

Abstract
Background
Inorganic nitrate () is a precursor of nitric oxide (NO) in the body and a large number of short-term studies with dietary nitrate supplementation in animals and humans show beneficial effects on cardiovascular health, exercise efficiency, host defense and ischemia reperfusion injury. In contrast, there is a long withstanding concern regarding the putative adverse effects of chronic nitrate exposure related to cancer and adverse hormonal effects. To address these concerns we performed in mice, a physiological and biochemical multi-analysis on the effects of long-term dietary nitrate supplementation.
Design
7 week-old C57BL/6 mice were put on a low-nitrate chow and at 20 weeks-old were treated with NaNO3 (1 mmol/L) or NaCl (1 mmol/L, control) in the drinking water. The groups were monitored for weight gain, food and water consumption, blood pressure, glucose metabolism, body composition and oxygen consumption until one group was reduced to eight animals due to death or illness. At that point remaining animals were sacrificed and blood and tissues were analyzed with respect to metabolism, cardiovascular function, inflammation, and oxidative stress.
Results
Animals were supplemented for 17 months before final sacrifice. Body composition, oxygen consumption, blood pressure, glucose tolerance were measured during the experiment, and vascular reactivity and muscle mitochondrial efficiency measured at the end of the experiment with no differences identified between groups. Nitrate supplementation was associated with improved insulin response, decreased plasma IL-10 and a trend towards improved survival.
Conclusions
Long term dietary nitrate in mice, at levels similar to the upper intake range in the western society, is not detrimental.
Keywords: Nitrate, Toxicity, Long-term, Supplementation, Treatment, Survival
Abbreviations: ACh, acetylcholine; ADMA, asymmetric dimethylarginine; AU, arbitrary units; AUC, area under the curve; BP, blood pressure; cGMP, cyclic guanidine monophosphate; DeXA, Dual energy X-ray absorptiometry; eNOS, endothelial nitric oxide synthase; HOMA-IR, homeostatic model assessment of insulin resistance; IL-6, interleukin-6; IL-10, interleukin-10; iNOS, inducible nitric oxide synthase; IPGTT, intraperitoneal glucose tolerance test; IPITT, intraperitoneal insulin tolerance test; MAP, Mean arterial pressure; NADPH, nicotinamide adenine dinucleotide phosphate; NO, nitric oxide; NOS, nitric oxide synthase; NaCl, sodium chloride; NaNO3, sodium nitrate; , nitrate; PE, phenylephrine; SDMA, symmetric dimethylarginine; SEM, standard error of measure; SNP, sodium nitroprusside; T3, triiodothyronine; T4, thyroxine
Graphical abstract
Highlights
-
•
Long term dietary nitrate supplementation for 17 months in mice.
-
•
Nitrate treatment in the upper range in the western society diet, has no adverse health effects.
-
•
Chronic nitrate intake in mice improves fasting insulin and insulin response.
-
•
Cardiovascular and inflammatory parameters were unchanged after long-term dietary nitrate treatment.
Introduction
Dietary intake of inorganic nitrate has for more than half a century been considered potentially harmful due to its bioconversion to nitrite and carcinogenic nitrosamines [1,2]. After the discovery of nitric oxide (NO) as a biological signaling molecule nitrate and nitrite have been used as stable surrogate markers of NO synthase-derived NO [3]. More recently an alternative pathway of NO generation was described in which serial reduction of nitrate and nitrite to NO and other bioactive nitrogen intermediates occurs [4]. This NO synthase (NOS) independent pathway relies on retention of nitrate anions through salivary concentration from the plasma. Nitrate is reduced to nitrite by oral commensal bacteria, swallowed, absorbed and further reduced to NO and other bioactive nitrogen oxides in blood and tissues [5,6]. A number of studies have demonstrated robust NO-like bioactivity after ingestion of nitrate. These include a reduction in blood pressure [7,8], inhibition of platelet aggregation [8,9], improved vascular function [10] and increased mitochondrial efficiency [11]. The crucial involvement of oral bacteria in nitrate bioactivation is evident from experiments using an antibacterial mouthwash, or if the test persons avoid swallowing of saliva after nitrate ingestion. Under these conditions the nitrate effects are greatly reduced or lost [8,12–14]. A paradox regarding the proposed harmful effects of dietary nitrate is the fact that vegetables are the greatest source of this anion. Long term longitudinal studies have specifically identified green leafy vegetable intake, which is the dominant nitrate source, as inversely proportional to the risk of diabetes in aging females and cardiovascular disease in males [15,16].
The recent studies showing beneficial cardiovascular and metabolic effects of dietary nitrate in animal models and humans have been short term or acute. Other long term dietary studies have been performed investigating the putative toxicity of nitrate and nitrite using high doses of these anions [17–21]. This study aimed at determining the long term effects of dietary nitrate intake in mice at doses similar to what can be achieved in humans by a diet rich in vegetables. During nitrate or matched sodium chloride supplementation of approximately 18 months, we looked at the parameters for metabolic, cardiovascular and inflammatory differences.
Materials and methods
Animals
All animal procedures in this study were in strict adherence to the Guide for the Care and Use of Laboratory Animals as adopted by the U.S. National Institutes of Health and were approved by the Regional Animal Care and Use Committee of Stockholm, Sweden. Six week-old male C57Bl/6 mice (n=27) were obtained from Charles River (Sulzfeld, Germany). After 1 week acclimatization the animals were fed low nitrate chow (0.06 µg/g NO3, D06041501DW, Research Diets, New Brunswick, NJ, USA) and housed in rooms at 22 °C with a 12/12 day-night cycle. After 3 months, the animals were separated into similar weight groups of 14 and 13 with 1 mM sodium chloride (NaCl) or 1 mM sodium nitrate (NaNO3) respectively in the drinking water. Over this period, weight and food and water uptake were monitored (Fig. 1). The health of the animals was monitored using an institutional physiological checklist and if behavior resulted in a combined score of 0.4 the animal was sacrificed and blood and organs collected. Seven animals were sacrificed due to poor health and four animals died of natural causes during the experiment.
Fig. 1.
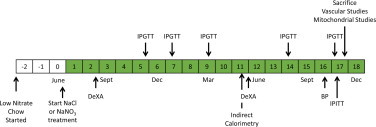
Experimental timeline. Diagram of the experimental procedures on the mice over the course of the experiment. The green squares indicate period of supplementation. NaCl, sodium chloride; NaNO3, sodium nitrate; DeXA, dual energy X-ray absorptiometry; IPGTT, intraperitoneal glucose tolerance test; BP, blood pressure; IPITT, intraperitoneal insulin tolerance test.
Materials
All chemicals not specified were obtained from Sigma-Aldrich at the highest grade available.
Body composition
Body composition by dual energy X-ray absorptiometry (DeXA) was measured under isoflurane anesthesia using the PIXImus imager (General Electric, Niskayuna, NJ, USA) according to manufacturer's instructions [22].
Intraperitoneal glucose tolerance test
Animals were starved for 6 h, separated individually, fasting blood glucose recorded (Freestyle Lite, Abbott Diabetes Care, Alameda, CA, USA). Following an intraperitoneal injection of d-glucose (2 g/kg BW) blood glucose was measured at 15, 30, 60 and 120 min [22].
Intraperitoneal insulin tolerance test
Under non-fasting conditions the mice were separated and initial blood glucose measurement was taken from the tail, an intraperitoneal insulin load (0.75 IE kg−1) was administered and blood glucose was measured at 15, 30, 45, 60, 75, 90, and 120 min [22].
Blood pressure
Blood pressure was monitored using the Coda High Throughput Noninvasive Tail Monitoring System (Kent Scientific, Torrington, CT, USA) following the manufacturer's protocol. Systolic, diastolic and mean arterial pressure measurements were collected over 5 consecutive days and averaged [22].
Indirect calorimetry
Individual mouse oxygen consumption was measured using an INCA indirect calorimeter (Somedic, Horby, Sweden). Animals were placed in the equipment between 5 pm and 6 pm and underwent 36 h at 24 °C and 24 h at the thermoneutral 34 °C maintaining a 12/12 light dark cycle consistent with animal room timing.
Mitochondrial isolation
After euthanizing the mouse, the soleus muscle was extracted, weighed and immediately added to 1 mL of ice-cold isolation medium: 100 mM sucrose, 100 mM KCl, 50 mM Trizma–HCl, 1 mM KH2PO4, 0.1 mM EGTA and 0.2% BSA (weight). The sample was homogenized with a pair of scissors (≈5 min) and left for sedimentation. The supernatant was removed and the sample was washed by adding new isolation medium again followed by sedimentation. This procedure was repeated twice whereupon isolation medium containing bacterial protease (0.2 mg/mL) was added and the sample was left on ice for 30 s followed by low intensity vortexing during 30 s. After repeating the procedure once more, the sample was transferred to a Potter-Elvehjem homogenizer and further homogenized at 125 rpm during approximately 2 min. The sample was then transferred to a falcon-tube pre-filled with 3 mL of isolation medium and centrifuged at 700g for 10 min (4 °C). The supernatant was collected and further centrifuged at 10,000g during 10 min. The buffy coat of extramitochondrial debris was carefully washed away with isolation medium followed by resuspension in isolation medium. After a new centrifugation step at 7000g for 5 min, the pellet was further washed and resuspended in mitochondrial preservation medium (0.6 µl/mg tissue sample) containing 0.5 mM EGTA, 3 mM MgCl2, 60 mM K-lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose, 1 g/L BSA, 20 mM histidine, 20 µM vitamin E succinate, 3 mM glutathione, 1 µM leupeptin, 2 mM glutamate, 2 mM malate and 2 mM Mg–ATP.
Mitochondrial respirometry
All mitochondrial experiments were performed at 37 °C in a 2-channel respirometer (Oroboros Oxygraph O2-K; Oroboros Instruments, Innsbruck, Austria). Respiration medium contained 0.5 mM EGTA, 3 mM MgCl2, 60 mM K-lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose and 1 mg/mL BSA. Mitochondrial substrates used for complex I respiration were pyruvate (5 mM) and malate (2 mM). Stepwise titration of carbonylcyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP) was performed in order to measure maximal uncoupled respiration. The produced adenosine triphosphate per consumed oxygen (P/O-ratio) was determined at 10 kPa in the presence of 2 mM ATP. Adenosine diphosphate was infused with a microinjection pump (Tip2K; Oroboros Instruments) at nonsaturating concentrations to produce a respiration rate corresponding to approximately half maximal state 3 respiration. The ADP infusion rate was divided by the oxygen consumption in order to calculate the P/O-ratio. Corrections were made for back diffusion of oxygen into the respiration chamber, oxygen dissolved in the infused ADP solution and oxygen consumption by the oxygen electrode.
Vascular reactivity
Isolation and dissection of carotids were performed as described previously [23]. In brief, bilateral carotid arteries were isolated and placed in physiological salt solution (PSS, composition in mM: 119 NaCl, 4.7 KCl, 1.6 CaCl2, 1.2 KH2PO4, 1.2 MgSO4, 25.1 NaHCO3, 5.5 glucose, 0.026 EDTA). Arteries were dissected and mounted in myograph chambers (model 620M, Danish Myo Technology, Denmark), and isometric tension was recorded (Powerlab 4/30, A D instruments, Australia). After mounting, vessels were equilibrated for 20 min in PSS bubbled with carbogen (95% O2; 5% CO2) at 37 °C, pH 7.4. Resting tension of the arteries was set in accordance with Mulvany's normalization procedure [24]. Following a second equilibration, 0.1 mol/L KCl was applied to assess arterial ring viability. Carotid arteries were then assessed for their dose response to acetylcholine (ACh, 10−9–10−4 mol/L), sodium nitroprusside (SNP, 10−9–10−4 mol/L), and 30 min contractile response to the non-selective NO-synthase inhibitor Nω-nitro-l-arginine methyl ester (L-NAME, 100 µM) after preconstriction with phenylephrine (PE, 0.1 or 1 µM). Arteries were washed three times and allowed to stabilize between experimental segments.
Plasma markers
Insulin ELISAs were purchased from Mercodia (Uppsala, Sweden). Multiplex proinflammatory 7 cytokine kit and custom made metabolic kit (Glucagon, active GLP-1, Insulin, and Leptin) were purchased from Mesoscale Discoveries (Rockville, MD, USA). T3 and T4 ELISA kits were purchased from Calbiotech (Spring Valley, CA, USA). cGMP EIA was purchased from GE Healthcare. All kits were run according to manufacturers' instructions. Plasma nitrite and nitrate and urinary nitrate were analyzed by HPLC (ENO-20) and autosampler (840, EiCom, Kyoto, Japan). Plasma was extracted using methanol (1:2) then centrifuged for 10 min 4 °C 10g. Urine samples were initially diluted (1:50) using carrier solution containing 10% methanol. Nitrate and nitrite were separated by reverse phase/ion exchange chromatography followed by nitrate reduction to nitrite by cadmium and reduced copper. The nitrite was then derivatized using Griess reagent to form diazo compounds and analyzed by detection at 540 nm.
Plasma amino acids
Urea cycle amino acids and methyl-arginines were analyzed as previously described with a few modifications [25]. Briefly, after thawing samples on ice, 25 µl of plasma were crashed with 225 µl of 0.2% formic acid in isopropanol containing the internal standard (0.73 µM of N4-Arginine). Afterwards samples were vortexed for 30 s and centrifuged at 8000g for 10 min. Finally 5 µl of the supernatant was analyzed by liquid chromatography tandem mass spectrometry (LC–MS/MS). Separation was performed on an ACQUITY UPLC System from Waters Corporation (Milford, MA, USA) using an Atlantis HILIC Silica 3 µm (150×2.1 mm2) column from Waters. Mobile phases consisted of 0.2% formic acid in ACN:MeOH (75:25, v/v) and 0.2% formic acid in water. The flowrate was set at 400 µl/min. The chromatographic gradient was set as follows: 0 min, 100% A; time range 0→8 min, 100→60% A (linear decrease); time range 8–9 min, 60→50% A (linear decrease); time range 9→9.8 min 50% A (isocratic range), time range 9.8→10.0, 50→100% A (linear increase). The column was then allowed to equilibrate at 100% A for 3 min. Detection was performed using a Waters Xevo® TQ triple quadrupole equipped with an Electrospray Ion Source working in positive mode. For the quantification, the following SRM transitions were used: Arginine (175.1→70.1), Ornithine (133.1→70.1), Citrulline (176.1→70.1), ADMA (203.1→46.0), SDMA (203.1→172.1) and N4-Arginine (179.1→71.1).
NADPH oxidase activity
Segments of the heart, liver and kidney were cut from frozen tissue and homogenized in PBS using a Bullet Blender (Next Advance, Inc. Averill Park, NY, USA) following the manufacturer's protocol. Particulate was centrifuged, supernatant was collected and diluted, and then 50 mM of lucigenin and 0.1 mM NADPH added. Superoxide production was measured every 3 s for 3 min with the AutoLumat LB953 Multi-Tube Luminometer (Berthold Technologies, Bad Wildbad, Germany) [22].
Statistics
Comparison between groups was calculated by student's t-test. For comparison of 2 groups over time a 2-way ANOVA followed by Bonferroni post-hoc test was used. Survival was calculated using Log-rank test. All statistical calculations were made using Graphpad Prism (La Jolla, CA, USA). A p value<0.05 was considered significant. All results are shown as mean±SEM.
Results
Metabolic changes
Weight and body composition
Past publications have found that nitrate intake inhibited weight gain [20,26–28]. In contrast with these findings we found no differences in average weight or weight gain over the 17 months of nitrate supplementation when compared to control (Fig. 2A). Only animals that survived until final sacrifice were included in this weight comparison. Food and water consumption, measured periodically over the length of the study, were similar between groups (not shown). There were no differences in fat and lean body composition between groups when measured at 2, 12, and 17 months of supplementation (Fig. 2B). At final sacrifice, there were no significant differences in the weight of the heart, liver, pancreas, kidney, and gonadal adipose tissue (Table 1). One mouse in each group had hepatomegaly when they were sacrificed due to poor health.
Fig. 2.
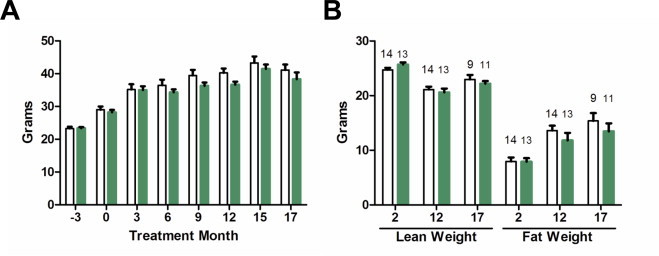
Weight and body composition. Average weight (A) and lean and fat body composition (B) at different time points over the course of the study. Control represented by open bars and nitrate by green bars. Average weight consists of the 8 animals per group. In the body composition graph the numbers above the bars indicate animals per group. Values are means ± SEM.
Table 1.
Final body weight and organ weight.
| Control (8) | Nitrate (8) | p-Value | |
|---|---|---|---|
| Final body weight | 38.46 ± 1.76 | 36.48 ± 1.99 | 0.469 |
| Heart | 0.205 ± 0.00 | 0.213 ± 0.00 | 0.310 |
| Liver | 1.993 ± 0.14 | 2.017 ± 0.20 | 0.924 |
| Pancreas | 0.175 ± 0.01 | 0.185 ± 0.01 | 0.576 |
| Kidney | 0.540 ± 0.02 | 0.534 ± 0.02 | 0.835 |
| Gonadal fat | 1.432 ± 0.17 | 1.382 ± 0.20 | 0.854 |
Glucose and insulin tolerance tests
Short-term dietary nitrate has previously been shown to improve glucose metabolism in endothelial NO synthase (eNOS) deficient mice with features of metabolic syndrome [29]. To see if there were any differences in glucose handling, intraperitoneal glucose tolerance tests (IPGTT) were run periodically throughout the experiment. There were no differences in the area under the curve of IPGTTs performed at 5, 7, 10, 14 and 17 months of treatment (Fig. 3A) (5 month results shown in Fig. 3B). This held true for the fasting glucose levels (Control 7.34±0.14 mmol/L vs Nitrate 7.45±0.14 mmol/L; p=0.563). An intraperitoneal insulin tolerance test at 17 months of treatment showed improved insulin responsiveness in the nitrate treated group as measured by percent change from basal glucose level (Fig. 3C) and area under the curve of the individual tests (Fig. 3D).
Fig. 3.
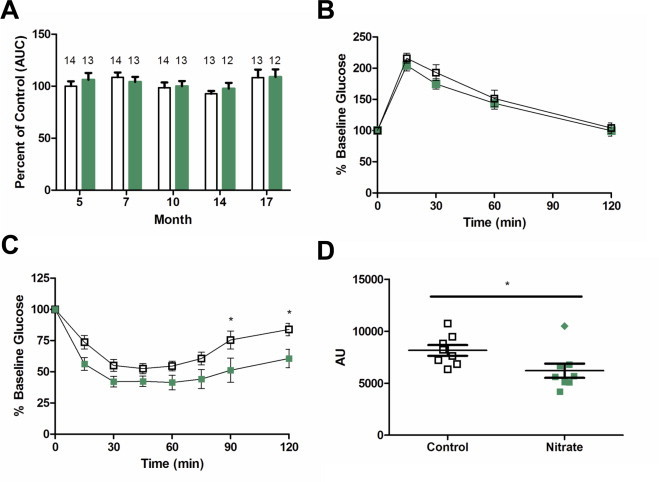
Glucose and insulin tolerance tests. Area under the curve (AUC) of the intraperitoneal glucose tolerance test (IPGTT) as a percentage of the 5 month IPGTT in control animals (A), and group average IPGTT curve at 5 month of supplementation (B). Intraperitoneal insulin tolerance test (IPITT) as percent of basal glucose (C) and area under the curve for each of the individual mice measured in arbitrary units (AU) (D). Control represented by open bars or symbols and nitrate by green bars or symbols. Values are means ± SEM. ⁎p<0.05.
Metabolic markers in plasma
The improved insulin tolerance test suggested changes in hormone levels, so we looked at plasma hormone levels related to metabolism by multiplex ELISA. There were no differences in plasma leptin, glucagon, active GLP-1 and insulin levels (Table 2). Closer look at the individual results indicated one obese nitrate animal skewed the insulin results away from significance (Fig. 4A). This mouse had insulin resistance as identified in the nitrate treated animals by the diamond in the individual IPITT AUC (Fig. 3D), fasting insulin level (Fig. 4A) and the HOMA-IR figures (Fig. 4B). Statistical calculation without this animal led to significantly decreased insulin and HOMA insulin resistance score in the dietary nitrate group.
Table 2.
Plasma hormone markers.
| Control (8) | Nitrate (8) | p-Value | |
|---|---|---|---|
| Leptin (pg/L) | 7.542 ± 1.55 | 8.399 ± 2.99 | 0.803 |
| Glucagon (pg/mL) | 187.9 ± 29.5 | 208.3 ± 43.2 | 0.702 |
| GLP-1 (pg/mL) | 28.72 ± 3.49 | 33.42 ± 7.77 | 0.589 |
| Insulin (pg/L) | 3.881 ± 0.61 | 2.627 ± 0.76 | 0.222 |
| Total T3 (nM) | 2.712 ± 0.12 | 2.199 ± 0.22 | 0.060 |
| Total T4 (nM) | 56.85 ± 3.75 | 54.98 ± 2.54 | 0.648 |
| T3/T4 Ratio | 0.048 ± 0.002 | 0.040 ± 0.003 | 0.052 |
Fig. 4.
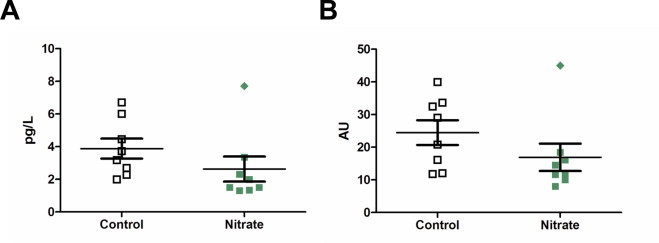
Plasma insulin and HOMA. Plasma insulin (A) and HOMA score for insulin resistance (B) calculated from blood glucose and insulin level at sacrifice and shown as arbitrary units (AU). Values are means ± SEM.
Prior research had identified nitrate intake as down regulating plasma levels of thyroid hormones thyroxine (T4) and triiodothyronine (T3) [20,26,30]. Our experiment found total T3 trended to being decreased in the nitrate treated group and this remained after calculating the ratio of T3 to T4 (Table 2).
Oxygen consumption
Research in humans has shown that acute nitrate intake reduces resting metabolic rate and oxygen cost during exercise, and these effects are coupled to improved skeletal muscle mitochondrial efficiency [11,31,32]. Indirect calorimetry was measured in a subset of animals at 11 months of treatment to see if there were changes in oxygen consumption under both ambient and thermoneutral conditions. Thermoneutral conditions remove the requirement for the animals to expend energy maintaining their body temperature. There were no significant differences in oxygen consumption, respiratory quotient, or delta oxygen under normal (24 °C, data not shown) or thermoneutral (34 °C) conditions (Table 3). At sacrifice, soleus muscle was isolated from the mice and mitochondrial efficiency was measured. Longterm nitrate treatment did not change the respiratory control ratio, the P/O ratio, leak, state 3, or 4 or uncoupled respiration (Table 4).
Table 3.
Metabolic cages: thermoneutral.
| Control (6) | Nitrate (5) | p-Value | |
|---|---|---|---|
| Inactive period | |||
| Oxygen consumption | 20.52±1.779 | 20.00±1.828 | 0.843 |
| Respiratory quotient | 0.726±0.054 | 0.778±0.026 | 0.438 |
| Delta oxygen | 8.602±0.727 | 8.379±0.813 | 0.842 |
| CO2 production | 14.43±0.624 | 15.36±1.113 | 0.468 |
| Active period | |||
| Oxygen consumption | 32.42±1.969 | 33.16±4.784 | 0.881 |
| Respiratory quotient | 0.596±0.027 | 0.677±0.075 | 0.299 |
| Delta oxygen | 13.63±1.025 | 13.70±1.812 | 0.974 |
| CO2 production | 19.33±1.207 | 21.76±2.385 | 0.362 |
Table 4.
Mitochondrial respiration.
| Control (5) | Nitrate (5) | p-Value | |
|---|---|---|---|
| RCR | 5.553±0.794 | 4.551±0.398 | 0.301 |
| P/O Ratio (pmol s−1) | 1.232±0.132 | 1.215±0.078 | 0.910 |
| Leak respiration | 0.410±0.190 | 0.328±0.067 | 0.538 |
| State 3 respiration | 6.414±0.952 | 4.471±0.787 | 0.154 |
| State 4 respiration | 0.497±0.117 | 0.467±0.082 | 0.839 |
| Uncoupled respiration | 6.933±1.888 | 5.180±0.629 | 0.404 |
RCR=Respiratory control ratio.
Leak, States 3 and 4, and uncoupled respiration units are pmol /s/µg.
Cardiovascular parameters
Blood pressure
Acute dietary nitrate has been shown to decrease blood pressure in humans [7,8]. Blood pressure was measured by tail cuff monitoring after 16 months of nitrate treatment. No differences between the treatment groups were identified in the systolic, diastolic or mean arterial pressure (MAP) (Table 5).
Table 5.
Blood pressure.
| Control (10) | Nitrate (12) | p-Value | |
|---|---|---|---|
| Systolic | 95.8±1.49 | 98.1±2.42 | 0.452 |
| Diastolic | 66.8±1.32 | 66.9±1.47 | 0.972 |
| MAP | 76.1±1.26 | 76.9±1.74 | 0.730 |
Vascular reactivity
At final sacrifice, ex vivo reactivity of the carotid artery was carried out in a myograph. There were no differences in the endothelium-independent dilatory response to sodium nitroprusside between controls and nitrate treated mice (Fig. 5A). The endothelium-dependent response to acetylcholine was also similar between the two groups (Fig. 5B). Treatment with non-selective NO synthase inhibitor L-NAME for half an hour induced a similar sized contraction in both groups (Fig. 5 C). Taken together, the results demonstrate that long term dietary nitrate had no adverse effects on vascular reactivity.
Fig. 5.
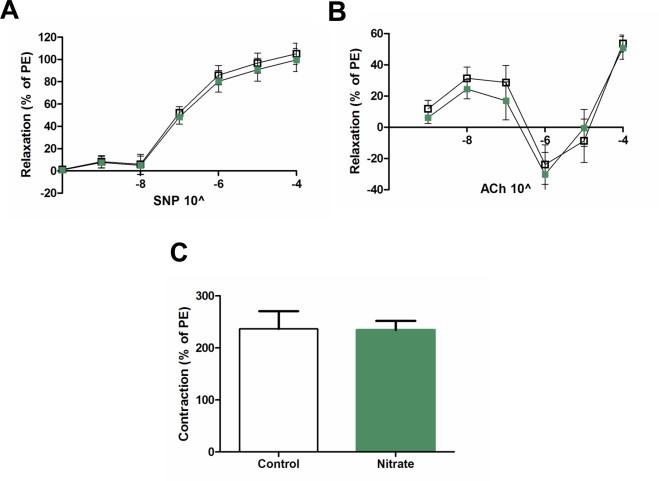
Ex vivo vascular reactivity. Carotid rings were analyzed for their response to sodium nitroprusside (SNP) (A), acetylcholine (ACh) (B) or 30 minute application of L-NAME (C). Control represented by open squares and nitrate by green squares. Values are means±SEM. PE=phenylephrine.
Vascular plasma markers
Analysis of nitrogen oxides in the final plasma collection indicated no difference in plasma nitrite or nitrate between the treatment groups (Fig. 6A and B). Two animals with liver nodules skewed the control group towards increased plasma nitrite levels inducing a trend towards significance (Fig. 6A). Plasma cGMP has been shown to be increased after acute nitrate dosing, though there were no differences after long term nitrate treatment (Fig. 6C). To confirm that the animals were ingesting nitrate, we measured urinary nitrate collected from the bladder at final sacrifice (Fig. 6D). Urine collected directly from the bladder at sacrifice contained similar amounts of nitrate in both groups. However, in the control group one mouse with liver nodules skewed the control data. Exclusion of this animal led to twice as high levels of urinary nitrate in the treated animals.
Fig. 6.
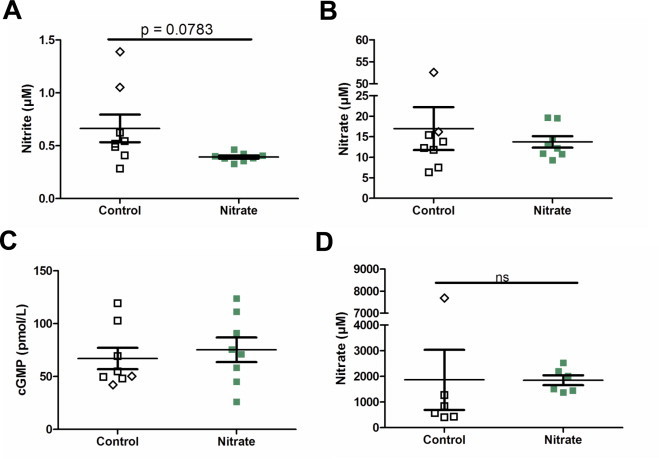
Nitrogen oxides and cGMP. Final plasma nitrite (A), nitrate (B), cGMP (C), and urinary nitrate (D). Note the split y-axis on graphs B and D. Data from two animals in the control groups with liver nodules are shown as diamonds. Values are means±SEM. Ns=non-significant.
Previous research has identified a crosstalk between nitrate and decreased eNOS function as evidenced by changes in plasma amino acid concentrations and aortic eNOS phosphorylation [33]. Nitric oxide is produced by the reaction of oxygen and l-arginine with the eNOS yielding l-citrulline. There were no significant differences in total arginine, citrulline and ornithine concentrations or in ratios of citrulline to arginine, or ornithine to citrulline (Table 6). Dimethyl arginines, which are inhibitors of eNOS function, were also analyzed with no significant differences in their plasma concentration (Table 6).
Table 6.
Plasma amino acids.
| Control (8) | Nitrate (8) | p-Value | |
|---|---|---|---|
| Arginine (µM) | 85.97±10.15 | 78.21±6.275 | 0.526 |
| Ornithine (µM) | 41.79±4.252 | 45.06±5.976 | 0.662 |
| Citrulline (µM) | 43.59±2.418 | 42.50±2.734 | 0.770 |
| Cit/Arg | 0.544±0.047 | 0.575±0.062 | 0.699 |
| Orn/Cit | 0.965±0.092 | 1.063±0.139 | 0.562 |
| SDMA (µM) | 0.302±0.036 | 0.266±0.009 | 0.344 |
| ADMA (µM) | 1.984±0.085 | 1.884±0.005 | 0.322 |
Inflammation and oxidative stress
Inflammatory markers in plasma
During aging there is a natural increase in plasma inflammatory markers [34]. Using a pro-inflammatory multiplex ELISA we found no differences in interferon-gamma, interleukin-1beta, interleukin-6, KC/GRO, (an interleukin-8 homolog found in mice), and interleukin 12p70 (Table 7). The anti-inflammatory interleukin-10 was significantly decreased in the nitrate supplemented group. The overall values were in the same range as we found in a batch of 6–9 month-old wildtype animals (unpublished data).
Table 7.
Plasma cytokines (pg/mL).
| Control (8) | Nitrate (8) | p-Value | |
|---|---|---|---|
| INF-gamma | 1.897±0.602 | 1.120±0.114 | 0.245 |
| IL-1beta | 5.708±0.298 | 6.824±0.616 | 0.125 |
| IL-6 | 49.07±14.68 | 32.52±3.421 | 0.309 |
| IL-10 | 57.04±5.991 | 35.79±5.329 | 0.018⁎ |
| KC/GRO | 60.26±12.83 | 54.97±4.552 | 0.708 |
| IL-12p70 | 216.9±83.63 | 135.6±8.981 | 0.360 |
p<0.05
NADPH oxidase activity in tissue
Oxidative stress due to increased NADPH oxidase activity is often increased with aging and chronic inflammation. NADPH oxidase activity was compared in heart, liver and kidney tissue. There were no differences between the treatment groups in tissue NADPH oxidase activity, supporting the lack of inflammation changes found in the plasma cytokine measurements (Fig. 7).
Fig. 7.
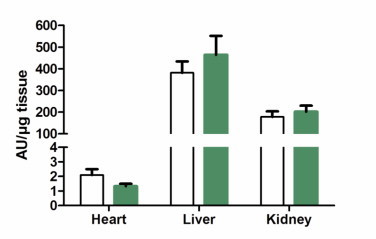
Tissue NAPDH oxidase activity. NADPH oxidase activity was measured by NADPH addition to heart, liver and kidney homogenates inducing superoxide production and reaction with lucigenin. Control represented by open bars and nitrate by green bars. Values are means ± SEM.
Survival
We made a survival curve of the animals that died or had to be euthanized due to poor health. The results suggested a slightly better median survival with nitrate supplementation vs the control group (Control 600.5 days vs Nitrate 640.5 days, p<0.05). This should be confirmed in a larger study.
Discussion
This study in normal healthy mice shows that dietary nitrate does not have any obvious adverse long-term effects, an important finding considering the major health concerns associated with nitrate intake. Though there are continued concerns of nitrate intake inducing adverse health effects in humans, our study found limited differences from control mice after close to a year and a half of high dietary nitrate. Our calculations extrapolating average mouse nitrate consumption to a 75 kg person would yield approximately 26.1 mg nitrate per day on the control diet and 350 mg/day on the nitrate diet. These doses fall at the ends of the average intake range of 40 mg to 120 mg/day published in World Health Organization advisory documents [35], combined from multiple national nitrate intake studies. A study from the Netherlands, analyzing nitrate and nitrite content in a duplicate sample of individual 24 h food and water intake in 1994, found a range of 1–322 mg/day with a median of 53 mg nitrate per day and approximately 20% of the volunteers exceeded 150 mg/day [36]. While the control is on the low side of published ranges, it is a plausible level due to increased societal reliance on processed food, often containing negligible nitrate content. Vegetarians often ingest more nitrate so the treatment regimen may be more representative of a vegetarian diet.
Prior research on the toxicity and carcinogenicity of nitrate in rats used doses 10–50 times that used in this study, without salt matched controls [17,18,20,21]. Even at such exorbitant doses Chow et al. (1980) saw no survival difference when compared to control, while Maekawa et al. (1982) saw more animals in the nitrate group surviving at termination but no differences in cancer incidence of various organs. A more recent study using a dose 10 times our nitrate dose over a shorter treatment period, found nitrate led to decreased weight gain and decreased plasma fasting insulin, T4 and T3 levels [20]. Our study showed no difference in group weight or weight gain over the course of 17 months of nitrate treatment.
The animals used in our study were healthy wildtype mice, which started to be afflicted by aging related issues and death around 20 months of age. The overall health of the mice may have prevented our finding beneficial effects due to long-term nitrate supplementation. The study, initially developed to document these beneficial effects, was modified towards a survival experiment due to lack of differences. Larsen et al. (2014) found acute nitrate did not change the glucose tolerance response in healthy volunteers, though there are other parameters such as blood pressure and oxygen consumption where acute nitrate induced changes. The long term supplementation may have also affected the nitrate homeostasis of the animals, which is supported by the lack of differences in final plasma nitrite, nitrate and cGMP. Urinary nitrate disposal increased from 7 months (data not shown) of treatment to 17 months reflecting a modified disposal potentially matching the nitrate intake. These changes could also have affected the ability to identify changes in blood pressure, vascular reactivity, indirect calorimetry and mitochondrial efficiency which were analyzed after 10 months of nitrate treatment. Conversion of nitrate to nitrite in the oral cavity and ingestion is imperative to achieve blood pressure improvement; which may be modified under extended nitrate treatment and homeostasis modifications. The lack of differences in nitrite concentration bears this out. In contrast to organic nitrate where continuous treatment leads to endothelial dysfunction, there were no adverse effects on vascular function as measured by blood pressure or ex vivo carotid response after long-term inorganic nitrate supplementation.
The plasma concentration of tri-iodothyronine (T3) and IL-10 were reduced, IL-10 significantly, compared to control after long term nitrate treatment. Whether these findings are coincidence or part of a program of overall signaling changes in the body remains to be determined. Previous studies analyzing T3 concentration and inflammatory cytokines in patients with euthyroid illness have conflicting results with one study finding no association between T3 and cytokines [37], while another reported decreased T3 and increased IL-10 and IL-6 [38].
T3 and T4 concentrations affect metabolic function and have been shown to be decreased in people with hypothyroidism and increased in hyperthyroidism. Body mass index changes inversely to plasma T3 concentration [39,40]. Weight loss studies looking at T3 concentration before and after weight loss show that plasma T3 was lower compared to controls [41–43]. Pourhassan et al. (2014) following a cohort of non-smoking adults between 2 and 4 years, stratified the volunteers by weight loss, weight gain or weight stable and found concomitant decreases in T3 and insulin in the weight loss group and a T3 concentration increase in the weight gain group. In contrast to studies using supra-physiological nitrate intake in rats which found decreased plasma T3 and T4 concentrations with decreased weight gain [20,26,27,30,44–46], there was no significant weight or weight gain difference between groups at any point during the 17 month period. Combining our findings of improved insulin response, decreased plasma T3 and plasma insulin in most of our mice, opens the possibility that nitrate treatment lowers the threshold for metabolic response.
The decrease in anti-inflammatory IL-10 is another counterintuitive event. In general IL-10 increases are described to lead to better insulin response, while IL-6 leads to insulin resistance [47]. IL-10 has been shown to be decreased in response to acute inflammation [48,49] and multiple sclerosis [50] inversely to the increased pro-inflammatory cytokines. There was no increase in inflammatory signaling in our experiments. Jadert et al. (2012) showed that acute nitrate decreased inflammatory response to signaling for neutrophil extravasation and recruitment without a change in overall clearance of a bacterial load [51]. Nitric oxide and nitrite have been shown to negatively modulate iNOS and NADPH oxidase activity [52]. As speculated with T3 maybe nitrate modulates inflammatory signaling lowering the basal cytokine expression. Future experiments will need to explore this possibility.
Conclusions
The findings in this study do not support any long-term toxic effects of dietary nitrate. Several of the beneficial effects described previously with short-term nitrate supplementation could not be reproduced in this model of natural aging. Intriguing results of decreased plasma insulin, and IL-10 and improved insulin tolerance and survival will need to be confirmed in larger follow up studies. Studies exploring different doses of nitrate supplementation will yield better indications for optimal supplementation.
Funding
This work was supported by Swedish Research Council, European Union FP7 research Grant (FP7-KBBE-2008-2B) FLAVIOLA, Swedish Heart and Lung Foundation, Stockholm City Council (ALF), David and Astrid Hagelén Foundation and Funds from Karolinska Institutet.
Acknowledgments
We would like to thank Margaretha Stensdotter, Annika Olsson and Carina Nihlén for their valuable technical assistance.
Contributor Information
Michael P. Hezel, Email: Michael.Hezel@ki.se.
Eddie Weitzberg, Email: Eddie.Weitzberg@ki.se.
References
- 1.Mensinga T.T., Speijers G.J., Meulenbelt J. Health implications of exposure to environmental nitrogenous compounds. Toxicological Reviews. 2003;22(1):41–51. doi: 10.2165/00139709-200322010-00005. 14579546 [DOI] [PubMed] [Google Scholar]
- 2.Tannenbaum S.R., Correa P. Nitrate and gastric cancer risks. Nature. 1985;317(6039):675–676. doi: 10.1038/317675b0. 4058577 [DOI] [PubMed] [Google Scholar]
- 3.Moncada S., Palmer R.M., Higgs E.A. Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacological Reviews. 1991;43(2):109–142. 1852778 [PubMed] [Google Scholar]
- 4.Lundberg J.O., Weitzberg E., Gladwin M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nature Reviews Drug Discovery. 2008;7(2):156–167. doi: 10.1038/nrd2466. 18167491 [DOI] [PubMed] [Google Scholar]
- 5.Lundberg J.O. Nitrate, bacteria and human health. Nature Reviews Microbiology. 2004;2(7):593–602. doi: 10.1038/nrmicro929. 15197394 [DOI] [PubMed] [Google Scholar]
- 6.Weitzberg E., Lundberg J.O. Novel aspects of dietary nitrate and human health. Annual Review of Nutrition. 2013;33:129–159. doi: 10.1146/annurev-nutr-071812-161159. 23642194 [DOI] [PubMed] [Google Scholar]
- 7.Larsen F.J. Effects of dietary nitrate on blood pressure in healthy volunteers. New England Journal of Medicine. 2006;355(26):2792–2793. doi: 10.1056/NEJMc062800. 17192551 [DOI] [PubMed] [Google Scholar]
- 8.Webb A.J. Acute blood pressure lowering, vasoprotective, and antiplatelet properties of dietary nitrate via bioconversion to nitrite. Hypertension. 2008;51(3):784–790. doi: 10.1161/HYPERTENSIONAHA.107.103523. 18250365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Richardson G. The ingestion of inorganic nitrate increases gastric S-nitrosothiol levels and inhibits platelet function in humans. Nitric Oxide. 2002;7(1):24–29. doi: 10.1016/s1089-8603(02)00010-1. 12175816 [DOI] [PubMed] [Google Scholar]
- 10.Rodriguez-Mateos A. Interactions between cocoa flavanols and inorganic nitrate: additive effects on endothelial function at achievable dietary amounts. Free Radical Biology and Medicine. 2015;80:121–128. doi: 10.1016/j.freeradbiomed.2014.12.009. 25530151 [DOI] [PubMed] [Google Scholar]
- 11.Larsen F.J. Dietary inorganic nitrate improves mitochondrial efficiency in humans. Cell Metabolism. 2011;13(2):149–159. doi: 10.1016/j.cmet.2011.01.004. 21284982 [DOI] [PubMed] [Google Scholar]
- 12.Govoni M. The increase in plasma nitrite after a dietary nitrate load is markedly attenuated by an antibacterial mouthwash. Nitric Oxide. 2008;19(4):333–337. doi: 10.1016/j.niox.2008.08.003. 18793740 [DOI] [PubMed] [Google Scholar]
- 13.Kapil V. Physiological role for nitrate-reducing oral bacteria in blood pressure control. Free Radical Biology and Medicine. 2013;55:93–100. doi: 10.1016/j.freeradbiomed.2012.11.013. 23183324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Petersson J. Gastroprotective and blood pressure lowering effects of dietary nitrate are abolished by an antiseptic mouthwash. Free Radical Biology and Medicine. 2009;46(8):1068–1075. doi: 10.1016/j.freeradbiomed.2009.01.011. 19439233 [DOI] [PubMed] [Google Scholar]
- 15.Bazzano L.A. Intake of fruit, vegetables, and fruit juices and risk of diabetes in women. Diabetes Care. 2008;31(7):1311–1317. doi: 10.2337/dc08-0080. 18390796 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Park S.K. Fruit, vegetable, and fish consumption and heart rate variability: the veterans administration normative aging study. American Journal of Clinical Nutrition. 2009;89(3):778–786. doi: 10.3945/ajcn.2008.26849. 19158214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Aly H.A. Potential testicular toxicity of sodium nitrate in adult rats. Food and Chemical Toxicology. 2010;48(2):572–578. doi: 10.1016/j.fct.2009.11.034. 19922758 [DOI] [PubMed] [Google Scholar]
- 18.Chow C.K., Chen C.J., Gairola C. Effect of nitrate and nitrite in drinking water on rats. Toxicology Letters. 1980;6(3):199–206. doi: 10.1016/0378-4274(80)90192-7. 7404601 [DOI] [PubMed] [Google Scholar]
- 19.Chow C.K., Hong C.B. Dietary vitamin E and selenium and toxicity of nitrite and nitrate. Toxicology. 2002;180(2):195–207. doi: 10.1016/s0300-483x(02)00391-8. 12324194 [DOI] [PubMed] [Google Scholar]
- 20.El-Wakf A.M. Fenugreek potent activity against nitrate-induced diabetes in young and adult male rats. Cytotechnology. 2015;67(3):437–447. doi: 10.1007/s10616-014-9702-7. 24615531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Maekawa A. Carcinogenicity studies of sodium nitrite and sodium nitrate in F−344 rats. Food and Chemical Toxicology. 1982;20(1):25–33. doi: 10.1016/s0278-6915(82)80005-7. 7200054 [DOI] [PubMed] [Google Scholar]
- 22.Yang T. Abrogation of adenosine A receptor signalling improves metabolic regulation in mice by modulating oxidative stress and inflammatory responses. Diabetologia. 2015 doi: 10.1007/s00125-015-3570-3. [DOI] [PubMed] [Google Scholar]
- 23.Gao X. NADPH oxidase in the renal microvasculature is a primary target for blood pressure-lowering effects by inorganic nitrate and nitrite. Hypertension. 2015;65(1):161–170. doi: 10.1161/HYPERTENSIONAHA.114.04222. 25312440 [DOI] [PubMed] [Google Scholar]
- 24.Mulvany M.J., Halpern W. Contractile properties of small arterial resistance vessels in spontaneously hypertensive and normotensive rats. Circulation Research. 1977;41(1):19–26. doi: 10.1161/01.res.41.1.19. 862138 [DOI] [PubMed] [Google Scholar]
- 25.Brown C.M. Simultaneous determination of 6 L-arginine metabolites in human and mouse plasma by using hydrophilic-interaction chromatography and electrospray tandem mass spectrometry. Clinical Chemistry. 2011;57(5):701–709. doi: 10.1373/clinchem.2010.155895. 21406573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chaoui A.A. Effects of inorganic nitrates on thyroid gland activity and morphology in female rats. Therapie. 2004;59(4):471–475. doi: 10.2515/therapie:2004079. 15559551 [DOI] [PubMed] [Google Scholar]
- 27.Jahreis G. Nitrate-induced hypothyroidism is associated with a reduced concentration of growth hormone-releasing factor in hypothalamic tissue of rats. Experimental and Clinical Endocrinology. 1991;97(1):109–112. doi: 10.1055/s-0029-1211049. 1864311 [DOI] [PubMed] [Google Scholar]
- 28.Oğur R., Korkmaz A., Hasde M. Effects of high nitrate intake in rats. Journal of Basic and Clinical Physiology and Pharmacology. 2000;11(1):47–56. doi: 10.1515/jbcpp.2000.11.1.47. 10851662 [DOI] [PubMed] [Google Scholar]
- 29.Carlström M. Dietary inorganic nitrate reverses features of metabolic syndrome in endothelial nitric oxide synthase-deficient mice. Proceedings of the National Acadamy of Science United States of America. 2010;107(41):17716–17720. doi: 10.1073/pnas.1008872107. 20876122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jahreis G. Growth impairment caused by dietary nitrate intake regulated via hypothyroidism and decreased somatomedin. Endocrinologia Experimentalis. 1987;21(3):171–180. 3499306 [PubMed] [Google Scholar]
- 31.Larsen F.J. Dietary nitrate reduces resting metabolic rate: a randomized, crossover study in humans. American Journal of Clinical Nutrition. 2014;99(4):843–850. doi: 10.3945/ajcn.113.079491. 24500154 [DOI] [PubMed] [Google Scholar]
- 32.Larsen F.J. Effects of dietary nitrate on oxygen cost during exercise. Acta Physiologica (Oxford) 2007;191(1):59–66. doi: 10.1111/j.1748-1716.2007.01713.x. 17635415 [DOI] [PubMed] [Google Scholar]
- 33.Carlström M. Cross-talk between nitrate–Nitrite-NO and NO synthase pathways in control of vascular NO homeostasis. Antioxidants and Redox Signaling. 2014 doi: 10.1089/ars.2013.5481. 24224525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Franceschi C., Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. Journals of Gerontology Series A: Biological Sciences and Medical Sciences. 2014;69(Suppl. 1):S4–S9. doi: 10.1093/gerona/glu057. 24833586 [DOI] [PubMed] [Google Scholar]
- 35.WHO, Nitrate and Nitrite in Drinking Water, Background Document for Guidelines for Drinking Water Quality [PDF] 20112011, Available from: 〈http://www.who.int/water_sanitation_health/dwq/chemicals/nitratesnitrite/en/〉 (cited 19.04.15).
- 36.van Vliet J.J. Twenty-four-hour duplicate diet study 1994; nitrate and nitrite: method development and intake per person per day. Cancer Letters. 1997;114(1–2):305–307. doi: 10.1016/s0304-3835(97)04688-0. 9103317 [DOI] [PubMed] [Google Scholar]
- 37.Boelen A., Schiphorst M.C., Wiersinga W.M. Relationship between serum 3,5,3′-triiodothyronine and serum interleukin-8, interleukin-10 or interferon gamma in patients with nonthyroidal illness. Journal of Endocrinological Investigation. 1996;19(7):480–483. doi: 10.1007/BF03349894. 8884543 [DOI] [PubMed] [Google Scholar]
- 38.Abo-Zenah H.A. Relating circulating thyroid hormone concentrations to serum interleukins-6 and −10 in association with non-thyroidal illnesses including chronic renal insufficiency. B.M.C. Endocrine Disorders. 2008;8:1. doi: 10.1186/1472-6823-8-1. 18211669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Iglesias P. Serum concentrations of adipocytokines in patients with hyperthyroidism and hypothyroidism before and after control of thyroid function. Clinical Endocrinology (Oxford) 2003;59(5):621–629. doi: 10.1046/j.1365-2265.2003.01897.x. 14616887 [DOI] [PubMed] [Google Scholar]
- 40.Li H. Clinical evaluation of various thyroid hormones on thyroid function. International Journal of Endocrinology. 2014;2014:618572. doi: 10.1155/2014/618572. 25548564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Agnihothri R.V. Moderate weight loss is sufficient to affect thyroid hormone homeostasis and inhibit its peripheral conversion. Thyroidology. 2014;24(1):19–26. doi: 10.1089/thy.2013.0055. 23902316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Astrup A. Low resting metabolic rate in subjects predisposed to obesity: a role for thyroid status. American Journal of Clinical Nutrition. 1996;63(6):879–883. doi: 10.1093/ajcn/63.6.879. 8644681 [DOI] [PubMed] [Google Scholar]
- 43.Pourhassan M. Impact of body composition during weight change on resting energy expenditure and homeostasis model assessment index in overweight nonsmoking adults. American Journal of Clinical Nutrition. 2014;99(4):779–791. doi: 10.3945/ajcn.113.071829. 24500156 [DOI] [PubMed] [Google Scholar]
- 44.Jahreis G. Effect of chronic dietary nitrate and different iodine supply on porcine thyroid function, somatomedin-C-level and growth. Experimental and Clinical Endocrinology. 1986;88(2):242–248. doi: 10.1055/s-0029-1210603. 3556413 [DOI] [PubMed] [Google Scholar]
- 45.Mukhopadhyay S. Evaluation of possible goitrogenic and anti-thyroidal effect of nitrate, a potential environmental pollutant. Indian Journal of Physiology and Pharmacology. 2005;49(3):284–288. 16440845 [PubMed] [Google Scholar]
- 46.Zaki A. Impact of nitrate intake in drinking water on the thyroid gland activity in male rat. Toxicology Letters. 2004;147(1):27–33. doi: 10.1016/j.toxlet.2003.10.010. 14700525 [DOI] [PubMed] [Google Scholar]
- 47.Aroor A.R. Maladaptive immune and inflammatory pathways lead to cardiovascular insulin resistance. Metabolism. 2013;62(11):1543–1552. doi: 10.1016/j.metabol.2013.07.001. 23932846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kedzierski L. Decreased IL-10 and IL-13 production and increased CD44hi T cell recruitment contribute to Leishmania major immunity induced by non-persistent parasites. European Journal of Immunology. 2008;38(11):3090–3100. doi: 10.1002/eji.200838423. 18924210 [DOI] [PubMed] [Google Scholar]
- 49.Petursdottir D.H., Olafsdottir I., Hardardottir I. Dietary fish oil increases tumor necrosis factor secretion but decreases interleukin-10 secretion by murine peritoneal macrophages. Journal of Nutrition. 2002;132(12):3740–3743. doi: 10.1093/jn/132.12.3740. 12468616 [DOI] [PubMed] [Google Scholar]
- 50.Hirotani M. Decreased IL-10 production mediated by Toll-like receptor 9 in B cells in multiple sclerosis. Journal of Neuroimmunology. 2010;221(1–2):95–100. doi: 10.1016/j.jneuroim.2010.02.012. 20227772 [DOI] [PubMed] [Google Scholar]
- 51.Jädert C. Decreased leukocyte recruitment by inorganic nitrate and nitrite in microvascular inflammation and NSAID-induced intestinal injury. Free Radical Biology and Medicine. 2012;52(3):683–692. doi: 10.1016/j.freeradbiomed.2011.11.018. 22178413 [DOI] [PubMed] [Google Scholar]
- 52.Yang T. Inorganic nitrite attenuates NADPH oxidase-derived superoxide generation in activated macrophages via a nitric oxide-dependent mechanism. Free Radical Biology and Medicine. 2015;83:159–166. doi: 10.1016/j.freeradbiomed.2015.02.016. 25724690 [DOI] [PubMed] [Google Scholar]