Kidd E, Matthews B, Fried K. Distribution of P2X3 receptors in the rat trigeminal ganglion after inferior alveolar nerve injury. Neurosci Lett 254: 37–40, 1998. [DOI] [PubMed] [Google Scholar]
- 51.Ersoz M, Akyuz M. Bladder-filling sensation in patients with spinal cord injury and the potential for sensation-dependent bladder emptying. Spinal Cord 42: 110–116, 2004. [DOI] [PubMed] [Google Scholar]
- 52.Fabbretti E, D'Arco M, Fabbro A, Simonetti M, Nistri A, Giniatullin R. Delayed upregulation of ATP P2X3 receptors of trigeminal sensory neurons by calcitonin gene-related peptide. J Neurosci 26: 6163–6171, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Fang M, Wang J, Huang JY, Ling SC, Rudd JA, Hu ZY, Yew DT, Han S. The neuroprotective effects of Reg-2 following spinal cord transection injury. Anat Rec (Hoboken) 294: 24–45, 2011. [DOI] [PubMed] [Google Scholar]
- 54.Fasanella KE, Christianson JA, Chanthaphavong RS, Davis BM. Distribution and neurochemical identification of pancreatic afferents in the mouse. J Comp Neurol 509: 42–52, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Fleming JC, Norenberg MD, Ramsay DA, Dekaban GA, Marcillo AE, Saenz AD, Pasquale-Styles M, Dietrich WD, Weaver LC. The cellular inflammatory response in human spinal cords after injury. Brain 129: 3249–3269, 2006. [DOI] [PubMed] [Google Scholar]
- 56.Fowler CJ, Griffiths D, de Groat WC. The neural control of micturition. Nat Rev Neurosci 9: 453–466, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gattone VH 2nd, Marfurt CF, Dallie S. Extrinsic innervation of the rat kidney: a retrograde tracing study. Am J Physiol Renal Fluid Electrolyte Physiol 250: F189–F196, 1986. [DOI] [PubMed] [Google Scholar]
- 58.Giniatullin R, Nistri A, Fabbretti E. Molecular mechanisms of sensitization of pain-transducing P2X3 receptors by the migraine mediators CGRP and NGF. Mol Neurobiol 37: 83–90, 2008. [DOI] [PubMed] [Google Scholar]
- 59.Gittins R, Harrison PJ. Neuronal density, size and shape in the human anterior cingulate cortex: a comparison of Nissl and NeuN staining. Brain Res Bull 63: 155–160, 2004. [DOI] [PubMed] [Google Scholar]
- 60.Gondim FA, Alencar HM, Rodrigues CL, da Graca JR, dos Santos AA, Rola FH. Complete cervical or thoracic spinal cord transections delay gastric emptying and gastrointestinal transit of liquid in awake rats. Spinal Cord 37: 793–799, 1999. [DOI] [PubMed] [Google Scholar]
- 61.Gondim FA, Rodrigues CL, da Graca JR, Camurca FD, de Alencar HM, dos Santos AA, Rola FH. Neural mechanisms involved in the delay of gastric emptying and gastrointestinal transit of liquid after thoracic spinal cord transection in awake rats. Auton Neurosci 87: 52–58, 2001. [DOI] [PubMed] [Google Scholar]
- 62.Gore RM, Mintzer RA, Calenoff L. Gastrointestinal complications of spinal cord injury. Spine (Phila Pa 1976) 6: 538–544, 1981. [DOI] [PubMed] [Google Scholar]
- 63.Green T, Dockray GJ. Calcitonin gene-related peptide and substance P in afferents to the upper gastrointestinal tract in the rat. Neurosci Lett 76: 151–156, 1987. [DOI] [PubMed] [Google Scholar]
- 64.Green T, Dockray GJ. Characterization of the peptidergic afferent innervation of the stomach in the rat, mouse and guinea-pig. Neuroscience 25: 181–193, 1988. [DOI] [PubMed] [Google Scholar]
- 65.Gris D, Hamilton EF, Weaver LC. The systemic inflammatory response after spinal cord injury damages lungs and kidneys. Exp Neurol 211: 259–270, 2008. [DOI] [PubMed] [Google Scholar]
- 66.Grundy D. Neuroanatomy of visceral nociception: vagal and splanchnic afferent. Gut 51, Suppl 1: i2–i5, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hammell KR. Spinal cord injury rehabilitation research: patient priorities, current deficiencies and potential directions. Disabil Rehabil 32: 1209–1218, 2010. [DOI] [PubMed] [Google Scholar]
- 68.Hayakawa T, Kuwahara S, Maeda S, Tanaka K, Seki M. Calcitonin gene-related peptide immunoreactive neurons innervating the soft palate, the root of tongue, and the pharynx in the superior glossopharyngeal ganglion of the rat. J Chem Neuroanat 39: 221–227, 2010. [DOI] [PubMed] [Google Scholar]
- 69.Helke CJ. Vagal Afferent Neurons: Neurotrophic Factors and Epigenetic Influences. Boca Raton, FL: CRC Press, Taylor and Francis Group, 2005. [Google Scholar]
- 70.Helke CJ, Adryan KM, Fedorowicz J, Zhuo H, Park JS, Curtis R, Radley HE, Distefano PS. Axonal transport of neurotrophins by visceral afferent and efferent neurons of the vagus nerve of the rat. J Comp Neurol 393: 102–117, 1998. [DOI] [PubMed] [Google Scholar]
- 71.Helke CJ, Hill KM. Immunohistochemical study of neuropeptides in vagal and glossopharyngeal afferent neurons in the rat. Neuroscience 26: 539–551, 1988. [DOI] [PubMed] [Google Scholar]
- 72.Helke CJ, Niederer AJ. Studies on the coexistence of substance P with other putative transmitters in the nodose and petrosal ganglia. Synapse 5: 144–151, 1990. [DOI] [PubMed] [Google Scholar]
- 73.Helke CJ, O'Donohue TL, Jacobowitz DM. Substance P as a baro- and chemoreceptor afferent neurotransmitter: immunocytochemical and neurochemical evidence in the rat. Peptides 1: 1–9, 1980. [DOI] [PubMed] [Google Scholar]
- 74.Helke CJ, Rabchevsky A. Axotomy alters putative neurotransmitters in visceral sensory neurons of the nodose and petrosal ganglia. Brain Res 551: 44–51, 1991. [DOI] [PubMed] [Google Scholar]
- 75.Herculano-Houzel S, Lent R. Isotropic fractionator: a simple, rapid method for the quantification of total cell and neuron numbers in the brain. J Neurosci 25: 2518–2521, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Herrera JJ, Haywood-Watson RJ 2nd, Grill RJ. Acute and chronic deficits in the urinary bladder after spinal contusion injury in the adult rat. J Neurotrauma 27: 423–431, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Herrity AN, Rau KK, Petruska JC, Stirling DP, Hubscher CH. Identification of bladder and colon afferents in the nodose ganglia of male rats. J Comp Neurol 522: 3667–3682, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hill CE, Harrison BJ, Rau KK, Hougland MT, Bunge MB, Mendell LM, Petruska JC. Skin incision induces expression of axonal regeneration-related genes in adult rat spinal sensory neurons. J Pain 11: 1066–1073, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Holmes GM. Upper gastrointestinal dysmotility after spinal cord injury: is diminished vagal sensory processing one culprit? Front Physiol 3: 277, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Holzer P. Afferent signalling of gastric acid challenge. J Physiol Pharmacol 54, Suppl 4: 43–53, 2003. [PubMed] [Google Scholar]
- 81.Hubscher CH, Berkley KJ. Spinal and vagal influences on the responses of rat solitary nucleus neurons to stimulation of uterus, cervix and vagina. Brain Res 702: 251–254, 1995. [DOI] [PubMed] [Google Scholar]
- 82.Hubscher CH, Johnson RD. Effects of acute and chronic midthoracic spinal cord injury on neural circuits for male sexual function. II. Descending pathways. J Neurophysiol 83: 2508–2518, 2000. [DOI] [PubMed] [Google Scholar]
- 83.Hubscher CH, Petruska JC, Rau KK, Johnson RD. Co-expression of P2X receptor subunits on rat nodose neurons that bind the isolectin GS-I-B4. Neuroreport 12: 2995–2997, 2001. [DOI] [PubMed] [Google Scholar]
- 84.Jancso G, Maggi CA. Distribution of capsaicin-sensitive urinary bladder afferents in the rat spinal cord. Brain Res 418: 371–376, 1987. [DOI] [PubMed] [Google Scholar]
- 85.Janig W. Neurobiology of visceral afferent neurons: neuroanatomy, functions, organ regulations and sensations. Biol Psychol 42: 29–51, 1996. [DOI] [PubMed] [Google Scholar]
- 86.Kaddumi EG, Hubscher CH. Urinary bladder irritation alters efficacy of vagal stimulation on rostral medullary neurons in chronic T8 spinalized rats. J Neurotrauma 24: 1219–1228, 2007. [DOI] [PubMed] [Google Scholar]
- 87.Kao CH, Ho YJ, Changlai SP, Ding HJ. Gastric emptying in spinal cord injury patients. Dig Dis Sci 44: 1512–1515, 1999. [DOI] [PubMed] [Google Scholar]
- 88.Kestler C, Neuhuber WL, Raab M. Distribution of P2X(3) receptor immunoreactivity in myenteric ganglia of the mouse esophagus. Histochem Cell Biol 131: 13–27, 2009. [DOI] [PubMed] [Google Scholar]
- 89.Khakh BS, North RA. P2X receptors as cell-surface ATP sensors in health and disease. Nature 442: 527–532, 2006. [DOI] [PubMed] [Google Scholar]
- 90.Klusman I, Schwab ME. Effects of pro-inflammatory cytokines in experimental spinal cord injury. Brain Res 762: 173–184, 1997. [DOI] [PubMed] [Google Scholar]
- 91.Koerber HR, Mirnics K, Kavookjian AM, Light AR. Ultrastructural analysis of ectopic synaptic boutons arising from peripherally regenerated primary afferent fibers. J Neurophysiol 81: 1636–1644, 1999. [DOI] [PubMed] [Google Scholar]
- 92.Komisaruk BR, Gerdes CA, Whipple B. ‘Complete’ spinal cord injury does not block perceptual responses to genital self-stimulation in women. Arch Neurol 54: 1513–1520, 1997. [DOI] [PubMed] [Google Scholar]
- 93.Kruse MN, Bray LA, de Groat WC. Influence of spinal cord injury on the morphology of bladder afferent and efferent neurons. J Auton Nerv Syst 54: 215–224, 1995. [DOI] [PubMed] [Google Scholar]
- 94.Kruse MN, de Groat WC. Changes in lower urinary tract function following spinal cord injury. Restor Neurol Neurosci 5: 79–80, 1993. [DOI] [PubMed] [Google Scholar]
- 95.Kuypers NJ, James KT, Enzmann GU, Magnuson DS, Whittemore SR. Functional consequences of ethidium bromide demyelination of the mouse ventral spinal cord. Exp Neurol 247: 615–622, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Lamb K, Bielefeldt K. Rapid effects of neurotrophic factors on calcium homeostasis in rat visceral afferent neurons. Neurosci Lett 336: 9–12, 2003. [DOI] [PubMed] [Google Scholar]
- 97.Lewis C, Neidhart S, Holy C, North RA, Buell G, Surprenant A. Coexpression of P2X2 and P2X3 receptor subunits can account for ATP-gated currents in sensory neurons. Nature 377: 432–435, 1995. [DOI] [PubMed] [Google Scholar]
- 98.Li H, Nomura S, Mizuno N. Binding of the isolectin I-B4 from Griffonia simplicifolia to the general visceral afferents in the vagus nerve: a light- and electron-microscope study in the rat. Neurosci Lett 222: 53–56, 1997. [DOI] [PubMed] [Google Scholar]
- 99.Li L, Zhou XF. Pericellular Griffonia simplicifolia I isolectin B4-binding ring structures in the dorsal root ganglia following peripheral nerve injury in rats. J Comp Neurol 439: 259–274, 2001. [DOI] [PubMed] [Google Scholar]
- 100.Liu F, Takahashi N, Yamaguchi O. Expression of P2X3 purinoceptors in suburothelial myofibroblasts of the normal human urinary bladder. Int J Urol 16: 570–575, 2009. [DOI] [PubMed] [Google Scholar]
- 101.Ma VY, Chan L, Carruthers KJ. Incidence, prevalence, costs, and impact on disability of common conditions requiring rehabilitation in the United States: stroke, spinal cord injury, traumatic brain injury, multiple sclerosis, osteoarthritis, rheumatoid arthritis, limb loss, and back pain. Arch Phys Med Rehabil 95: 986–995, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Maley BE. Immunohistochemical localization of neuropeptides and neurotransmitters in the nucleus solitarius. Chem Senses 21: 367–376, 1996. [DOI] [PubMed] [Google Scholar]
- 103.Michael GJ, Priestley JV. Differential expression of the mRNA for the vanilloid receptor subtype 1 in cells of the adult rat dorsal root and nodose ganglia and its downregulation by axotomy. J Neurosci 19: 1844–1854, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Molliver DC, Wright DE, Leitner ML, Parsadanian AS, Doster K, Wen D, Yan Q, Snider WD. IB4-binding DRG neurons switch from NGF to GDNF dependence in early postnatal life. Neuron 19: 849–861, 1997. [DOI] [PubMed] [Google Scholar]
- 105.Mullen RJ, Buck CR, Smith AM. NeuN, a neuronal specific nuclear protein in vertebrates. Development 116: 201–211, 1992. [DOI] [PubMed] [Google Scholar]
- 106.Myers AC, Kajekar R, Undem BJ. Allergic inflammation-induced neuropeptide production in rapidly adapting afferent nerves in guinea pig airways. Am J Physiol Lung Cell Mol Physiol 282: L775–L781, 2002. [DOI] [PubMed] [Google Scholar]
- 107.Neumann S, Doubell TP, Leslie T, Woolf CJ. Inflammatory pain hypersensitivity mediated by phenotypic switch in myelinated primary sensory neurons. Nature 384: 360–364, 1996. [DOI] [PubMed] [Google Scholar]
- 108.North RA. P2X3 receptors and peripheral pain mechanisms. J Physiol 554: 301–308, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Novakovic SD, Kassotakis LC, Oglesby IB, Smith JA, Eglen RM, Ford AP, Hunter JC. Immunocytochemical localization of P2X3 purinoceptors in sensory neurons in naive rats and following neuropathic injury. Pain 80: 273–282, 1999. [DOI] [PubMed] [Google Scholar]
- 110.Oddiah D, Anand P, McMahon SB, Rattray M. Rapid increase of NGF, BDNF and NT-3 mRNAs in inflamed bladder. Neuroreport 9: 1455–1458, 1998. [DOI] [PubMed] [Google Scholar]
- 111.Ortega-Villalobos M, Garcia-Bazan M, Solano-Flores LP, Ninomiya-Alarcon JG, Guevara-Guzman R, Wayner MJ. Vagus nerve afferent and efferent innervation of the rat uterus: an electrophysiological and HRP study. Brain Res Bull 25: 365–371, 1990. [DOI] [PubMed] [Google Scholar]
- 112.Ozaki N, Sengupta JN, Gebhart GF. Mechanosensitive properties of gastric vagal afferent fibers in the rat. J Neurophysiol 82: 2210–2220, 1999. [DOI] [PubMed] [Google Scholar]
- 113.Pakkenberg B, Gundersen HJ. Total number of neurons and glial cells in human brain nuclei estimated by the disector and the fractionator. J Microsc 150: 1–20, 1988. [DOI] [PubMed] [Google Scholar]
- 114.Pan XQ, Gonzalez JA, Chang S, Chacko S, Wein AJ, Malykhina AP. Experimental colitis triggers the release of substance P and calcitonin gene-related peptide in the urinary bladder via TRPV1 signaling pathways. Exp Neurol 225: 262–273, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Peters JH, Gallaher ZR, Ryu V, Czaja K. Withdrawal and restoration of central vagal afferents within the dorsal vagal complex following subdiaphragmatic vagotomy. J Comp Neurol 521: 3584–3599, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Petruska JC, Cooper BY, Gu JG, Rau KK, Johnson RD. Distribution of P2X1, P2X2, and P2X3 receptor subunits in rat primary afferents: relation to population markers and specific cell types. J Chem Neuroanat 20: 141–162, 2000. [DOI] [PubMed] [Google Scholar]
- 117.Petruska JC, Napaporn J, Johnson RD, Cooper BY. Chemical responsiveness and histochemical phenotype of electrophysiologically classified cells of the adult rat dorsal root ganglion. Neuroscience 115: 15–30, 2002. [DOI] [PubMed] [Google Scholar]
- 118.Peyronnard JM, Charron L, Messier JP, Lavoie J, Leger C, Faraco-Cantin F. Changes in lectin binding of lumbar dorsal root ganglia neurons and peripheral axons after sciatic and spinal nerve injury in the rat. Cell Tissue Res 257: 379–388, 1989. [DOI] [PubMed] [Google Scholar]
- 119.Phillips RJ, Powley TL. Tension and stretch receptors in gastrointestinal smooth muscle: re-evaluating vagal mechanoreceptor electrophysiology. Brain Res Brain Res Rev 34: 1–26, 2000. [DOI] [PubMed] [Google Scholar]
- 120.Plato M, Kummer W, Haberberger RV. Structural and neurochemical comparison of vagal and spinal afferent neurons projecting to the rat lung. Neurosci Lett 395: 215–219, 2006. [DOI] [PubMed] [Google Scholar]
- 121.Popovich PG, Wei P, Stokes BT. Cellular inflammatory response after spinal cord injury in Sprague-Dawley and Lewis rats. J Comp Neurol 377: 443–464, 1997. [DOI] [PubMed] [Google Scholar]
- 122.Qiao LY, Vizzard MA. Spinal cord injury-induced expression of TrkA, TrkB, phosphorylated CREB, and c-Jun in rat lumbosacral dorsal root ganglia. J Comp Neurol 482: 142–154, 2005. [DOI] [PubMed] [Google Scholar]
- 123.Rajendran SK, Reiser JR, Bauman W, Zhang RL, Gordon SK, Korsten MA. Gastrointestinal transit after spinal cord injury: effect of cisapride. Am J Gastroenterol 87: 1614–1617, 1992. [PubMed] [Google Scholar]
- 124.Rau KK, Jiang N, Johnson RD, Cooper BY. Heat sensitization in skin and muscle nociceptors expressing distinct combinations of TRPV1 and TRPV2 protein. J Neurophysiol 97: 2651–2662, 2007. [DOI] [PubMed] [Google Scholar]
- 125.Segal JL, Milne N, Brunnemann SR. Gastric emptying is impaired in patients with spinal cord injury. Am J Gastroenterol 90: 466–470, 1995. [PubMed] [Google Scholar]
- 126.Seki S, Sasaki K, Fraser MO, Igawa Y, Nishizawa O, Chancellor MB, de Groat WC, Yoshimura N. Immunoneutralization of nerve growth factor in lumbosacral spinal cord reduces bladder hyperreflexia in spinal cord injured rats. J Urol 168: 2269–2274, 2002. [DOI] [PubMed] [Google Scholar]
- 127.Sharkey KA, Williams RG. Extrinsic innervation of the rat pancreas: demonstration of vagal sensory neurones in the rat by retrograde tracing. Neurosci Lett 42: 131–135, 1983. [DOI] [PubMed] [Google Scholar]
- 128.Sharkey KA, Williams RG, Dockray GJ. Sensory substance P innervation of the stomach and pancreas. Demonstration of capsaicin-sensitive sensory neurons in the rat by combined immunohistochemistry and retrograde tracing. Gastroenterology 87: 914–921, 1984. [PubMed] [Google Scholar]
- 129.Silverman JD, Kruger L. Selective neuronal glycoconjugate expression in sensory and autonomic ganglia: relation of lectin reactivity to peptide and enzyme markers. J Neurocytol 19: 789–801, 1990. [DOI] [PubMed] [Google Scholar]
- 130.Singh A, Balasubramanian S, Murray M, Lemay M, Houle J. Role of spared pathways in locomotor recovery after body-weight-supported treadmill training in contused rats. J Neurotrauma 28: 2405–2416, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Staikopoulos V, Sessle BJ, Furness JB, Jennings EA. Localization of P2X2 and P2X3 receptors in rat trigeminal ganglion neurons. Neuroscience 144: 208–216, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Steers WD, Tuttle JB. Mechanisms of disease: the role of nerve growth factor in the pathophysiology of bladder disorders. Nat Clin Pract Urol 3: 101–110, 2006. [DOI] [PubMed] [Google Scholar]
- 133.Stinneford JG, Keshavarzian A, Nemchausky BA, Doria MI, Durkin M. Esophagitis and esophageal motor abnormalities in patients with chronic spinal cord injuries. Paraplegia 31: 384–392, 1993. [DOI] [PubMed] [Google Scholar]
- 134.Stirling DP, Yong VW. Dynamics of the inflammatory response after murine spinal cord injury revealed by flow cytometry. J Neurosci Res 86: 1944–1958, 2008. [DOI] [PubMed] [Google Scholar]
- 135.Stucky CL, Lewin GR. Isolectin B(4)-positive and -negative nociceptors are functionally distinct. J Neurosci 19: 6497–6505, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Sugiura T, Dang K, Lamb K, Bielefeldt K, Gebhart GF. Acid-sensing properties in rat gastric sensory neurons from normal and ulcerated stomach. J Neurosci 25: 2617–2627, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Sun Y, Keay S, De Deyne PG, Chai TC. Augmented stretch activated adenosine triphosphate release from bladder uroepithelial cells in patients with interstitial cystitis. J Urol 166: 1951–1956, 2001. [PubMed] [Google Scholar]
- 138.Tanaka M, Uchiyama M, Kitano M. Gastroduodenal disease in chronic spinal cord injuries. An endoscopic study. Arch Surg 114: 185–187, 1979. [DOI] [PubMed] [Google Scholar]
- 139.Tang HB, Li YS, Arihiro K, Nakata Y. Activation of the neurokinin-1 receptor by substance P triggers the release of substance P from cultured adult rat dorsal root ganglion neurons. Mol Pain 3: 42, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Tong M, Holmes GM. Gastric dysreflexia after acute experimental spinal cord injury in rats. Neurogastroenterol Motil 21: 197–206, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Tsujino H, Kondo E, Fukuoka T, Dai Y, Tokunaga A, Miki K, Yonenobu K, Ochi T, Noguchi K. Activating transcription factor 3 (ATF3) induction by axotomy in sensory and motoneurons: A novel neuronal marker of nerve injury. Mol Cell Neurosci 15: 170–182, 2000. [DOI] [PubMed] [Google Scholar]
- 142.Tsuzuki K, Kondo E, Fukuoka T, Yi D, Tsujino H, Sakagami M, Noguchi K. Differential regulation of P2X(3) mRNA expression by peripheral nerve injury in intact and injured neurons in the rat sensory ganglia. Pain 91: 351–360, 2001. [DOI] [PubMed] [Google Scholar]
- 143.Undem BJ, Chuaychoo B, Lee MG, Weinreich D, Myers AC, Kollarik M. Subtypes of vagal afferent C-fibres in guinea-pig lungs. J Physiol 556: 905–917, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Verge VM, Merlio JP, Grondin J, Ernfors P, Persson H, Riopelle RJ, Hokfelt T, Richardson PM. Colocalization of NGF binding sites, trk mRNA, and low-affinity NGF receptor mRNA in primary sensory neurons: responses to injury and infusion of NGF. J Neurosci 12: 4011–4022, 1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Vizzard MA. Alterations in growth-associated protein (GAP-43) expression in lower urinary tract pathways following chronic spinal cord injury. Somatosens Mot Res 16: 369–381, 1999. [DOI] [PubMed] [Google Scholar]
- 146.Vizzard MA. Alterations in neuropeptide expression in lumbosacral bladder pathways following chronic cystitis. J Chem Neuroanat 21: 125–138, 2001. [DOI] [PubMed] [Google Scholar]
- 147.Vizzard MA. Changes in urinary bladder neurotrophic factor mRNA and NGF protein following urinary bladder dysfunction. Exp Neurol 161: 273–284, 2000. [DOI] [PubMed] [Google Scholar]
- 148.Vizzard MA. Neurochemical plasticity and the role of neurotrophic factors in bladder reflex pathways after spinal cord injury. Prog Brain Res 152: 97–115, 2006. [DOI] [PubMed] [Google Scholar]
- 149.Vizzard MA, Brisson M, de Groat WC. Transneuronal labeling of neurons in the adult rat central nervous system following inoculation of pseudorabies virus into the colon. Cell Tissue Res 299: 9–26, 2000. [DOI] [PubMed] [Google Scholar]
- 150.Vulchanova L, Arvidsson U, Riedl M, Wang J, Buell G, Surprenant A, North RA, Elde R. Differential distribution of two ATP-gated channels (P2X receptors) determined by immunocytochemistry. Proc Natl Acad Sci U S A 93: 8063–8067, 1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Vulchanova L, Olson TH, Stone LS, Riedl MS, Elde R, Honda CN. Cytotoxic targeting of isolectin IB4-binding sensory neurons. Neuroscience 108: 143–155, 2001. [DOI] [PubMed] [Google Scholar]
- 152.Vulchanova L, Riedl MS, Shuster SJ, Buell G, Surprenant A, North RA, Elde R. Immunohistochemical study of the P2X2 and P2X3 receptor subunits in rat and monkey sensory neurons and their central terminals. Neuropharmacology 36: 1229–1242, 1997. [DOI] [PubMed] [Google Scholar]
- 153.Vulchanova L, Riedl MS, Shuster SJ, Stone LS, Hargreaves KM, Buell G, Surprenant A, North RA, Elde R. P2X3 is expressed by DRG neurons that terminate in inner lamina II. Eur J Neurosci 10: 3470–3478, 1998. [DOI] [PubMed] [Google Scholar]
- 154.Wang L, Feng D, Yan H, Wang Z, Pei L. Comparative analysis of P2X1, P2X2, P2X3, and P2X4 receptor subunits in rat nodose ganglion neurons. PLos One 9: e96699, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Wang T, Molliver DC, Jing X, Schwartz ES, Yang FC, Samad OA, Ma Q, Davis BM. Phenotypic switching of nonpeptidergic cutaneous sensory neurons following peripheral nerve injury. PLos One 6: e28908, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Wang YH, Tache Y, Sheibel AB, Go VL, Wei JY. Two types of leptin-responsive gastric vagal afferent terminals: an in vitro single-unit study in rats. Am J Physiol Regul Integr Comp Physiol 273: R833–R837, 1997. [DOI] [PubMed] [Google Scholar]
- 157.Wank M, Neuhuber WL. Local differences in vagal afferent innervation of the rat esophagus are reflected by neurochemical differences at the level of the sensory ganglia and by different brainstem projections. J Comp Neurol 435: 41–59, 2001. [DOI] [PubMed] [Google Scholar]
- 158.Ward PJ, Herrity AN, Smith RR, Willhite A, Harrison BJ, Petruska JC, Harkema SJ, Hubscher CH. Novel multi-system functional gains via task specific training in spinal cord injured male rats. J Neurotrauma 31: 819–833, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Williams RE 3rd, Bauman WA, Spungen AM, Vinnakota RR, Farid RZ, Galea M, Korsten MA. SmartPill technology provides safe and effective assessment of gastrointestinal function in persons with spinal cord injury. Spinal Cord 50: 81–84, 2012. [DOI] [PubMed] [Google Scholar]
- 160.Wolf C, Meiners TH. Dysphagia in patients with acute cervical spinal cord injury. Spinal Cord 41: 347–353, 2003. [DOI] [PubMed] [Google Scholar]
- 161.Wu XY, Zhu JX, Gao J, Owyang C, Li Y. Neurochemical phenotype of vagal afferent neurons activated to express C-FOS in response to luminal stimulation in the rat. Neuroscience 130: 757–767, 2005. [DOI] [PubMed] [Google Scholar]
- 162.Wyndaele JJ. Investigation of the afferent nerves of the lower urinary tract in patients with “complete” and “incomplete” spinal cord injury. Paraplegia 29: 490–494, 1991. [DOI] [PubMed] [Google Scholar]
- 163.Wynn G, Ma B, Ruan HZ, Burnstock G. Purinergic component of mechanosensory transduction is increased in a rat model of colitis. Am J Physiol Gastrointest Liver Physiol 287: G647–G657, 2004. [DOI] [PubMed] [Google Scholar]
- 164.Xiang Z, Bo X, Oglesby I, Ford A, Burnstock G. Localization of ATP-gated P2X2 receptor immunoreactivity in the rat hypothalamus. Brain Res 813: 390–397, 1998. [DOI] [PubMed] [Google Scholar]
- 165.Yoshimura N. Bladder afferent pathway and spinal cord injury: possible mechanisms inducing hyperreflexia of the urinary bladder. Prog Neurobiol 57: 583–606, 1999. [DOI] [PubMed] [Google Scholar]
- 166.Yoshimura N, de Groat WC. Plasticity of Na+ channels in afferent neurones innervating rat urinary bladder following spinal cord injury. J Physiol 503: 269–276, 1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Yoshimura N, Erdman SL, Snider MW, de Groat WC. Effects of spinal cord injury on neurofilament immunoreactivity and capsaicin sensitivity in rat dorsal root ganglion neurons innervating the urinary bladder. Neuroscience 83: 633–643, 1998. [DOI] [PubMed] [Google Scholar]
- 168.Yoshimura N, Seki S, Erickson KA, Erickson VL, Hancellor MB, de Groat WC. Histological and electrical properties of rat dorsal root ganglion neurons innervating the lower urinary tract. J Neurosci 23: 4355–4361, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Young RL, Cooper NJ, Blackshaw LA. Chemical coding and central projections of gastric vagal afferent neurons. Neurogastroenterol Motil 20: 708–718, 2008. [DOI] [PubMed] [Google Scholar]
- 170.Yu S, Undem BJ, Kollarik M. Vagal afferent nerves with nociceptive properties in guinea-pig oesophagus. J Physiol 563: 831–842, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Zhang X, Ji RR, Arvidsson J, Lundberg JM, Bartfai T, Bedecs K, Hokfelt T. Expression of peptides, nitric oxide synthase and NPY receptor in trigeminal and nodose ganglia after nerve lesions. Exp Brain Res 111: 393–404, 1996. [DOI] [PubMed] [Google Scholar]
- 172.Zhong F, Christianson JA, Davis BM, Bielefeldt K. Dichotomizing axons in spinal and vagal afferents of the mouse stomach. Dig Dis Sci 53: 194–203, 2008. [DOI] [PubMed] [Google Scholar]
- 173.Zhou XF, Chie ET, Deng YS, Zhong JH, Xue Q, Rush RA, Xian CJ. Injured primary sensory neurons switch phenotype for brain-derived neurotrophic factor in the rat. Neuroscience 92: 841–853, 1999. [DOI] [PubMed] [Google Scholar]
- 174.Zhuo H, Helke CJ. Presence and localization of neurotrophin receptor tyrosine kinase (TrkA, TrkB, TrkC) mRNAs in visceral afferent neurons of the nodose and petrosal ganglia. Brain Res Mol Brain Res 38: 63–70, 1996. [DOI] [PubMed] [Google Scholar]
- 175.Zhuo H, Ichikawa H, Helke CJ. Neurochemistry of the nodose ganglion. Prog Neurobiol 52: 79–107, 1997. [DOI] [PubMed] [Google Scholar]
- 176.Zvarova K, Dunleavy JD, Vizzard MA. Changes in pituitary adenylate cyclase activating polypeptide expression in urinary bladder pathways after spinal cord injury. Exp Neurol 192: 46–59, 2005. [DOI] [PubMed] [Google Scholar]