

Abstract
The frequency of H5N1 avian influenza outbreaks in China and Eastern Europe has raised concern in the world health community regarding the potential for an influenza pandemic. Efforts to monitor the disease will only provide minimal warning in a global society, and steps must be taken to prevent the morbidity and mortality associated with past pandemics. The current stockpiling of antibody-inducing “bird flu” vaccines assumes the strain that emerges will be the same as strains currently circulating. We propose a novel consensus-based approach to vaccine development, employing a DNA vaccine strategy that can provide more highly cross-reactive cellular immunity against lethal influenza infection. We show such constructs can induce strong cellular immunity against H5 influenza antigens.
Keywords: Avian influenza, DNA vaccine, consensus
Introduction
Current vaccination strategies against influenza rely on surface antigen-directed antibody responses to prevent infection. The ability of strain-specific antibodies to prevent infection has been well-documented, and will likely continue to be the used to provide protection from annual influenza infection. However, reliance on humoral immunity to prevent infection requires that the vaccine strain be matched identically to the strain causing infection, which must be carefully predicted from circulating viruses and prepared for the following flu season. While this strategy is largely effective, there are several drawbacks. The length of time required to produce these vaccines is such that they would be largely useless during a rapidly spreading pandemic. In addition, as occurred during the 2003-2004 influenza season, the virus can drift between the time a vaccine strain is chosen and produced [1]. As mentioned above, recent reports indicate that a new strain of H5N1 avian influenza has become the dominant strain circulating among poulty in China, having spread to a majority of provinces [2]. Current stockpiles of vaccine, and those in production, may already be outdated.
Antibody-based influenza vaccines have been employed successfully since the 1940s. However, pandemics in 1957 and 1968 resulted in 1-2 million deaths and 700,000 deaths worldwide, respectively [3]. These events do not question the ability of antibodies to prevent influenza infection, but rather our exclusive reliance on these vaccines.
While the ability of antibodies to prevent influenza infection is well-established, the importance and potential of cellular immunity against influenza is still being investigated. Several studies have shown that cell-mediated immunity can provide protection from the morbidity and mortality associated with pathogenic influenza infection [4]. Mice immunologically primed with sub-lethal doses of Influenza A show a dramatically enhanced ability to recover from lethal challenge with heterologous virus – to which antibodies would give no protection. This is seen when the internal genes of the challenge virus are highly homologous [5] or more divergent [6].
Cellular immune responses have two advantages over antibody-dependent protection. While antibody responses are highly specific, and so susceptible to evasion by antigenic drift and shift [7], cellular immune responses are far more cross-reactive. The promiscuity of CD4 and CD8 T cell receptors could allow them to recognize epitopes even after the addition of a point mutation. While escape from CTLs is possible and observable [8-11], also observable are T cells that recognize a range of peptide sequences . In addition, the functional constraints placed on internal structural genes, inaccessible to antibodies, limit their ability to mutate. These functional constraints, linked to immunodominant epitopes, have been well documented for the internal influenza proteins [12-14]. In addition, the presence of pre-existing cross-reactive CD4+ helper cells allows for the more rapid generation of a type-specific antibody response, which can aid in neutralizing remaining virus [15].
Current vaccine strategies completely ignore the cellular aspect of protection from influenza. FluMist, the only currently approved vaccine that drives CTL responses, in addition to emerging reverse genetic approaches to vaccine design, all rely on an identical genetic background for the generation of cellular immunity. This avoidance of generating cellular immune responses against the evolving pool of influenza viruses could be a significant error.
Our lab and others have previously published work on consensus DNA immunogens [16-19]. These constructs have been shown to not only induce strong CD8+ and CD4+ T cell responses, but that these responses are in fact more cross-reactive than their individual component constructs. We present here several novel consensus immunogens against avian influenza. These constructs were designed by analyzing a large number of circulating avian influenza viruses in order to generate a construct with the potential to induce highly cross-reactive cellular immune responses. By definition, these sequences should retain the most highly conserved regions, providing appropriate targets for a cellular immune response.
Materials & Methods
Construct and Synthesis
Consensus sequences were optimized for expression, including codon and RNA optimization (GeneArt, Regensburg, Germany). These constructs were then synthesized and inserted into the pVAX1 expression vector (Invitrogen).
In vitro expression
Construct expression was confirmed by utilizing a T7-promoter in the pVAX backbone and T7-based coupled transcription/translation system containing S35-methionine (Promega, Madison, WI) [22]. The synthesized protein was immunoprecipitated using an anti-HA tag antibody (Abgent). The immunoprecipitated protein was electrophoresed on a 12% SDS-PAGE gel (NuPage) and subsequently fixed and dried. Autoradiography was performed to detect incorporated an S35-labeled gene product.
Generation of Recombinant HA
Recombinant protein was generated by Abgent. Briefly, constructs were subcloned into a bacterial expression vector (pET21a(+)), sequence verified, and used to transform the BL21(DE3) strain. The protein was then isolated and purified on an Ni-NTA affinity chromatography column and eluted with imidazole, which was then dialyzed into PBS.
Mice and immunizations
For DNA immunizations, the quadricep muscle of 6- to 8-wk-old female BALB/c or C57BL/6 mice (The Jackson Laboratory) were injected 2-3 times, two weeks apart, and sacrificed one week following the final immunization. For all experiments, the mice were immunized with 50μg (HA) or 100ug (NA and M) of antigen-encoding plasmid or vector control. All DNA was made using Qiagen columns (endotoxin free) [20]. All animals were housed in a temperature-controlled, light-cycled facility at the University of Pennsylvania, and they were cared for under the guidelines of the National Institutes of Health and the University of Pennsylvania. For protein immunizations, mice were injected i.p. with 5μg recombinant protein in 250μL Complete (prime) or Incomplete (boost) Freund’s Adjuvant (Sigma) two weeks apart.
Splenocyte Purification
Spleens from each mouse were harvested and pooled in a 15-ml conical containing RPMI 1640 (one tube for each experimental group). In a sterile tissue culture hood, the pooled spleens from each experimental group were placed in a petri dish and crushed using the plunger from a 3-ml syringe. The cells were then incubated for 5–10 min at room temperature in ACK lysing buffer (Biosource). The splenocytes were washed and put through a 70-μm cell strainer to remove any remaining spleen organ stroma. The splenocytes were washed in RPMI 1640 twice, resuspended in R10 medium (RPMI 1640 plus 10% FBS), and counted (cell viability is determined using trypan blue stain) using a hemocytometer [20].
ELISpot assay
An ELISPOT assay was conducted as previously described [21]. Briefly, ELISPOT 96-well plates (Millipore) were coated with anti-mouse IFN-γ capture Ab and incubated for 24 h at 4°C (R&D Systems). The following day, plates were washed and blocked for 2 h with 1% BSA. Two hundred thousand splenocytes from the immunized mice were added to each well and stimulated overnight at 37°C in 5% CO2 in the presence of RPMI 1640 (negative control), Con A (positive control), or specific peptide Ags (10 μg/ml) (Invitrogen). Peptide pools consist of 15-mer peptides overlapping by 11 amino acids. After 24 h of stimulation, the cells were washed and incubated for 24 h at 4°C with biotinylated anti-mouse IFN-γ Ab (R&D Systems). The plates were washed, and streptavidin-alkaline phosphatase (R&D Systems) was added to each well and incubated for 2h at room temperature. The plate was washed, and 5-bromo-4-chloro-3′-indolylphosphate p-toluidine salt and nitro blue tetrazolium chloride (chromogen color reagent; R&D Systems) were added to each well. The plate was then rinsed with distilled water and dried at room temperature. Spots were counted by an automated ELISPOT reader (CTL Limited).
Antibody ELISA assay
Antigen-specific antibodies were measured by ELISA. Briefly, 96-well high-binding polystyrene plates (Corning, New York) plates were coated overnight at 4°C with recombinant protein (2 μg/ml, generated by Abgent in a bacterial expression system) for diluted in PBS. The next day, plates were washed with PBST (PBS, 0.05% Tween 20), blocked for one hour with 3% BSA in PBST, and incubated with serial dilutions of serum from immunized and naïve mice for 1h at 37°C. Bound IgG was detected using goat anti-mouse IgG-HRP (Research Diagnostics, NJ) at a dilution of 1:10000. Bound enzyme was detected by the addition of the chromogen substrate solution TMB (R&D Systems), and read at 450 nm on a Biotek EL312e Bio-Kinetics reader. All serum samples were tested in duplicate.
Virus Challenge
Isoflurane-anaesthetized mice were intranasally inoculated with 100 LD50 of A/Hanoi/30408/2005 (obtained from CDC) in 50 μl MEM/3% BSA. LD50 was determined to be 15.6 PFU/mouse, determined by Reed and Muench method
Results
Construction of a Novel Consensus Immunogen
In an effort to develop an immunogen with the ability to induce highly cross-reactive cellular responses, we took two approaches to designing several consensus constructs against avian influenza. Presented here are constructs against H5N1 hemagglutinin (HA), neuraminidase (NA), and matrix 1 protein. For the hemagglutinin construct, 16 sequences were chosen from avian viruses isolated between 1997 and 2005 that caused infection and death in humans [23]. The amino acid sequences were aligned, and the most common amino acid at each position was chosen. Figure 1b shows a phylogenetic analysis of the 16 component proteins and the placement of the consensus construct among them. After generation of the consensus sequence, an HA tag was added to the C-terminus for expression verification. For the neuraminidase (NA) and matrix 1 (M) proteins, 40 and 45 primary sequences, respectively, were chosen as component viruses. The DNA sequences chosen for each gene were from both H1N1 and H5N1 viruses, and chosen from varying countries in order to avoid sampling bias. The DNA sequences were aligned, and the most common nucleotide chosen. Deduced amino acid sequences were used to guide the introduction of alignment gaps so that they were introduced between codons. Figures 2b and 3b depict phylogenetic analyses of these constructs, including the placement of their consensus constructs. Both the neuraminidase and matrix 1 proteins have N-ternimal IgE leaders to enhance expression [24], in addition to C-terminal HA tags for expression verification.
Figure 1.
Hemagglutinin (HA) construct characterization. Figure 1a shows S35-labeled in vitro translation of the synthesized construct. The antigen was translated and run on a 12% SDS gel, and subject to radiographic analysis. The antigen runs at its predicted molecular weight, confirming expression. Figure 1b shows a phylogenetic analysis of the consensus construct. The star represents the consensus construct, relative to its component viruses.
Figure 2.

Neuraminidase (NA) construct characterization. Figure 2a shows S35-labeled in vitro translation of the synthesized construct. The antigen was translated and run on a 12% SDS gel, and subject to radiographic analysis. The antigen runs at its predicted molecular weight, confirming expression. Figure 2b shows a phylogenetic analysis of the consensus construct. The star represents the consensus construct, relative to its component viruses.
Figure 3.
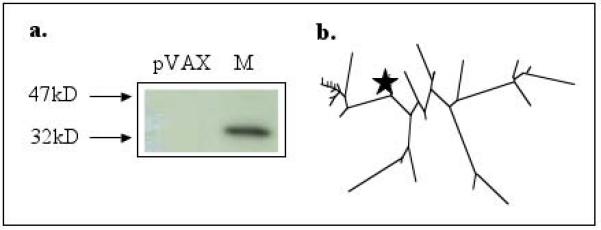
Matrix 1 (M) construct characterization. Figure 3a shows S35-labeled in vitro translation of the synthesized construct. The antigen was translated and run on a 12% SDS gel, and subject to radiographic analysis. The antigen runs at its predicted molecular weight, confirming expression. Figure 3b shows a phylogenetic analysis of the consensus construct. The star represents the consensus construct, relative to its component viruses.
Consensus Immunogen Expression
Expression of our synthesized genes was verified by S35-labeled in vitro transcription and translation. Translation products were immunoprecipitated using an anti-HA tag antibody. SDS-PAGE and radiographic analysis shows that each construct runs at its theoretically predicted molecular weight (Figures 1a-3a). After expression verification, the HA tag was removed from the hemagglutinin construct in order to avoid interfering with intracellular trafficking [25].
Cellular Immunogenicity
We first looked at the ability of our hemmaglutinin construct to induce CD8+ CTL responses as determined by IFN-γ ELISpot assays. As shown in figure 4, the consensus construct was able to induce strong CTL responses in BALB/C mice after two immunizations. Worth noting, while we have not yet mapped the dominant epitopes of the individual constructs, there is a clear bias towards peptide pool 4 in the H5 HA-immunized mice. This region includes an amino acid sequence identical to the dominant epitope of Influenza A/PR/8/34 (IYSTVASSL), which may reveal immunogenically useful constraints on the antigen.
Figure 4.

Interferon-γ ELISpot. Balb/c mice were immunized two weeks apart with 50ug pVAX vector or pH5HA and sacrificed one week later. Splenocytes were harvested and cultured overnight in the presence of R10 (negative control) or 10μg/mL of one of four peptide pools, made up of 15-mer peptides overlapping by 11 amino acids, spanning the length of the hemmaglutinin antigen. Spot forming units were quantified by an automated ELISPOT reader, and the raw values were normalized to SFU per million splenocytes. Values represent the mean (±SD) of duplicate wells.
We then looked at our NA and matrix 1 constructs in more detail. Both BALB/C and C57BL/6 mice were immunized, and IFN-γ ELISpot assays were performed. As shown in figures 5 and 6, these constructs were able to induce potent immune responses in both mouse strains – indicating an ability to be recognized across multiple MHCs. In addition, CD8-depleted controls were analyzed to assess the contribution of CD4+ T cells to the interferon response. As shown in figures 5 and 6, CD8+ T cells are responsible for the majority of interferon secretion.
Figure 5.

Interferon-γ ELISpot. BALB/C or C57BL/6 mice were immunized three times, each two weeks apart, with 100ug pVAX vector or pNA and sacrificed one week later. Splenocytes were harvested and cultured overnight in the presence of R10 (negative control) or 10μg/mL of one of three peptide pools, made up of 15-mer peptides overlapping by 11 amino acids, spanning the length of the neuraminidase antigen. Also shown are CD8-depleted controls. Spot forming units were quantified by an automated ELISPOT reader, and the raw values were normalized to SFU per million splenocytes. Values represent the mean (±SD) of triplicate wells.
Figure 6.

Interferon-γ ELISpot. BALB/C or C57BL/6 mice were immunized three times, each two weeks apart, with 100ug pVAX vector or pM and sacrificed one week later. Splenocytes were harvested and cultured overnight in the presence of R10 (negative control) or 10μg/mL of one of two peptide pools, made up of 15-mer peptides overlapping by 11 amino acids, spanning the length of the matrix 1 antigen. Also shown are CD8-depleted controls. Spot forming units were quantified by an automated ELISPOT reader, and the raw values were normalized to SFU per million splenocytes. Values represent the mean (±SD) of triplicate wells.
Humoral Immunogenicity
While we hypothesize that the strength of our consensus immunogens to protect from lethal influenza will lie with the cellular arm of the immune system, cross-reactive but non-neutralizing antibodies can provide a certain degree of protection against disease severity. In order to determine if our constructs induce antibody responses, we performed an antibody ELISA on HA-immunized mouse serum to determine antibody titer, as show in figure 7. While low-titer antibodies are present, we believe we can improve upon this with alternative means of plasmid delivery, specifically electroporation.
Figure 7.

Hemmaglutinin antibody ELISA. Balb/c mice were immunized two weeks apart with 50ug pVAX vector or pH5HA and sacrificed one week later. Serum was collected and subject to serial dilutions. Serum was incubated for one hour at 37°C on 96-well plates coated with 2μg/mL of recombinant H5HA, and antibody was detected using anti-mouse IgG-HRP. Values represent the mean (±SD) of duplicate wells.
Challenge Study
Recombinant protein of our H5 HA construct was produced in a bacterial expression system for use in immune assays. As an initial experiment to test the activity of our consensus HA immunogen, we used this protein, in addition to Complete (prime) and Incomplete (boost) Freund’s adjuvant to immunize mice. These mice were allowed to rest for four weeks, and then infected with an avian influenza (A/Hanoi/30408/05). While these mice were considerably older (presumably more immunologically competent) than those used to determine virus LD50, allowing for high survival in the naïve group, this initial survival data, shown in figure 8, does show that this protein was able to confer a degree of protection to immunized mice. We believe that optimization of our DNA constructs, individually or in combination, will be able to build upon this protection with the induction of more potent cytotoxic immune responses, which are not seen with protein immunizations.
Figure 8.
Avian influenza challenge. 10 BALB/C mice were primed with 5μg of recombinant H5HA in Complete Freund’s adjuvant and boosted two weeks later with 5μg recombinant H5HA in Incomplete Freund’s adjuvant. Mice were allowed to rest for four weeks and then challenged, in addition to a control group of 10 BALB/C mice, with Avian Influenza A/Hanoi/30408/05. Shown here are the survival rates of the different groups of mice.
Discussion
Our current understanding of avian influenza, including the mutations that might allow it to transform into a virus transmissible from person-to-person, remains limited. However, the ability of avian influenza to cause disease and death in humans is well documented. The absence of herd immunity to any strain of avian influenza requires that we be prepared with means to fight an epidemic and prevent recurrences of past pandemics. Current strategies of vaccinating against influenza will not be sufficient. Avian influenza aside, current strategies of influenza vaccination leave much to be desired.
While a vaccine based on cellular immunity may not prevent infection, we do believe it could significantly reduce the morbidity and mortality associated with lethal influenza. By decreasing the duration of infection, it could also decrease transmission of the virus. Instead of replacing existing and emerging vaccines based on humoral immunity, these could provide a supplement to guard against rapidly spreading, pathogenic influenza. With the advantage of representing a large number of contemporary primary virus isolates, we aim to generate a vaccine that does not need to be redesigned on an annual basis, but rather generates responses against more highly conserved regions that functional constraints will keep in place over time.
Our work here extends our earlier work with consensus HIV vaccines, which show that constructs, designed to reflect a large number of primary influenza sequences, have the ability to induce potent and highly cross-reactive cellular immune responses. Subsequent studies detailing the cross-reactivity of these responses, in addition to the ability of these vaccines to drive protection against a broadarray of lethal virus challenges, is currently in progress.
Acknowledgements
Vaccine studies were supported by grants awarded to D.B.W. Challenges were supported by G.P.K.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Referenecs
- 1.Fauci AS. Pandemic influenza threat and preparedness. Emerg Infect Dis. 2006;12:73–7. doi: 10.3201/eid1201.050983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Smith G, Fan X, Wang J, Li K, Qin K, Zhang K, Vijaykrishna D, Cheung C, Huang K, Rayner J, Peiris J, Chen H, Webster R, Guan Y. Emergence and predominance of an H5N1 influenza variant in China. PNAS. 2006;103:16936–16941. doi: 10.1073/pnas.0608157103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ghendon Y. Introduction to pandemic influenza through history. European Journal of Epidemiology. 2005;10:451–453. doi: 10.1007/BF01719673. [DOI] [PubMed] [Google Scholar]
- 4.Kreijtz JH, Bodewes R, van Amerongen G, Kuiken T, Fouchier RA, Osterhaus AD, Rimmelzwaan GF. Primary influenza A virus infection induces cross-protective immunity against a lethal infection with a heterosubtypic virus strain in mice. Vaccine. 2006 doi: 10.1016/j.vaccine.2006.08.036. [DOI] [PubMed] [Google Scholar]
- 5.O’Neill E, Krauss SL, Riberdy JM, Webster RG, Woodland DL. Heterologous protection against lethal A/HongKong/156/97 (H5N1) influenza virus infection in C57BL/6 mice. J Gen Virol. 2000;81:2689–96. doi: 10.1099/0022-1317-81-11-2689. [DOI] [PubMed] [Google Scholar]
- 6.Thomas PG, Keating R, Hulse-Post DJ, Doherty PC. Cell-mediated protection in influenza infection. Emerg Infect Dis. 2006;12:48–54. doi: 10.3201/eid1201.051237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Scholtissek C. Source for influenza pandemics. Eur J Epidemiol. 1994;10:455–8. doi: 10.1007/BF01719674. [DOI] [PubMed] [Google Scholar]
- 8.Berkhoff EG, Boon AC, Nieuwkoop NJ, Fouchier RA, Sintnicolaas K, Osterhaus AD, Rimmelzwaan GF. A mutation in the HLA-B*2705-restricted NP383-391 epitope affects the human influenza A virus-specific cytotoxic T-lymphocyte response in vitro. J Virol. 2004;78:5216–22. doi: 10.1128/JVI.78.10.5216-5222.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Boon AC, de Mutsert G, Graus YM, Fouchier RA, Sintnicolaas K, Osterhaus AD, Rimmelzwaan GF. Sequence variation in a newly identified HLA-B35-restricted epitope in the influenza A virus nucleoprotein associated with escape from cytotoxic T lymphocytes. J Virol. 2002;76:2567–72. doi: 10.1128/jvi.76.5.2567-2572.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rimmelzwaan GF, Boon AC, Voeten JT, Berkhoff EG, Fouchier RA, Osterhaus AD. Sequence variation in the influenza A virus nucleoprotein associated with escape from cytotoxic T lymphocytes. Virus Res. 2004;103:97–100. doi: 10.1016/j.virusres.2004.02.020. [DOI] [PubMed] [Google Scholar]
- 11.Voeten JT, Bestebroer TM, Nieuwkoop NJ, Fouchier RA, Osterhaus AD, Rimmelzwaan GF. Antigenic drift in the influenza A virus (H3N2) nucleoprotein and escape from recognition by cytotoxic T lymphocytes. J Virol. 2000;74:6800–7. doi: 10.1128/jvi.74.15.6800-6807.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Berkhoff EG, de Wit E, Geelhoed-Mieras MM, Boon AC, Symons J, Fouchier RA, Osterhaus AD, Rimmelzwaan GF. Functional constraints of influenza A virus epitopes limit escape from cytotoxic T lymphocytes. J Virol. 2005;79:11239–46. doi: 10.1128/JVI.79.17.11239-11246.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Berkhoff EG, de Wit E, Geelhoed-Mieras MM, Boon AC, Symons J, Fouchier RA, Osterhaus AD, Rimmelzwaan GF. Fitness costs limit escape from cytotoxic T lymphocytes by influenza A viruses. Vaccine. 2006 doi: 10.1016/j.vaccine.2006.05.051. [DOI] [PubMed] [Google Scholar]
- 14.Boon AC, de Mutsert G, Fouchier RA, Osterhaus AD, Rimmelzwaan GF. The hypervariable immunodominant NP418-426 epitope from the influenza A virus nucleoprotein is recognized by cytotoxic T lymphocytes with high functional avidity. J Virol. 2006;80:6024–32. doi: 10.1128/JVI.00009-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Brown DM, Dilzer AM, Meents DL, Swain SL. CD4 T cell-mediated protection from lethal influenza: perforin and antibody-mediated mechanisms give a one-two punch. J Immunol. 2006;177:2888–98. doi: 10.4049/jimmunol.177.5.2888. [DOI] [PubMed] [Google Scholar]
- 16.Krohn K, Stanescu I, Blazevic V, Vesikari T, Ranki A, Ustav M. A DNA HIV-1 vaccine based on a fusion gene expressing non-structural and structural genes of consensus sequence of the A-C subtypes and the ancestor sequence of the F-H subtypes. Preclinical and clinical studies. Microbes Infect. 2005;7:1405–13. doi: 10.1016/j.micinf.2005.07.023. [DOI] [PubMed] [Google Scholar]
- 17.Malm M, Rollman E, Ustav M, Hinkula J, Krohn K, Wahren B, Blazevic V. Cross-clade protection induced by human immunodeficiency virus-1 DNA immunogens expressing consensus sequences of multiple genes and epitopes from subtypes A, B, C, and FGH. Viral Immunol. 2005;18:678–88. doi: 10.1089/vim.2005.18.678. [DOI] [PubMed] [Google Scholar]
- 18.Weaver EA, Lu Z, Camacho ZT, Moukdar F, Liao HX, Ma BJ, Muldoon M, Theiler J, Nabel GJ, Letvin NL, Korber BT, Hahn BH, Haynes BF, Gao F. Cross-subtype T-cell immune responses induced by a human immunodeficiency virus type 1 group m consensus env immunogen. J Virol. 2006;80:6745–56. doi: 10.1128/JVI.02484-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yan J, Yoon H, Kumar S, Ramanathan M, Corbitt N, Kutzler M, Dai A, Weiner JBD. Enhanced Diversity and Magnitude of Cellular Immune Responses Elicited by a Novel Engineered HIV-1 Subtype B Consensus-based Envelope DNA Vaccine. Molecular Therapy. 2006 doi: 10.1038/sj.mt.6300036. [DOI] [PubMed] [Google Scholar]
- 20.Kim JJ, Nottingham LK, Sin JI, Tsai A, Morrison L, Oh J, Dang K, Hu Y, Kazahaya K, Bennett M, Dentchev T, Wilson DM, Chalian AA, Boyer JD, Agadjanyan MG, Weiner DB. CD8 positive T cells influence antigen-specific immune responses through the expression of chemokines. J Clin Invest. 1998;102:1112–24. doi: 10.1172/JCI3986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kutzler MA, Robinson TM, Chattergoon MA, Choo DK, Choo AY, Choe PY, Ramanathan MP, Parkinson R, Kudchodkar S, Tamura Y, Sidhu M, Roopchand V, Kim JJ, Pavlakis GN, Felber BK, Waldmann TA, Boyer JD, Weiner DB. Coimmunization with an optimized IL-15 plasmid results in enhanced function and longevity of CD8 T cells that are partially independent of CD4 T cell help. J Immunol. 2005;175:112–23. doi: 10.4049/jimmunol.175.1.112. [DOI] [PubMed] [Google Scholar]
- 22.Kumar S, Yan J, Muthumani K, Ramanathan MP, Yoon H, Pavlakis GN, Felber BK, Sidhu M, Boyer JD, Weiner DB. Immunogenicity testing of a novel engineered HIV-1 envelope gp140 DNA vaccine construct. DNA Cell Biol. 2006;25:383–92. doi: 10.1089/dna.2006.25.383. [DOI] [PubMed] [Google Scholar]
- 23.Evolution of H5N1 avian influenza viruses in Asia. Emerg Infect Dis. 2005;11:1515–21. doi: 10.3201/eid1110.050644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yang JS, Kim JJ, Hwang D, Choo AY, Dang K, Maguire H, Kudchodkar S, Ramanathan MP, Weiner DB. Induction of potent Th1-type immune responses from a novel DNA vaccine for West Nile virus New York isolate (WNV-NY1999) J Infect Dis. 2001;184:809–16. doi: 10.1086/323395. [DOI] [PubMed] [Google Scholar]
- 25.Doyle C, Roth MG, Sambrook J, Gething MJ. Mutations in the cytoplasmic domain of the influenza virus hemagglutinin affect different stages of intracellular transport. J Cell Biol. 1985;100:704–14. doi: 10.1083/jcb.100.3.704. [DOI] [PMC free article] [PubMed] [Google Scholar]