

Abstract
Gene manipulation using the Cre/loxP-recombinase system has been successfully employed in zebrafish to study gene functions and lineage relationships. Recently, gene trapping approaches have been applied to produce large collections of transgenic fish expressing conditional alleles in various tissues. However, the limited number of available cell- and tissue-specific Cre/CreERT2-driver lines still constrains widespread application in this model organism. To enlarge the pool of existing CreERT2-driver lines, we performed a genome-wide gene trap screen using a Tol2-based mCherry-T2a-CreERT2 (mCT2aC) gene trap vector. This cassette consists of a splice acceptor and a mCherry-tagged variant of CreERT2 which enables simultaneous labeling of the trapping event, as well as CreERT2 expression from the endogenous promoter. Using this strategy, we generated 27 novel functional CreERT2-driver lines expressing in a cell- and tissue-specific manner during development and adulthood. This study summarizes the analysis of the generated CreERT2-driver lines with respect to functionality, expression, integration, as well as associated phenotypes. Our results significantly enlarge the existing pool of CreERT2-driver lines in zebrafish and combined with Cre–dependent effector lines, the new CreERT2-driver lines will be important tools to manipulate the zebrafish genome.
Introduction
Zebrafish has become an excellent model system to understand gene function in vertebrate development and disease. Several advantages, such as the optical clarity of its embryos, short generation time and large number of offspring enable for large-scale forward mutagenesis [1–6] screens as well as real-time in vivo imaging [7–9]. Furthermore, reverse genetic techniques including morpholino-mediated gene knock-down [10], Targeting Induced Local Lesions IN Genomes (TILLING) [11], and targeted gene modification using engineered endonucleases like TALENs or CRISPR/Cas systems [12–16] allow to interfere with zebrafish gene function. In addition, site-specific recombinases (SSRs), which have been an invaluable tool for altering the mouse and fly genome [17–20], have been successfully applied in zebrafish [21–24]. Cre (Causes recombination of the bacteriophage P1 genome) and other SSRs permit for effective conditional mutagenesis and genetic fate mapping, using a common mechanism of DNA recombination including strand cleavage, exchange and ligation [25–27], which is mediated through defined target sites (loxP sites). To achieve temporal control of the recombination process, ligand-inducible forms have been developed. To this end, ligand-binding domains (LBDs) from homodimeric nuclear receptors, such as the human estrogen receptor (ER), have been used to generate CreER [28] fusions. At the moment, CreERT2 shows the best properties in terms of ligand sensitivity and inducible recombination efficiency [29]. Upon administration of Tamoxifen (TAM) or its metabolite 4-hydroxy-Tamoxifen (4-OHT), a conformational change of the LBD mediates translocation of the fusion protein from the cytoplasm into the nucleus and leads to subsequent site-specific recombination. Depending on the nature of the Cre-effector constructs, application of site-specific approaches allows e.g. for cell lineage tracing [22], genetic ablation [30, 31], misexpression studies [32] or conditional gene activity [33–35].
Whereas genome-wide approaches have been conducted to create Cre-effector lines [33–35], the limited number of available cell- and tissue-specific Cre/CreERT2-driver lines still restricts its widespread application in zebrafish [36]. Broad expression of Cre/CreERT2 can be achieved using the inducible heat shock cognate 70-kd protein, like (hsp70l) [21, 37] or the ubiquitin b (ubb) promoter [38] and tissue-restricted Cre/CreERT2-driver lines have been reported that allow genetic lineage labeling studies or transgene overexpression. However, at present only 59 Cre/CreERT2-driver lines have been described in zebrafish (Table 1), whereas more than 2000 driver lines have been created in the mouse model so far [39–41]. Recently, trapping approaches have been applied to produce large collections of transgenic fish expressing conditional alleles in various tissues [34, 35, 42, 43]. Enhancer trapping screens yielded a large library of tissue-specific reporter and driver lines [44]. However, non-specific background expression and also promoter-dependent integration biases reveal limitations of this trapping strategy [45–48]. In addition to enhancer trapping, multiple gene trapping approaches have been applied to dissect the zebrafish genome [5, 33–35, 43]. Gene trapping enables for transgene expression driven by the endogenous promoter and can also be advantageous over transposon-mediated transgenesis using promoter fragments, which often do not faithfully recapitulate the endogenous expression level [22, 49]. In contrast, gene traps fully recapitulate the endogenous gene expression pattern without background expression unless the trapping event interferes with post‐transcriptional regulation of gene expression [49, 50]. Thus in general, gene trapping provides a fast and unbiased method to create tissue-specific driver lines on a large scale basis.
Table 1. Summary of currently available Cre/CreERT2-driver lines in zebrafish.
| Nr. | Cre-driver line | Conditional Cre-driver line | Reference |
|---|---|---|---|
| 1 | Tg(hsp70l:EGFP-cre)ku1 | Thummel R, Burket CT, Brewer JL, Sarras MP, Jr., Li L, et al. (2005) Cre-mediated site-specific recombination in zebrafish embryos. Dev Dyn 233: 1366–1377. | |
| 2 | Tg(hsp70l:Cre)zdf13 | [Feng H, Langenau DM, Madge JA, Quinkertz A, Gutierrez A, et al. (2007) Heat-shock induction of T-cell lymphoma/leukaemia in conditional Cre/lox-regulated transgenic zebrafish. Br J Haematol 138: 169–175 | |
| 3 | Tg(zp3:cre; krt8:rfp)gz14 | [Liu X, Li Z, Emelyanov A, Parinov S, Gong Z (2008) Generation of oocyte-specifically expressed cre transgenic zebrafish for female germline excision of loxP-flanked transgene. Dev Dyn 237: 2955–2962. | |
| 4 | Tg(pax2a:CreERT2)tud101 | Hans S, Kaslin J, Freudenreich D, Brand M (2009) Temporally-controlled site-specific recombination in zebrafish. PLoS One 4: e4640 | |
| 5 | Tg(pax2a:CreERT2)tud102 | Hans S, Kaslin J, Freudenreich D, Brand M (2009) Temporally-controlled site-specific recombination in zebrafish. PLoS One 4: e4640 | |
| 6 | Tg(-1.5ins:Cre,-.58cryaa:Venus)s924 | [Hesselson D, Anderson RM, Beinat M, Stainier DY (2009) Distinct populations of quiescent and proliferative pancreatic beta-cells identified by HOTcre mediated labeling. Proc Natl Acad Sci U S A 106: 14896–14901. | |
| 7 | Tg(-1.5hsp70l:Cre)vu297 | [6 Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W (2009) FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis 47: 484–491. | |
| 8 | Tg(-1.8myl7:Cre)vu300 | [Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W (2009) FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis 47: 484–491. | |
| 9 | Tg(-3her4.1:ERT2-CreERT2)vu298a | [Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W (2009) FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis 47: 484–491. | |
| 10 | Tg(-3her4.1:ERT2-CreERT2)vu298b | [Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W (2009) FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis 47: 484–491. | |
| 11 | Tg(-3her4.1:ERT2-CreERT2)vu298c | Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W (2009) FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis 47: 484–491. | |
| 12 | Tg(-3her4.1:CreERT2)vu299 | Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W (2009) FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis 47: 484–491. | |
| 13 | Tg(-1myl7:ERT2-CreERT2-IRES-mCherry)be1 | Jopling C, Sleep E, Raya M, Marti M, Raya A, et al. (2010) Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature 464: 606–609. | |
| 14 | Tg(Cau.Tuba1a:CreERT2,Cau.Tuba1a:CFP)mi19/+ | Ramachandran R, Reifler A, Parent JM, Goldman D (2010) Conditional gene expression and lineage tracing of tuba1a expressing cells during zebrafish development and retina regeneration. J Comp Neurol 518: 4196–4212 | |
| 15 | Tg(kdrl:Cre)s898 | Bertrand JY, Chi NC, Santoso B, Teng S, Stainier DY, et al. (2010) Haematopoietic stem cells derive directly from aortic endothelium during development. Nature 464: 108–111. | |
| 16 | Tg(cryaa:DsRed,-5.1myl7:CreERT2)pd12 | Liu J, Bressan M, Hassel D, Huisken J, Staudt D, et al. (2010) A dual role for ErbB2 signaling in cardiac trabeculation. Development 137: 3867–3875. | |
| 17 | Tg(-14.8gata4:ERT2-CreERT2)pd39 | Kikuchi K, Holdway JE, Werdich AA, Anderson RM, Fang Y, et al. (2010) Primary contribution to zebrafish heart regeneration by gata4(+) cardiomyocytes. Nature 464: 601–605. | |
| 18 | Tg(actb2:Cerulean-Cre)ct5000 | Trinh le A, Hochgreb T, Graham M, Wu D, Ruf-Zamojski F, et al. (2011) A versatile gene trap to visualize and interrogate the function of the vertebrate proteome. Genes Dev 25: 2306–2320. | |
| 19 | Tg(lmo2:Cre)rj5 | Zhou T, Wang L, Zhu KY, Dong M, Xu PF, et al. (2011) Dominant-negative C/ebpalpha and polycomb group protein Bmi1 extend short-lived hematopoietic stem/progenitor cell life span and induce lethal dyserythropoiesis. Blood 118: 3842–3852. | |
| 20 | TgBAC(-25ltbp3:TagRFP-Cre)fb1 | Zhou T, Wang L, Zhu KY, Dong M, Xu PF, et al. (2011) Dominant-negative C/ebpalpha and polycomb group protein Bmi1 extend short-lived hematopoietic stem/progenitor cell life span and induce lethal dyserythropoiesis. Blood 118: 3842–3852. | |
| 21 | Tg(ela3l:Cre,cryaa:Venus)s932 | Hesselson D, Anderson RM, Stainier DY (2011) Suppression of Ptf1a activity induces acinar-to-endocrine conversion. Curr Biol 21: 712–717. | |
| 22 | Tg(-3.5ubb:CreERT2, myl7:EGFP)cz1702 | Mosimann C, Kaufman CK, Li P, Pugach EK, Tamplin OJ, et al. (2011) Ubiquitous transgene expression and Cre-based recombination driven by the ubiquitin promoter in zebrafish. Development 138: 169–177. | |
| 23 | Tg(EPV.Tp1-Ocu.Hbb2:CreERT2)jh12 | Wang Y, Rovira M, Yusuff S, Parsons MJ (2011) Genetic inducible fate mapping in larval zebrafish reveals origins of adult insulin-producing beta-cells. Development 138: 609–617. | |
| 24 | Tg(actb2:GFP-CreERT2)jh29 | Wang Y, Rovira M, Yusuff S, Parsons MJ (2011) Genetic inducible fate mapping in larval zebrafish reveals origins of adult insulin-producing beta-cells. Development 138: 609–617. | |
| 25 | TgBAC(cryaa:EGFP,tcf21:CreERT2)pd42 | Kikuchi K, Gupta V, Wang J, Holdway JE, Wills AA, et al. (2011) tcf21+ epicardial cells adopt non-myocardial fates during zebrafish heart development and regeneration. Development 138: 2895–2902.] | |
| 26 | Tg(cryaa:DsRed,-5.1myl7:CreERT2)pd10 | Kikuchi K, Gupta V, Wang J, Holdway JE, Wills AA, et al. (2011) tcf21+ epicardial cells adopt non-myocardial fates during zebrafish heart development and regeneration. Development 138: 2895–2902. | |
| 27 | Tg(Ola.Sp7:CreERT2-2A-mCherry)tud8 | Knopf F, Hammond C, Chekuru A, Kurth T, Hans S, et al. (2011) Bone regenerates via dedifferentiation of osteoblasts in the zebrafish fin. Dev Cell 20: 713–724. | |
| 28 | Tg(hsp70l:mCherry,CreERT2)tud104 | Hans S, Freudenreich D, Geffarth M, Kaslin J, Machate A, et al. (2011) Generation of a non-leaky heat shock-inducible Cre line for conditional Cre/lox strategies in zebrafish. Dev Dyn 240: 108–115. | |
| 29 | Tg(hsp70l:mCherry,CreERT2)tud105 | Hans S, Freudenreich D, Geffarth M, Kaslin J, Machate A, et al. (2011) Generation of a non-leaky heat shock-inducible Cre line for conditional Cre/lox strategies in zebrafish. Dev Dyn 240: 108–115. | |
| 30 | Tg(her4.1:mCherry,CreERT2)tud106 | Kroehne V, Freudenreich D, Hans S, Kaslin J, Brand M (2011) Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors. Development 138: 4831–4841. | |
| 31 | TgBAC(dbx1b:Cre-mCherry)nns13a | [Satou C, Kimura Y, Higashijima S (2012) Generation of multiple classes of V0 neurons in zebrafish spinal cord: progenitor heterogeneity and temporal control of neuronal diversity. J Neurosci 32: 1771–1783. | |
| 32 | Tg(-2.8fabp10a:Cre,cryaa:Venus)s955 | Ni TT, Lu J, Zhu M, Maddison LA, Boyd KL, et al. (2012) Conditional control of gene function by an invertible gene trap in zebrafish. Proc Natl Acad Sci U S A. | |
| 33 | Tg(Mmu.Sox10-Mmu.Fos:Cre)zf384 | [Kague E, Gallagher M, Burke S, Parsons M, Franz-Odendaal T, et al. (2012) Skeletogenic fate of zebrafish cranial and trunk neural crest. PLoS One 7: e47394. | |
| 34 | Tg(-4.7sox10:Cre)ba73 | Rodrigues FS, Doughton G, Yang B, Kelsh RN (2012) A novel transgenic line using the Cre-lox system to allow permanent lineage-labeling of the zebrafish neural crest. Genesis 50: 750–757. | |
| 35 | Tg(-4.7sox10:Cre)ba74 | Hammond CL, Moro E (2012) Using transgenic reporters to visualize bone and cartilage signaling during development in vivo. Front Endocrinol (Lausanne) 3: 91. | |
| 36 | Tg(-4.7sox10:Cre)ba101 | Rodrigues FS, Doughton G, Yang B, Kelsh RN (2012) A novel transgenic line using the Cre-lox system to allow permanent lineage-labeling of the zebrafish neural crest. Genesis 50: 750–757. | |
| 37 | Tg(hsp70l.1:mcherry,CreERT2)jk67 | Yoshinari N, Ando K, Kudo A, Kinoshita M, Kawakami A (2012) Colored medaka and zebrafish: transgenics with ubiquitous and strong transgene expression driven by the medaka beta-actin promoter. Dev Growth Differ 54: 818–828. | |
| 38 | Tg(Ola.Sp7:TagBFP,CreERT2)pd45/+ | Singh SP, Holdway JE, Poss KD (2012) Regeneration of amputated zebrafish fin rays from de novo osteoblasts. Dev Cell 22: 879–886. | |
| 39 | Tg(dusp6:CreERT2,myl7:ECFP)b1230 | Stewart S, Stankunas K (2012) Limited dedifferentiation provides replacement tissue during zebrafish fin regeneration. Dev Biol 365: 339–349. | |
| 40 | TgBAC(-25ltbp3:TagRFP-Cre)fb1 | Guner-Ataman B, Paffett-Lugassy N, Adams MS, Nevis KR, Jahangiri L, et al. (2013) Zebrafish second heart field development relies on progenitor specification in anterior lateral plate mesoderm and nkx2.5 function. Development 140: 1353–1363. | |
| 41 | Tg(kop:Cre-UTRnanos3,CMV:EGFP)ihb7 | Xiong F, Wei ZQ, Zhu ZY, Sun YH (2013) Targeted expression in zebrafish primordial germ cells by Cre/loxP and Gal4/UAS systems. Mar Biotechnol (NY) 15: 526–539. | |
| 42 | TgBAC(nkx2.5:ERT2-CreERT2)fb8 | Guner-Ataman B, Paffett-Lugassy N, Adams MS, Nevis KR, Jahangiri L, et al. (2013) Zebrafish second heart field development relies on progenitor specification in anterior lateral plate mesoderm and nkx2.5 function. Development 140: 1353–1363. | |
| 43 | Tg(sox10:Cre,myl7:EGFP)sq5 | Ho Lee RT, Thiery JP, Carney TJ (2013) Dermal fin rays and scales derive from mesoderm, not neural crest. Curr Biol 23: R336-337. | |
| 44 | Tg(tbx6l:Cre,myl7:EGFP)sq6 | Ho Lee RT, Thiery JP, Carney TJ (2013) Dermal fin rays and scales derive from mesoderm, not neural crest. Curr Biol 23: R336-337. | |
| 45 | Tg(tbx6l:CreERT2,myl7:EGFP)sq7 | Ho Lee RT, Thiery JP, Carney TJ (2013) Dermal fin rays and scales derive from mesoderm, not neural crest. Curr Biol 23: R336-337. | |
| 46 | Tg(sox10:CreERT2,myl7:GFP)t007 | Mongera A, Singh AP, Levesque MP, Chen YY, Konstantinidis P, et al. (2013) Genetic lineage labeling in zebrafish uncovers novel neural crest contributions to the head, including gill pillar cells. Development 140: 916–925. | |
| 47 | Tg2(hsp70l:Cre)a134 | Pan YA, Freundlich T, Weissman TA, Schoppik D, Wang XC, et al. (2013) Zebrabow: multispectral cell labeling for cell tracing and lineage analysis in zebrafish. Development 140: 2835–2846. | |
| 48 | Tg(amhc:CreERT2)sd20 | Zhang R, Han P, Yang H, Ouyang K, Lee D, et al. (2013) In vivo cardiac reprogramming contributes to zebrafish heart regeneration. Nature 498: 497–501. | |
| 49 | Tg(pax2a:CreERT2)tud110 | Hans S, Irmscher A, Brand M (2013) Zebrafish Foxi1 provides a neuronal ground state during inner ear induction preceding the Dlx3b/4b-regulated sensory lineage. Development 140: 1936–1945. | |
| 50 | Tg(LOXP-CMV:Cre,CMV:EGFP) | Lin HJ, Lee SH, Wu JL, Duann YF, Chen JY (2013) Development of Cre-loxP technology in zebrafish to study the regulation of fish reproduction. Fish Physiol Biochem 39: 1525–1539. | |
| 51 | TgBAC(gsx1:Cre) | Satou C, Kimura Y, Hirata H, Suster ML, Kawakami K, et al. (2013) Transgenic tools to characterize neuronal properties of discrete populations of zebrafish neurons. Development 140: 3927–3931. | |
| 52 | Tg(myl7:YFP-Cre) | Ding Y, Liu W, Deng Y, Jomok B, Yang J, et al. (2013) Trapping cardiac recessive mutants via expression-based insertional mutagenesis screening. Circ Res 112: 606–617. | |
| 53 | Tg(fabp10a:CreERT2) | Choi TY, Ninov N, Stainier DY, Shin D (2013) Extensive conversion of hepatic biliary epithelial cells to hepatocytes after near total loss of hepatocytes in zebrafish. Gastroenterology 146: 776–788. | |
| 54 | Tg(TP1:CreRT2) | Ninov N, Hesselson D, Gut P, Zhou A, Fidelin K, et al. (2013) Metabolic regulation of cellular plasticity in the pancreas. Curr Biol 23: 1242–1250. | |
| 55 | Tg(krt4:CreERT2,myl7:EGFP) | Lee RT, Asharani PV, Carney TJ (2014) Basal keratinocytes contribute to all strata of the adult zebrafish epidermis. PLoS One 9: e84858. | |
| 56 | Tg(krtt1c19e:CreERT2,myl7:EGFP) | Lee RT, Asharani PV, Carney TJ (2014) Basal keratinocytes contribute to all strata of the adult zebrafish epidermis. PLoS One 9: e84858. | |
| 57 | Tg(krtt1c19e:CreERT2) | Fischer B, Metzger M, Richardson R, Knyphausen P, Ramezani T, et al. (2014) p53 and TAp63 promote keratinocyte proliferation and differentiation in breeding tubercles of the zebrafish. PLoS Genet 10: e1004048. | |
| 58 | Tg(krt4:CreERT2) | Fischer B, Metzger M, Richardson R, Knyphausen P, Ramezani T, et al. (2014) p53 and TAp63 promote keratinocyte proliferation and differentiation in breeding tubercles of the zebrafish. PLoS Genet 10: e1004048. | |
| 59 | Tg(kdrl:Cre-ERT2) | Zhao L, Borikova AL, Ben-Yair R, Guner-Ataman B, MacRae CA, et al. (2014) Notch signaling regulates cardiomyocyte proliferation during zebrafish heart regeneration. Proc Natl Acad Sci U S A 111: 1403–1408. |
Cre- and CreERT2-driver lines available in zebrafish are listed in chronological order including citation.
To expand the existing pool of CreERT2-driver lines in zebrafish, we performed a genome-wide gene trap screen, using a vector that reports gene expression via the fluorescence protein mCherry and simultaneously drives CreERT2 under the endogenous promoter. To this end, we applied the mCherry-T2a-CreERT2 (mCT2aC) gene trap construct consisting of a splice acceptor (SA) and a single open reading frame coding for mCherry and CreERT2, separated by the viral T2a peptide sequence (Fig 1A) [21, 24, 49]. In total, we used three different mCT2aC gene trap vectors, yielding 27 novel CreERT2-driver lines expressing in various tissues in the developing and adult zebrafish. All lines were analyzed with respect to integration, expression, functionality and potential phenotype caused by the insertion.
Fig 1. Generation of CreERT2-driver lines via gene trapping.
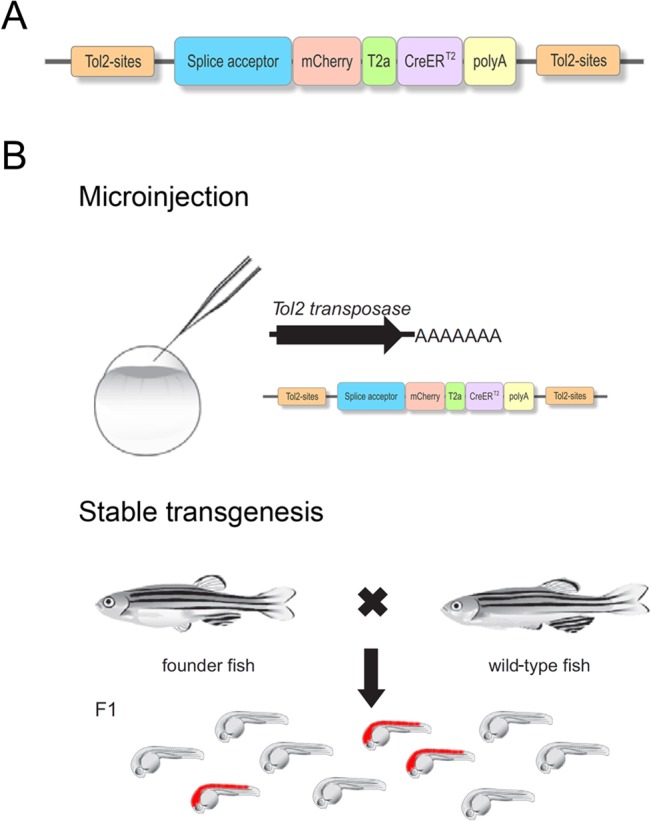
(A) Tol2-based mCT2aC gene trap vector comprising a splice acceptor, the mCherry sequence separated from CreERT2 via T2a. (B) Work schedule for the generation of CreERT2-driver lines using a gene trap approach.
Material and Methods
Cloning of the pTol-SA x -mCT2aC gene trap cassettes
To generate the rabbit β-globin SA containing plasmid pTol-SA1-mCT2aC, the mCherry-T2a-CreERT2 cassette (mCT2aC) was cloned into the Tol2 transposon-based gene trap vector pT2KSAG plasmid [5]. To create the pTol-SA2-mCT2aC plasmid the zebrafish bcl2 SA was amplified from the T2BGS plasmid [51] using the following primers flanked by the indicated restriction sites: Bcl2-for (Apa1) atatGGGCCCtagcagtttcatgcaccatagaccgc; Egfp r4-rev (Fse1) atatGGCCGGCCgatgggcaccaccccggtga that allowed substitution of the SA1 of the pTol-SA1-mCT2aC plasmid. Similarly, to generate the pTol-SA3-mCT2aC plasmid the zebrafish gata6 SA was amplified from 24 hpf wild-type AB cDNA using the following primers flanked by the indicated restriction sites: GATA6-for (Apa1) atatGGGCCCtataagtagactgttaggttggggttaggat; GATA6-5’-rev (Fse1) atatGGCCGGCCcctggatcagagcagagaatgtccgtg that allowed substitution of the SA1 of the pTol-SA1-mCT2aC plasmid.
Zebrafish husbandry, germ line transformation and screening of F1 progeny
Zebrafish embryos were obtained by natural spawnings of adult wild-type AB fish maintained at 28.5°C on a 14-hr light, 10-hr dark cycle and staged as described [52, 53]. For germ line transformation, 30 pg plasmid DNA and 30 pg transposase mRNA were injected into fertilized eggs (F0), raised to adulthood and crossed to wild-type AB fish as previously described [5]. To identify transgenic carriers, F1 embryos were screened for mCherry under a fluorescent microscope (Olympus MVX10) at various developmental stages (1–5 dpf). mCherry positive embryos were raised and re-identified in the F2 generation.
Insertion mapping using 5’RACE and inverse PCR (iPCR)
Mapping of insertions was done by 5’RACE on the cDNA level. RNA was isolated from 24 to 48 hpf mCherry positive 10–15 embryos using Trizol (Ambion, Life Technologies) according to the manufacturer’s protocol. 5’RACE was performed according to the manufacturer’s protocol of the SMARTer RACE cDNA Amplification Kit (Clontech) with the following primers: (mcherry rev 5’- AGTTCATCACGCGCTCCCACTTGAAGCC and mcherry rev 2 5’- CGTAGGCCTTGGAGCCGTAC (as nested primer)). Mapping of gene trap insertions on DNA level was done by inverse PCR as previously published [54] with modification of primers (1st PCR: Tol for1 3‘ TTTACTCAAGTAAGATTCTAG; Tol rev1 3‘ CTCCATTAAAATTGTACTTG; Tol for1 5‘ CTTGAGTACAATTAAAAATCAATAC; Tol rev1 5‘ GTAAAAATCCCCAAAAATAATAC; 2nd PCR: Tol for2 3‘ ACTTGTACTTTCACTTGAGTA; Tol rev2 3‘ GCAAGAAAGAAAACTAGAGA; Tol for2 5‘ CTCCTTACAATTTTATTTACAGTC; Tol rev2 5‘ GTAAAATTACTCAAGTACTTTACACC (communication with J.Bessa).
Expression analysis of transgenic lines
Expression patterns of respective CreERT2-driver lines were analyzed using native mCherry fluorescence as well as in situ hybridization (ISH) analysis for CreERT2. Probe synthesis and ISH was performed essentially as previously described [55, 56] using the vector pCs2+-CreERT2 [22]. Native mCherry fluorescence and stainings were analyzed using a Zeiss Axiophot 2 or an Olympus MVX10 microscope.
Pharmacological treatments and functionality assay
For Tamoxifen (TAM) and 4-hydroxy-Tamoxifen (4-OHT) (Sigma, St. Louis, MO;T5648 and H7904) treatments, a 50 mM and 25 mM stock solution was made in DMSO and ethanol and stored at -20°C. To test the functionality of the respective CreERT2-driver lines, the individual CreERT2-driver line was crossed with the Cre-dependent reporter line Tg(hsp70l:loxP-DsRed-loxP-EGFP) which expresses DsRed2 under the control of the ubiquitous, temperature inducible hsp70l promoter, but switches permanently to EGFP after a successful recombination event [24]. For embryonic treatment, progeny of this cross were exposed to 5 μM TAM from 6 hpf to 24 hpf to elicit recombination, heat shocked at 24 hpf for 1 hour to activate reporter expression and analyzed at 28 hpf. CreERT2-driver lines with an onset of CreERT2 beyond 24 hpf were exposed to 5 μM TAM from 36 hpf to 48 hpf, heat shocked at 48 hpf and analyzed at 52 hpf. For larval treatment, progeny of this cross were exposed to 1 μM 4-OHT from 80 hpf to 96 hpf to elicit recombination, heat shocked at 96 hpf for 1 hour to activate reporter expression and analyzed at 100 hpf.
Nomenclature guidelines
CreERT2-driver lines are designated based on ZFIN nomenclature. Full names are presented as e.g. Gt(SA3-mCT2aC)tud37, which reflects the utilized splice acceptor (SA3), the mCherry-T2a-CreERT2-cassette (mCT2aC) and the individual line designation number (e.g. 37) with respect to our institute (tud). To increase readability CreERT2-driver lines are abbreviated throughout the manuscript (e.g. CreERT2-driver line Gt(SA3-mCT2aC)tud37: tud37Gt).
Ethics statement
All experiments were conducted in accordance with the Animal Welfare Act and with permission of the federal authorities (Landesdirektion Sachsen AZ 24–9168.11-1/2013-14, Germany). Moreover, according to the EU Directive 2010/63/EU on the protection of animals used for scientific purposes, early life-stages of zebrafish are not protected as animals until the stage of being capable of independent feeding (5 days post fertilization). In this study the experiments did not exceed an exposure time of 4 days post fertilization, thus, the zebrafish utilized were not capable of independent feeding and not protected as animals according to the EU Directive mentioned above.
Results
The mCT2aC gene trap screen
In order to obtain a wide variety of CreERT2-driver lines a gene trap approach was chosen using a vector containing a splice acceptor (SA) and a mCherry-tagged variant of CreERT2 (consisting of a single open reading frame coding for mCherry and CreERT2 separated by the viral T2A peptide sequence) followed by a polyadenylation (p(A)) signal (Fig 1A). To avoid any SA site-specific integration bias three different trapping vectors were generated containing different SA sites [22] [49] (pTol-SA1-mCT2aC: rabbit β-globin SA; pTol-SA2-mCT2aC: zebrafish bcl2 SA; pTol-SA3-mCT2aC: zebrafish gata6 SA). Upon random integration of the gene trap vector into an endogenous locus, a fusion between the 5’-located exons and the gene trap cassette is generated. The p(A) signal within the gene trap cassette mediates transcriptional termination, resulting in a truncation of the endogenous gene sequence. The separation of the bicistronic message using T2a allows equimolar production of both mCherry and CreERT2. Depending on the insertion site, information on the subcellular localization of the trapped protein is provided by the mCherry tag. To generate mCherry-tagged CreERT2-driver lines, we injected one-cell-stage wild-type embryos with Tol2 transposase mRNA together with the respective gene trap construct. Injected fish were raised and outcrossed to wild-type fish as previously described [5]. The resulting F1 embryos were examined under a fluorescent microscope at various developmental stages (1–5 dpf) and mCherry positive embryos were selected and raised (Fig 1B). Co-transmission of gene trap integrations was resolved in subsequent generations and only carriers of single insertion events were raised further. In total, 1479 fish were screened for the different gene trap constructs (pTol-SA1-mCT2aC: 1034 fish; pTol-SA2-mCT2aC: 177 fish; pTol-SA3-mCT2aC: 268 fish) yielding trapping rates of 8,7%, 14,1% and 11,6% respectively, which is consistent with previous data [5, 35, 57]. Out of 148 insertions, 42 were selected and established as stable transgenic lines and further analyzed including a recombination functionality assay, embryonic expression profiling using CreERT2 ISH and native mCherry fluorescence, transgene mapping and investigation of integration-related phenotypes. In all 42 transgenic lines, inheritance rates of approximately 50% were consistent with the ratio of Mendelian segregation and observed through additional generations indicating integration into only one active locus per line. Southern blot analysis to determine multi-copy integration at the active locus or the overall copy number integrated in the genomes was not performed. A variety of mCherry expression patterns were observed in transgenic F1 embryos, indicating that the mCT2aC cassette was inserted into various loci in the genome and is expressed under the control of various endogenous promoters. In addition to ubiquitous expression (Fig 2A–2C) native mCherry expression could be detected e.g. in the anlagen of the neural tube (Fig 2D–2G), somites (Fig 2H–2K), inner ear (Fig 2L and 2M), heart (Fig 2N and 2O), tail bud (Fig 2P), fin bud (Fig 2Q) or kidney (Fig 2R).
Fig 2. Native mCherry expression of CreERT2-driver lines in a variety of embryonic tissues.

(A-C) Ubiquitous mCherry expression in tud20Gt at 24 and 48 hpf, and tud38Gt at 48 hpf. (D-G) CreERT2-driver lines expressing mCherry in restricted patterns of the neural tube: (D) hindbrain and spinal cord expression in tud17Gt at 48 hpf, (E-G) fore-, mid- and hindbrain expression in tud19Gt and tud27Gt at 48 hpf and fore- and midbrain expression in tud37Gt at 24 hpf. (H-K) Somitic mCherry expression in tud11Gt and tud13Gt at 48 hpf as well as in tud14Gt and tud38Gt at 24 hpf. (L, M) mCherry expression in the developing inner ear in tud37Gt at 48 hpf and tud38Gt at 24 hpf. (N, O) CreERT2-driver lines expressing mCherry in the embryonic heart in tud11Gt and tud14Gt at 48 hpf. (P) mCherry expression in the tail bud in tud27Gt at 24 hpf. (Q) Fin bud expression in tud28Gt at 48 hpf. (R) CreERT2-driver line tud28Gt shows mCherry expression in the kidney anlagen at 24 hpf; Bold letters indicate CreERT2-driver lines with known gene trap integrations.
Taken together, we find that gene trapping provides a rapid and unbiased method to create tissue-specific CreERT2-driver lines expressing in multiple tissues of the developing zebrafish.
Functional analysis of mCT2aC gene trap lines
To demonstrate the functionality of each CreERT2-driver line, we applied a standardized functionality assay [49]. In this assay the respective CreERT2-driver line (e.g. tud28Gt) was crossed to the Cre-dependent reporter line Tg(hsp70l:loxP-DsRed-loxP-EGFP) which expresses DsRed2 under the control of the ubiquitous, temperature inducible hsp70l promoter, but switches permanently to EGFP after a successful recombination event [24]. For embryonic treatment, progeny of this cross were exposed to 5 μM TAM from 6 hpf to 24 hpf to elicit recombination, heat shocked at 24 hpf for 1 hour to activate reporter expression and analyzed at 28 hpf (Fig 3A). Analysis of the selected 42 CreERT2-driver lines using this functionality assay showed successful recombination in 27 CreERT2-driver lines, or 64% of the gene trap lines (Fig 3B). In contrast, 15 CreERT2-driver lines, or 36% of the tested lines, did not show any successful recombination indicated by lack of green fluorescence (data not shown). In most cases, positive recombination (shown as native EGFP fluorescence) recapitulated the endogenous expression pattern detected by the CreERT2 ISH signal. For example, tud11Gt shows somitic CreERT2 expression at 24 hpf and native EGFP fluorescence is present in the same tissue at 28 hpf. Other examples are represented by CreERT2-driver lines tud17Gt or tud19Gt, where both endogenous CreERT2 expression and native EGFP fluorescence was observed in the spinal cord or telencephalon, respectively. However, the functionality assay also revealed positive recombination events in tissues devoid of CreERT2-expression at the selected time point, which most likely represent lineage tracings of cells expressing CreERT2 at earlier stages of development. For example, tud21Gt and tud23Gt show CreERT2 expression at 24 hpf only in the brain, whereas native EGFP fluorescence can be observed in the entire neural tube at 28 hpf. Similarly, CreERT2 expression in tud32Gt can only be detected in the developing brain at 24 hpf but native EGFP fluorescence is present in the entire neural tube as well as the somites.
Fig 3. Identification of functional CreERT2-driver lines.

(A) Scheme of the embryonic functionality assay. (B) 27 CreERT2-driver lines are shown with respect to CreERT2 expression at 24 hpf (CreERT2) and the respective embryonic functionality assay indicated by native EGFP fluorescence.
In order to test recombination at larval stages we randomly selected 5 CreERT2-driver lines (tud12Gt, tud29Gt, tud31Gt, tud37Gt, tud38Gt) and repeated the cross to the before mentioned Cre-dependent reporter line. Because treatment with 5 μM TAM from 80 hpf to 96 hpf followed by heat shock resulted in severe developmental abnormalities, we applied 1 μM 4-hydroxy-Tamoxifen (4-OHT) prior to heat treatment which did not cause any change in proper development. Analysis of tud12Gt and tud31Gt did not result in any successful recombination under these conditions (data not shown). In contrast, successful recombination reported by robust and strong EGFP fluorescence was observed in tud29Gt, tud37Gt and tud38Gt recapitulating the endogenous expression pattern detected by CreERT2 ISH (Fig 4). Furthermore, no EGFP expression could be detected in the absence of 4-OHT indicating tight regulation of CreERT2 in the respective CreERT2-driver lines and absence of any non-conditional recombination (leakiness).Taken together, our analysis revealed that the new gene trap lines are functional CreERT2-driver lines allowing for CreERT2-mediated transactivation in various tissues at various developmental stages. However, as it has been previously shown that recombination depends on the expression strength of CreERT2 [22] TAM or 4-OHT conditions need to be tested and optimized for each CreERT2 driver line.
Fig 4. Functionality of CreERT2-driver lines at larval stages.
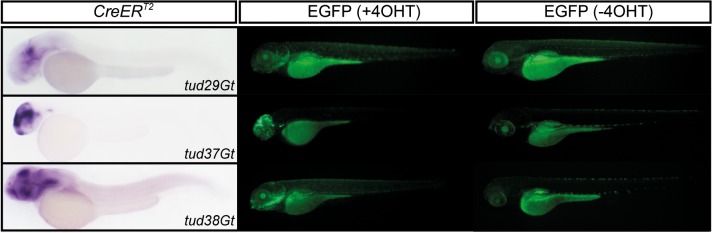
tud29Gt, tud37Gt and tud38Gt are shown with respect to CreERT2 expression at 48 hpf (CreERT2) and EGFP fluorescence in the presence and absence of 4-hydroxy-Tamoxifen (4-OHT) at 100 hpf.
Molecular characterization of CreERT2-driver lines
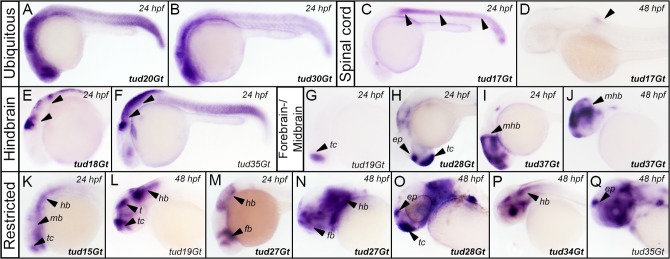
In order to map the gene trapping events, 5’RACE of mCherry positive embryos was performed. If insertions could not be identified with 5’RACE, inverse PCR was carried out, which has been successfully applied previously [33]. In total 17 (63%) out of 27 functional CreERT2-driver lines could be mapped. 11 traps (65%) were identified using 5’RACE and 6 traps (35%) by inverse PCR. The remaining 10 integrations (27%) were inconclusive, consistent with observations by Kawakami and colleagues, who reported that only 50–70% of Tol2-integrations can be mapped using 5’RACE [5]. Molecularly identified CreERT2-driver lines are summarized in Table 2, including gene trap integration, NCBI gene ID, linkage group as well as insertion site. Transgene mapping revealed vector integration into genes involved in a variety of biological processes and protein classes, such as cytoskeletal proteins (1 gene), extracellular matrix proteins (1 gene), signaling receptors (1 gene), transcription factors (5 genes) or nucleic acid binding proteins (6 genes) (source PANTHER database http://www.pantherdb.org [58]). Consistent with previous reports, no integration bias into any chromosomal location could be detected for the Tol2 transposable system [59, 60]. However, most of the gene trap integrations are biased towards the 5’ end of genes. 12 of the mapped insertions integrated into the 5’UTR or exon/intron 1 of the trapped gene. In tud25Gt, tud27Gt and tud38Gt the gene trap cassette was inserted into intron 2 or intron 4 of genes with 19 exons, respectively. Only in tud34Gt and tud37Gt vector integration was observed into the 3’ end of the genes.
Table 2. Molecular identification of functional CreERT2-driver lines.
| CreERT2-driver line | Gene trap integration | NCBI Gene ID | Linkage group | Insertion site |
|---|---|---|---|---|
| Gt(SA1-mCherry-T2A-CreERT2)tud11Gt | ptk2.2 | 386705 | 19 | 5'UTR |
| Gt(SA1-mCherry-T2A-CreERT2)tud12Gt | mapre1a | 334135 | 8 | 5'UTR |
| Gt(SA1-mCherry-T2A-CreERT2)tud13Gt | pvalb1 | 402805 | 3 | 5'UTR |
| Gt(SA1-mCherry-T2A-CreERT2)tud15Gt | klhl17 | 336187 | 23 | Intron 1 |
| Gt(SA1-mCherry-T2A-CreERT2)tud16Gt | LOC100002938 | 100002938 | 12 | Intron 1 |
| Gt(SA1-mCherry-T2A-CreERT2)tud17Gt | hoxb1b | 30374 | 2 | 5'UTR |
| Gt(SA1-mCherry-T2A-CreERT2)tud18Gt | msxc | 30526 | 13 | Exon 1 |
| Gt(SA1-mCherry-T2A-CreERT2)tud20Gt | si:ch73-248e17.1 | Not identified | 22 | 5'UTR |
| Gt(SA1-mCherry-T2A-CreERT2)tud21Gt | baz2ba | 561095 | 6 | Intron 1 |
| Gt(SA1-mCherry-T2A-CreERT2)tud25Gt | vldlr | 393897 | 10 | Intron 2 |
| Gt(SA2-mCherry-T2A-CreERT2)tud27Gt | epha7 | 562195 | 20 | Intron 4 |
| Gt(SA2-mCherry-T2A-CreERT2)tud28Gt | si:ch211-263k4.2 | 796720 | 8 | Intron 1 |
| Gt(SA2-mCherry-T2A-CreERT2)tud29Gt | ccdc102a | 325319 | 7 | Intron 1 |
| Gt(SA3-mCherry-T2A-CreERT2)tud30Gt | srsf1b | 393565 | 21 | Intron 1 |
| Gt(SA3-mCherry-T2A-CreERT2)tud34Gt | ywhaba | 323055 | 6 | 3'UTR |
| Gt(SA3-mCherry-T2A-CreERT2)tud37Gt | otx1b | 30500 | 17 | Exon 4 |
| Gt(SA3-mCherry-T2A-CreERT2)tud38Gt | sox6 | 567154 | 4 | Intron 4 |
Full names of functional CreERT2-driver lines are designated as transgenic lines according to ZFIN nomenclature. Information of the gene trap integrations are shown with gene names, NCBI gene ID, chromosomal insertion (linkage group) and the insertion site relative to the gene architecture.
Expression profile of tissue-specific CreERT2-driver lines
In order to determine the embryonic CreERT2 expression profile in more detail CreERT2 in situ hybridization (ISH) at 24 and 48 hpf was performed. This analysis corroborated that CreERT2 is expressed under the control of various endogenous promoters in various tissues, which were categorized using the “Phenotype Attribute and Trait Ontology” (PATO)-compliant terms to describe expression patterns with respect to anatomic regions [61], such as neural tube (Fig 5), eye (Fig 6), somites (Fig 7), fin bud, inner ear, ubiquitous, heart, blood island, reproductive system or kidney (Fig 8). To analyze whether gene trap integrations indeed resemble the endogenous gene transcripts, ISH was performed for both CreERT2 and respective endogenous genes. For example the insertion of the mCT2aC cassette into the orthodenticle homolog 1b (otx1b) locus in tud37Gt shows that CreERT2 expression faithfully recapitulates the endogenous otx1b expression. Expression of both CreERT2 and otx1b in tud37Gt transgenics or otx1b in wild-type siblings is restricted to the fore- and midbrain, as well as the developing inner ear at 24 and 48 hpf (S1 Fig). Another example is the insertion of the mCT2aC cassette into the kelch-like 17 (klhl17) locus in tud15Gt. Expression was observed in specific regions of the neural tube for CreERT2 transcripts of tud15Gt and klhl17 transcripts in wild-type siblings (data not shown). Thus, we conclude that pTol-SA x -mCT2aC gene trap integrations are able to report the endogenous expression pattern of trapped genes.
Fig 5. CreERT2-driver lines expressing in the embryonic neural tube.
(A, B) Pan-neural expression of CreERT2 revealed by in situ hybridization in tud20Gt and tud30Gt at 24 hpf. (C,D) Expression of CreERT2 in the spinal cord in tud17Gt at 24 and 48 hpf. (E,F) Hindbrain expression of CreERT2 in tud18Gt and tud35Gt at 24 hpf. (G-J) Fore-/Midbrain expression and (K-Q) other restricted patterns of CreERT2 in tud15Gt, tud19Gt, tud27Gt, tud28Gt, tud34Gt, tud35Gt and tud37Gt at 24 and 48 hpf, respectively. Bold letters indicate CreERT2-driver lines with known gene trap integrations. (See text for detailed description of expression patterns.); ep: epiphysis; fb: forebrain; hb: hindbrain; mb: midbrain; mhb: mid-hindbrain-boundary; t: tectum; tc: telencephalon.
Fig 6. CreERT2-driver lines expressing in the embryonic eye.
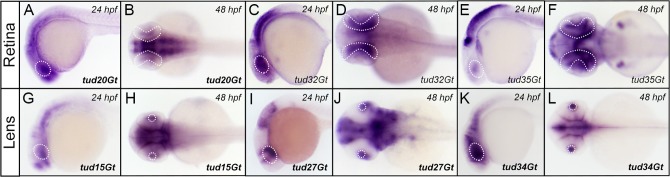
(A-F) Expression of CreERT2 in the developing retina in tud20Gt, tud32Gt and tud35Gt at 24 and 48 hpf, respectively. (G-L) CreERT2 is expressed in the developing lens in tud15Gt, tud27Gt and tud34Gt at 24 and 48 hpf, respectively. White dotted circles mark the retina (A-F) and lens (G-H). Bold letters indicate CreERT2-driver lines with known gene trap integrations. (See text for detailed description of expression patterns.)
Fig 7. CreERT2-driver lines expressing in the somites.
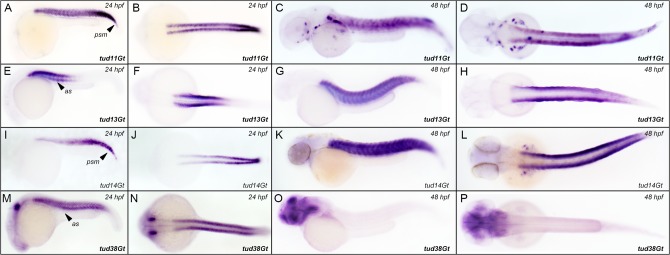
Expression of CreERT2 in (A-D) tud11Gt, (E-H) tud13Gt, (I-L) tud14Gt and (M-P) tud38Gt at 24 and 48 hpf.; Bold letters indicate CreERT2-driver lines with known gene trap integration. (See text for detailed description of expression patterns.) as: anterior somites; psm: presomitic mesoderm.
Fig 8. CreERT2-driver lines expressing in various embryonic tissues.

(A) Ubiquitous expression of CreERT2 in tud20Gt at 24 hpf. (B-E) CreERT2 is expressed in the developing heart in tud35Gt and tud36Gt at 24 and 48 hpf. (F-P) Expression of CreERT2 can be detected in the anlagen of (F-I) the inner ear in tud37Gt and tud38Gt (J) blood island in tud16Gt, (K,L) kidney in tud28Gt (M-O) fin buds in tud28Gt, tud29Gt and tud35Gt and (P) the reproductive system in tud29Gt at 24 and 48 hpf. Bold letters indicate CreERT2-driver lines with known gene trap integrations. (See text for description of expression patterns.)
CreERT2-driver lines expressing in the developing neural tube
24 functional CreERT2-driver lines show expression in the developing neural tube. Examples are depicted in Fig 5 with respect to different categories including ubiquitous expression in the neural tube (Fig 5A and 5B), spinal cord (Fig 5C and 5D), but also expression patterns restricted to the hindbrain (Fig 5E and 5F), fore- and midbrain (Fig 5G–5J), as well as other restricted neuronal patterns (Fig 5K–5Q). Broad expression in the neural tube is exemplarily shown in tud20Gt and tud30Gt at 24 hpf (Fig 5A and 5B) where integration of the mCherry-T2a-CreERT2 (mCT2aC) cassette occurred into the uncharacterized si:ch73-248e17.1 locus and the serine/arginine-rich splicing factor 1b (srsf1b) gene, respectively. Expression of CreERT2 in the entire spinal cord was detected in tud17Gt at 24 hpf (Fig 5C), which becomes restricted to the anterior spinal cord at 48 hpf (Fig 5D). Mapping of this insertion revealed integration of the mCT2aC cassette into the homeobox B1b (hoxb1b) locus, which was recently reported to control cell division, cell shape and microtubule dynamics during neural tube morphogenesis in zebrafish [62]. CreERT2-driver line tud18Gt shows expression in the hindbrain region at 24 hpf and was mapped to the muscle segment homeobox C (msxc) locus which has been previously described to be expressed in the hindbrain of early embryos [63]. Strong hindbrain expression was also detected in tud35Gt at 24 hpf (Fig 5F), which disperses into the fore- and midbrain at later stages (Fig 5Q). Unfortunately, integration mapping of this line was inconclusive. Restricted telencephalic expression was detected in tud19Gt in 24 hpf old embryos (Fig 5G), which expands into the mid- and hindbrain at 48 hpf (Fig 5L). Strong expression of CreERT2 was observed in the epiphyseal region as well as in the telencephalon in tud28Gt at 24 hpf with weaker expression in the hindbrain (Fig 5H). At 48 hpf, strong expression in the forebrain and epiphysis is maintained but hindbrain expression has significantly increased (Fig 5O). Insertion mapping revealed integration into the si:ch211-263k4.2 (novel protein similar to H.sapiens PRDM16, PR domain containing 16) locus. Another interesting CreERT2-driver line is represented by tud37Gt, where gene trapping has occurred into the orthodenticle homolog 1b (otx1b) locus. Expression is restricted to the fore- and midbrain abutting the mid-hindbrain-boundary (mhb) (Fig 5I and 5J). This pattern is consistent with previous reports detecting otx1b transcripts at high levels in a triangular patch already at mid-gastrula stage, which gives rise to fore- and midbrain structures [64]. Other restricted expression patterns include tud15Gt (Fig 5K), where gene trapping has occurred into the kelch-like family member 17 (klhl17) gene, which is also known as actinfilin. Interestingly, expression analysis in rat brain indicated actinfilin to be expressed in neurons of most brain regions [65]. In addition, tud27Gt displays expression in the fore- and hindbrain, excluding the midbrain region at 24 and 48 hpf (Fig 5M and 5N). Molecular mapping revealed integration of the mCT2aC cassette into the eph receptor A7 (epha7) locus. In tud34Gt CreERT2 expression is present in the entire brain, excluding the telencephalon and tectum at 48 hpf (Fig 5P). Molecular mapping revealed integration into tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, beta polypeptide a (ywhaba).
CreERT2-driver lines expressing in the developing eye
16 functional CreERT2-driver lines show expression in the embryonic eye, including the developing retina (Fig 6A–6F) and lens (Fig 6G–6L). Retinal expression is exemplarily shown in transgenic lines tud20Gt (Fig 6A and 6B), tud32Gt (Fig 6C and 6D) and tud35Gt (Fig 6E and 6F). CreERT2 is expressed broadly in the optic cup at 24 hpf in tud20Gt (Fig 6A) and tud32Gt (Fig 6C), whereas CreERT2 expression is restricted to a subdomain in tud35Gt (Fig 6E). At 48 hpf, CreERT2 transcripts are detected in a small region of the retina in tud20Gt (Fig 6B). Broader retinal expression is observed in tud32Gt (Fig 6D) and tud35Gt (Fig 6F). Mapping of these lines revealed either integration into an uncharacterized gene locus (tud20Gt) or remained inconclusive (tud32Gt, tud35Gt). In addition, we observed CreERT2-driver lines expressing in the embryonic lens, which develops at 30 to 36 hpf. Fairly broad CreERT2 expression in the eye field at 24 hpf was detected for all lines shown (Fig 6G, 6I and 6K). At 48 hpf expression was observed in both lens and retina for tud15Gt (Fig 6H). Interestingly, CreERT2 expression in tud27Gt at 48 hpf was restricted to the lens with adjacent expression in the temporal retina (Fig 6J). Restricted lens expression was also detected in tud34Gt at 48 hpf. Insertion mapping of these CreERT2-driver lines revealed integration into kelch-like 17 (klhl17) (tud15Gt), eph receptor A7 (epha7) (tud27Gt) and tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, beta polypeptide a (ywhaba) (tud34Gt).
CreERT2-driver lines expressing in the developing somites
We observed somitic CreERT2 expression in 9 functional CreERT2-driver lines (Fig 7). tud11Gt shows broad somitic CreERT2 expression at 24 and 48 hpf (Fig 7A–7D), including the presomitic mesoderm (PSM) at 24 hpf (Fig 7A). Gene trap integration occurred into protein tyrosine kinase 2aa (ptk2aa), a human ortholog of Focal adhesion kinase 1 (Fak1). Recent lineage tracing data using tud11Gt revealed that early ptk2aa expressing cells give rise to scale structures in zebrafish [50]. CreERT2 expression restricted to anterior somites at 24 hpf was observed in tud13Gt (Fig 7E and 7F), which expands into the posterior somites at 48 hpf (Fig 7G and 7H). Insertion mapping revealed integration into parvalbumin 1 (pvalb1), which is known to be expressed in zebrafish skeletal muscle [66]. In tud14Gt, CreERT2 expression gradually increases from anterior to posterior somites and the PSM (Fig 7I and 7J) at 24 hpf. In contrast, strong and robust somatic CreERT2 expression can be found at 48 hpf (Fig 7K and 7L). Gene mapping of tud14Gt, however, remained inconclusive. Interestingly, in tud38Gt CreERT2 somitic expression is detected only at 24 hpf but is completely absent at 48 hpf (Fig 7M–7P). Molecular mapping revealed integration of the mCT2aC cassette into SRY-box containing gene 6 (sox6), which plays, amongst others, important roles in zebrafish muscle fibre type specification and differentiation [67].
CreERT2-driver lines expressing in other embryonic tissues
CreERT2 expression was also detected in various other embryonic tissues such as the anlagen of the heart (Fig 8B–8E), inner ear (Fig 8F–8I), blood island (Fig 8J), kidney (Fig 8K and 8L), fin bud (Fig 8M–8O) or reproductive system (Fig 8P). Ubiquitous CreERT2 expression was observed in tud20Gt at 24 hpf (Fig 8A). We detected expression of CreERT2 in the embryonic heart in tud35Gt and tud36Gt at 24 as well as 48 hpf (Fig 8B–8E). Unfortunately, gene mapping of these CreERT2-driver lines remained inconclusive. Transgenic lines tud37Gt (Fig 8F and 8G) and tud38Gt (Fig 8H and 8I) showed transgene expression in the anlagen of the inner ear at 24 and 48 hpf. Whereas CreERT2 expression is restricted to ventral cells of the otic vesicle in tud37Gt, the transgene is expressed throughout the otic vesicle in tud38Gt. Gene trap integration in tud37Gt occurred into the orthodenticle homolog 1b (otx1b) locus, which is involved in normal development of the zebrafish inner ear [68]. Also in mouse expression of otx1 was detected in specific subdomains of the inner ear such as the lateral canal, the ampulla or the cochlea [69]. In tud38Gt the mCT2aC cassette was trapped into the sox6 locus, which also has been described in the otic vesicle during mouse inner ear development [70]. Integration of the mCT2aC cassette occurred into the cdc42 effector protein 4-like gene locus in tud16Gt. Interestingly, CreERT2 expression is detected in the embryonic blood island, also known as the caudal hematopoietic tissue at 24 hpf (Fig 8J). Expression of the mCT2aC cassette in the embryonic kidney was detected in tud28Gt at 24 and 48 hpf (Fig 8K and 8L). Additionally, we found six gene trap lines expressing CreERT2 in the fin bud (Fig 8M–8O). In tud29Gt (Fig 8N) and tud35Gt (Fig 8O) CreERT2 expression is observed in restricted domains of the developing fin bud at 48 hpf. In tud28Gt CreERT2 is expressed in a fairly broad expression domain of the fin bud at 48 hpf (Fig 8M). Gene trap integration in tud11Gt occurred into protein tyrosine kinase 2aa (ptk2aa). Molecular identification of tud29Gt revealed integration into coiled-coil domain containing 102A (ccdc102a). Interestingly, tud29Gt shows CreERT2 expression, amongst others, in the reproductive system.
Overall, to date 59 Cre/CreERT2-driver lines have been described (Table 1). Addition of 27 novel functional CreERT2-driver lines described here, increases the number of available lines significantly (> 30%), and hence these lines represent important new tools to study zebrafish development, homeostasis and regeneration.
Phenotype analysis
Because of their ability to interfere with gene function, gene trap approaches have also been previously applied for insertional mutagenesis [44, 71] [72–74]. In most cases, vector insertion results in null or hypomorphic mutant phenotypes when inserted into the 5’ regions of a gene. To test if any insertion resulted in a developmental phenotype, all functional CreERT2-driver lines were inbred to obtain homozygous individuals which could be identified due to stronger mCherry fluorescence and were examined for morphological phenotypes in the first 5 days after fertilization. Of all 27 tested integrations, only homozygous tud37Gt animals showed an apparent phenotype. tud37Gt was mapped to the otx1b locus which is located on linkage group 17. The locus comprises four exons coding for a homeobox domain transcription factor (Fig 9A) [75]. Integration of the gene trap cassette occurred into exon four, resulting in a truncated Otx1b-mCherry fusion protein lacking parts of the transcription factor domain. Homozygous animals of tud37Gt display a variable phenotype affecting the development of eye, inner ear, forebrain, midbrain and heart (Fig 9B). Additionally, homozygous animals exhibit a bent body axis [76]. Analysis of more than 40 intercrosses revealed that the observed abnormalities are variable in strength, indicating additional genetic factors. Interestingly, knockdown of otx1b using antisense-morpholinos is less severe than the mutant phenotype observed in homozygous tud37Gt embryos/larvae [76, 77]. However, because the insertion occurred in the coding region of the transcription factor domain, the phenotype might also result from a dominant negative form of Otx1b protein.
Fig 9. Gene trap insertion into the otx1b locus of tud37Gt.
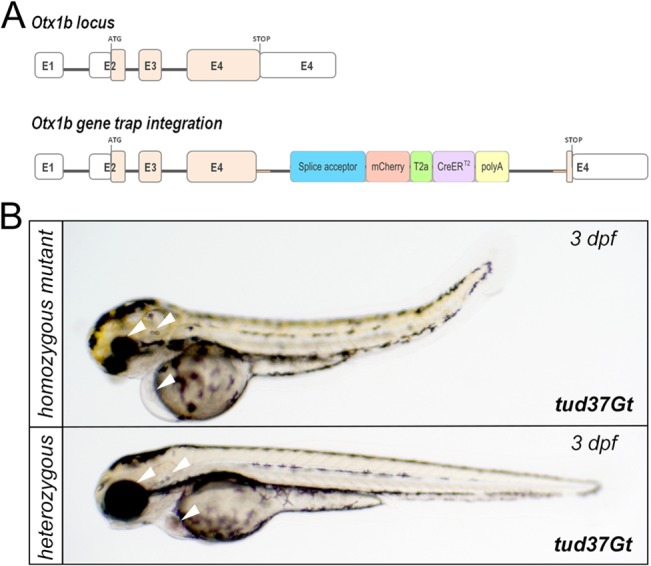
(A) Schematic drawing of the otx1b locus comprising of four exons (E1-E4) encoding a homeobox domain transcription factor. White boxes represent the 5’ and 3’ untranslated regions separated by the open reading frame in pink. The mCT2aC-cassette integrated into E4. (B) Bright field images of homozygous mutant tud37Gt embryos and heterozygous siblings. In comparison to heterozygous siblings, homozygous mutant tud37Gt embryos show defects in the developing eye, fore-/midbrain, ear and heart (white arrowheads) as well as a bend body shape.
Altogether, we conclude that integration of our pTol-SA x -mCT2aC gene trap constructs rarely interferes with endogenous gene function and results only rarely in overtly apparent mutant phenotypes. In agreement with these results, previous reports in zebrafish have shown that integrations of other gene trap vectors such as the T2KSAG vector did not result in any mutant phenotypes [5].
Discussion
Cre/loxP-technology has been successfully applied to dissect the zebrafish genome and genome-wide approaches have been conducted to create various Cre-effector lines [33–35]. However, currently the number of available cell- and tissue-specific Cre-driver lines to transactivate the before mentioned Cre-effector lines is limited [36]. By 2009, only 12 different Cre/CreERT2-driver lines had been published expressing Cre recombinase in a ubiquitous manner using the temperature inducible hsp70l promoter or tissue-specific promoter fragments driving expression in oocytes [78], pancreas [79], heart or neural tube [80]. In order to increase the existing pool of conditional Cre-driver lines we performed a genome-wide trapping screen using the mCherry-T2a-CreERT2 (mCT2aC) [21, 22] gene trap vector that yielded 27 new, fully functional CreERT2-driver lines expressing in various tissues in the developing zebrafish. The use of the mCT2aC cassette allows temporal control of Cre-mediated recombination using CreERT2. Initial conditional approaches in zebrafish have been carried out using Cre recombinase driven by the ubiquitous, temperature inducible hsp70l promoter [37, 81]. However, basal leakiness of the hsp70l promoter resulted in non-conditional Cre-mediated recombination even at permissive temperatures, limiting the usefulness of this approach [21, 22]. Non-conditional recombination has also been reported in cases when using CreERT2-constructs where high levels of CreERT2 might overwhelm the cellular machinery, preventing retention in the cytoplasm [24, 80]. To overcome this problem, Cre constructs fused with two LBD-domains have been generated [80, 82, 83]. However, although these constructs are more tightly regulated and display no non-conditional recombination, incomplete CreERT2-mediated recombination has been observed [80]. Therefore, Cre constructs with a single LBD-domain (CreERT2) currently provide the best option to achieve temporal control of Cre-mediated recombination. In addition to our gene trap lines, several promoter-fragment driven tissue-specific Cre/CreERT2-driver lines have been generated recently. To date, in total 63 Cre/CreERT2-driver lines have been described expressing either ubiquitously or in a tissue-specific manner [84] (data not shown). Thus, our new gene trap lines described here will increase this number by about 30% to 90 Cre/CreERT2-driver lines. The gene trapping approach using the pTol-SA x -mCT2aC constructs provides an efficient method for generating numerous CreERT2-driver lines expressing in various tissues. Whereas promoter fragments often do not faithfully recapitulate the endogenous expression pattern [22, 85], gene trapping enables transgene expression driven by the endogenous promoter. Gene trapping is also faster and less expensive compared to the generation of Cre/CreERT2-driver lines using BAC (bacterial artificial chromosome) transgenesis [86]. Another promising tool to create Cre/CreERT2-driver lines will be the use of sequence specific transcription activator-like effector nucleases (TALENs) and the clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) 9 system (RNA-guided nucleases, RGNs). These endonuclease systems enable the targeted insertion of open reading frames [87] or whole plasmid vectors [88] [89], thus allowing specific knock-ins into any desired gene locus in zebrafish.
Although our gene trap approach offers several advantages for creating new CreERT2-driver lines, we also observed some difficulties. According to our functionality assay, only 64% of the selected CreERT2-driver lines showed successful recombination, whereas 36% remained non-functional. In general, gene trapping results in fusion transcripts of the N-terminal endogenous exons followed by the open reading frame of the gene trap vector. Consequently, signal sequences encoded by the N-terminus are included into the fusion protein and result in sorting of the fusion protein to various intracellular compartments. The viral T2a peptide of the mCT2aC gene trap cassette mediates cleavage only after translation [90]. Hence, cleavage of CreERT2 from the mCherry tagged truncated protein occurs only after a potential trapping event into an intracellular compartment and translocation of CreERT2 into the nucleus after TAM application is prevented, rendering it non-functional. Alternatively, internal ribosomal entry site (IRES) sequences could be used to achieve bicistronic expression of mCherry and CreERT2. IRES sequences have been successfully applied in mouse and recently also in zebrafish [91, 92]. In contrast to T2a peptides, IRES sequences allow independent protein translation of a bicistronic mRNA containing two open reading frames. As a result, subcellular localization would only affect the first, upstream cistron (mCherry), but not the second (CreERT2). However, non-zebrafish derived IRES-based gene expression is notoriously non-stoichiometric, creating disproportionate transgene translation levels [93], whereas the viral T2A peptide sequence allows the production of mCherry and CreERT2 proteins in equimolar ratios [90]. Recent identification of IRES sequences derived from zebrafish might overcome this problem by generating equal amounts of gene product from both cistrons [94]. Indeed, successful application of a zebrafish IRES for the generation of CreERT2-driver expressing in the heart has been reported recently by Jopling and colleagues [83]. The identification of the insertion site using 5’RACE was not always successful, which might be explained by low amounts of fusion transcript or by integration far away from the 5’ end. However, Trinh le and colleagues reported transgene mapping efficiencies up to 92% using 5’RACE [35]. Still, other methods might prove useful to detect gene trap integrations. For example splinkerette PCR has been successfully applied to assess the genomic integration of the FlipTrap vector [35, 95]. Furthermore linker-mediated PCR (LM-PCR) has been used to detect gene trap integrations on genomic level [34].
Previously, gene trap vectors have been reported to create mutant phenotypes upon insertion, [44, 71] [33, 72–74]. Due to an internal p(A) signal of the gene trap cassette, transcription of the endogenous gene is terminated, which leads to a truncated transcript. Generation of mutation-linked CreERT2-driver lines might be disadvantageous when studying biological processes, e.g. when studying lineage relationships. However, of all tested integrations only homozygous tud37Gt animals with an integration into the otx1b locus showed an overt phenotype, consistent with previous results showing that integrations of the gene trap vector T2KSAG vector did not result in mutant phenotypes [5]. This observation could be explained by weakness of the SA that might lead to alternative splicing of the gene trap cassette and hence, allow low level production of endogenous full-length transcript. For example, the application of another SA derived from the first intron of the carp β-actin gene shows high mutagenic potential [33, 74]. However, homozygous CreERT2 integrations are not required because single alleles are sufficient to elicit efficient CreERT2-mediated recombination. Thus, transactivation studies using CreERT2 gene trap lines can, in most cases, be expected not to interfere with insertion-related phenotypes.
The analysis of all selected CreERT2-driver lines produced during this gene trap screen, but also of other promoter-fragment based CreERT2-driver lines generated in our lab, have been summarized in an online database, the zebrafish CreZoo (http://crezoo.crt-dresden.de) [49]. CreZoo lines can be requested from the European Zebrafish Resource Center in Karlsruhe (EZRC) (http://www.ezrc.kit.edu/).
Supporting Information
Comparison of the endogenous otx1b expression in wild-type embryos with the CreERT2 expression pattern in tud37Gt embryos which has been mapped to the otx1b locus at 24 and 48 hpf. fb: forebrain; mb: midbrain; ov: otic vesicle.
(TIF)
Acknowledgments
We thank members of the Brand lab for generous support and helpful discussions, Claudia Selck for help with the analysis of tud37Gt, and Victoria Splith for technical assistance. Furthermore, we thank Dr. Anming Meng for providing the T2BGS vector and José Bessa for help with the iPCR protocol.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This project is supported by the Deutsche Forschungsgemeinschaft (BR 1746/3-1 and SFB 655, project A3) and the European Union (ZF Health F-002339-535-000-2301201). In addition, Michael Brand is supported by funding of the Excellence Initiative by the German Federal and State Governments (Institutional Strategy, measure "support the best"). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Driever W, Solnica-Krezel L, Schier AF, Neuhauss SC, Malicki J, Stemple DL, et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development (Cambridge, England). 1996;123:37–46. . [DOI] [PubMed] [Google Scholar]
- 2. Haffter P, Granato M, Brand M, Mullins MC, Hammerschmidt M, Kane DA, et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development (Cambridge, England). 1996;123:1–36. . [DOI] [PubMed] [Google Scholar]
- 3. Amsterdam A, Burgess S, Golling G, Chen W, Sun Z, Townsend K, et al. A large-scale insertional mutagenesis screen in zebrafish. Genes & development. 1999;13(20):2713–24. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Wang D, Jao LE, Zheng N, Dolan K, Ivey J, Zonies S, et al. Efficient genome-wide mutagenesis of zebrafish genes by retroviral insertions. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(30):12428–33. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Kawakami K, Takeda H, Kawakami N, Kobayashi M, Matsuda N, Mishina M. A transposon-mediated gene trap approach identifies developmentally regulated genes in zebrafish. Developmental cell. 2004;7(1):133–44. . [DOI] [PubMed] [Google Scholar]
- 6. Parinov S, Kondrichin I, Korzh V, Emelyanov A. Tol2 transposon-mediated enhancer trap to identify developmentally regulated zebrafish genes in vivo. Dev Dyn. 2004;231(2):449–59. . [DOI] [PubMed] [Google Scholar]
- 7. Herbomel P, Levraud JP. Imaging early macrophage differentiation, migration, and behaviors in live zebrafish embryos. Methods in molecular medicine. 2005;105:199–214. . [DOI] [PubMed] [Google Scholar]
- 8. Weber M, Huisken J. Light sheet microscopy for real-time developmental biology. Current opinion in genetics & development. 2011;21(5):566–72. . [DOI] [PubMed] [Google Scholar]
- 9.Keller PJ. In vivo imaging of zebrafish embryogenesis. Methods (San Diego, Calif. 2013. . [DOI] [PMC free article] [PubMed]
- 10. Nasevicius A, Ekker SC. Effective targeted gene 'knockdown' in zebrafish. Nature genetics. 2000;26(2):216–20. . [DOI] [PubMed] [Google Scholar]
- 11. Wienholds E, van Eeden F, Kosters M, Mudde J, Plasterk RH, Cuppen E. Efficient target-selected mutagenesis in zebrafish. Genome research. 2003;13(12):2700–7. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Doyon Y, Vo TD, Mendel MC, Greenberg SG, Wang J, Xia DF, et al. Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures. Nature methods. 2008;8(1):74–9. . [DOI] [PubMed] [Google Scholar]
- 13. Huang P, Xiao A, Zhou M, Zhu Z, Lin S, Zhang B. Heritable gene targeting in zebrafish using customized TALENs. Nature biotechnology. 2011;29(8):699–700. 10.1038/nbt.1939 [DOI] [PubMed] [Google Scholar]
- 14. Hwang WY, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD, et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nature biotechnology. 2013;31(3):227–9. 10.1038/nbt.2501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Meng X, Noyes MB, Zhu LJ, Lawson ND, Wolfe SA. Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases. Nature biotechnology. 2008;26(6):695–701. 10.1038/nbt1398 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Sander JD, Cade L, Khayter C, Reyon D, Peterson RT, Joung JK, et al. Targeted gene disruption in somatic zebrafish cells using engineered TALENs. Nature biotechnology. 2011;29(8):697–8. 10.1038/nbt.1934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Branda CS, Dymecki SM. Talking about a revolution: The impact of site-specific recombinases on genetic analyses in mice. Developmental cell. 2004;6(1):7–28. . [DOI] [PubMed] [Google Scholar]
- 18. Glaser S, Anastassiadis K, Stewart AF. Current issues in mouse genome engineering. Nature genetics. 2005;37(11):1187–93. . [DOI] [PubMed] [Google Scholar]
- 19. Siegal ML, Hartl DL. Application of Cre/loxP in Drosophila. Site-specific recombination and transgene coplacement. Methods in molecular biology (Clifton, NJ. 2000;136:487–95. . [DOI] [PubMed] [Google Scholar]
- 20. Bischof J, Basler K. Recombinases and their use in gene activation, gene inactivation, and transgenesis. Methods in molecular biology (Clifton, NJ. 2008;420:175–95. 10.1007/978-1-59745-583-1_10 [DOI] [PubMed] [Google Scholar]
- 21. Hans S, Freudenreich D, Geffarth M, Kaslin J, Machate A, Brand M. Generation of a non-leaky heat shock-inducible Cre line for conditional Cre/lox strategies in zebrafish. Dev Dyn. 2011;240(1):108–15. 10.1002/dvdy.22497 [DOI] [PubMed] [Google Scholar]
- 22. Hans S, Kaslin J, Freudenreich D, Brand M. Temporally-controlled site-specific recombination in zebrafish. PloS one. 2009;4(2):e4640 10.1371/journal.pone.0004640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Knopf F, Hammond C, Chekuru A, Kurth T, Hans S, Weber CW, et al. Bone regenerates via dedifferentiation of osteoblasts in the zebrafish fin. Developmental cell. 2011;20(5):713–24. 10.1016/j.devcel.2011.04.014 [DOI] [PubMed] [Google Scholar]
- 24. Kroehne V, Freudenreich D, Hans S, Kaslin J, Brand M. Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors. Development (Cambridge, England). 2011;138(22):4831–41. 10.1242/dev.072587 [DOI] [PubMed] [Google Scholar]
- 25. Sadowski PD. The Flp recombinase of the 2-microns plasmid of Saccharomyces cerevisiae. Progress in nucleic acid research and molecular biology. 1995;51:53–91. . [PubMed] [Google Scholar]
- 26. Hirano N, Muroi T, Takahashi H, Haruki M. Site-specific recombinases as tools for heterologous gene integration. Applied microbiology and biotechnology. 2011;92(2):227–39. 10.1007/s00253-011-3519-5 [DOI] [PubMed] [Google Scholar]
- 27. Turan S, Bode J. Site-specific recombinases: from tag-and-target- to tag-and-exchange-based genomic modifications. Faseb J. 2011;25(12):4088–107. 10.1096/fj.11-186940 [DOI] [PubMed] [Google Scholar]
- 28. Metzger D, Chambon P. Site- and time-specific gene targeting in the mouse. Methods (San Diego, Calif. 2001;24(1):71–80. . [DOI] [PubMed] [Google Scholar]
- 29. Feil R, Wagner J, Metzger D, Chambon P. Regulation of Cre recombinase activity by mutated estrogen receptor ligand-binding domains. Biochemical and biophysical research communications. 1997;237(3):752–7. . [DOI] [PubMed] [Google Scholar]
- 30. Kurita R, Sagara H, Aoki Y, Link BA, Arai K, Watanabe S. Suppression of lens growth by alphaA-crystallin promoter-driven expression of diphtheria toxin results in disruption of retinal cell organization in zebrafish. Developmental biology. 2003;255(1):113–27. . [DOI] [PubMed] [Google Scholar]
- 31. Curado S, Anderson RM, Jungblut B, Mumm J, Schroeter E, Stainier DY. Conditional targeted cell ablation in zebrafish: a new tool for regeneration studies. Dev Dyn. 2007;236(4):1025–35. . [DOI] [PubMed] [Google Scholar]
- 32. Bailey JM, Creamer BA, Hollingsworth MA. What a fish can learn from a mouse: principles and strategies for modeling human cancer in mice. Zebrafish. 2009;6(4):329–37. 10.1089/zeb.2009.0626 [DOI] [PubMed] [Google Scholar]
- 33. Clark KJ, Balciunas D, Pogoda HM, Ding Y, Westcot SE, Bedell VM, et al. In vivo protein trapping produces a functional expression codex of the vertebrate proteome. Nature methods. 2011;8(6):506–15. 10.1038/nmeth.1606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ni TT, Lu J, Zhu M, Maddison LA, Boyd KL, Huskey L, et al. Conditional control of gene function by an invertible gene trap in zebrafish. Proceedings of the National Academy of Sciences of the United States of America. 2012. . [DOI] [PMC free article] [PubMed]
- 35. Trinh le A, Hochgreb T, Graham M, Wu D, Ruf-Zamojski F, Jayasena CS, et al. A versatile gene trap to visualize and interrogate the function of the vertebrate proteome. Genes & development. 2012;25(21):2306–20. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Mosimann C, Zon LI. Advanced zebrafish transgenesis with Tol2 and application for Cre/lox recombination experiments. Methods in cell biology. 2011;104:173–94. 10.1016/B978-0-12-374814-0.00010-0 [DOI] [PubMed] [Google Scholar]
- 37. Thummel R, Burket CT, Brewer JL, Sarras MP Jr, Li L, Perry M, et al. Cre-mediated site-specific recombination in zebrafish embryos. Dev Dyn. 2005;233(4):1366–77. . [DOI] [PubMed] [Google Scholar]
- 38. Mosimann C, Kaufman CK, Li P, Pugach EK, Tamplin OJ, Zon LI. Ubiquitous transgene expression and Cre-based recombination driven by the ubiquitin promoter in zebrafish. Development (Cambridge, England). 2011;138(1):169–77. 10.1242/dev.059345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Chandras C, Zouberakis M, Salimova E, Smedley D, Rosenthal N, Aidinis V. CreZOO—the European virtual repository of Cre and other targeted conditional driver strains. Database (Oxford). 2012;2012:bas029 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Smedley D, Salimova E, Rosenthal N. Cre recombinase resources for conditional mouse mutagenesis. Methods (San Diego, Calif. 2011;53(4):411–6. 10.1016/j.ymeth.2010.12.027 [DOI] [PubMed] [Google Scholar]
- 41. Nagy A, Mar L, Watts G. Creation and use of a cre recombinase transgenic database. Methods in molecular biology (Clifton, NJ. 2009;530:365–78. 10.1007/978-1-59745-471-1_19 [DOI] [PubMed] [Google Scholar]
- 42. Clark BS, Winter M, Cohen AR, Link BA. Generation of Rab-based transgenic lines for in vivo studies of endosome biology in zebrafish. Dev Dyn. 2011;240(11):2452–65. 10.1002/dvdy.22758 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Clark KJ, Geurts AM, Bell JB, Hackett PB. Transposon vectors for gene-trap insertional mutagenesis in vertebrates. Genesis. 2004;39(4):225–33. . [DOI] [PubMed] [Google Scholar]
- 44. Trinh le A, Fraser SE. Enhancer and gene traps for molecular imaging and genetic analysis in zebrafish. Development, growth & differentiation. 2013;55(4):434–45. . [DOI] [PubMed] [Google Scholar]
- 45. Asakawa K, Kawakami K. Targeted gene expression by the Gal4-UAS system in zebrafish. Development, growth & differentiation. 2008;50(6):391–9. . [DOI] [PubMed] [Google Scholar]
- 46. Asakawa K, Suster ML, Mizusawa K, Nagayoshi S, Kotani T, Urasaki A, et al. Genetic dissection of neural circuits by Tol2 transposon-mediated Gal4 gene and enhancer trapping in zebrafish. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(4):1255–60. 10.1073/pnas.0704963105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Scott EK, Baier H. The cellular architecture of the larval zebrafish tectum, as revealed by gal4 enhancer trap lines. Frontiers in neural circuits. 2009;3:13 10.3389/neuro.04.013.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Scott EK, Mason L, Arrenberg AB, Ziv L, Gosse NJ, Xiao T, et al. Targeting neural circuitry in zebrafish using GAL4 enhancer trapping. Nature methods. 2007;4(4):323–6. . [DOI] [PubMed] [Google Scholar]
- 49.Jungke P, Hans S, Brand M. The Zebrafish CreZoo: An Easy-to-Handle Database for Novel CreER-Driver Lines. Zebrafish. 2013. . [DOI] [PubMed]
- 50. Mongera A, Nusslein-Volhard C. Scales of fish arise from mesoderm. Curr Biol. 2013;23(9):R338–9. 10.1016/j.cub.2013.02.056 [DOI] [PubMed] [Google Scholar]
- 51. Tian T, Zhao L, Zhao X, Zhang M, Meng A. A zebrafish gene trap line expresses GFP recapturing expression pattern of foxj1b. Journal of genetics and genomics = Yi chuan xue bao. 2009;36(10):581–9. 10.1016/S1673-8527(08)60150-2 [DOI] [PubMed] [Google Scholar]
- 52. Brand M, Granato M, Nüsslein-Volhard C. Keeping and raising zebrafish Zebrafish—A Practical Approach: Oxford University Press, Oxford, UK.; 2002. [Google Scholar]
- 53. Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embryonic development of the zebrafish. Dev Dyn. 1995;203(3):253–310. . [DOI] [PubMed] [Google Scholar]
- 54. Abe G, Suster ML, Kawakami K. Tol2-mediated transgenesis, gene trapping, enhancer trapping, and the Gal4-UAS system. Methods in cell biology. 2004;104:23–49. . [DOI] [PubMed] [Google Scholar]
- 55. Reifers F, Bohli H, Walsh EC, Crossley PH, Stainier DY, Brand M. Fgf8 is mutated in zebrafish acerebellar (ace) mutants and is required for maintenance of midbrain-hindbrain boundary development and somitogenesis. Development (Cambridge, England). 1998;125(13):2381–95. . [DOI] [PubMed] [Google Scholar]
- 56. Westerfield M. The zebrafish book. A guide for the laboratory use ofzebrafish (Danio rerio) Univ of Oregon Press; 2000;4th edition. [Google Scholar]
- 57. Balciunas D, Davidson AE, Sivasubbu S, Hermanson SB, Welle Z, Ekker SC. Enhancer trapping in zebrafish using the Sleeping Beauty transposon. BMC genomics. 2004;5(1):62 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Mi H, Muruganujan A, Casagrande JT, Thomas PD. Large-scale gene function analysis with the PANTHER classification system. Nature protocols. 2013;8(8):1551–66. 10.1038/nprot.2013.092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Kawakami K. Tol2: a versatile gene transfer vector in vertebrates. Genome biology. 2007;8 Suppl 1:S7 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Kondrychyn I, Garcia-Lecea M, Emelyanov A, Parinov S, Korzh V. Genome-wide analysis of Tol2 transposon reintegration in zebrafish. BMC genomics. 2009;10:418 10.1186/1471-2164-10-418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Knowlton MN, Li T, Ren Y, Bill BR, Ellis LB, Ekker SC. A PATO-compliant zebrafish screening database (MODB): management of morpholino knockdown screen information. BMC bioinformatics. 2008;9:7 10.1186/1471-2105-9-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Zigman M, Laumann-Lipp N, Titus T, Postlethwait J, Moens CB. Hoxb1b controls oriented cell division, cell shape and microtubule dynamics in neural tube morphogenesis. Development (Cambridge, England). 2014;141(3):639–49. 10.1242/dev.098731 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Ekker M, Akimenko MA, Allende ML, Smith R, Drouin G, Langille RM, et al. Relationships among msx gene structure and function in zebrafish and other vertebrates. Molecular biology and evolution. 1997;14(10):1008–22. . [DOI] [PubMed] [Google Scholar]
- 64. Li Y, Allende ML, Finkelstein R, Weinberg ES. Expression of two zebrafish orthodenticle-related genes in the embryonic brain. Mechanisms of development. 1994;48(3):229–44. . [DOI] [PubMed] [Google Scholar]
- 65. Chen Y, Derin R, Petralia RS, Li M. Actinfilin, a brain-specific actin-binding protein in postsynaptic density. The Journal of biological chemistry. 2002;277(34):30495–501. . [DOI] [PubMed] [Google Scholar]
- 66. Cambier S, Gonzalez P, Durrieu G, Maury-Brachet R, Boudou A, Bourdineaud JP. Serial analysis of gene expression in the skeletal muscles of zebrafish fed with a methylmercury-contaminated diet. Environmental science & technology. 2010;44(1):469–75. . [DOI] [PubMed] [Google Scholar]
- 67. Jackson HE, Ingham PW. Control of muscle fibre-type diversity during embryonic development: the zebrafish paradigm. Mechanisms of development. 2013;130(9–10):447–57. 10.1016/j.mod.2013.07.002 [DOI] [PubMed] [Google Scholar]
- 68. Hammond KL, Whitfield TT. The developing lamprey ear closely resembles the zebrafish otic vesicle: otx1 expression can account for all major patterning differences. Development (Cambridge, England). 2006;133(7):1347–57. . [DOI] [PubMed] [Google Scholar]
- 69. Morsli H, Tuorto F, Choo D, Postiglione MP, Simeone A, Wu DK. Otx1 and Otx2 activities are required for the normal development of the mouse inner ear. Development (Cambridge, England). 1999;126(11):2335–43. . [DOI] [PubMed] [Google Scholar]
- 70. Murakami A, Ishida S, Thurlow J, Revest JM, Dickson C. SOX6 binds CtBP2 to repress transcription from the Fgf-3 promoter. Nucleic acids research. 2001;29(16):3347–55. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Stanford WL, Cohn JB, Cordes SP. Gene-trap mutagenesis: past, present and beyond. Nature reviews. 2001;2(10):756–68. . [DOI] [PubMed] [Google Scholar]
- 72. Friedel RH, Soriano P. Gene trap mutagenesis in the mouse. Methods in enzymology. 2010;477:243–69. 10.1016/S0076-6879(10)77013-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Schnutgen F, De-Zolt S, Van Sloun P, Hollatz M, Floss T, Hansen J, et al. Genomewide production of multipurpose alleles for the functional analysis of the mouse genome. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(20):7221–6. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Sivasubbu S, Balciunas D, Davidson AE, Pickart MA, Hermanson SB, Wangensteen KJ, et al. Gene-breaking transposon mutagenesis reveals an essential role for histone H2afza in zebrafish larval development. Mechanisms of development. 2006;123(7):513–29. . [DOI] [PubMed] [Google Scholar]
- 75.Ensembl. http://www.ensembl.org/Danio_rerio/Transcript/ProteinSummary?db=core;g=ENSDARG00000094992;r=17:24146910-24152572;t=ENSDART00000064083 2013. Available from: http://www.ensembl.org/Danio_rerio/Transcript/ProteinSummary?db=core;g=ENSDARG00000094992;r=17:24146910-24152572;t=ENSDART00000064083.
- 76. Lane BM, Lister JA. Otx but not Mitf transcription factors are required for zebrafish retinal pigment epithelium development. PloS one. 2012;7(11):e49357 10.1371/journal.pone.0049357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Foucher I, Mione M, Simeone A, Acampora D, Bally-Cuif L, Houart C. Differentiation of cerebellar cell identities in absence of Fgf signalling in zebrafish Otx morphants. Development (Cambridge, England). 2006;133(10):1891–900. . [DOI] [PubMed] [Google Scholar]
- 78. Liu X, Li Z, Emelyanov A, Parinov S, Gong Z. Generation of oocyte-specifically expressed cre transgenic zebrafish for female germline excision of loxP-flanked transgene. Dev Dyn. 2008;237(10):2955–62. 10.1002/dvdy.21701 [DOI] [PubMed] [Google Scholar]
- 79. Hesselson D, Anderson RM, Beinat M, Stainier DY. Distinct populations of quiescent and proliferative pancreatic beta-cells identified by HOTcre mediated labeling. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(35):14896–901. 10.1073/pnas.0906348106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Boniface EJ, Lu J, Victoroff T, Zhu M, Chen W. FlEx-based transgenic reporter lines for visualization of Cre and Flp activity in live zebrafish. Genesis. 2009;47(7):484–91. 10.1002/dvg.20526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Le X, Langenau DM, Keefe MD, Kutok JL, Neuberg DS, Zon LI. Heat shock-inducible Cre/Lox approaches to induce diverse types of tumors and hyperplasia in transgenic zebrafish. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(22):9410–5. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Guner-Ataman B, Paffett-Lugassy N, Adams MS, Nevis KR, Jahangiri L, Obregon P, et al. Zebrafish second heart field development relies on progenitor specification in anterior lateral plate mesoderm and nkx2.5 function. Development (Cambridge, England). 2013;140(6):1353–63. 10.1242/dev.088351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Jopling C, Sleep E, Raya M, Marti M, Raya A, Izpisua Belmonte JC. Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature. 2010;464(7288):606–9. 10.1038/nature08899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.ZFIN. Cre transgenics http://zfin.org/action/fish/do-search?geneOrFeatureName=cre&mutationType=Any&anatomyTermIDs=&anatomyTermNames=&searchTerm=&filter1=showAll&sortBy=BEST_MATCH&maxDisplayRecords=&page=1 2013. Available from: http://zfin.org/action/fish/do-search?geneOrFeatureName=cre&mutationType=Any&anatomyTermIDs=&anatomyTermNames=&searchTerm=&filter1=showAll&sortBy=BEST_MATCH&maxDisplayRecords=&page=1.
- 85. Amsterdam A, Becker TS. Transgenes as screening tools to probe and manipulate the zebrafish genome. Dev Dyn. 2005;234(2):255–68. . [DOI] [PubMed] [Google Scholar]
- 86. Copeland NG, Jenkins NA, Court DL. Recombineering: a powerful new tool for mouse functional genomics. Nature reviews. 2001;2(10):769–79. . [DOI] [PubMed] [Google Scholar]
- 87. Zu Y, Tong X, Wang Z, Liu D, Pan R, Li Z, et al. TALEN-mediated precise genome modification by homologous recombination in zebrafish. Nature methods. 2013;10(4):329–31. 10.1038/nmeth.2374 [DOI] [PubMed] [Google Scholar]
- 88. Auer TO, Duroure K, De Cian A, Concordet JP, Del Bene F. Highly efficient CRISPR/Cas9-mediated knock-in in zebrafish by homology-independent DNA repair. Genome research. 2014;24(1):142–53. 10.1101/gr.161638.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Auer TO, Del Bene F. CRISPR/Cas9 and TALEN-mediated knock-in approaches in zebrafish. Methods (San Diego, Calif. 2014. . [DOI] [PubMed]
- 90. Provost E, Rhee J, Leach SD. Viral 2A peptides allow expression of multiple proteins from a single ORF in transgenic zebrafish embryos. Genesis. 2007;45(10):625–9. . [DOI] [PubMed] [Google Scholar]
- 91. Wang X, Wan H, Korzh V, Gong Z. Use of an IRES bicistronic construct to trace expression of exogenously introduced mRNA in zebrafish embryos. BioTechniques. 2000;29(4):814–6, 8, 20. . [DOI] [PubMed] [Google Scholar]
- 92.Weber T, Koster R. Genetic Tools for Multicolor Imaging in zebrafish larvae. Methods (San Diego, Calif. 2013. . [DOI] [PubMed]
- 93. Wong ET, Ngoi SM, Lee CG. Improved co-expression of multiple genes in vectors containing internal ribosome entry sites (IRESes) from human genes. Gene therapy. 2002;9(5):337–44. . [DOI] [PubMed] [Google Scholar]
- 94. Ul-Hussain M, Dermietzel R, Zoidl G. Characterization of the internal IRES element of the zebrafish connexin55.5 reveals functional implication of the polypyrimidine tract binding protein. BMC molecular biology. 2008;9:92 10.1186/1471-2199-9-92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Uren AG, Mikkers H, Kool J, van der Weyden L, Lund AH, Wilson CH, et al. A high-throughput splinkerette-PCR method for the isolation and sequencing of retroviral insertion sites. Nature protocols. 2009;4(5):789–98. 10.1038/nprot.2009.64 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Comparison of the endogenous otx1b expression in wild-type embryos with the CreERT2 expression pattern in tud37Gt embryos which has been mapped to the otx1b locus at 24 and 48 hpf. fb: forebrain; mb: midbrain; ov: otic vesicle.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.