

Abstract
There have been studies on health beneficial effects of ginger and its components. However, there still remain certain aspects that are not well defined in their anti-hyperglycemic effects. Our aims were to find evidence of possible mechanisms for antidiabetic action of [6]-gingerol, a pungent component of ginger, employing a rat skeletal muscle-derived cell line, a rat-derived pancreatic β-cell line, and type 2 diabetic model animals. The antidiabetic effect of [6]-gingerol was investigated through studies on glucose uptake in L6 myocytes and on pancreatic β-cell protective ability from reactive oxygen species (ROS) in RIN-5F cells. Its in vivo effect was also examined using obese diabetic db/db mice. [6]-Gingerol increased glucose uptake under insulin absent condition and induced 5′ adenosine monophosphate-activated protein kinase phosphorylation in L6 myotubes. Promotion by [6]-gingerol of glucose transporter 4 (GLUT4) translocation to plasma membrane was visually demonstrated by immunocytochemistry in L6 myoblasts transfected with glut4 cDNA-coding vector. [6]-Gingerol suppressed advanced glycation end product-induced rise of ROS levels in RIN-5F pancreatic β-cells. [6]-Gingerol feeding suppressed the increases in fasting blood glucose levels and improved glucose intolerance in db/db mice. [6]-Gingerol regulated hepatic gene expression of enzymes related to glucose metabolism toward decreases in gluconeogenesis and glycogenolysis as well as an increase in glycogenesis, thereby contributing to reductions in hepatic glucose production and hence blood glucose concentrations. These in vitro and in vivo results strongly suggest that [6]-gingerol has antidiabetic potential through multiple mechanisms.
Electronic supplementary material
The online version of this article (doi:10.1007/s10616-014-9730-3) contains supplementary material, which is available to authorized users.
Keywords: AMPK, Gingerol, GLUT4, Immunocytochemistry, L6 myocytes, RIN-5F cells
Introduction
Type 2 diabetes and pre-diabetes are dramatically increasing worldwide. The pathophysiological factors of type 2 diabetes mellitus consist of insulin resistance and pancreatic β-cell failure. Thus, the available anti-hyperglycemic agents related to these causes are necessary to treat and prevent type 2 diabetes (Del Prato and Marchetti 2004). Insulin resistance causes a decrease of glucose uptake and its utilization in insulin sensitive tissues. The majority (~75 %) of glucose uptake in the post-prandial state is accomplished in the skeletal muscle by the translocation of glucose transporter 4 (GLUT4) toward plasma membrane (Saltiel and Kahn 2001). GLUT4 translocation in muscle cells is known to depend on two divergent signaling pathways. The first pathway involves the binding of insulin to insulin receptor and the subsequent activation of phosphatidylinositol-3 kinase (PI3K) and Akt (Imamura et al. 2003). On the other hand, 5′ adenosine monophosphate-activated protein kinase (AMPK), regarded as cellular energy sensor, has been shown to promote GLUT4 translocation independently of insulin (Huang and Czech 2007).
Hyperglycemia stimulates the endogenous nonenzymatic reaction of glucose with proteins and this reaction results in the generation of advanced glycation end products (AGEs). AGEs interact with certain receptors for them, thus leading to enhanced oxidative stress in various tissues. In particular, an important part of pancreatic β-cell damage related to diabetes is mediated by free radical, suggesting the protection of pancreatic β-cell from reactive oxygen species (ROS) has a crucial role in improving pancreatic β-cell damage (Lin et al. 2012).
Natural plants contain various medicinal compounds for metabolic syndrome including diabetes. Ginger, which is the rhizome of the plant Zingiber officinale Roscoe, is widely used in several food products such as soft beverages and also in many types of pharmaceutical formulations (Kubra and Rao 2012). Among the various derivatives of ginger, [6]-gingerol has been identified as, for instance, antioxidant, anti-inflammatory and anti-cancer principle (Dugasani et al. 2010; Yagihashi et al. 2008). Although there have been studies concerned with anti-hyperglycemic and antioxidant effects of ginger and [6]-gingerol (Al-Amin et al. 2006; Chakraborty et al. 2012), there still remain certain aspects that are not well defined in their anti-hyperglycemic effects and mechanisms.
In the present study, we investigated the effect of [6]-gingerol through the studies on molecular mechanisms using cultured rat skeletal muscle cells (L6 myocytes). We also examined whether or not [6]-gingerol would protect pancreatic β-cells from reactive oxygen species (ROS)-induced stress employing cultured pancreatic β-cells (RIN-5F). To confirm anti-hyperglycemic effect of [6]-gingerol in vivo, we also examined its effect on the fasting blood glucose level, glucose intolerance and gene expression of hepatic enzymes related to glucose metabolism in type 2 diabetic model db/db mice.
Materials and methods
Materials
[6]-Gingerol was purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). A rat skeletal muscle-derived cell line of L6 myoblasts and a rat pancreatic β-cell-derived cell line of RIN-5F cells were obtained from American Type Culture Collection (Manassas, VA, USA; ATCC® numbers: CRL-1458 and CRL-2058, respectively). Dulbecco’s modified Eagle medium (DMEM) and RPMI 1640 medium were purchased from Nissui Pharmaceutical Co. (Tokyo, Japan). Fetal bovine serum (FBS) was from JRH Biosciences (Lenexa, KS, USA). Streptomycin and penicillin G were from Nacalai Tesque, Inc. (Kyoto, Japan). Bovine serum albumin (BSA, fatty acid free) and Triton X-100 were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Glucose, triglyceride (TG) and total cholesterol (TC) assay kits (Glucose CII-Test Wako, Triglyceride E-Test Wako and Cholesterol E-Test Wako, respectively) were from Wako Pure Chemical Industries. Serum lipid peroxide was estimated as thiobarbituric acid-reactive substances (TBARS) with a commercial kit (TBARS Assay Kit), ZeptoMetrix Corporation (Buffalo, NY, USA). Tumor necrosis factor (TNF)-α assay kit was from Shibayagi Co., Ltd. (Gunma, Japan). Compound C was from Wako Pure Chemical Industries. The anti-phospho-AMPKα (Thr172) and anti-AMPK antibodies were obtained from Cell Signaling Technology, Inc. (Beverly, MA, USA). Horseradish peroxidase-conjugated anti-rabbit IgG antibody was from Amersham Biosciences (Little Chalfont, UK). Anti-HaloTag® rabbit polyclonal antibody was from Promega KK (Tokyo, Japan). Anti-caveolin-3 goat polyclonal IgG was from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). Alexa Fluor 555-conjugated anti-rabbit IgG was from Invitrogen (Carlsbad, CA, USA). FITC-conjugated anti-goat IgG was from Santa Cruz Biotechnology Inc. All other chemicals were of the best grade commercially available, unless otherwise noted. Plastic multiwell plates and tubes were obtained from Nunc A/S (Roskilde, Denmark) or Iwaki brand, Asahi Glass Co., Ltd. (Tokyo, Japan).
Determination of glucose uptake by cultured L6 myotubes
Stock cultures of L6 myoblasts were maintained in DMEM supplemented with 10 % (v/v) FBS, streptomycin (100 µg/ml), and penicillin G (100 U/ml) (10 % FBS/DMEM) under an atmosphere of 5 % CO2/95 % humidified air at 37 °C as described previously (Yagasaki et al. 2003). Effect of [6]-gingerol on glucose uptake was examined by the procedure described previously (Kawano et al. 2009; Ha et al. 2012) with slight modifications. Briefly, L6 myoblasts (5 × 104 cells/well) were cultured in Nunc 24-place multiwell plates and grown for 11 days to form myotubes in 0.4 ml of 10 % FBS/DMEM. The 11-day-old myotubes were preincubated for 2 h in Krebs-Henseleit buffer (pH 7.4) containing 0.1 % BSA, 10 mM Hepes and 2 mM sodium pyruvate (KHH buffer). The myotubes were then incubated in KHH buffer containing 11 mM glucose without or with [6]-gingerol (0–100 μM) for 4 h. [6]-Gingerol was dissolved in dimethyl sulfoxide. Glucose concentrations in KHH buffer were determined with a glucose assay kit, and the amounts of glucose uptake were calculated from the differences in glucose concentrations between before and after incubation with [6]-gingerol.
Western blotting analysis
Western blotting analysis was carried out as described previously (Minakawa et al. 2011). Briefly, L6 myoblasts (5 × 105 cells) were cultured in Nunc 60 mm dishes and grown for 11 days to form myotubes in 3 ml of 10 % FBS/DMEM. The myotubes were incubated for 2 h in KHH buffer, and they were then incubated in KHH buffer containing 11 mM glucose without or with [6]-gingerol for 30, 60, 120 and 240 min. The lysates were subjected to Western blotting analysis for AMPK and phospho-AMPK.
Transfection of pFN21A (HaloTag®7)-glut4 vector into L6 myoblasts and immunocytochemistry
Effect of [6]-gingerol on glucose uptake and GLUT4 translocation was examined by the procedure described previously using HaloTag®-mock vector or HaloTag®-glut4 vector (Minakawa et al. 2012). Briefly, L6 myoblasts (5 × 104) were cultured in a 24-well culture plate (Nunc) for glucose uptake assay or an 8-well chamber slide (Nunc) for immunocytochemistry. L6 myoblasts were transfected with HaloTag®-mock or HaloTag®-glut4 vector. Cells were used for the glucose uptake assay at 48 h after transfection and for immunocytochemistry at 36 h after transfection. Glucose uptake assay was conducted as mentioned above, and immunocytochemistry was conducted to detect GLUT4 and caveolin-3. Anti-HaloTag rabbit polyclonal antibody and anti-caveolin-3 goat polyclonal antibody were used for the first antibodies. Alexa Fluor 555-conjugated anti-rabbit IgG and FITC-conjugated anti-goat IgG were used for second antibodies.
Synthesis of advanced glycation end product
Artificial advanced glycation end product (AGE) was generated by incubation of BSA with d-glyceraldehyde according to the method of Kume et al. (2005). BSA was incubated at 37 °C for 2 weeks in the presence of d-glyceraldhyde (designated as AGE2). BSA alone was similarly incubated at 37 °C for 2 weeks under conditions without any carbohydrates, and employed as the control for AGE2.
Measurement of intracellular reactive oxygen species in RIN-5F cells
Intracellular reactive oxygen species (ROS) levels were measured as described previously (Minakawa et al. 2011) with slight modifications. Briefly, RIN-5F cells derived from rat pancreatic β-cells were maintained in RPMI 1640 supplemented with 10 % (v/v) FBS, streptomycin (100 µg/ml), and penicillin G (100 U/ml) (10 % FBS/RPMI 1640) under an atmosphere of 5 % CO2/95 % humidified air at 37 °C. The cells (5 × 105 cells/well) were cultured in Nunc 12-place multiwell plates. After being cultured for 72 h in 1 ml of 10 % FBS/RPMI 1640, the medium in each well was removed. Thereafter, RIN-5F cells received 1 ml of fresh medium (1 % FBS/RPMI 1640) without or with [6]-gingerol and AGE2. After the 4 h incubation, RIN-5F cells were incubated with DCFH-DA in 1 % FBS/RPMI 1640 at 37 °C for 20 min. At the end of the incubation, DCFH fluorescence of the cells from each well was measured at an emission wavelength of 530 nm and an excitation wavelength of 488 nm with a flow cytometer (EPICS ELITE EPS, Beckman-Coulter, Hialeah, FL).
Effect of [6]-gingerol on blood glucose levels in db/db mice
All animal experiments were conducted in accordance with guidelines established by the Animal Care and Use Committee of Tokyo University of Agriculture and Technology and were approved by this committee. Male db/db and its misty (m/m) control (normal) mice (5 weeks old) were obtained from Charles River Japan (Kanagawa, Japan). Animals were individually housed in stainless-steel cages with wire bottoms in an air-conditioned room with a temperature of 22 ± 2 °C, a relative humidity of 60 ± 5 %, and an 8:00–20:00 light cycle. All mice were maintained on a stock CE-2 pellet diet (CLEA Japan, Tokyo, Japan) for 3 days and thereafter on a basal 20 % casein diet (20C) for 4 days. The composition of the 20C diet (AIN-93 formula) was described elsewhere (Minakawa et al. 2011). After preliminary feeding for 1 week, mice were deprived of their diet at 9:00 but allowed free access to water until blood collection from tail vein 3 h later. After determination of blood glucose concentrations with a commercial kit as described previously (Minakawa et al. 2011), db/db mice were divided into two groups of similar fasting blood glucose levels and body weights (0 week). Diabetic mice of each of the two groups were given either the 20C as the control (CNT) group or the 20C supplemented with 0.05 % [6]-gingerol as the test (GIN) group for 4 weeks. Likewise, misty mice were given the 20C as the normal (NOR) group for 4 weeks. Water and each diet were available at all times except for the experiments to determine fasting blood glucose levels, which were carried out every week. After 4 weeks of feeding, whole blood was collected from all mice by cardiac puncture under somnopentyl anesthesia. Concentrations of serum lipids, TBARS and TNF-α were measured with commercial kits.
Intraperitoneal glucose tolerance test
Intraperitoneal glucose tolerance test (IPGTT) was performed as described previously (Kawano et al. 2009) after the determination of fasting blood glucose levels at the 3rd week of feeding. Two groups of db/db mice were deprived of their diet at 20:00 but allowed free access to water. After fasted for 15 h, [6]-gingerol (test group) was orally given at a dose of 5 mg/ml of 0.1 % carboxymethyl cellulose sodium salt (CMC) solution/100 g body weight. Similarly, 0.1 % CMC solution alone (1 ml/100 g body weight) was orally given to the control group as a vehicle. Two hours later, blood was collected from the tail vein of db/db mice (0 min). Immediately after blood collection, diabetic mice received an intraperitoneal injection of glucose (0.2 g/ml/100 g body weight). Blood samples were successively collected at appropriate time intervals (30, 60, 90 and 120 min), and blood glucose levels were determined as described above.
Quantitative RT-PCR analyses
Quantitative RT-PCR analyses were conducted as described previously (Son et al. 2013). Total RNA was isolated from the liver using modified AGPC method (Chomczynski and Sacchi 1987). cDNA was synthesized using SuperScript™ III Reverse Transcriptase kit (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. Real-time PCR was performed with FastStart SYBR green Master Mix (Roche Diagnostics, Mannheim, Germany) in Thermal Cycler Dice® Real Time System II (Takara Bio, Inc., Tokyo, Japan). The PCR reaction was performed in duplicate. The relative expression levels of the target genes to the expression level of the endogenous reference gene β-actin were calculated using the delta cycle threshold method. The primer sequences are listed in supplementary Table 1.
Statistical analyses
Data are presented as means ± standard errors of means (SEM). Multigroup comparisons were conducted by one-way analysis of variance followed by Tukey–Kramer multiple comparisons test and differences between two group means were compared by Student’s t test. Values of P < 0.05 were considered statistically significant.
Results
Effect of [6]-gingerol on glucose uptake in cultured L6 myotubes
[6]-Gingerol dose dependently and significantly increased glucose uptake in L6 myotubes at concentrations up to 100 μM in the absence of insulin (Fig. 1a). The addition of insulin (100 nM) did not affect glucose uptake in L6 myotubes, indicating that L6 myotubes were insulin-insensitive under our culture conditions. However, the treatment of L6 myotubes with [6]-gingerol alone and combination of insulin and [6]-gingerol significantly increased glucose uptake in the cells (Fig 1b). In many cases, the differentiation to myotubes from myoblasts is performed under a concentration of 2 % FBS like in our previous study (Ha et al. 2012). In the present study, L6 myoblasts were cultured and differentiated to L6 myotubes in the medium containing a higher concentration (10 %) of FBS, because suppliers and hence lots of FBS were different. Previous reports have shown that sustained high glucose and insulin exposure decreases insulin-stimulated GLUT4 translocation in L6 myotubes (Huang et al. 2002). Thus, relatively high concentration of insulin present in FBS might be considered as one of reasons for insulin-insensitivity as seen in Fig. 1b, although precise reasons remain to be elucidated. Compound C, an AMPK inhibitor (Zhou et al. 2001), significantly canceled the [6]-gingerol-induced increase in glucose uptake at a concentration of 10 μM, where compound C alone exerted no influence on glucose uptake (Fig. 1c). In our preliminary experiment, we have examined the dose-dependent effect of compound C on glucose uptake to find an appropriate concentration where compound C itself exerts no influence, because we considered that the inhibitor alone should not affect glucose uptake in this kind of experiment. As a result, we found that 10 μM of compound C was optimum.
Fig. 1.
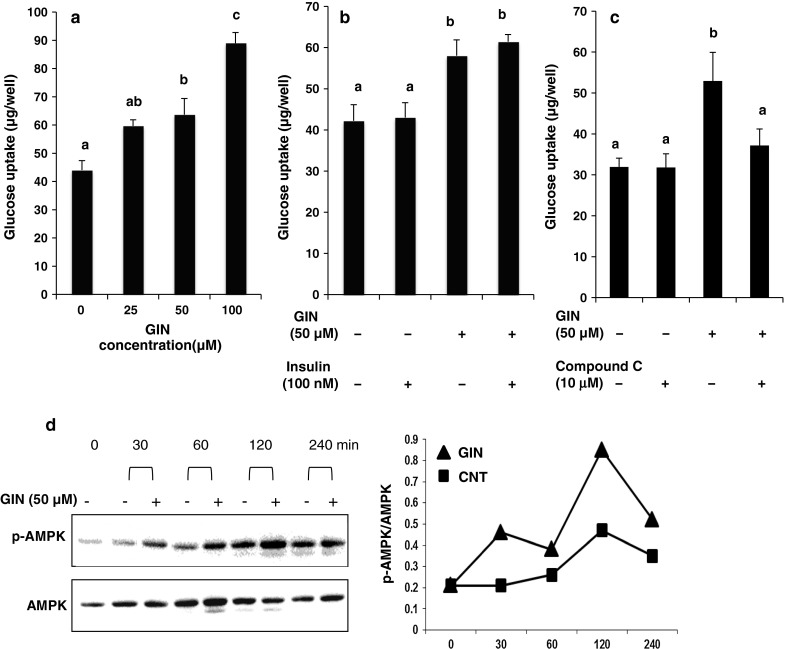
Effect of [6]-gingerol on glucose uptake and AMPK phosphorylation in L6 myotubes. a, b, c Glucose uptake assay was conducted without or with [6]-gingerol (0, 25, 50 and 100 µM), insulin (100 nM) and compound C (10 µM). Each value represents the mean ± SEM for 6 wells. Values not sharing a common letter are significantly different at P < 0.05 by Tukey–Kramer multiple comparisons test. d Western blotting analysis for AMPK phosphorylation. Western blotting was conducted with anti-phospho-AMPK and anti-AMPK antibodies
Effect of [6]-gingerol on phosphorylation of AMPK in cultured L6 myotubes
L6 myotubes were treated with 50 μM [6]-gingerol for 30–240 min. [6]-Gingerol stimulated the phosphorylation of AMPK (the ratio of phosphorylated AMPK to total AMPK, p-AMPK/AMPK) from 30 min after treatment, which peaked after 120 min (Fig. 1d).
Effect of [6]-gingerol on glucose uptake in cultured L6 myoblasts transfected with HaloTag®-mock vector or HaloTag®-glut4 vector
Glucose uptake for 4 h was examined in L6 myoblasts 48 h after transfection of HaloTag®-mock vector or HaloTag®-glut4 vector (Fig. 2a). Glucose uptake was significantly higher in HaloTag®-glut4 vector-transfected cells than in HaloTag®-mock vector-transfected cells (mock GIN− vs. glut4 GIN−), suggesting that Halo-GLUT4 protein derived from transfected HaloTag®-glut4 could operate in glucose uptake. [6]-Gingerol (50 μM) significantly promoted glucose uptake in L6 myoblasts transfected with both the HaloTag®-mock and HaloTag®-glut4 vectors (mock GIN− vs. mock GIN+, glut4 GIN− vs. glut4 GIN+), indicating that [6]-gingerol could promote glucose uptake even under the condition of GLUT4 overexpression.
Fig. 2.

Effect of [6]-gingerol on glucose uptake and glucose transporter 4 (GLUT4) translocation in cultured L6 myoblasts transfected with HaloTag-mock or HaloTag-glut4 vector and on ROS levels in RIN-5F cells. a Glucose uptake in L6 myoblasts transfected with HaloTag-mock or HaloTag-glut4 vector. L6 myoblasts (mock GIN+ and glut4 GIN+) were exposed to [6]-gingerol (50 µM) for 4 h. Each value represents the mean ± SEM for 4 wells. Values not sharing a common letter are significantly different at P < 0.05 by Tukey–Kramer multiple comparisons test. b GLUT4 translocation in L6 myoblasts transfected with HaloTag-glut4 vector. Immunocytochemistry was processed using anti-HaloTag and anti-caveolin-3 antibodies. c Basal ROS levels in RIN-5F cells and d AGE2-induced ROS level in RIN-5F cells. Each value represents the mean ± SEM for 3 wells. Values not sharing a common letter are significantly different at P < 0.05 by Tukey–Kramer multiple comparisons test
Bioimaging analysis of [6]-gingerol effect on GLUT4 translocation to plasma membrane
To visually observe the translocation of GLUT4 in cells, we employed L6 myoblasts transfected with HaloTag®-glut4 vector, because glucose uptake was higher in L6 myoblasts transfected with HaloTag®-glut4 vector than in those transfected with HaloTag®-mock vector (Fig. 2a). Caveolin-3 is involved in spatial and temporal regulation of GLUT4 translocation to plasma membrane and hence glucose uptake in skeletal muscle cells (Fecchi et al. 2006). Figure 2b (top, red) shows cellular localization of Halo-GLUT4 protein (glut4 GIN− and glut4 GIN+), Fig. 2b (center, green) shows cellular localization of caveolin 3, and Fig. 2b (bottom, yellow) shows their merging. In the absence of [6]-gingerol, Halo-GLUT4 protein and caveolin-3 were expressed in whole area and partially in the cell surface regions in L6 myoblasts transfected with HaloTag®-glut4 vector as shown in Fig. 2b (left, GIN−). [6]-Gingerol treatment at 50 µM for 30 min strengthened co-localization of GLUT4 protein and caveolin-3 in the plasma membrane compartment as shown by yellowish color in Fig. 2b (right, GIN+).
Changes of intracellular ROS levels in cultured RIN-5F cells by [6]-gingerol treatment
[6]-Gingerol dose-dependently and significantly reduced the basal ROS levels of RIN-5F cells (Fig. 2c) as compared with control cells (0 μM [6]-gingerol). As shown in Fig. 2d, artificial AGE2 was demonstrated to significantly and strikingly enhance the production of ROS in RIN-5F cells. However, [6]-gingerol significantly suppressed the AGE2-induced rise in the ROS level.
Effect of [6]-gingerol on fasting blood glucose levels and glucose intolerance in db/db mice
Based on the in vitro results, we examined whether or not [6]-gingerol feeding would improve glucose metabolism in db/db mice. As shown in Fig. 3a, the fasting blood glucose concentrations were significantly higher in control db/db mice (CNT) than in normal mice (NOR). However, [6]-gingerol treatment for 4 weeks significantly suppressed the rise (CNT vs. GIN). In IPGTT, [6]-gingerol significantly suppressed the rise in the blood glucose levels at 60 and 90 min after glucose administration as compared with the CNT group, suggesting that [6]-gingerol could improve glucose intolerance in db/db mice (Fig. 3b). Blood glucose concentrations of diabetic control groups in Fig. 3b (IPGTT) were lower than those in Fig. 3a. This difference might be due to a longer fasting period in Fig. 3b (15 h) than in Fig. 3a (3 h).
Fig. 3.
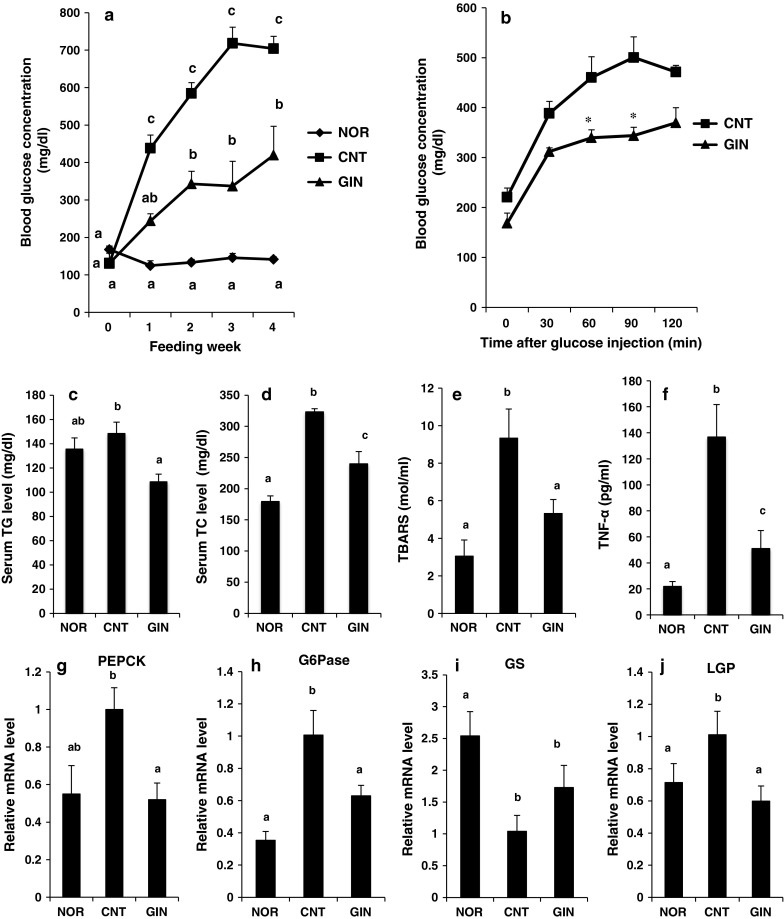
Effect of [6]-gingerol on fasting blood glucose levels, glucose intolerance, serum levels of lipids and TNF-α and on hepatic gene expression of enzymes related to glucose metabolism in db/db mice. a Fasting blood glucose level. During 4 weeks of [6]-gingerol feeding, fasting blood glucose levels were measured once a week, after mice were fasted for 3 h before blood collection. Each value represents the mean ± SEM for 6 mice. b IPGTT test after the determination of fasting blood glucose levels at 3rd week of feeding. After fasted for 15 h, mice of test group (GIN) were given orally [6]-gingerol at a dose of 5 mg/ml CMC/100 g body weight (n = 5), while those of diabetic control group (CNT) were given orally CMC solution alone at a volume of 1 ml/100 g body weight (n = 6). Asterisk means statistical significance at P < 0.05 by Student’s t test as compared with CNT group. c The levels of serum TG, d TC, e TBARS, and f TNF-α were measured after 4 weeks of [6]-gingerol feeding. The relative mRNA levels of g PEPCK, h G6Pase, i GS and j LGP in the liver were measured after 4 weeks of [6]-gingerol feeding. Each value represents the mean ± SEM for 6 mice (c–j). Values not sharing a common letter are significantly different at P < 0.05 by Tukey–Kramer multiple comparisons test. TG triglyceride, TC total cholesterol, TBARS thiobarbituric acid-reactive substances, TNF-α tumor necrosis factor-α, PEPCK phosphoenolpyruvate carboxykinase, G6Pase glucose-6-phosphatase, GS glycogen synthase, LGP liver glycogen phosphorylase
Effect of [6]-gingerol on serum levels of lipids and TNF-α in db/db mice
The serum TG, TC, TBARS and TNF-α levels significantly (TC, TBARS and TNF-α) increased or tended (TG) to increase in the CNT group as compared with those in the NOR group, whereas the treatment with [6]-gingerol significantly suppressed these rises (CNT vs. GIN in Fig. 3c–f).
Effect of [6]-gingerol on hepatic gene expression of enzymes related to glucose metabolism
Hepatic gene expression of phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase) increased in the CNT group when compared to the NOR group. Treatment of db/db mice with [6]-gingerol significantly suppressed these rises (Fig. 3g, h). Furthermore, the gene expression of glycogen synthase (GS) and liver glycogen phosphorylase (LGP) was significantly lower and higher, respectively, in the CNT group than in the NOR group. In contrast, [6]-gingerol treatment tended to increase the gene expression of GS and significantly suppressed that of LGP, respectively, as compared with control db/db mice (CNT vs. GIN in Fig. 3i, j).
Discussion
The majority of glucose uptake is accomplished in the skeletal muscle by the translocation of GLUT4 toward plasma membrane (Saltiel and Kahn 2001). In muscle cells, GLUT4 translocation is known to depend on two divergent signaling pathways: insulin signaling pathway and AMPK pathway (Imamura et al. 2003; Huang and Czech 2007). A molecule that promotes glucose uptake in muscle cells through GLUT4 translocation to plasma membrane via AMPK activation is considered to be capable of overcoming insulin resistance (Krook et al. 2004). Thus, we constructed a simple and rapid glucose uptake assay system with L6 myotubes (Kawano et al. 2009) which is comparable to that with soleus muscles of rats and [14C] 2-deoxyglucose (Nishitani et al. 2002). So far, we have found some food components which promote glucose uptake and GLUT4 translocation through AMPK activation in L6 myocytes and possess anti-hyperglycemic effect in type 2 diabetic model mice (Kawano et al. 2009; Minakawa et al. 2011, 2012; Ha et al. 2012; Son et al. 2013; Cheong et al. 2014). We have further found [6]-gingerol in the assay system as an antidiabetic candidate molecule in insulin resistant L6 myotubes. Pancreatic β-cells are known to be vulnerable to oxidative stress (Tiedge et al. 1997), leading to the reduction of insulin secretion. In the present study, we studied modes of stimulatory action of [6]-gingerol on glucose uptake in L6 myocytes and examined whether or not [6]-gingerol, an antioxidant (Dugasani et al. 2010), would protect RIN-5F pancreatic β-cells from oxidative stress. In addition, the effect of [6]-gingerol was studied at the whole body level using type 2 diabetic model db/db mice.
Glucose uptake promoted by [6]-gingerol was entirely eliminated by the addition of compound C. This result suggested that promotion of glucose uptake by [6]-gingerol might depend on the AMPK pathway. Indeed, [6]-gingerol could directly increase AMPK phosphorylation in L6 myotubes. Activation of AMPK has emerged as the critical event that upregulates GLUT4 translocation to the plasma membrane independently of insulin (Krook et al. 2004). Recently, [6]-gingerol has been reported to elevate intracellular Ca2+ concentration and subsequently induce an enhancement of phosphorylated AMPKα in L6 myotubes via modulation by Ca2+/calmodulin-dependent protein kinase kinase (CamKK) (Li et al. 2013), this being consistent with the present results. Park et al. (2012) recently show that resveratrol directly inhibits cAMP-dependent phosphodiesterases (PDEs), leading to elevated cAMP levels. The resulting activation of Epac1, a cAMP effector protein, increases intracellular Ca2+ levels and activates the CamKK-AMPK pathway via phospholipase C and the ryanodine receptor Ca2+-release channel. As a consequence, resveratrol increases AMPK phosphorylation. Thus, it seems worthy from the aspect of direct target identification to examine whether or not [6]-gingerol could inhibit PDEs.
As was expected from promoted AMPK phosphorylation (= activation) by [6]-gingerol, its treatment strengthened co-localization of GLUT4 protein and caveolin-3 in the plasma membrane compartment of L6 myoblasts transfected with HaloTag®-glut4 vector (Fig. 2b). These results visually verified that [6]-gingerol did promote GLUT4 translocation to plasma membrane in L6 myoblasts, as previously demonstrated for the first time in piceatannol-treated L6 myoblasts as well as L6 myotubes (Minakawa et al. 2012).
Hyperglycemia augments the accumulation of advanced glycation end products (AGEs). It is well recognized that AGEs produce ROS through their receptor and play a significant role in the development of diabetic complications (Wautier and Guillausseau 2001; Fiorentino et al. 2013). The serum level of AGEs is reported to be associated with insulin resistance even in non-obese, non-diabetic subjects (Tan et al. 2011). Therefore, reducing AGEs-induced ROS as well as the circulating AGEs appears to protect against the development of diabetes and its complications. We examined whether or not [6]-gingerol would protect β-cells from AGEs-induced ROS level using artificial AGE (AGE2). [6]-Gingerol was demonstrated to reduce the artificial AGE2-induced increase in the intracellular ROS level in RIN-5F cells (Fig. 2d). It has been suggested that AGEs can impair insulin sensitivity in skeletal muscle cells and adipocytes (Miele et al. 2003; Unoki et al. 2007). We only have demonstrated that [6]-gingerol has AGE2-induced ROS scavenging activity. Further intensive studies such as measurement of the circulating level of AGEs are required to learn if [6]-gingerol improves diabetic complications caused by AGEs.
[6]-Gingerol feeding to db/db mice was found to suppress the increase in fasting blood glucose concentrations (Fig. 3a). Food intake during the experimental period of 4 weeks was similar between diabetic control and [6]-gingerol-treated db/db mice (data not shown), indicating the suppressive effect of [6]-gingerol on the rise in fasting blood glucose concentrations was not due to reduced food consumption but due to its pharmacological action. [6]-Gingerol administration to db/db mice was also demonstrated to improve glucose intolerance in IPGTT (Fig. 3b). Oral glucose tolerance test (OGTT) has been reported to show similar results in the mice received [6]-gingerol (Singh et al. 2009), suggesting another possibility that [6]-gingerol may act on the intestinal enzyme and/or cells. Further studies are required to clarify this aspect.
Hyperglycemia is particularly related to hepatic glucose synthesis. Excessive hepatic gluconeogenesis is responsible for the high blood glucose level. Thus, suppression of gluconeogenesis in the liver is suggested as an intervention for type 2 diabetes. In the present experiment, [6]-gingerol suppressed the gene expression of hepatic enzymes related to gluconeogenesis (PEPCK, G6Pase) and glycogenolysis (LGP) and tended to enhance the gene expression of hepatic enzyme related to glycogenesis (GS) in db/db mice, like recently demonstrated in ob/ob mice treated with aspalathin (Son et al. 2013) and equol, a metabolite of daidzein (Cheong et al. 2014). Further intensive studies such as measurement of activities of above-mentioned hepatic enzymes related to glucose metabolism and identification of direct target of [6]-gingerol in hepatocytes are required to know precise mechanisms for antidiabetic action of [6]-gingerol.
The serum TG and TC levels increased in diabetic control group as compared with those in normal group. However, [6]-gingerol significantly suppressed these rises, indicating hypolipidemic action of [6]-gingerol, being consistent with other reports in diabetic rats and mice (Al-Amin et al. 2006; Singh et al. 2009). TBARS concentration is widely used as an indicator of lipid peroxidation process and oxidative stress (Bełtowski et al. 2000). [6]-Gingerol also suppressed the rise in the serum TBARS level in db/db mice. In the present study, [6]-gingerol is demonstrated to protect animals from oxidative stress at the whole body level as well as at the cellular level as above-mentioned. From these results, [6]-gingerol is demonstrated to possess its antioxidative function both in vitro and in vivo. Increased level of TNF-α has been reported to be associated with insulin resistance (Winkler et al. 2003). The serum TNF-α level was remarkably higher in diabetic control mice than in normal mice, while [6]-gingerol significantly suppressed this rise. This result suggests a possibility that [6]-gingerol may play a role in reducing insulin resistance in the obese diabetic state.
According to Reagan-Shaw et al. (2008), effective doses in humans are suggested to be one-eighth to one-twelfth of those in mice. Although necessary amounts of [6]-gingerol to apply its hypoglycemic effect in the human body have not been established yet, our research provided the importance and basis of its glucose-lowering effect for further researches in humans as well as animal models.
In summary, [6]-gingerol increased glucose uptake through promotion of GLUT4 translocation via AMPK activation in L6 myocytes. [6]-Gingerol could protect pancreatic β-cells from oxidative stress. The feeding of [6]-gingerol to db/db mice suppressed the increase in the fasting blood glucose level and improved glucose intolerance. [6]-Gingerol was demonstrated for the first time to regulate in vivo hepatic gene expression of enzymes related to glucose metabolism, toward a decrease in glucose production and an increase in glycogen synthesis, these changes contributing to the anti-hyperglycemic effect of [6]-gingerol. Thus, [6]-gingerol may provide therapeutic as well as preventive possibilities for type 2 diabetes.
Electronic supplementary material
Acknowledgments
This work was supported in part by the Japan Science Society and in part by the Asahi Group Foundation, Tokyo, Japan.
Conflict of interest
Authors declare that they have no conflict of interest.
Abbreviations
- AMPK
5′ Adenosine monophosphate-activated protein kinase
- GLUT4
Glucose transporter 4
- G6Pase
Glucose-6-phosphatase
- GS
Glycogen synthase
- LGP
Liver glycogen phosphorylase
- PEPCK
Phosphoenolpyruvate carboxykinase
- TBARS
Thiobarbituric acid-reactive substances
- TC
Total cholesterol
- TG
Triglyceride
References
- Al-Amin ZM, Thomson M, Al-Qattan KK, Peltonen-Shalaby R, Ali M. Anti-diabetic and hypolipidaemic properties of ginger (Zingiber officinale) in streptozotocin-induced diabetic rats. Br J Nutr. 2006;96:660–666. doi: 10.1079/BJN20061849. [DOI] [PubMed] [Google Scholar]
- Bełtowski J, Wójcicka G, Górny D, Marciniak A. The effect of dietary-induced obesity on lipid peroxidation, antioxidant enzymes and total plasma antioxidant capacity. J Physiol Pharmacol. 2000;51:883–896. [PubMed] [Google Scholar]
- Chakraborty D, Mukherjee A, Sikdar S, Paul A, Ghosh S, Khuda-Bukhsh AR. [6]-Gingerol isolated from ginger attenuates sodium arsenite induced oxidative stress and plays a corrective role in improving insulin signaling in mice. Toxicol Lett. 2012;210:34–43. doi: 10.1016/j.toxlet.2012.01.002. [DOI] [PubMed] [Google Scholar]
- Cheong SH, Furuhashi K, Ito K, Nagaoka M, Yonezawa T, Miura Y, Yagasaki K. Antihyperglycemic effect of equol, a daidzein derivative, in cultured L6 myocytes and ob/ob mice. Mol Nutr Food Res. 2014;58:267–277. doi: 10.1002/mnfr.201300272. [DOI] [PubMed] [Google Scholar]
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. doi: 10.1016/0003-2697(87)90021-2. [DOI] [PubMed] [Google Scholar]
- Del Prato S, Marchetti P. Targeting insulin resistance and beta-cell dysfunction: the role of thiazolidinediones. Diabetes Technol Ther. 2004;6:719–731. doi: 10.1089/dia.2004.6.719. [DOI] [PubMed] [Google Scholar]
- Dugasani S, Pichika MR, Nadarajah VD, Balijepalli MK, Tandra S, Korlakunta JN. Comparative antioxidant and anti-inflammatory effects of [6]-gingerol, [8]-gingerol, [10]-gingerol and [6]-shogaol. J Ethnopharmacol. 2010;127:515–520. doi: 10.1016/j.jep.2009.10.004. [DOI] [PubMed] [Google Scholar]
- Fecchi K, Volonte D, Hezel MP, Schmeck K, Galbiati F. Spatial and temporal regulation of GLUT4 translocation by flotillin-1 and caveolin-3 in skeletal muscle cells. FASEB J. 2006;20:705–707. doi: 10.1096/fj.05-4661fje. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiorentino TV, Prioletta A, Zuo P, Folli F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr Pharm Des. 2013;19:5695–5703. doi: 10.2174/1381612811319320005. [DOI] [PubMed] [Google Scholar]
- Ha BG, Nagaoka M, Yonezawa T, Tanabe R, Woo JT, Kato H, Chung UI, Yagasaki K. Regulatory mechanism for the stimulatory action of genistein on glucose uptake in vitro and in vivo. J Nutr Biochem. 2012;23:501–509. doi: 10.1016/j.jnutbio.2011.02.007. [DOI] [PubMed] [Google Scholar]
- Huang S, Czech MP. The GLUT4 glucose transporter. Cell Metab. 2007;5:237–252. doi: 10.1016/j.cmet.2007.03.006. [DOI] [PubMed] [Google Scholar]
- Huang C, Somwar R, Patel N, Niu W, Török D, Klip A. Sustained exposure of L6 myotubes to high glucose and insulin decreases insulin-stimulated GLUT4 translocation but upregulates GLUT4 activity. Diabetes. 2002;51:2090–2098. doi: 10.2337/diabetes.51.7.2090. [DOI] [PubMed] [Google Scholar]
- Imamura T, Huang J, Usui I, Satoh H, Bever J, Olefsky JM. Insulin-induced GLUT4 translocation involves protein kinase C-λ-mediated functional coupling between Rab4 and the motor protein kinesin. Mol Cell Biol. 2003;23:4892–4900. doi: 10.1128/MCB.23.14.4892-4900.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawano A, Nakamura H, Hata S, Minakawa M, Miura Y, Yagasaki K. Hypoglycemic effect of aspalathin, a rooibos tea component from Aspalathus linearis, intype 2 diabetic model db/db mice. Phytomedicine. 2009;16:437–443. doi: 10.1016/j.phymed.2008.11.009. [DOI] [PubMed] [Google Scholar]
- Krook A, Wallberg-Henriksson H, Zierath JR. Sending the signal: molecular mechanisms regulating glucose uptake. Med Sci Sports Exerc. 2004;36:1212–1217. doi: 10.1249/01.MSS.0000132387.25853.3B. [DOI] [PubMed] [Google Scholar]
- Kubra IR, Rao LJ. An impression on current developments in the technology, chemistry, and biological activities of ginger (Zingiber officinale Roscoe) Crit Rev Food Sci Nutr. 2012;52:651–688. doi: 10.1080/10408398.2010.505689. [DOI] [PubMed] [Google Scholar]
- Kume S, Kato S, Yamagishi S, Inagaki Y, Ueda S, Arima N, Okawa T, Kojiro M, Nagata K. Advanced glycation end-products attenuate human mesenchymal stem cells and prevent cognate differentiation into adipose tissue, cartilage, and bone. J Bone Miner Res. 2005;20:1647–1658. doi: 10.1359/JBMR.050514. [DOI] [PubMed] [Google Scholar]
- Li Y, Tran VH, Koolaji N, Duke C, Roufogalis BD. (S)-[6]-Gingerol enhances glucose uptake in L6 myotubes by activation of AMPK in response to [Ca2+]i. J Pharm Pharm Sci. 2013;16:304–312. doi: 10.18433/j34g7p. [DOI] [PubMed] [Google Scholar]
- Lin N, Zhang H, Su Q. Advanced glycation end-products induce injury to pancreatic beta cells through oxidative stress. Diabetes Metab. 2012;38:250–257. doi: 10.1016/j.diabet.2012.01.003. [DOI] [PubMed] [Google Scholar]
- Miele C, Riboulet A, Maitan MA, Oriente F, Romano C, Formisano P, Giudicelli J, Beguinot F, Van Obberghen E. Human glycated albumin affects glucose metabolism in L6 skeletal muscle cells by impairing insulin-induced insulin receptor substrate (IRS) signaling through a protein kinase C alpha-mediated mechanism. J Biol Chem. 2003;278:47376–47387. doi: 10.1074/jbc.M301088200. [DOI] [PubMed] [Google Scholar]
- Minakawa M, Kawano A, Miura Y, Yagasaki K. Hypoglycemic effect of resveratrol in type 2 diabetic model db/db mice and its actions in cultured L6 myotubes and RIN-5F pancreatic β-cells. J Clin Biochem Nutr. 2011;48:237–244. doi: 10.3164/jcbn.10-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minakawa M, Miura Y, Yagasaki K. Piceatannol, a resveratrol derivative, promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and suppresses blood glucose levels in type 2 diabetic model db/db mice. Biochem Biophys Res Commun. 2012;422:469–475. doi: 10.1016/j.bbrc.2012.05.017. [DOI] [PubMed] [Google Scholar]
- Nishitani S, Matsumura T, Fujitani S, Sonaka I, Miura Y, Yagasaki K. Leucine promotes glucose uptake in skeletal muscles of rats. Biochem Biophys Res Commun. 2002;299:693–696. doi: 10.1016/S0006-291X(02)02717-1. [DOI] [PubMed] [Google Scholar]
- Park SJ, Ahmad F, Philp A, Baar K, Williams T, Luo H, Ke H, Rehmann H, Taussig R, Brown AL, Kim MK, Beaven MA, Burgin AB, Manganiello V, Chung JH. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell. 2012;148:421–433. doi: 10.1016/j.cell.2012.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reagan-Shaw S, Nihal M, Ahmad N. Dose translation from animal to human studies revisited. FASEB J. 2008;22:659–661. doi: 10.1096/fj.07-9574LSF. [DOI] [PubMed] [Google Scholar]
- Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature. 2001;414:799–806. doi: 10.1038/414799a. [DOI] [PubMed] [Google Scholar]
- Singh AB, Akanksha Singh N, Maurya R, Srivastava AK. Anti-hyperglycaemic, lipid lowering and anti-oxidant properties of [6]-gingerol in db/db mice. Int J Med Med Sci. 2009;1:536–544. [Google Scholar]
- Son MJ, Minakawa M, Miura Y, Yagasaki K. Aspalathin improves hyperglycemia and glucose intolerance in obese diabetic ob/ob mice. Eur J Nutr. 2013;52:1607–1619. doi: 10.1007/s00394-012-0466-6. [DOI] [PubMed] [Google Scholar]
- Tan KC, Shiu SW, Wong Y, Tam X. Serum advanced glycation end products (AGEs) are associated with insulin resistance. Diabetes Metab Res Rev. 2011;27:488–492. doi: 10.1002/dmrr.1188. [DOI] [PubMed] [Google Scholar]
- Tiedge M, Lortz S, Drinkgern J, Lenzen S. Relation between antioxidant enzyme gene expression and antioxidative defense status of insulin-producing cells. Diabetes. 1997;46:1733–1742. doi: 10.2337/diab.46.11.1733. [DOI] [PubMed] [Google Scholar]
- Unoki H, Bujo H, Yamagishi S, Takeuchi M, Imaizumi T, Saito Y. Advanced glycation end products attenuate cellular insulin sensitivity by increasing the generation of intracellular reactive oxygen species in adipocytes. Diabetes Res Clin Pract. 2007;76:236–244. doi: 10.1016/j.diabres.2006.09.016. [DOI] [PubMed] [Google Scholar]
- Wautier JL, Guillausseau PJ. Advanced glycation end products, their receptors and diabetic angiopathy. Diabetes Metab. 2001;27:535–542. [PubMed] [Google Scholar]
- Winkler G, Kiss S, Keszthelyi L, Sápi Z, Ory I, Salamon F, Kovács M, Vargha P, Szekeres O, Speer G, Karádi I, Sikter M, Kaszás E, Dworak O, Gerö G, Cseh K. Expression of tumor necrosis factor (TNF)-α protein in the subcutaneous and visceral adipose tissue in correlation with adipocyte cell volume, serum TNF-α soluble serum TNF-receptor-2 concentrations and C-peptide level. Eur J Endocrinol. 2003;149:129–135. doi: 10.1530/eje.0.1490129. [DOI] [PubMed] [Google Scholar]
- Yagasaki K, Morisaki N, Kitahara Y, Miura A, Funabiki R. Involvement of proteinkinase C activation in l-leucine-induced stimulation of protein synthesis in L6 myotubes. Cytotechnology. 2003;43:97–103. doi: 10.1023/B:CYTO.0000039898.44839.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yagihashi S, Miura Y, Yagasaki K. Inhibitory effect of gingerol on the proliferation and invasion of hepatoma cells in culture. Cytotechnology. 2008;57:129–136. doi: 10.1007/s10616-008-9121-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou G, Myers R, Li Y, Chen Y, Shen X, Fenyk-Melody J, Wu M, Ventre J, Doebber T, Fujii N, Musi N, Hirshman MF, Goodyear LJ, Moller DE. Role of AMP-activated protein kinase in mechanism of metformin action. J Clin Invest. 2001;108:1167–1174. doi: 10.1172/JCI13505. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.