

Abstract
Activation of the immune system can impair cognitive function, particularly on hippocampus dependent tasks. Several factors such as normal aging and prenatal experiences can modify the severity of these cognitive deficits. One additional factor that may modulate the behavioral response to immune activation is obesity. Prior work has shown that obesity alters the activity of the immune system. Whether diet-induced obesity (DIO) influences the cognitive deficits associated with inflammation is currently unknown. The present study explored whether DIO alters the behavioral response to the bacterial endotoxin, lipopolysaccharide (LPS). Female C57BL/6J mice were fed a high-fat (60% fat) or control diet (10% fat) for a total of five months. After consuming their respective diets for four months, mice received an LPS or saline injection and were assessed for alterations in spatial learning. One month later, mice received a second injection of LPS or saline and tissue samples were collected to assess the inflammatory response within the periphery and central nervous system (CNS). Results showed that LPS administration impaired spatial learning in the control diet mice, but had no effect in DIO mice. This lack of a cognitive deficit in the DIO female mice is likely due to a blunted inflammatory response within the brain. While cytokine production within the periphery (i.e., plasma, adipose, and spleen) was similar between the DIO and control mice, the DIO mice failed to show an increase in IL-6 and CD74 in the brain following LPS administration. Collectively, these data indicate that DIO can reduce aspects of the neuroinflammatory response as well as blunt the behavioral reaction to an immune challenge.
Keywords: High-fat diet, LPS, CD74, CD86, spatial learning, water maze, cytokines, IL-6
1. INTRODUCTION
A wealth of evidence indicates the obesity alters immune function [1–4]. These alterations in immune activity may contribute to the increased incidence of several diseases in obese individuals including cardiovascular disease and Alzheimer’s disease [5–7]. In addition, obesity is associated with a higher rate of infection following surgery or an injury [2, 8]. Further research has shown that obese individuals require a longer recovery time after surgery [8]. During influenza season, obese individuals have higher rates of hospitalization than normal weight individuals [9]. Given the substantial increase in the number of obese individuals, understanding the nature of obesity-related changes in immune function and their potential relevance to disease and infection susceptibility is of great importance.
Investigations using an animal model of diet-induced obesity (DIO) confirm that obesity modifies the response to an immune challenge. For instance, DIO has been shown to increase peripheral (i.e., plasma, spleen, and adipose tissue) levels of the cytokines interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and IL-1 receptor antagonist (IL-1ra) in response to the endotoxin, lipopolysaccharide (LPS) [3, 4, 10, 11]. Further, DIO rats show higher expression of IL-6 and TNF-α in the hypothalamus following LPS administration when compared to control rats [3]. In contrast to these data, several reports have shown that DIO can suppress aspects of the response to an immune challenge. For example, Lawerence et al [12] reported that DIO mice show reduced neural activation, as measured by the immediate early gene c-Fos, following LPS administration in select brain regions. In addition, macrophages from DIO animals show reduced interleukin-1β (IL-1β), TNF-α, nuclear factor-κB (NF-κB), Toll-like receptor-4 (TLR-4) and inducible nitric oxide synthase (iNOS) levels following an immune challenge when compared to macrophages from control diet animals [13–16]. In addition, we found attenuated levels of IL-6 in the cortex of DIO males mice following LPS administration [17]. Presently, the reason for these divergent findings is unknown, but variations in the experimental procedure including differences in the animal’s age, species, timing of tissue sampling, as well as the type and duration of the diet, among other factors, likely contribute to these differences. Despite these differences, the data collectively indicate the DIO alters the response to immune activation.
The body’s response to an immune challenge includes a complementary shift in behavior. This behavioral response, commonly known as sickness behavior, includes a reduction in locomotor activity, alterations in sleep, depression of social and sexual behavior, anorexia, fever, as well as suppression of other species typical behaviors [18–20]. Induction of sickness behavior is mediated by the activity of several inflammatory molecules including prostaglandins and the proinflammatory cytokines IL-1β, IL-6, and TNF-α [18, 20, 21]. Prior work indicates that obesity, through alterations in immune function, may exacerbate the sickness behavior response. For example, Pohl et al. [22] report that rats fed a high-fat diet showed prolonged suppression of social behavior following LPS administration. However, DIO has no effect on LPS-induced reductions in locomotor behavior, as DIO animals show a similar reduction and recovery rate when compared to control animals [17, 22]. Alterations in the fever response have also been observed, as some report an enhanced fever in DIO animals following an immune challenge [3, 22]. However, this appears to be dose dependent as Lawrence et al. [12] report a blunted fever in DIO mice following a low LPS dose and an exaggerated fever following a higher dose. Taken together, these data indicate that DIO modifies aspects of the sickness behavioral response to an immune challenge.
Beyond the development of sickness behavior, immune activation can disrupt cognitive function. Processes dependent on the hippocampus are particularly sensitive to disruption following an immune challenge. For instance, LPS administration impairs contextual, but not auditory, fear conditioning [23, 24]. In addition, immune activation has been shown to disrupt spatial learning in the water maze [25–27]. Similar to sickness behavior, these cognitive deficits result from proinflammatory cytokines acting within the brain, as cytokine administration mimics the deficits induced by LPS and inhibition of IL-1β and IL-6 have been shown to block memory deficits following infection [28–32]. There has been some suggestion that the cognitive deficits associated with DIO may reflect basal increases in inflammation, however, this has not been consistently reported and may depend on the age of the animal [17, 33, 34]. While data exists on how DIO may alter the sickness behavior response to an immune challenge, whether DIO alters the development of cytokine-induced cognitive deficits is presently unknown. The objective of the current study was to assess alterations in spatial learning in DIO and control mice following an LPS challenge along with corresponding changes in the systemic and central inflammatory response.
2. MATERIAL AND METHODS
2.1. Animals
Subjects were 52 female C57BL/6J mice bred in the University of North Carolina Wilmington (UNCW) animal facility, with breeding stock obtained from The Jackson Laboratory (Bar Harbor, Maine). Mice were 5 weeks old at the beginning of the study. Animals were treated in compliance with the Guide for the Care and Use of Laboratory Animals and the experiments were conducted in accordance with a protocol approved by the Institutional Animal Care and Use Committee (IACUC) at UNCW.
2.2. Diets and housing
Mice were assigned to the high-fat diet (DIO; Open source diets, D12492) or control diet (Open source diets, D12450B). The high-fat diet contained 60% of calories from fat and the control diet contained 10% of calories from fat. Mice were group housed with two to four mice per cage. Mice started consuming the high-fat or control diet at 5 weeks of age and continued for 5 months until the mice were approximately 6 months old. Mice were weighed once a week throughout the experiment as well as assessed for weight loss following administration of LPS or saline (described below). The treatment-induced changes in total body weight (i.e., grams lost or gained) was determined by calculating a difference score for each mouse by subtracting the animal’s body weight on the day of the injection from the animal’s body weight twenty-four hours after an LPS or saline injection.
2.3. Effect of DIO on LPS-induced spatial learning deficits
After consuming the high-fat or control diet for four months mice received a single intraperitoneal (ip) injection of LPS (250 μg/kg, obtained from E. coli, serotype 0111:B4; Sigma, St. Louis, MO) or saline. Four hours later, mice were tested in the water maze to assess spatial learning. The maze consisted of a circular tub (121 cm diameter) and a white circular platform (12.7 cm). The platform was submerged 1 cm under the surface of the water. The water was made opaque with white tempera paint to conceal the platform. Water temperature was maintained at 19° ± 1° C throughout testing. Extra-maze cues were located around the maze. Mice received three trials (up to 60 sec) per day from different start locations for five consecutive days. If a mouse failed to locate the platform within the 60 sec they were gently guided to the platform. All mice remained on the platform for 10 seconds at the end of each trial. A video tracking system (Topscan, Clever Systems, Reston, VA) was used to measure distance swam (mm), latency to locate the platform (sec), and swim speed (mm/sec). A single 60 sec probe trial was conducted approximately two hours after the subject’s last trial on day 5. The platform was removed and the number of times the animal crossed the original location of the platform as well as percent time spent in the target quadrant that contained the platform during testing was recorded by the tracking system.
2.4. Effect of DIO on LPS-induced cytokine expression
One month following behavioral testing mice received a second ip injection of LPS (250 μg/kg) or saline. Mice received the same treatment as they received during the behavioral testing. Four hours after LPS or saline administration mice were sacrificed and tissue was collected to assess changes in gene expression via quantitative real-time reverse-transcription polymerase chain reaction (qRT-PCR) and protein levels of proinflammatory cytokines via ELISA.
2.4.1. Tissue collection
Mice were sacrificed by rapid decapitation. Blood was collected in heparin-treated vacutainer tubes (Becton-Dickinson, Rutherford, NJ), centrifuged (2000 rpm for 30 min at 4°C), and plasma collected and stored at −70°C until assayed. Hippocampal samples were dissected on a chilled glass dish and immediately placed into RNAlater solution (Qiagen, Valencia, CA) and stored at −20°C until RNA isolation. The remaining brain (i.e., everything, but the hippocampus) was placed in a separate tube and snap frozen. Additionally, the spleen and gonadal adipose pads from the abdominal cavity were collected and snap frozen for analysis with ELISA and qRT-PCR, respectively.
2.4.2. qRT-PCR
Hippocampal samples were sonicated and RNA purified by the RNeasy Mini kit (Qiagen, Valencia, CA), then quantified and assessed for purity using a Gen5 Epoch spectrophotometer (BioTek Instruments, Highland Park, VT). All hippocampal samples had a 260/280 ratio of 1.99 or above. Conversion of RNA into cDNA was completed by following the instructions of the High-capacity cDNA reverse transcription kit that included an RNase inhibitor step (Life Technologies) using the following cycling conditions: 10 min at 25°C, 120 min at 37°C and 5 min at 85°C. After reverse transcription, cDNA was held at 2–8° C in a refrigerator overnight before conducting two-step qRT-PCR to determine the amount of specific mRNA transcript present in each sample. Hippocampal samples were assessed for expression of IL-1β (Mm00434228_m1), IL-6 (Mm00446190_m1), CD74 (Mm00658576_m1) and CD86 (Mm00444543_m1). β-actin (Mm00607939_s1) served as the endogenous control gene. RNA was also extracted from adipose samples and analyzed for expression levels of IL-6 and IL-1β normalized against β-actin levels. Each sample was run in triplicate for each gene in a 10 μl reaction that contained 80 ng of cDNA and 0.5 μl of a 20X probe/primer mix in one 384 well plate. The amount of specific mRNA present was determined by utilizing TaqMan™ probe and primer chemistry (Applied Biosystems, Foster City, CA) specifically designed to bind to reverse-transcribed cDNA of the genes of interest using an Applied Biosystems Viia7 PCR instrument (Applied Biosystems, Foster City, CA) using the following cycle parameters: 2 min at 50°C, 10 min at 95o, followed by 40 cycles of 15 sec at 95oC and 1 min at 60oC. Florescence data (ΔRn) was exported from the Applied Biosystems SDS software and analyzed by DART (Data Analysis for RT-PCR) [35]. Gene expression data were normalized by dividing the Ro (florescent intensity) values of the target genes by the Ro values of the endogenous control gene β-actin. Levels of β-actin expression did not differ across groups.
2.4.3. ELISA
Brain and spleen samples were homogenized in a 1mM solution of the proteinase inhibitor phenylmethylsulfonyl fluoride (PMSF in 0.1M phosphate buffer) and centrifuged for 30 min at 3500 rpm at 4° C. Spleen samples were assayed for protein levels of IL-1β (BD Biosciences, San Diego, CA; Detection limits 15.6 to 1000 pg/ml) and IL-6 (BD Biosciences, San Diego, CA; Detection limits 15.6 to 1000 pg/ml) with ELISA kits. Brain and plasma samples were assayed for IL-6. Total protein levels were determined in each sample by a protein assay (Pierce Thermo Scientific, Rockford, IL). Spleen and brain ELISA results are expressed as picograms (pg) of cytokine protein per milligram (mg) of total protein. Plasma results are expressed as pg of cytokine protein per milliliter.
2.5. Statistical analyses
Body weight data were analyzed by repeated measures ANOVA with diet (DIO or control diet) and week as the within-subject factor. Water maze data were analyzed by repeated measures ANOVA with diet (DIO or control diet) and treatment (LPS or saline) as the between-subject factors and test day (days 1–5) as the within-subject factor. Probe trial water maze data, brain cytokine ELISA data, plasma cytokine data, splenic cytokine data, gene expression data, and change in body weight following treatment were analyzed by two-way ANOVA with diet (DIO or control diet) and treatment (LPS or saline) as the between-subject variables. When appropriate, Fisher’s least significant difference (LSD) was used as a post hoc test to determine significant differences between groups. A p<0.05 was considered statistically significant.
3. RESULTS
3.1. Body weight
There was a significant main effect of diet and a diet x week interaction (F(1, 48)=125.42, p<0.0001; F(20, 980)=100.31, p<0.0001, respectively, see Figure 1A) for body weight. Results show that DIO and control mice had similar weights at the start of the study. While both groups gained weight throughout the experiment the DIO mice (average weight 42.1 grams, range of 28–57 grams) weighted more than the control mice (average weight 24.2 grams, range of 20–29 grams). In response to LPS administration, a significant main effect of treatment (F(1, 48)=90.03, p<0.0001, see Figure 1B) showed that both the DIO and control mice lost weight when compared the saline-treated mice. There was no difference between the DIO and control diet mice in terms of the grams lost 24 hours after LPS administration, as both the DIO and control diet mice lost a similar amount of weight.
Figure 1.
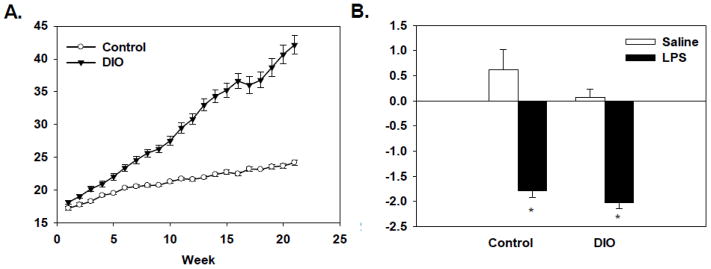
Data are shown as mean body weight in grams ± standard error of the means (SEMs) across the experiment in weeks (A). There was no difference in body weight initially, but the DIO mice gained significantly more weight than control mice throughout the experiment (A). LPS administration caused a significant reduction in body weight relative to saline-treated mice 24 hours after treatment (B). There was no difference in the LPS-induced weight loss between the DIO and control mice. * indicates a significant difference from diet-matched saline-treated mice.
3.2. Water maze
For distance swam (i.e., path length) there was a significant diet x treatment x test day interaction (F(4, 192)=2.94, p<0.05). Post hoc testing showed that for mice fed the control diet, LPS-treated mice swam a shorter distance on day 1 and longer distance on days 4 and 5 of testing when compared with the control mice treated with saline (p<0.05, see Figure 2A). For the DIO mice, LPS reduced distance swam on day 1 (p<0.05), but no other differences were detected (see Figure 2A). No differences in distance swam were detected between the saline-treated control and DIO mice. Analysis of the latency to locate the platform showed significant main effects of treatment and test day (F(1, 48)=7.45, p<0.005; F(4, 192)= 55.12, p<0.0001; see Figure 2B). Post hoc analysis revealed that LPS-treated control diet mice had longer latencies to locate the platform on days 4 and 5 of testing compared to saline-treated control mice (p<0.05). For the DIO mice, LPS administration had no effect on latency to locate the platform, as they did not differ from saline-treated DIO mice on any of the test days. No differences in latency to locate the platform were detected between the saline-treated control and DIO mice. For swim speed there was a significant main effect of diet (F(1, 48)=4.701, p<0.05) that showed the DIO mice swam slower than the control mice. In addition, a significant treatment x day interaction (F(4, 192)=4.963, p<0.05, see Figure 2C) for swim speed showed that LPS-treated mice swam slower than saline-treated mice on day 1 (p<0.05). Analysis of the number of platform crossings and percent time spent in the target quadrant during the probe trial showed no significant diet or treatment differences (data not shown).
Figure 2.
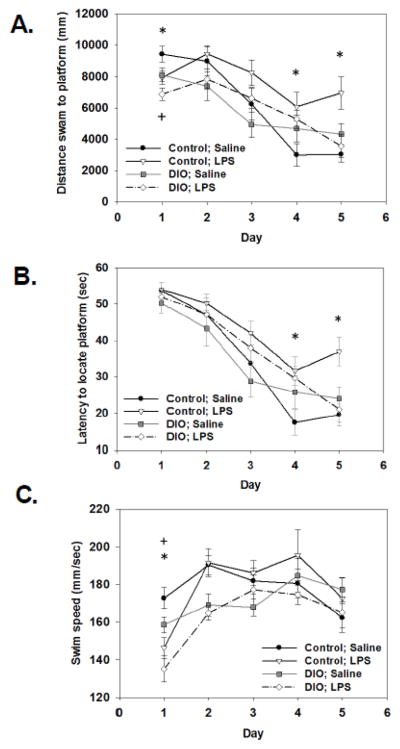
In the control diet mice, LPS administration 4 hours prior to day 1 of testing reduced distance swam on day 1 and increased distance swam to locate the platform on days 4 and 5 of testing (A). The DIO LPS-treated mice swam a shorter distance on day 1 relative to saline-treated DIO mice, but had no effect during later days of testing (A). LPS increased mean latency to locate the platform on days 4 and 5 in control, but not DIO, mice (B). Overall DIO mice swam slower than control mice (C). LPS administration significantly decreased swim speed in both the DIO and control diet mice on day 1 of testing compared to saline-treated mice (C). Lines represent group averages of three trials per test day ±SEMs. * indicates significant difference between control diet LPS- and saline-treated mice. + indicates significant difference between DIO LPS- and saline-treated mice.
3.3. Hippocampus RNA data
Significant main effects of treatment for hippocampal expression of IL-1β, IL-6, and CD86 (F(1, 48)=31.43, p<0.0001; F(1, 48)= 25.22, p<0.0001; F(1, 48)=11.21, p<0.005, respectively, see Figures 3B, 3C, and 3D) showed that LPS administration, regardless of diet, increased expression of IL-1β, IL-6 and CD86 in the hippocampus relative to saline-treated mice. For CD74 expression there was a significant diet x treatment interaction (F(1, 48)=4.25, p<0.05, see Figure 3A). Post hoc testing showed that LPS administration increased CD74 expression in the control mice (p<0.05), but had no effect in the DIO mice. However, DIO saline-treated mice had higher expression of CD74 relative to saline-treated control mice (p<0.05).
Figure 3.

DIO mice showed higher basal expression of CD74 in the hippocampus (A). In response to LPS control mice showed the expected increase in CD74 expression, but the DIO mice failed to show a response to LPS (A). Administration of LPS increased hippocampal expression of CD86 (B), IL-6 (C), and IL-1β (D), but diet did not influence this response. Bars represent means ± SEMs. + indicates a significant difference from the saline-treated control diet mice. * indicates a significant difference from diet-matched saline-treated mice.
3.4. Brain IL-6 protein data
Analysis of protein levels of IL-6 in the brain samples revealed a significant diet x treatment interaction (F(1, 48)= 6.90, p<0.05, see Figure 4). LPS administration increased brain levels of IL-6 in the control mice (p<0.05). However, LPS administration had no effect on IL-6 levels in the DIO mice as LPS- and saline-treated DIO mice did not differ from one another.
Figure 4.

LPS increased protein levels of IL-6 in brain samples 4 hours after treatment in the control diet mice, but not in the DIO mice. Bars represent means ± SEMs. * indicates a significant difference from diet-matched saline-treated mice.
3.5. Adipose RNA data
Significant main effects of treatment for adipose expression of IL-6 and IL-1β (F(1, 48)=17.94, p<0.0001; F(1, 48)= 14.35, p<0.005, respectively, see Figures 5A and 5B) showed that LPS administration, regardless of diet, increased expression of IL-6 and IL-1β in the adipose tissue relative to saline-treated mice.
Figure 5.
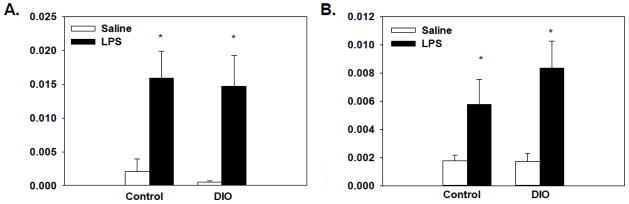
Expression of IL-6 (A) and IL-1β (B) in adipose tissue was increased 4 hours after LPS administration as compared to saline treated mice. No differences or interactions were found with diet. Bars represent means ± SEMs. * indicates a significant difference from diet-matched saline-treated mice.
3.6. Plasma IL-6 protein data
There was a significant main effect of treatment for plasma levels of IL-6 (F(1, 48)=46.86, p<0.0001, see Figure 6A) that showed LPS administration, regardless of diet, increased levels of IL-6 compared to saline-treated mice.
Figure 6.
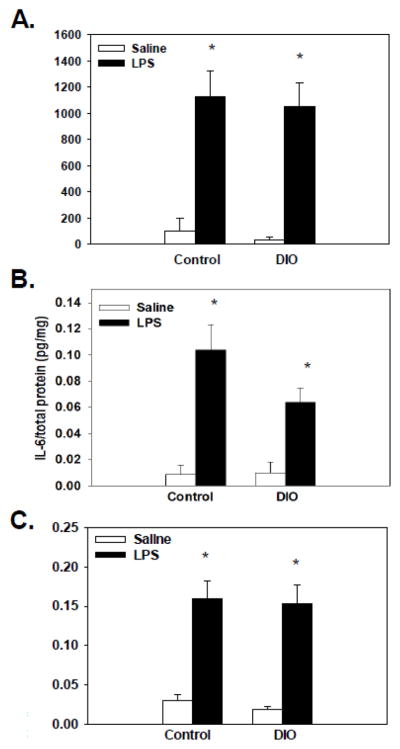
LPS increased plasma levels of IL-6 (A), regardless of diet, when compared to saline-treated mice. Splenic protein levels of IL-6 (B) and IL-1β (C) were increased following LPS administration as compared to saline. No significant differences or interactions were found with diet. Bars represent means ± SEMs. * indicates a significant difference from diet-matched saline-treated mice.
3.7. Spleen protein data
Significant main effects of treatment for splenic levels of IL-6 and IL-1β (F(1, 48)= 35.96, p<0.0001; F(1, 48)=53.99, p<0.0001, respectively, see Figures 6B and 6C) showed that LPS administration, regardless of diet, increased protein levels of IL-6 and IL-1β in the spleen relative to saline-treated mice.
4. DISCUSSION
Results from the present study indicate that DIO female mice show a differential behavioral response to an immune challenge compared to control diet female mice. While the control mice showed impaired spatial learning in the water maze following LPS administration, the DIO mice showed no impairments in acquiring the task following LPS administration. The lack of a cognitive deficit following LPS administration in the DIO mice is likely due to attenuation of select aspects of the neuroinflammatory response, as DIO mice failed to show an increase in IL-6 and CD74 in response to LPS. On the surface the lack of cytokine-induced cognitive deficits may be viewed as a beneficial response, however, these behavioral data in conjunction with the physiological measurements clearly indicate that DIO alters the immune system’s ability to adequately respond to an immune challenge.
The present study is the first to assess alterations in cognitive function in DIO mice following an immune challenge. In agreement with prior reports, we observed that LPS administration impaired spatial learning in the control diet mice [26, 27], as the LPS-treated controls swam significantly longer path lengths to locate the hidden platform during the later days of testing. However, we found no difference in path length or latency to locate the platform between the LPS- and saline-treated DIO mice. The lack of a difference is not simply the result of impaired learning in the DIO group as both the LPS- and saline-treated DIO mice showed reductions in path length across the five days of testing, indicating the DIO mice acquired the task. To date, the existing literature on the effects of DIO on the behavioral response to an immune challenge has been limited to sickness behavior and has shown mixed results. DIO was reported to prolong the LPS-induced suppression of social behavior in rats, but had no effect on the LPS-induced reductions in locomotor behavior [17, 22]. In agreement we found that LPS administration reduced swim speed on day 1 of testing in both the DIO and control diet mice. Prior work has shown that very low doses of LPS can inhibit activity [20]. While the DIO mice show a similar performance deficit following LPS administration, they showed no evidence of impaired acquisition, potentially indicating that the underlying inflammatory mediators that induce the cognitive and performance deficits may differ or that there is a difference in the sensitivity to express sickness behavior versus cognitive impairments. Though more work is needed to fully characterize the impact of DIO on the behavioral response to immune activation, our data provide novel evidence that the cognitive deficits following an immune challenge are blunted in DIO mice.
Attenuated levels of IL-6 may contribute to the absence of cognitive deficits following immune activation in the DIO mice. In humans, increased levels of IL-6 are associated with a greater incidence of cognitive decline [36]. Further, animal research has shown that inhibiting IL-6 prevents the expression of LPS-induced cognitive deficits [31, 32]. For example, Sparkman et al. [32] found that wild type mice show impairments in working memory following LPS administration, but these deficits were absent in IL-6 knockout mice. In agreement, we found attenuated levels of IL-6 in the brains of DIO mice who failed to show cognitive deficits following LPS administration. While it is unlikely that IL-6 completely mediates the cognitive deficits that result from an immune challenge, as other cytokines such as IL-1β are likely involved, the present data in conjunction with prior reports emphasizes the role of IL-6 in impairing cognitive function [28, 29, 31, 32, 36].
Prior work has shown that DIO can impair cognitive performance in the absence of any additional manipulations [33, 34, 37, 38]. In the present study, we found no evidence that DIO impaired spatial learning, as we found no difference in acquisition between the saline-treated control and DIO female mice. Both DIO and control saline-treated mice acquired the task, as evidenced by a significant reduction in path length and latency across the five days of testing. These data appear to contrast with numerous prior reports that show DIO disrupts spatial learning [33, 37, 38], including our prior work in male mice using the same procedure [17]. However, the lack of a spatial learning deficit in the DIO mice in the present study is likely due to the use of female mice. Prior work has shown that male and female mice respond differently to DIO. For example, Gallou-Kabani [39] reported that while both male and female C57BL/6J mice show symptoms of type 2 diabetes when fed a high-fat diet the changes were less pronounced in the female mice. Further, DIO was reported to decrease hippocampal neurogenesis in males, but not females [40]. Similarly, high-fat diet consumption impaired long-term potentiation in male mice, but had no effect in female mice [41]. Analysis of diet-induced changes in hippocampal gene expression via a microarray revealed that consumption of a high-fat diet altered expression of several genes in both male and female mice [42]. However, there were no common genes altered by high-fat diet consumption when males and females are compared, indicating a differential response to high-fat diet exists between the sexes [42]. Most relevant to the present data is the work of Hwang et al. [41] who reports that high-fat diet consumption impaired acquisition of a passive avoidance task in male, but not female, mice. The current data are in agreement with these findings, as we report female DIO mice do not develop deficits in cognitive function. Taken together, the data indicate that while DIO induces physiological changes in both male and female mice, male mice appear more susceptible to the effects of DIO particularly in regards to impairments in cognitive function and measures of neural plasticity.
Understanding the nature of the immune alterations induced by obesity is a necessary step to help reduce the increased risk of infection as well as improve recovery following infection. In the current study, DIO mice showed an array of changes in immune activity, particularly in the brain, when compared to the control diet mice. We did not observe a simple enhancement or suppression of immune activity, but rather a combination of basal increases as well as a blunted reaction to an immune challenge when compared to the control diet mice. Specifically, we found that within the periphery DIO had no effect on basal or LPS-induced cytokine levels when assessed in plasma, spleen, and adipose tissue, as both DIO and control mice showed a similar increase in IL-6 and IL-1β following LPS exposure. In the CNS we found that DIO increased basal expression of CD74, a molecule involved in trafficking major histocompatibility complex II (MHC II) proteins, compared to control diet mice. The MHC II protein is known to play a central role in cellular communication and initiation of the innate immune response [43]. However, in response to an immune challenge the DIO mice failed to show an increase in CD74 expression. Given the basal difference, the lack of an increase in CD74 following LPS administration in the DIO mice may result from higher basal expression. In addition, we observed a deficit in IL-6 production in the brains of DIO mice following LPS administration. These findings are in agreement with prior data that show DIO can attenuate aspects of the inflammatory response [12, 13, 16, 17]. For instance, Lawerence et al. [12] report that DIO mice show dampened neural activation in select brain regions following LPS administration when compared to control mice. Further, we reported attenuated IL-6 production in the brain of DIO mice following LPS administration [17]. However, others report exaggerated cytokine production following an immune challenge. For example, Pohl et al. [3] report higher levels of IL-6 and IL-1β in the hypothalamus of DIO rats following LPS administration. A recent report found DIO increased LPS-induced IL-1β and TNF-α expression in the hippocampus of juvenile, but not adult, mice [34]. These data indicate that DIO modulates the neuroinflammatory response to an immune challenge, but that the nature of these changes may be affected by several factors including age, species, and the brain region assessed.
Taken together the current findings indicate that obesity does not simply exaggerate or inhibit immune function, but rather induces a more complex dysregulation in immune activity. This contention is supported by a recent report by Tilton et al. [1] who observed that DIO mice showed a unique gene expression profile in lung tissue following inhalation of LPS when assessed via a microarray. While many genes showed a similar response to LPS exposure in the obese and control mice there were over 3,000 genes that were differentially regulated in the obese mice that did not change in the control mice. In addition, they report that DIO mice showed a 50% increase in the number of macrophages in the lungs in response to LPS inhalation when compared to control mice, but lower levels of IL-6 and TNF-α were produced in the DIO mice [1]. Collectively, these data indicate that DIO alters immune function, but depending on the immune parameter assessed results may show suppression or enhancement of immune activity.
Collectively, the current study demonstrates that DIO modifies immune function and attenuates the response to immune activation. DIO had the greatest impact on immune activity within the brain. Further, the data indicate that DIO female mice fail to show cognitive deficits following LPS administration that likely relates to alterations in the neuroinflammatory response.
Highlights.
Diet-induced obesity blunts the cognitive deficits associated with inflammation.
Diet-induced obesity attenuates LPS-induced interleukin-6 production in brain.
Basal expression of CD74 in the hippocampus is increased in diet-induced obese mice.
Acknowledgments
The authors would like to thank our funding sources for their support. This work was supported by grants from National Institute on Aging R00AG040194 and UNCW Charles Cahill awarded to R.A.K. Funding sources had no involvement in the experimental design or interpretation of the results.
Footnotes
Conflict of interests: All authors declare that there are no conflicts of interest
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Sharay E. Setti, Email: ses8535@uncw.edu.
Alyssa M. Littlefield, Email: aml8440@uncw.edu.
Samantha W. Johnson, Email: swj4531@uncw.edu.
Rachel A. Kohman, Email: kohmanr@uncw.edu.
References
- 1.Tilton SC, Waters KM, Karin NJ, Webb-Robertson BJ, Zangar RC, Lee KM, Bigelow DJ, Pounds JG, Corley RA. Diet-induced obesity reprograms the inflammatory response of the murine lung to inhaled endotoxin. Toxicol Appl Pharmacol. 2013;267(2):137–48. doi: 10.1016/j.taap.2012.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vilar-Compte D, Mohar A, Sandoval S, de la Rosa M, Gordillo P, Volkow P. Surgical site infections at the National Cancer Institute in Mexico: a case-control study. Am J Infect Control. 2000;28(1):14–20. doi: 10.1016/s0196-6553(00)90006-3. [DOI] [PubMed] [Google Scholar]
- 3.Pohl J, Woodside B, Luheshi GN. Changes in hypothalamically mediated acute-phase inflammatory responses to lipopolysaccharide in diet-induced obese rats. Endocrinology. 2009;150(11):4901–10. doi: 10.1210/en.2009-0526. [DOI] [PubMed] [Google Scholar]
- 4.Kim MS, Choi MS, Han SN. High fat diet-induced obesity leads to proinflammatory response associated with higher expression of NOD2 protein. Nutr Res Pract. 2011;5(3):219–23. doi: 10.4162/nrp.2011.5.3.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hubert HB, Feinleib M, McNamara PM, Castelli WP. Obesity as an independent risk factor for cardiovascular disease: a 26-year follow-up of participants in the Framingham Heart Study. Circulation. 1983;67(5):968–77. doi: 10.1161/01.cir.67.5.968. [DOI] [PubMed] [Google Scholar]
- 6.Whitmer RA, Gustafson DR, Barrett-Connor E, Haan MN, Gunderson EP, Yaffe K. Central obesity and increased risk of dementia more than three decades later. Neurology. 2008;71(14):1057–64. doi: 10.1212/01.wnl.0000306313.89165.ef. [DOI] [PubMed] [Google Scholar]
- 7.Calabro P, Golia E, Maddaloni V, Malvezzi M, Casillo B, Marotta C, Calabro R, Golino P. Adipose tissue-mediated inflammation: the missing link between obesity and cardiovascular disease? Intern Emerg Med. 2009;4(1):25–34. doi: 10.1007/s11739-008-0207-2. [DOI] [PubMed] [Google Scholar]
- 8.Falagas ME, Kompoti M. Obesity and infection. Lancet Infect Dis. 2006;6(7):438–46. doi: 10.1016/S1473-3099(06)70523-0. [DOI] [PubMed] [Google Scholar]
- 9.Kwong JC, Campitelli MA, Rosella LC. Obesity and respiratory hospitalizations during influenza seasons in Ontario, Canada: a cohort study. Clin Infect Dis. 2011;53(5):413–21. doi: 10.1093/cid/cir442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Huang H, Liu T, Rose JL, Stevens RL, Hoyt DG. Sensitivity of mice to lipopolysaccharide is increased by a high saturated fat and cholesterol diet. J Inflamm (Lond) 2007;4:22. doi: 10.1186/1476-9255-4-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lavin DN, Joesting JJ, Chiu GS, Moon ML, Meng J, Dilger RN, Freund GG. Fasting induces an anti-inflammatory effect on the neuroimmune system which a high-fat diet prevents. Obesity (Silver Spring) 2011;19(8):1586–94. doi: 10.1038/oby.2011.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lawrence CB, Brough D, Knight EM. Obese mice exhibit an altered behavioural and inflammatory response to lipopolysaccharide. Dis Model Mech. 2012;5(5):649–59. doi: 10.1242/dmm.009068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen S, Lin G, Lei L, You X, Wu C, Xu W, Huang M, Luo L, Wang Z, Li Y, Zhao X, Yan F. Hyperlipidemia modifies innate immune responses to lipopolysaccharide via the TLR-NF-kappaB signaling pathway. Inflammation. 2013;36(4):968–76. doi: 10.1007/s10753-013-9628-9. [DOI] [PubMed] [Google Scholar]
- 14.Amar S, Zhou Q, Shaik-Dasthagirisaheb Y, Leeman S. Diet-induced obesity in mice causes changes in immune responses and bone loss manifested by bacterial challenge. Proc Natl Acad Sci U S A. 2007;104(51):20466–71. doi: 10.1073/pnas.0710335105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhou Q, Leeman SE, Amar S. Signaling mechanisms involved in altered function of macrophages from diet-induced obese mice affect immune responses. Proc Natl Acad Sci U S A. 2009;106(26):10740–5. doi: 10.1073/pnas.0904412106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Borges MC, Vinolo MA, Crisma AR, Fock RA, Borelli P, Tirapegui J, Curi R, Rogero MM. High-fat diet blunts activation of the nuclear factor-kappaB signaling pathway in lipopolysaccharide-stimulated peritoneal macrophages of Wistar rats. Nutrition. 2013;29(2):443–9. doi: 10.1016/j.nut.2012.06.008. [DOI] [PubMed] [Google Scholar]
- 17.Baumgarner KM, Setti S, Diaz C, Littlefield A, Jones A, Kohman RA. Diet-induced obesity attenuates cytokine production following an immune challenge. Behav Brain Res. 2014;267:33–41. doi: 10.1016/j.bbr.2014.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dantzer R. Cytokine-induced sickness behaviour: a neuroimmune response to activation of innate immunity. Eur J Pharmacol. 2004;500(1–3):399–411. doi: 10.1016/j.ejphar.2004.07.040. [DOI] [PubMed] [Google Scholar]
- 19.Hart BL. Biological basis of the behavior of sick animals. Neurosci Biobehav Rev. 1988;12(2):123–37. doi: 10.1016/s0149-7634(88)80004-6. [DOI] [PubMed] [Google Scholar]
- 20.Teeling JL, Felton LM, Deacon RM, Cunningham C, Rawlins JN, Perry VH. Sub-pyrogenic systemic inflammation impacts on brain and behavior, independent of cytokines. Brain Behav Immun. 2007;21(6):836–50. doi: 10.1016/j.bbi.2007.01.012. [DOI] [PubMed] [Google Scholar]
- 21.Bluthé RM, Dantzer R, Kelley KW. Effects of interleukin-1 receptor antagonist on the behavioral effects of lipopolysaccharide in rat. Brain Res. 1992;573(2):318–20. doi: 10.1016/0006-8993(92)90779-9. [DOI] [PubMed] [Google Scholar]
- 22.Pohl J, Sheppard M, Luheshi GN, Woodside B. Diet-induced weight gain produces a graded increase in behavioral responses to an acute immune challenge. Brain Behav Immun. 2014;35:43–50. doi: 10.1016/j.bbi.2013.09.002. [DOI] [PubMed] [Google Scholar]
- 23.Pugh CR, Kumagawa K, Fleshner M, Watkins LR, Maier SF, Rudy JW. Selective effects of peripheral lipopolysaccharide administration on contextual and auditory-cue fear conditioning. Brain Behav Immun. 1998;12(3):212–29. doi: 10.1006/brbi.1998.0524. [DOI] [PubMed] [Google Scholar]
- 24.Kranjac D, McLinden KA, Deodati LE, Papini MR, Chumley MJ, Boehm GW. Peripheral bacterial endotoxin administration triggers both memory consolidation and reconsolidation deficits in mice. Brain Behav Immun. 2012;26(1):109–21. doi: 10.1016/j.bbi.2011.08.005. [DOI] [PubMed] [Google Scholar]
- 25.Gibertini M. IL1 beta impairs relational but not procedural rodent learning in a water maze task. Adv Exp Med Biol. 1996;402:207–17. doi: 10.1007/978-1-4613-0407-4_27. [DOI] [PubMed] [Google Scholar]
- 26.Sparkman NL, Kohman RA, Scott VJ, Boehm GW. Bacterial endotoxin-induced behavioral alterations in two variations of the Morris water maze. Physiol Behav. 2005;86(1–2):244–51. doi: 10.1016/j.physbeh.2005.07.016. [DOI] [PubMed] [Google Scholar]
- 27.Shaw KN, Commins S, O’Mara SM. Lipopolysaccharide causes deficits in spatial learning in the watermaze but not in BDNF expression in the rat dentate gyrus. Behav Brain Res. 2001;124(1):47–54. doi: 10.1016/s0166-4328(01)00232-7. [DOI] [PubMed] [Google Scholar]
- 28.Palin K, Bluthe RM, Verrier D, Tridon V, Dantzer R, Lestage J. Interleukin-1beta mediates the memory impairment associated with a delayed type hypersensitivity response to bacillus Calmette-Guerin in the rat hippocampus. Brain Behav Immun. 2004;18(3):223–30. doi: 10.1016/j.bbi.2003.09.002. [DOI] [PubMed] [Google Scholar]
- 29.Barrientos RM, Higgins EA, Sprunger DB, Watkins LR, Rudy JW, Maier SF. Memory for context is impaired by a post context exposure injection of interleukin-1 beta into dorsal hippocampus. Behav Brain Res. 2002;134(1–2):291–8. doi: 10.1016/s0166-4328(02)00043-8. [DOI] [PubMed] [Google Scholar]
- 30.Frank MG, Barrientos RM, Hein AM, Biedenkapp JC, Watkins LR, Maier SF. IL-1RA blocks E. coli-induced suppression of Arc and long-term memory in aged F344xBN F1 rats. Brain Behav Immun. 2010;24(2):254–62. doi: 10.1016/j.bbi.2009.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Burton MD, Johnson RW. Interleukin-6 trans-signaling in the senescent mouse brain is involved in infection-related deficits in contextual fear conditioning. Brain Behav Immun. 2012;26(5):732–8. doi: 10.1016/j.bbi.2011.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sparkman NL, Buchanan JB, Heyen JR, Chen J, Beverly JL, Johnson RW. Interleukin-6 facilitates lipopolysaccharide-induced disruption in working memory and expression of other proinflammatory cytokines in hippocampal neuronal cell layers. J Neurosci. 2006;26(42):10709–16. doi: 10.1523/JNEUROSCI.3376-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Pistell PJ, Morrison CD, Gupta S, Knight AG, Keller JN, Ingram DK, Bruce-Keller AJ. Cognitive impairment following high fat diet consumption is associated with brain inflammation. J Neuroimmunol. 2010;219(1–2):25–32. doi: 10.1016/j.jneuroim.2009.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Boitard C, Cavaroc A, Sauvant J, Aubert A, Castanon N, Laye S, Ferreira G. Impairment of hippocampal-dependent memory induced by juvenile high-fat diet intake is associated with enhanced hippocampal inflammation in rats. Brain Behav Immun. 2014 doi: 10.1016/j.bbi.2014.03.005. [DOI] [PubMed] [Google Scholar]
- 35.Peirson SN, Butler JN, Foster RG. Experimental validation of novel and conventional approaches to quantitative real-time PCR data analysis. Nucleic Acids Res. 2003;31(14):e73. doi: 10.1093/nar/gng073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Weaver JD, Huang MH, Albert M, Harris T, Rowe JW, Seeman TE. Interleukin-6 and risk of cognitive decline: MacArthur studies of successful aging. Neurology. 2002;59(3):371–8. doi: 10.1212/wnl.59.3.371. [DOI] [PubMed] [Google Scholar]
- 37.Molteni R, Wu A, Vaynman S, Ying Z, Barnard RJ, Gomez-Pinilla F. Exercise reverses the harmful effects of consumption of a high-fat diet on synaptic and behavioral plasticity associated to the action of brain-derived neurotrophic factor. Neuroscience. 2004;123(2):429–40. doi: 10.1016/j.neuroscience.2003.09.020. [DOI] [PubMed] [Google Scholar]
- 38.Molteni R, Barnard RJ, Ying Z, Roberts CK, Gomez-Pinilla F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience. 2002;112(4):803–14. doi: 10.1016/s0306-4522(02)00123-9. [DOI] [PubMed] [Google Scholar]
- 39.Gallou-Kabani C, Vige A, Gross MS, Rabes JP, Boileau C, Larue-Achagiotis C, Tome D, Jais JP, Junien C. C57BL/6J and A/J mice fed a high-fat diet delineate components of metabolic syndrome. Obesity (Silver Spring) 2007;15(8):1996–2005. doi: 10.1038/oby.2007.238. [DOI] [PubMed] [Google Scholar]
- 40.Lindqvist A, Mohapel P, Bouter B, Frielingsdorf H, Pizzo D, Brundin P, Erlanson-Albertsson C. High-fat diet impairs hippocampal neurogenesis in male rats. Eur J Neurol. 2006;13(12):1385–8. doi: 10.1111/j.1468-1331.2006.01500.x. [DOI] [PubMed] [Google Scholar]
- 41.Hwang LL, Wang CH, Li TL, Chang SD, Lin LC, Chen CP, Chen CT, Liang KC, Ho IK, Yang WS, Chiou LC. Sex differences in high-fat diet-induced obesity, metabolic alterations and learning, and synaptic plasticity deficits in mice. Obesity (Silver Spring) 2010;18(3):463–9. doi: 10.1038/oby.2009.273. [DOI] [PubMed] [Google Scholar]
- 42.Martin B, Pearson M, Brenneman R, Golden E, Keselman A, Iyun T, Carlson OD, Egan JM, Becker KG, Wood W, 3rd, Prabhu V, de Cabo R, Maudsley S, Mattson MP. Conserved and differential effects of dietary energy intake on the hippocampal transcriptomes of females and males. PLoS One. 2008;3(6):e2398. doi: 10.1371/journal.pone.0002398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Beswick EJ, V, Reyes E. CD74 in antigen presentation, inflammation, and cancers of the gastrointestinal tract. World J Gastroenterol. 2009;15(23):2855–61. doi: 10.3748/wjg.15.2855. [DOI] [PMC free article] [PubMed] [Google Scholar]