

Abstract
Rhodopsin is a well-characterized structural model of a G protein-coupled receptor. Photoisomerization of the covalently bound retinal triggers activation. Surprisingly, the x-ray crystal structure of the active Meta-II state has a 180° rotation about the long-axis of the retinal polyene chain. Unbiased microsecond-timescale all-atom molecular dynamics simulations show that the retinal cofactor can flip back to the orientation observed in the inactive state of rhodopsin under conditions favoring the Meta-I state. Our results provide, to our knowledge, the first evidence from molecular dynamics simulations showing how rotation of the retinal ligand within its binding pocket can occur in the activation mechanism of rhodopsin.
Main Text
Understanding the specific intermolecular interactions that lead to G protein-coupled receptor (GPCR) function is one of the most pressing needs in the design of targeted chemotherapeutics. Rhodopsin, a prototypical GPCR, is activated when its covalently bound chromophore, retinal, absorbs a photon and undergoes an 11-cis → all-trans isomerization, culminating in an equilibrium between inactive Meta-I and active Meta-II forms (Fig. 1, a–c and Fig. S1 in the Supporting Material) (1). Most NMR spectroscopic and crystallographic structures of various photointermediates reveal the retinal ligand acting as an inverse agonist (2–5). However, recent x-ray structures of the active Meta-II state are very surprising, in that retinal is rotated 180° about its long axis versus inactive photointermediates (6,7). This has led to uncertainty as to the role of the retinal orientation: does the polyene chain undergo a long-axis rotation in the transition from Meta-I to Meta-II—and if so, what is the biological relevance of this striking large-scale movement?
Figure 1.
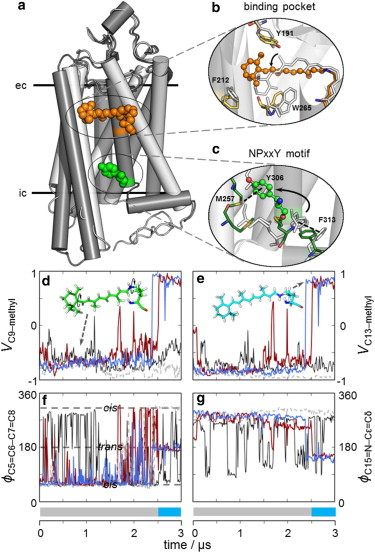
Microsecond timescales are required for retinal flip in rhodopsin. (a) Retinal undergoes an 11-cis to all-trans isomerization upon photon absorption, initiating large-scale conformational changes in rhodopsin (dark gray, dark state (PDB:1U19); light gray, Meta-II-like (PDB:3PXO)). An outward tilt and elongation of TM5 and outward rotation of TM6 are conserved in Class A GPCRs. (b) A 180° rotation occurs about the long axis of the retinylidene chain in Meta-II, changing the orientation of the C9- and C13-methyl groups from the extracellular (ec) (gray) to the cytoplasmic (ic) side (orange) of the protein. (c) Conserved residue Tyr3067.53 of the NPxxY motif toggles between π-π interactions with Phe3137.60 (dark) and Met2576.40 (Meta-II) (gray, dark; green, Meta-II). (d–g) MD trajectories for rhodopsin in various lipid bilayers (black, Meta-II; blue, DOPC/ DOPE (3:1); gray, DOPC/DOPE (1:1); and red, DOPC/DOPE (1:3)). (d) The C9-CH3 and (e) C13-CH3 bond orientational time evolution Vi defined as the cosine angle of the bond vector with respect to the membrane normal. At ∼2.5 μs, the retinylidene moiety undergoes a long-axis 180° rotation in two trajectories. (Insets) Retinal cofactor before (d) and after (e) the flip. The C5=C6–C7=C8 dihedral (f) and the C15–NZ–Cε–Cδ dihedral (g) limit rotation to the polyene chain; the β-ionone ring undergoes a 6-s-cis to trans conformational transition. To see this figure in color, go online.
We used unbiased μs-timescale, all-atom molecular dynamics (MD) simulations of rhodopsin in a series of lipid bilayers, taking advantage of the microscopic reversibility of the Meta-I to Meta-II transition to investigate the behavior of the retinal orientation. Our results clearly show that upon photoactivation retinal possesses remarkable flexibility: we present what is, to our knowledge, the first theoretical evidence of a long-axis flip of the retinal cofactor in the rhodopsin activation process. In two of three simulations, long-axis rotation of the retinal polyene chain was clearly observed, leading to a reorientation toward the extracellular side of the protein. These results create an important bridge between spectroscopic and crystallographic studies (1,3). Furthermore, these results provide evidence for a loose coupling between agonist and receptor upon GPCR activation, as suggested from experimental and computational studies (1,8,9).
We utilized the fact that the lipid bilayer composition can significantly influence the Meta-I to Meta-II equilibrium (1,10). Our hypothesis was that long-axis rotation of the polyene chain occurs in Meta-II due to an ensemble of activated conformations that all possess a larger binding pocket than inactive forms of rhodopsin. To test this hypothesis, we modeled the Meta-II structure of rhodopsin under Meta-I protonation states embedded in a bilayer of lipids known to forward- or back-shift the Meta-I to Meta-II equilibrium using μs-long all-atom MD simulations (1) (for details, see the Supporting Material). Meta-II conformations are defined by the state of two protonation switches involving the protonated Schiff base (SB) with its complex counterion, as well as Glu1343.49 (superscript denotes Ballesteros-Weinstein numbering) of the conserved E(D)RY motif (1). Furthermore, the reversible transition to an inactive conformation initially requires the polyene chain to rotate with the C9- and C13-methyl groups pointing toward extracellular loop-2 (EL2) as in the dark and Meta-I states.
All-atom MD simulations have proven invaluable to the characterization of the structure-function relationship in rhodopsin activation. In particular, MD studies on the μs-timescale have reproduced solid-state 2H NMR spectra of the Meta-I state (11), and showed an increase in both hydration and retinal flexibility versus the dark state (12). In this study, the most remarkable event occurs at roughly 2.5 μs in two of the three Meta-II to Meta-I simulations: the polyene chain in retinal undergoes a 180° rotation about its long axis, in which the C9- and C13-methyl groups shift orientation from the cytoplasmic side toward the extracellular side of rhodopsin (Fig. 1, d and e). Once the reorientation occurs, the retinal polyene chain remains stable for the final 500 ns of the simulation. The rotation is notable because our simulations are unbiased, and our control simulation with Meta-II protonation states showed no rotation of the polyene chain. The long-axis rotation is directly related to the dihedral fluctuations that connect both ends of the chromophore, i.e., the β-ionone ring and the retinylidene SB linkage. In particular, the distribution of the dihedral angle for the C6–C7 bond connecting the β-ionone ring to the polyene chain correlates well with previous studies that identified three minima for retinal with a protonated SB (Figs. 1, f and g, and S2) (13). The final orientation of the β-ionone ring is the 6-s-trans conformation, a local minimum for the C5=C6–C7=C8 dihedral. The β-ionone ring remains essentially fixed during the polyene chain rotation, with a transition from 6-s-cis in the starting structure to 6-s-trans. It has been shown that the 6-s-trans conformation raises the pKa of the SB, favoring the protonated state (13), as in Meta-I. Moreover, the pathway that occurs from cis to trans is very similar to the one observed in previous long-timescale MD studies that examined the dark → Meta-I transition (11,12).
Hydration plays a crucial role in rhodopsin function, in which water is necessary for rearrangement of the chromophore (12,14). The retinal flip is accompanied by a decrease in hydration of the binding pocket (Fig. 2 a). Expulsion of water through switching to a Meta-I-like orientation stems from efficient packing between the side chains within the binding pocket and the methyl groups of the retinal (Fig. 2, b and c). The C9-methyl is directly coupled to the protonation switch of Glu1343.49 in fully activated rhodopsin. Removal of the C9-methyl back-shifts rhodopsin to the preactive Meta-I state (15). The back-rotation of the polyene chain reestablishes these interactions, in particular between the C9-methyl and Tyr191EL2. In addition to these interactions, the CWxP motif is part of a transmission switch that is common to many Class A GPCRs, which involves a rotation of Trp2656.48 and Phe2125.47 that facilitates the outward rotation of TM6 from the inactive to the active state (16). Both flipping events were promptly followed by rotation of Trp2656.48 around its χ1 dihedral, leading to a rearrangement of the indole ring to maintain stabilizing interactions with the retinal (Fig. 2 d). Another microdomain critical to GPCR activation involves the conserved NPxxY motif, where Tyr3067.53 toggles between aromatic stacking with Phe3137.60 in the inactive state and participating in a water-mediated hydrogen-bonding network with Met2576.40 in the active state. Both the Tyr3067.53–Phe3137.60 and Tyr3067.53–Met2576.40 interactions occur in our simulations, with no correlation to the flip. Although in the Meta-II state Tyr3067.53 interacts with Met2576.40, this coupling disrupts a staggered interaction among Met2536.36, Met2576.40, and Met3097.56 that is present in the dark state. After ∼0.5 μs in two simulations, Tyr3067.53 rotates outward to reestablish a π-π stacking interaction with Phe3137.60 (Fig. S3, a–d). This outward rotation also allows the three methionine side chains to reform the staggered interactions present in the dark state (Fig. S3 e).
Figure 2.
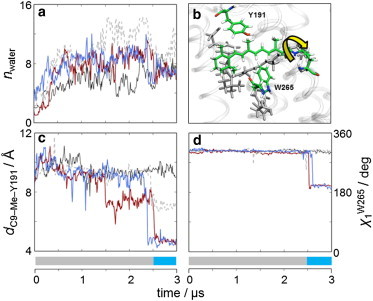
Retinal polyene flip is linked to changes in binding pocket hydration and conserved GPCR motifs. (a) Back-flip of the retinal polyene chain leads to an expulsion of water from the binding pocket (black, Meta-II; blue, DOPC/DOPE (3:1); gray, DOPC/DOPE (1:1); and red, DOPC/DOPE (1:3)). (b) Visualization of the retinal binding pocket before and after the flip from the DOPC/DOPE (3:1) trajectory. (c) Nonbonded interactions between the C9-methyl and Tyr191EL2 are reestablished after the polyene chain flip. (d) Rotation of Trp2656.48 in the CWxY motif is correlated with the retinal flip. To see this figure in color, go online.
The flip about the polyene long axis was an unanticipated result from our attempt to model the deactivation of rhodopsin. Interestingly, however, it provides essential insights into the dynamics of the retinal binding pocket (7). The Meta-II structure was obtained from opsin crystals soaked with all-trans retinal (7), and this characterization was later supported by a structure of a constitutively active rhodopsin (6). However, comparisons between the Meta-II crystal structure and solid-state NMR studies of frozen detergent-solubilized rhodopsin in the Meta-II state (17) reveal several discrepancies. If one assumes that trapping of the Meta-II photointermediate was equivalent in both NMR and crystallography, then the orientation of retinal in the NMR studies represents an average of those in equilibrium. By starting from the crystal structure and observing a stable 180° rotation about the long axis of the polyene chain, our simulations provide a plausible explanation for these differences. Furthermore, our results provide compelling support for our proposal that the increased ligand flexibility observed in Meta-I could lead to multiple retinal conformations in the active state (12).
Very few computational studies have been carried out on rhodopsin in the Meta-II state (18). This is the first attempt, to our knowledge, to use microscopic reversibility together with μs-long simulations to model the deactivation of rhodopsin. Our attempt to perturb this equilibrium starting from active Meta-II has led to the surprising discovery of a back-flip of retinal to an orientation found in the inactive photointermediates. The timescale of the flip occurs so quickly that it suggests the retinal binding pocket has evolved to stabilize the polyene chain in either of two conformations (i.e., up or down). In addition, these results provide compelling support for previous studies on GPCRs that identified loose coupling between agonist and protein upon activation (1,8; see Fig. S4 and Tables S1 and S2 in the Supporting Material). The role of the retinal flip in the interaction of transducin with rhodopsin, and the correspondence to other members of Class A GPCRs, remain as important questions for future research.
Author Contributions
J.F. and B.M. designed research; J.F. performed research; J.F. and B.M. analyzed results; and J.F., M.F.B., and B.M. wrote the article.
Acknowledgments
The authors thank M. Pitman, A. Struts, and T. Sakmar for discussions.
This work was supported by the National Institutes of Health (to M.F.B.) and West Virginia University (to B.M.). Computational time was provided through XSEDE allocation No. TG-MCB090200.
Editor: Scott Feller.
Supporting Material
Time evolution from the DOPC:DOPE (3:1) trajectory in which the polyene chain flips from the cytoplasmic side of the protein and Trp2656.48 (bottom) towards the extracellular lid of the protein and Tyr191EL2 (top). Note the two-state conformation of the polyene chain (“down” to “up”) and the fluctuation of the Trp2656.48 χ1 dihedral (part of the “trans-mission switch” (33)).
References
- 1.Mahalingam M., Martínez-Mayorga K., Vogel R. Two protonation switches control rhodopsin activation in membranes. Proc. Natl. Acad. Sci. USA. 2008;105:17795–17800. doi: 10.1073/pnas.0804541105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Standfuss J., Edwards P.C., Schertler G.F.X. The structural basis of agonist-induced activation in constitutively active rhodopsin. Nature. 2011;471:656–660. doi: 10.1038/nature09795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nakamichi H., Okada T. Crystallographic analysis of primary visual photochemistry. Angew. Chem. Int. Ed. Engl. 2006;45:4270–4273. doi: 10.1002/anie.200600595. [DOI] [PubMed] [Google Scholar]
- 4.Palczewski K., Kumasaka T., Miyano M. Crystal structure of rhodopsin: a G protein-coupled receptor. Science. 2000;289:739–745. doi: 10.1126/science.289.5480.739. [DOI] [PubMed] [Google Scholar]
- 5.Han M., Groesbeek M., Smith S.O. The C9 methyl group of retinal interacts with glycine-121 in rhodopsin. Proc. Natl. Acad. Sci. USA. 1997;94:13442–13447. doi: 10.1073/pnas.94.25.13442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Deupi X., Edwards P., Standfuss J. Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. Proc. Natl. Acad. Sci. USA. 2012;109:119–124. doi: 10.1073/pnas.1114089108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Choe H.-W., Kim Y.J., Ernst O.P. Crystal structure of metarhodopsin II. Nature. 2011;471:651–655. doi: 10.1038/nature09789. [DOI] [PubMed] [Google Scholar]
- 8.Dror R.O., Arlow D.H., Shaw D.E. Activation mechanism of the β2-adrenergic receptor. Proc. Natl. Acad. Sci. USA. 2011;108:18684–18689. doi: 10.1073/pnas.1110499108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Deupi X., Kobilka B.K. Energy landscapes as a tool to integrate GPCR structure, dynamics, and function. Physiology (Bethesda) 2010;25:293–303. doi: 10.1152/physiol.00002.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zaitseva E., Brown M.F., Vogel R. Sequential rearrangement of interhelical networks upon rhodopsin activation in membranes: the Meta IIa conformational substate. J. Am. Chem. Soc. 2010;132:4815–4821. doi: 10.1021/ja910317a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Martínez-Mayorga K., Pitman M.C., Brown M.F. Retinal counterion switch mechanism in vision evaluated by molecular simulations. J. Am. Chem. Soc. 2006;128:16502–16503. doi: 10.1021/ja0671971. [DOI] [PubMed] [Google Scholar]
- 12.Leioatts N., Mertz B., Brown M.F. Retinal ligand mobility explains internal hydration and reconciles active rhodopsin structures. Biochemistry. 2014;53:376–385. doi: 10.1021/bi4013947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhu S., Brown M.F., Feller S.E. Retinal conformation governs pKa of protonated Schiff base in rhodopsin activation. J. Am. Chem. Soc. 2013;135:9391–9398. doi: 10.1021/ja4002986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Angel T.E., Gupta S., Chance M.R. Structural waters define a functional channel mediating activation of the GPCR, rhodopsin. Proc. Natl. Acad. Sci. USA. 2009;106:14367–14372. doi: 10.1073/pnas.0901074106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Vogel R., Lüdeke S., Sheves M. Agonists and partial agonists of rhodopsin: retinal polyene methylation affects receptor activation. Biochemistry. 2006;45:1640–1652. doi: 10.1021/bi052196r. [DOI] [PubMed] [Google Scholar]
- 16.Deupi X., Standfuss J. Structural insights into agonist-induced activation of G-protein-coupled receptors. Curr. Opin. Struct. Biol. 2011;21:541–551. doi: 10.1016/j.sbi.2011.06.002. [DOI] [PubMed] [Google Scholar]
- 17.Ahuja S., Crocker E., Smith S.O. Location of the retinal chromophore in the activated state of rhodopsin. J. Biol. Chem. 2009;284:10190–10201. doi: 10.1074/jbc.M805725200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rose A.S., Elgeti M., Hildebrand P.W. Position of transmembrane helix 6 determines receptor G protein coupling specificity. J. Am. Chem. Soc. 2014;136:11244–11247. doi: 10.1021/ja5055109. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Time evolution from the DOPC:DOPE (3:1) trajectory in which the polyene chain flips from the cytoplasmic side of the protein and Trp2656.48 (bottom) towards the extracellular lid of the protein and Tyr191EL2 (top). Note the two-state conformation of the polyene chain (“down” to “up”) and the fluctuation of the Trp2656.48 χ1 dihedral (part of the “trans-mission switch” (33)).