

Abstract
Background
Synergy between drugs manifests with increased potency and/or efficacy of the combination relative to either agonist given alone. Synergy is typically observed between drugs of different classes, as is the case with the alpha-adrenergic-opioid receptor synergy often observed in preclinical studies. However, rare studies report synergy between agonists of the same class. The present study examined the analgesic interaction between two intrathecally injected alpha-2 adrenergic receptor (AR) agonists previously thought to act at the same receptor subtype when given spinally.
Methods
Mice were administered clonidine, dexmedetomidine, or the combination spinally to evaluate the interaction between these two agonists. The ED50 values were calculated and the interactions tested by isobolographic analysis. The rotarod test was performed in the same mice following the completion of analgesic assessment to assess motor or sedative effects. These experiments were performed in outbred mice as well as in mice with mutant alpha2A-ARs, alpha2C-AR-knock-out or wildtype controls. Finally, analgesic cross-tolerance between clonidine and dexmedetomidine was evaluated.
Results
Clonidine and dexmedetomidine interacted synergistically in all lines except the alpha2C-AR knockout line, implicating alpha2C-ARs in the interaction. Additionally, clonidine and dexmedetomidine did not show analgesic cross-tolerance in the outbred strain, suggesting that the two drugs have distinct mechanisms of action.
Conclusions
The present study introduces a new synergistic agonist pair, clonidine – dexmedetomidine. These two drugs appear to require the alpha2A-AR for spinal analgesia when given separately; when delivered as a combination, the resultant synergistic interaction requires the alpha2C-AR as well.
Introduction
Synergistic drug interactions result in enhanced potency and/or efficacy when one agent is given together with another. Therapeutic application of synergistic combinations carries the expectation of efficacy at reduced doses and, theoretically, reduced side effects. Although the mechanisms underlying synergistic interactions are not well understood, synergy is thought to result from simultaneous action of the two agents at two distinct sites, such as a common receptor located at disparate anatomical sites or distinct receptors co-residing at a common anatomical location. Examples of well-described synergistic agonist pairs include selective agonists of the mu and delta opioid receptor subtypes as well as either of those subtypes combined with agonists targeting the α2 adrenergic receptors (α2ARs).
The analgesic and anesthetic properties of α2AR-selective agonists have been known for decades. Development of clinical applications of these agonists remains an area of interest, particularly as adjuvants for pain management and as anesthestic-sparing agents1. In contrast to the opioid receptor-selective agonists, definition of each α2AR agonist’s pharmacological profile has been limited due to poor ligand selectivity across the three α2AR subtypes, α2AAR, α2BAR, and α2CAR2. The α2AR subtypes are differentially expressed in specific regions of the central nervous system. For example, in the spinal cord, α2AARs appear to be principally of primary afferent neuron origin whereas α2CARs appear to be expressed primarily on neurons intrinsic to the spinal cord3. The evidence for α2BAR expression in spinal cord nerve terminals and intrinsic spinal neurons is not conclusive. Activation of both α2AARs4,5 and α2CARs6 has been reported to result in antinociception. Therefore, it is reasonable to propose that concurrent participation of α2AARs and α2CARs could result in analgesic synergy. Support for a positive interaction between α2AARs4,5 and α2CARs is provided in a previous report that evaluated interactions between two α2-adrenergic agonists7 that were thought to act at different α2AR subtypes based on differences in the pharmacology of their antagonist-sensitivity. To approach this question systematically, we have initiated a broad evaluation of several α2AR agonist combinations in mouse lines deficient in α2AAR or α2CAR function. As part of this larger program, the present study evaluated the interaction between intrathecally administered clonidine and dexmedetomidine. Prior studies of α2AAR mutant mice have been interpreted to indicate that the potency and/or efficacy of both of these agonists are primarily dependent on α2AAR activation, particularly when administered intrathecally. Because of this prevailing view, we did not expect co-administration of clonidine with dexmedetomidine would result in a synergistic analgesic interaction. Our observations indicate, however, that this combination produces definitive and replicable synergistic analgesia in several separate strains of mice: CD-1 Institute of Cancer Research (ICR) outbred mice, mice deficient in the α2AAR or the α2CAR subtype, and their wild type controls. Further, the potential for cross-tolerance between the agonists was assessed following chronic intrathecal delivery of either agonist. Finally, the interaction between clonidine and dexmedetomidine on a measure of sedation and motor coordination (accelerating rotarod) was also evaluated.
Methods and Materials
Animals
Experimental subjects were 20- to 25-g male ICR mice (Harlan, Madison, WI) or 15- to 20-g male and female mice (gender-matched) with either a mixed C57BL/6-129/Sv genetic background (α2AAR-WT or α2AAR-D79N) or a pure C57BL/6 background (α2CAR-WT or α2CAR-Knock-out (KO)). Animals were maintained on a 12 hour light/dark cycle and had unlimited access to food and water. The α2AAR-D79N mutant mice had been generated by hit-and-run gene targeting as previously described8 on a hybrid C57BL/6-129/Sv background. Wild-type animals of the same mixed background were used as controls (α2AAR-WT). The α2CAR KO mice (α2CAR-KO) had been developed at Stanford University (Palo Alto, California)9 and purchased from Jackson Labs following 17 generations of backcrossing to C57BL/6 background. C57BL/6 mice pair-bred within our facility were used as wild-type controls (α2CAR-WT). Breeding pairs were established and pups were weaned between two and three weeks of age. Within each experiment, animals were age- and gender-matched across groups. Animals were used no more than twice. In each case, a rest period of at least one week was used and the animals were randomized across treatment groups. Although the use of transgenic or knock-out mice may result in compensatory changes, we chose to use these mouse lines because we have extensively characterized their spinal neuropharmacology4–6 and they have been widely used by other groups with interest in α2AR-mediated antinociception and antihypertensive effects (for review, please see Kable and colleagues10). Therefore, the results presented in this study are directly comparable to the prior literature. These experiments were approved by the Institutional Animal Care and Use Committee of the University of Minnesota (Minneapolis, Minnesota). Subjects were housed in groups of 4 in 25 × 48 × 15-cm plastic cages in a temperature- and humidity-controlled environment and maintained on a 12-h light/dark cycle and had free access to food and water.
Chemicals
Clonidine HCl (2-[2,6–dichloroaniline]-2-imidazoline) and Substance P (SP) were purchased from Sigma Chemical Co. (St. Louis, MO). SP was dissolved in acidified saline. Zeneca (Wilmington, DE) donated the dexmedetomidine [(1)-(S)-4-[1-(2,3-dimethylphenyl) ethyl]1H-imidazole]. Clonidine and dexmedetomidine were dissolved in 0.9% saline. All drugs were administered intrathecally by direct lumbar puncture in a 5 µL volume in conscious mice11.
Nociceptive Assay
Nociceptive responsiveness was tested in the SP nociceptive test. The SP assay is a sensitive indicator of milder analgesics12. SP (10–20 ng) was injected intrathecally to produce approximately 40 to 60 behaviors (scratches and bites directed to the hindquarters) in the first minute after injection. The dose of SP required to produce this number of behaviors was confirmed with each new experiment. Coadministration of opioid or adrenergic analgesics dose-dependently inhibits those behaviors13. To test the ability of dexmedetomidine and clonidine to inhibit SP-induced behavior, the drugs were co-administered with SP and inhibition was expressed as a percent of the mean response of the control group (determined with each new experiment) according to the following equation:
% Inhibition = [(control − experimental)/control] × 100.
Sedation/Motor Impairment Assay
In the same mice that received SP stimulation, doses of clonidine, dexmedetomidine, and their combination were tested for impairment of rotorod performance. In such experiments, the animals were trained the day before experimentation to walk 300 seconds on the accelerating rotarod, typically requiring three trials to learn the behavior. The following day, the drugs were administered with SP. Following completion of the 1 minute SP-evoked scratching and biting analysis, the mice were run on the rotarod test.
Motor impairment or sedation was expressed as inhibition of the subjects’ ability to remain on the accelerating rotarod; baseline latencies to fall were typically at or near the cutoff of 300 seconds. Percent inhibition (% Inhibition) was expressed as a percent of the baseline latency of each mouse (determined prior to each new experiment) according to the following equation:
% Inhibition = [(Baseline – experimental)/Baseline] × 100.
Dose-Response Analysis
Individual dose and/or time points are expressed as means with standard error of the mean (SEM). ED50 values and confidence limits were calculated according to the graded dose-response method of Tallarida and Murray14 on the linear portion of each dose-response curve. Statistical comparisons of potencies are based on the confidence limits of the ED50 values. A minimum of three doses were used for each drug or combination of drugs. A minimum of 50% was set for a drug to be classified as efficacious.
Isobolographic Analysis
Dose-response curves were constructed for each agonist administered alone; the ED50 values were calculated and used to determine the potency ratio between the agonists (Example, fig. 1A). This ratio was then maintained when both agonists were administered in combination, a third dose-response curve was constructed and an experimentally derived combination ED50 was calculated. To test for interactions between agonists, the ED50 values and standard error for all dose-response curves were arithmetically arranged around the ED50 value using the equation: (ln(10)×ED50) X (S.E. of log ED50)15. Isobolographic analysis (the “gold standard” for the evaluation of drug interactions14,15) necessitates this manipulation. When testing an interaction between two drugs, a theoretical additive ED50 value is calculated for the combination based on the dose-response curves of each drug administered separately. This theoretical value is then compared by a t-test with the observed experimental ED50 value of the combination. These values are based on the total dose of both drugs. An interaction is considered synergistic if the experimental ED50 is significantly less (p<0.05) than the calculated theoretical additive ED50 value.
Figure 1. Clonidine and dexmedetomidine interact synergistically when given spinally to ICR (Institute of Cancer Research) mice.

A. Clonidine (●) and dexmedetomidine (■) inhibited substance P behavior in a dose-dependent manner. The agonists were then co-administered at a constant clonidine:dexmedetomidine dose ratio of 1:1 (□ clonidine (+dexmedetomidine)) based on the potency ratio between agonists. Note that the combination dose-response curves are plotted as the doses of clonidine used in the presence of dex. The corresponding “Dex (+ Clon)” curve is identical and not shown.. B. Isobolographic analysis applied to the data from Figure 1A. The y-intercept represents the ED50 for clonidine and the x-intercept represents the ED50 for dexmedetomidine. The observed combination ED50 (●) was significantly lower (p<0.05; t-test) than the theoretical additive ED50 (○), indicating that the interaction is synergistic in ICR mice. See Table 1 for ED50 values. Group sizes ranged from 5–8 mice.
Visualization of drug interactions can be facilitated and enhanced by graphical representation of isobolographic analysis. This representation depicts the ED50 of each agent as the x- or y-intercept. For example, figure 1B presents the ED50 of clonidine as the y-intercept and the ED50 of dexmedetomidine as the x-intercept. The line connecting these two points depicts the dose combinations expected to yield 50% efficacy if the interaction is purely additive and is called the theoretical additive line. The theoretical additive ED50 and its confidence interval are determined mathematically and plotted spanning this line. The observed ED50 for the combination is plotted at the corresponding x,y co-ordinates along with its 95% confidence interval for comparison to the theoretical additive ED50. All dose-response and isobolographic analyses were performed with the FlashCalc 4.5.3 pharmacological statistics software package16,17generously supplied by Michael Ossipov, Ph.D. (Professor, University of Arizona, Tucson, Arizona).
Chronic Clonidine or Dexmedetomedine Tolerance Induction
To induce spinal clonidine or dexmedetomidine tolerance, clonidine or dexmedetomidine (10 nmol in 5 µL) was delivered intrathecally once on experimental day 1, and twice daily on experimental days 2 and 3. Repeated injections were separated by at least eight hours. A separate group of mice received an equal number of injections of saline as a control group. On experimental day 4, full dose-response curves were constructed for each agonist in each pretreatment group. The antinociceptive potencies (ED50 values) of clonidine and dexmedetomidine to inhibit SP-evoked behaviors were compared between mice pretreated with saline or clonidine or dexmedetomidine.
Results
Clonidine – Dexmedetomidine Analgesic Synergy
Clonidine produces analgesic synergy with dexmedetomidine in ICR mice
We first determined the potency of each agonist to inhibit SP-evoked behavior in ICR mice. As expected, clonidine and dexmedetomidine inhibited the behavior with comparable potency and efficacy (fig. 1A). The calculated ED50 values of these dose-response curves formed the basis for the equi-effective dose ratios used in the respective combinations (table 1). Co-administration of clonidine with dexmedetomidine resulted in combination dose-response curves shifted approximately 700-fold to the left compared to each agonist given separately (fig. 1, table 1). The isobologram in figure 1B illustrates that the ED50 value of the observed combination differs significantly from the calculated theoretical additive ED50 value indicating a synergistic interaction (fig. 1B, table 1, Student’s t test, p<0.05). This experiment was replicated in a separate group of mice with comparable outcomes (synergism). The robust synergistic interaction of the clonidine-dexmedetomidine combination suggests a second spinal site of action for one of the two agonists.
Table 1.
Summary of Clonidine–Dexmedetomidine Antinociceptive Interactions
| Probe Drug, Intrathecal, nmol | ED50 Clonidine (95% CI) |
ED50 Dexmedetomidine (95% CI) |
Interaction | |
| Figure 1, ICR mice | Single drug | 3.0 (2.1–3.9) | 3.8 (1.3–6.3) | |
| Clonidine–dexmedetomidine, 1:1 ratio | Synergistic | |||
| Observed combination | 0.0045 (0.0001–0.0183)* | |||
| Theoretical additive | 1.7 (1.3–2.1) | |||
| Figure 2, α2AAR-WT | Single drug | 1.2 (0.45–1.9) | 1.0 (0.43–1.7) | |
| Clonidine–dexmedetomidine, 1:1 ratio | Synergistic | |||
| Observed combination | 0.18 (0.1–0.26)* | |||
| Theoretical additive | 0.54 (0.34–0.74) | |||
| Figures 4A and B, α2CAR-WT | Single drug | 1.7 (1.3–2.1) | 1.9 (1.3–2.5) | |
| Clonidine–dexmedetomidine, 1:1 ratio | Synergistic | |||
| Observed combination | 0.16 (0.11–0.23)* | |||
| Theoretical additive | 0.90 (0.73–1.1) | |||
| Figures 4C and D, α2CAR-zKO | Single drug | 5.3 (4.1–6.5) | 4.4 (3.5–5.3) | |
| Clonidine–dexmedetomidine, 1:1 ratio | Subadditive | |||
| Observed combination | 3.6 (2.2–5.0)* | |||
| Theoretical additive | 1.71 (1.3–2.1) | |||
Significant difference from theoretical additive by Student t test, P < 0.05.
AR = adrenergic receptor; CI = confidence interval; ICR = Institute of Cancer Research; KO = knockout; WT = wild type.
Clonidine and dexmedetomidine co-administration in α2AAR-WT mice
The objectives for testing the clonidine-dexmedetomidine combination in α2AAR-WT and α2AAR-D79N mice were 1) to determine whether the synergistic interaction was observable across mouse strains, and 2) to determine whether the combination demonstrated any efficacy in mice lacking α2AAR. Because clonidine consistently demonstrates no efficacy in α2AAR-D79N mice and dexmedetomidine is only efficacious at high doses, the expectation was that the combination would not yield significant efficacy in those mice; nonetheless, it was important to test the possibility that the combination resulted in a significantly different pharmacological profile than either agonist alone. We first determined the potency of each agonist to inhibit SP-evoked behavior in α2AAR-WT mice. As expected, clonidine and dexmedetomidine inhibited the behavior with comparable potency and efficacy (fig. 2A). Co-administration of clonidine with dexmedetomidine resulted in combination dose-response curves shifted about 7-fold to the left compared to each agonist given separately (fig. 2A, table 1). The isobologram in figure 2B illustrates that the ED50 value of the observed combination differs significantly from the calculated theoretical additive ED50 value indicating a synergistic interaction (fig. 2B, table 1, Student’s t test, p<0.05). The synergistic interaction of the clonidine-dexmedetomidine combination in α2AAR-WT mice confirms that the observation was not unique to ICR mice. Although the magnitude of synergism is significantly different (100-fold) across these two strains, the observation of significant synergy for this combination is consistent. This difference also profiles the importance of evaluating combinations across multiple strains. Consistent with our previous reports, neither clonidine nor dexmedetomidine demonstrates antinociceptive efficacy in the α2AAR-D79N mice when given either alone or as a 1:1 combination, even at relatively high doses (10, 30, 100 nmol, intrathecally - fig. 2C).
Figure 2. Clonidine produces antinociceptive synergy with dexmedetomidine in α2AAR-WT (wildtype) mice.
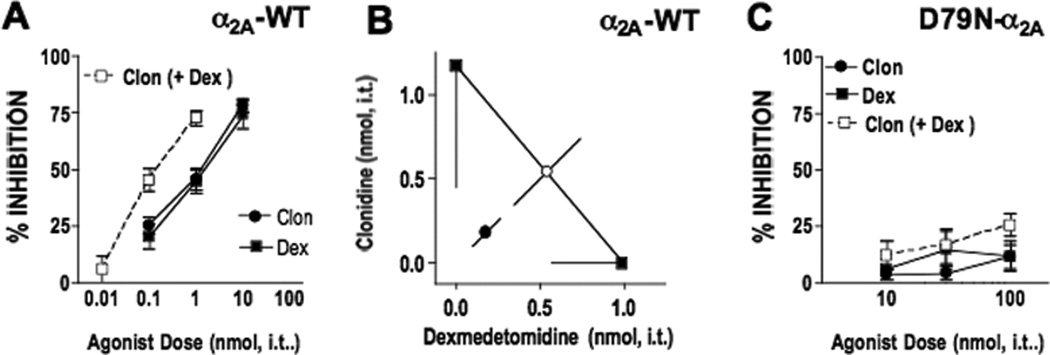
A. Clonidine (●) and dexmedetomidine (■) inhibited substance P behavior in a dose-dependent manner. The agonists were then co-administered at a constant clonidine:dexmedetomidine dose ratio of 1:1 (□ clon (+dex)) based on the potency ratio between agonists. Note that the combination dose-response curves are plotted as the doses of clonidine used in the presence of dex. The corresponding “Dex (+ Clon)” curve is identical and not shown. B. Isobolographic analysis applied to the data from Figure 2A. The y-intercept represents the ED50 for clonidine and the x-intercept represents the ED50 for dexmedetomidine. The observed combination ED50 (●) was significantly lower (p<0.05; t-test) than the theoretical additive ED50 (○), indicating that the interaction is synergistic in α2AAR-WT mice. See Table 1 for ED50 values. C. SP-induced behavior was challenged by intrathecally administered clonidine, dexmedetomidine or both in α2AAR-D79N mice. Neither clonidine (●) nor dexmedetomidine (■) inhibited the behavior. The co-administration of the agonists in a dose ratio of 1:1 (□ clonidine+dexmedetomidine) did not produce appreciable inhibition of the behavior. Group sizes ranged from 5–8 mice.
Mechanism of Clonidine – Dexmedetomidine Analgesic Synergism
Clonidine and dexmedetomidine do not evoke chronic analgesic cross-tolerance
The observation of synergy between clonidine and dexmedetomidine suggests that a receptor other than the α2AAR is involved in the interaction. In situations where two agonists act primarily at the same receptor, chronic administration of one agonist usually elicits cross-tolerance to the other18. Conversely, in cases where two agonists act at different receptor sites, chronic exposure to one agonist typically fails to invoke chronic tolerance to the other (e.g., μ opioid receptor (MOP); δ opioid receptor (DOP)19,20), although minor cross-tolerance is sometimes observed perhaps due to changes in convergent downstream signaling pathways (e.g., MOP-α2AAR20–22). Therefore, to evaluate whether clonidine and dexmedetomidine may act upon the same or different receptors, we conducted an evaluation of analgesic tolerance to clonidine or dexmedetomidine following repeated chronic exposure to spinally administered clonidine (fig. 3A) or dexmedetomidine (fig. 3B) in ICR mice. Whereas three-day spinal pretreatment with clonidine significantly reduced the potency of “probe” doses of clonidine (16-fold tolerance), the analgesic dose-response curve for dexmedetomidine remained largely unchanged (fig. 3A, table 2). Similarly, a three-day spinal pretreatment with dexmedetomedine significantly reduced the potency of “probe” doses of dexmedetomedine (21-fold tolerance), but the analgesic response to clonidine was not significantly altered (fig. 3B, table 2). This lack of cross-tolerance suggests that, despite their apparent shared reliance on spinal α2AARs when given separately, clonidine-evoked or dexmedetomidine-evoked antinociception requires participation of a second distinct receptor.
Figure 3. Chronic intrathecal clonidine or dexmedetomidine do not evoke mutual cross-tolerance.

A. Clonidine Intrathecal Tolerance. The potency of clonidine was significantly reduced in mice pre-treated (Ptx) with repeated injections of clonidine (○) relative to saline-pretreatment (●) indicating the development of analgesic tolerance. In contrast, the potency of dexmedetomidine in mice pre-treated with repeated injections of clonidine (□) did not differ relative to mice pre-treated with saline-pretreatment (■) confirming the lack development of analgesic cross-tolerance. B. Dexmedetomidine (Dex) Intrathecal Tolerance. The potency of dexmedetomidine was significantly reduced in mice pre-treated with repeated injections of dexmedetomidine (□) relative to saline-pretreatment (■) indicating the development of analgesic tolerance. In contrast, the potency of clonidine in mice pre-treated with repeated injections of dexmedetomidine (○) did not differ relative to mice pre-treated with saline-pretreatment (●) confirming the lack development of analgesic cross-tolerance. The ED50 values for the dose-response groups are presented in Table 2. Group sizes were 8 mice dose group.
Table 2.
Summary of Clonidine and Dexmedetomidine Tolerance ED50 Values and Potency Shifts
| Pretreatment Probe | Saline–Clonidine | Clonidine–Clonidine | Tolerance | Saline–Dexmedetomidine | Clonidine–Dexmedetomidine | Cross-tolerance |
| Figure 3A, clonidine tolerance | 0.93 (0.0–1.3) | 16 (14–18)* | 16-Fold reduced potency | 1.33 (1.0–1.8) | 1.26 (0.91–1.73) | No significant potency shift |
| Pretreatment Probe | Saline–Dexmedetomidine | Dexmedetomidine– Dexmedetomidine | Tolerance | Saline–Clonidine | Dexmedetomidine–Clonidine | Cross-tolerance |
| Figure 3B, dexmedetomidine tolerance | 0.71 (0.54–0.93) | 15 (13–17)* | 21-Fold reduced potency | 0.98 (7.3–1.3) | 1.5 (1.1–2.1) | No significant potency shift |
Significant difference in relative potency.
Clonidine produces analgesic synergy with dexmedetomidine in C57Bl/6, but not α2CAR-KO mice
A logical candidate for the second receptor is the α2CAR given its localization in spinal cord and previous studies illustrating that α2CAR activation can result in antinociception6,23. We therefore tested for clonidine-dexmedetomidine synergy in α2CAR-KO mice and their WT controls (C57BL/6 mice). The clonidine-dexmedetomidine combination demonstrated significant analgesic synergy in α2CAR-WT mice (fig. 4A,B, table 1) as was the case in ICR and α2AAR-WT mice. In contrast to the lack of efficacy observed in the α2AAR-D79N mice (fig. 2C), clonidine's and dexmedetomidine's analgesic potency decreased only two- to three-fold (though significantly) in α2CAR-KO mice relative to that in α2CAR-WT mice. These data indicate that, when given separately, neither agonist demonstrates an absolute requirement for the α2CAR (in contrast to that seen in α2AAR mutant mice), but that the α2CAR may participate in the full antinociceptive potential of the two agonists. However, despite this moderate KO effect on the individual dose-response curves of clonidine and dexmedetomidine, the synergistic interaction of their combination was clearly absent in the α2CAR-KO mice (fig. 4C, D and table 1). The potency of the clonidine-dexmedetomidine combination was not altered relative to that of either agonist given alone; the combination ED50 value was significantly higher than that of the theoretical additive ED50 value. This result suggests that the clonidine-dexmedetomidine synergistic interaction requires the presence of α2CARs and that in the absence of α2CARs the two drugs may act at the same receptor, presumably the α2AAR.
Figure 4. Clonidine and dexmedetomidine interact synergistically in α2CAR-WT (wildtype) but not α2CAR-KO (knock-out) mice.

A. Clonidine (●) and dexmedetomidine (■) inhibited the substance P behavior in a dose-dependent manner. The agonists were then co-administered at a constant clonidine:dexmedetomidine dose ratio of 1:1 (□ clon (+dex)) based on the potency ratio between agonists. Note that the combination dose-response curves are plotted as the doses of clonidine used in the presence of dex. The corresponding “Dex (+ Clon)” curve is equivalent. B. Isobolographic analysis applied to the data from Figure 4A. The y-intercept represents the ED50 for clonidine and the x-intercept represents the ED50 for dexmedetomidine. The observed combination ED50 (●) was significantly lower (p<0.05; t-test) than the theoretical additive ED50 (○), indicating that the interaction is synergistic in α2CAR-WT mice. C. SP-induced behavior was challenged by intrathecally administered clonidine, dexmedetomidine or both in α2CAR-KO mice. Clonidine (●) and dexmedetomidine (■) inhibited the behavior in a dose-dependent manner. The agonists were then co-administered at a constant clonidine:dexmedetomidine dose ratio of 1:1 □ clon (+dex)) based on the potency ratio between agonists. Note that the combination dose-response curves are plotted as the doses of clonidine used in the presence of dex. The corresponding “Dex (+ Clon)” curve is identical and not shown. D. Isobolographic analysis applied to the data from Figure 4C. The y-intercept represents the ED50 for clonidine and the x-intercept represents the ED50 for dexmedetomidine. The observed combination ED50 (●) was not significantly (p>0.05; t-test) different from the theoretical additive ED50 (○), indicating that the interaction is additive in α2CAR-KO mice. See Table 1 for ED50 values. Group sizes ranged from 5–8 mice.
Clonidine – dexmedetomidine interactions in other assays
Clonidine-dexmedetomidine interactions in the rotarod assay of sedation and motor impairment
In addition to their analgesic effects, α2AR agonists affect multiple physiological systems, including the central nervous system (sedation, cardiovascular effects, addiction and withdrawal responses). In the present study, the rotarod test, which has been previously used as a measure of both sedation and motor impairment24, was used to assess the sedative and/or motoric effects of the agonists or their combination immediately after SP nociceptive testing. In outbred ICR mice (fig. 5A) clonidine and dexmedetomidine each produced a mild reduction in rotarod performance at the highest dose tested (10 nmol); higher doses were not tested. The clonidine-dexmedetomidine combination reduced rotarod performance only 30% at the highest combination dose (1 nmol of each drug) tested which produced ~90% antinociception (fig. 1A); potentiation was evident at 0.01 and 1 nmol. We distinguish this interaction in rotarod from the synergistic analgesic interaction by referring to the former as potentiation. In α2AAR-WT mice (fig. 5B), clonidine reduced rotarod performance ~70% at the highest dose tested (10 nmol) whereas dexmedetomidine produced only partial reduction (~50%). The combination showed a moderate (< 10-fold) but significant increase in each agonist’s potency when co-administered, the interaction of which was statistically synergistic (isobole not shown). In α2CAR-WT mice (Fig. 5C), both clonidine and dexmedetomidine inhibited rotarod performance at about 10-fold lower potency relative to inhibition of SP behavior. Further, the clonidine-dexmedetomidine combination demonstrates substantially increased potency (~100-fold relative to each given alone) for reduction of rotarod performance. Isobolographic analysis confirmed a significant synergistic interaction (isobole not shown). Figure 5D reflects a minimal effect (< 25%) in α2AAR-D79N mutant mice (consistent with the lack of analgesic effect); Figure 5E also shows moderate (< 50%) rotarod impairment in α2CAR-KO mice. In summary, using the rotarod assay as a model of sedation and/or motor impairment, the clonidine-dexmedetomidine combination resulted in differential pharmacological outcomes across the three lines of mice tested in terms of relative potency and efficacy. Specifically, while the combination significantly impaired rotarod performance in the C57Bl/6 line, it impaired motor performance only moderately in the ICR line; the effect in the α2AAR-WT line was intermediate. These results that differ across mouse lines contrast with the concurrent antinociceptive measures in that the antinociceptive potency and efficacy and synergism were consistent (albeit of differing magnitude) across all wildtype mouse lines. Further analysis of other potential side effects of the combination in mice and other species will be needed to determine the utility of the clonidine-dexmedetomidine combination in pain management or anesthesia.
Figure 5. Clonidine- and dexmedetomidine-induced sedation/motor impairment.

A–E. Rotarod performance was challenged by intrathecally administered clonidine, dexmedetomidine or both in ICR (A), α2AAR-WT (B), α2CAR-WT (C), α2AAR-D79N (D) and α2CAR-KO mice (E). Neither clonidine (●), dexmedetomidine (■) nor the 1:1 combination (□) exhibited greater than 40% efficacy up to the highest doses used in the SP test. The combination did, however, result in the production of this modest efficacy at lower doses, representing significant potentiation. B. In α2AAR-WT mice, clonidine exhibited full efficacy at 10 nmol (●) whereas dexmedetomidine's maximum efficacy at that dose fell short of 50% (■). The 1:1 combination (□) dose-response curve shifted significantly to the left (~10-fold) relative to clonidine alone with comparable efficacy and the interaction was found to be synergistic (isobologram not shown; p<0.05; t-test). C. In α2CAR-WT mice both clonidine and dexmedetomidine inhibited rotarod performance with full efficacy. The 1:1 combination (□) dose-response curve shifted significantly to the left (~100-fold) relative to either drug alone with comparable efficacy and the interaction was found to be synergistic (isobologram not shown; p<0.05; t-test). D. In α2AAR-D79N mice, neither clonidine (●), dexmedetomidine (■) nor the 1:1 combination (◊) reduced rotarod performance more than 30%. E. α2CAR-KO mice, neither clonidine (●), dexmedetomidine (■) nor the 1:1 combination (□) reduced rotarod performance more than 50%. Group sizes were 5 mice/group. Abbreviations: CLON, clonidine; DEX, dexmedetomidine. ICR, Institute of Cancer Research; KO, knock-out; WT, wildtype.
Discussion
The present study reveals that two spinally active α2 adrenergic analgesics, clonidine and dexmedetomidine, interact synergistically in the production of antinociception in mice. These two agonists have previously been thought to act primarily on α2AARs to exert their various physiological effects10,25. Because clonidine requires α2AAR5 and dexmedetomidine's analgesic potency is dramatically reduced in mice in the absence of functional α2AARs4, the observation of synergism was an unexpected and novel finding. Upon further investigation in the present study, the participation of a second target has become apparent, likely to be α2CARs. The concept of α2CAR as a synergistic partner with α2AARs is supported by previous anatomical3 and pharmacological7 evidence.
Synergistic analgesic pairs
Historically, synergistic analgesic partners have implicated the activation of two distinct receptors or receptor subtypes. Opioid receptor pairs with synergistic interactions include MOP-DOP and MOP-KOP26; both pairings involve agonists acting at separate receptor subtypes in the same G protein-coupled receptor family (opioid). Others have demonstrated synergy between agonists that activate receptors in different G protein-coupled receptor families: examples include MOP and α2AR agonists27,28, DOP and α2AAR agonists4,13, and DOP and α2CAR agonists4. Studies evaluating interactions between agonists acting on the same opioid receptor subtype have only reported additive interactions29.
One previous report studied the interactions between two α2AR agonists, dexmedetomidine and ST917. The rationale for assessing that combination for synergy derived from observations that, whereas dexmedetomidine had been largely thought to activate α2AAR, ST91 appeared to be independent of α2AARs. These assertions did not derive from binding studies because the affinities of these ligands do not differ appreciably among α2AR subtypes. Rather, the proposed selectivity was derived from pharmacological studies using antagonists with differential affinity for the three receptor subtypes30,31. Dexmedetomidine's selectivity was subsequently validated by studies using genetically altered mice32, but ST91 did not show substantial dependence on either α2A or α2CARs in genetically altered mice32. However, the observation that synergy was detected between dexmedetomidine and ST91 is consistent with the participation of two distinct receptor subtypes. The distinct localizations of α2AAR (thought to be restricted to the spinal terminals of primary afferent neurons) and α2CAR (thought to be restricted to spinal neurons)3 in spinal cord positions this pair to operate in such a synergistic manner.
Clonidine-Dexmedetomidine Analgesic Synergy
The total lack of clonidine efficacy in α2AAR functional knock-out mice suggested that clonidine acts only at α2AARs to produce antinociception5. Although dexmedetomidine's potency was dramatically reduced in the same mice, dexmedetomidine retained analgesic efficacy, albeit at thousand-fold higher doses4. This distinction between clonidine and dexmedetomidine leaves open the possibility that the latter acts on another adrenergic receptor, such as the α2CAR. It is also conceivable that clonidine acts upon α2CAR with an effect below the threshold of detection in our nociceptive assay. These possibilities in turn suggested that clonidine-dexmedetomidine synergy may result from the participation of both α2AARs and α2CARs. Two experimental tests of this hypothesis yielded concurrent results. First, cross-tolerance did not occur between clonidine and dexmedetomidine, indicating that the two agonists act at different receptors when given as a combination. Second, clonidine-dexmedetomidine synergy was not observed in α2CAR-KO mice but did occur in WT mice. Therefore, whereas clonidine and dexmedetomidine given separately by the intrathecal route appear to rely primarily on activation of α2AAR, their spinal synergistic interaction requires the recruitment of α2CAR as well. Activity at both receptors is consistent with competition binding studies where both agonists bind with comparable affinity to both receptors2,33. However, competition binding studies are incongruent with functional assays (e.g., GTPγS binding) in transfected cell lines where dexmedetomidine has shown a rank order preference for α2BAR>α2CAR>α2AAR and clonidine was a partial agonist at α2BAR>α2AAR and inactive at α2CAR34. It is clear that in vitro binding or functional studies may not model the in vivo condition adequately. Furthermore, the participation of α2CAR may not be at the level of direct agonist-receptor interaction but rather could represent an indirect contribution within a more complex pathway. The present study indicates that the efficacy of single agonists delivered spinally may not adequately predict the efficacy, potency or mechanism of combined agonists given spinally.
Interaction studies of sedation and motor impairment
Assessing the analgesic utility of the clonidine-dexmedetomidine combination warrants determination of the effects of the combination on at least one non-analgesic dependent measure. Accordingly, sedation and motor impairment were assessed using the accelerating rotarod test immediately after antinociceptive testing. Unlike the antinociceptive measure, the sedative efficacy of the agonists and their combination varied across the strains studied. The individual agonists produced moderate (less than 50%) sedation in the outbred ICR strain, intermediate effects in the α2AAR-WT (mixed strain: C57BL/6-129sv), and pronounced sedation in the α2CAR-WT (inbred strain: C57BL/6). Interestingly, the individual agonists produced minimal sedation in the two mutant lines of mice, α2AAR-D79N and α2CAR-KO, indicating that both receptor subtypes contribute to the sedative effects. The clonidine-dexmedetomidine combination showed a small sedative effect at lower doses in ICR mice, synergistic sedation in both α2AAR-WT and α2CAR-WT mice, and minimal to no sedation in both α2AAR-D79N and α2CAR-KO mice. Interestingly, a prior study of α2AAR-D79N heterozygous mice revealed a clear difference between the antihypertensive and sedative effects of dexmedetomidine; dexmedetomidine's cardiovascular effects were fully manifest in heterozygous α2AAR-D79N mice, whereas its sedative effect was absent35. The authors attributed this difference in response to a different receptor occupancy requirement for lowering blood pressure versus sedation. They postulated that partial α2AAR agonists might provide that separation of effect in WT mice, and in fact observed a similar separation of effects in WT mice with the partial agonist moxonidine. Conceivably, the separation of analgesia and sedation in the outbred ICR strain results from a similar partial agonist character of the clonidine-dexmedetomidine combination. Whereas dexmedetomidine is considered a full agonist at both α2A and α2CARs, clonidine is considered a partial agonist at both2. We speculate that the relationship between receptor occupancy and sedation could be a strain-dependent effect and account for the difference in sedative effects in the strains studied; however, further testing is required to address this hypothesis. Further study is needed to refine the combination to optimize clinical outcomes for either analgesia with moderate sedation or improved sedative/anesthetic efficacy, depending on the target therapeutic application.
Clinical Relevance
Clinical application of interdrug synergy between G protein-coupled receptor agonists carries the potential for reduced dose and side effect profiles of drug combinations compared to the drugs given alone. There is an expectation that the dose reduction enabled by a synergistic interaction might reduce side effects. The utility of clonidine as a monotherapy36–41 or combined with spinal opioids42,43 and/or local anesthetics has been studied for decades44,45. Although the primary clinical use of dexmedetomidine has been as a sedative and anesthetic agent46,47, the combination of intrathecal dexmedetomidine with bupivacaine has recently been shown to be effective for analgesic control, comparing favorably with the combination of intrathecal clonidine and bupivacaine48. Further, a recent case report documents the use of intrathecal dexmedetomidine combined with morphine to restore analgesic control in a morphine-tolerant cancer patient49. Thus, both clonidine and dexmedetomidine produce antinociception when given intrathecally both in animal models4,5,50–52 and humans36,48,49.
However, prior to clinical application of any single agent or novel combination of spinal analgesics, the conduct of preclinical animal neurotoxicity studies53 and controlled clinical trials to establish safety of the singly delivered agents54 and the synergistic combinations (a requisite separate study from that of the singly delivered agents)55 is imperative56. The importance of neurotoxicity evaluation of potential neuraxial therapeutics cannot be overemphasized57,58. Whereas the safety profile of intrathecally delivered clonidine has been previously established53, the neurotoxicity of intrathecally delivered dexmedetomedine is largely unknown. A recent evaluation59 of toxicity of epidurally delivered commercial dexmedetomedine formulation in rabbits found white matter demyelination in the spinal cord, potentially attributable to the pH (4.5–7.0) of the current formulation. For the novel combination of clonidine-dexmedetomidine to be considered useful for application, substantially more work would be needed54. A further consideration is that the anatomical organization of α2ARs subtypes, while well defined in rodent, has not been evaluated in human spinal cord. Differences between species in receptor subtype expression pattern in the spinal cord could ultimately account for differences in agonist combination interactions. Isobolographic analysis of a combination (fentanyl-clonidine) well established to be synergistic in rodents did not demonstrate statistically significant synergism in one clinical evaluation42; the reason for the difference between rodent and human is not clear. Regardless of these considerations, the current study reveals an unexpected interaction between two α2AR agonists and suggests further evaluation of other α2AR agonists as potentially useful synergistic partners.
Conclusion
Application of interdrug synergy between G protein-coupled receptor agonists carries the potential for reduced dose and side effect profiles of drug combinations compared to the drugs given alone. The potential of such positive interactions encourages the continued search for novel useful combinations. The opportunities of therapeutic application of α2AR agonists either as single agents or as combinations (particularly with opioids and local anesthetics) continues to expand with recent clinical studies1. In the present study spinally coadministered clonidine and dexmedetomidine demonstrated a replicable and consistent synergistic interaction that was not predicted by prior pharmacological studies of the agonists in genetic knock-out mice. The application of isobolographic analysis to this unexpected combination in genetic knock-out mice revealed an interaction between α2AAR and α2CAR that would be otherwise difficult to identify60. Therefore, the combination of these two agonists or other co-activators of this α2A-α2CAR pair may have utility in the fields of pain management and sedative anesthesia.
ACKNOWLEDGMENTS
We extend our great appreciation to Zeneca Pharmaceuticals (Wilmington, Delware) for the gift of dexmedetomidine.
SUPPORT: This work was conducted with the support of the National Institute on Drug Abuse (Bethesda, Maryland), R01-DA-15438 to Dr. Wilcox and K01-DA-00509 to Dr. Fairbanks.
Footnotes
Work Attributed to Departments of Pharmaceutics, Pharmacology, and Neuroscience at the University of Minnesota.
References
- 1.Sanders RD, Maze M. Alpha2-adrenoceptor agonists. Curr Opin Investig Drugs. 2007;8:25–33. [PubMed] [Google Scholar]
- 2.Jasper JR, Lesnick JD, Chang LK, Yamanishi SS, Chang TK, Hsu SA, Daunt DA, Bonhaus DW, Eglen RM. Ligand efficacy and potency at recombinant alpha2 adrenergic receptors: agonist-mediated [35S]GTPgammaS binding. Biochem Pharmacol. 1998;55:1035–1043. doi: 10.1016/s0006-2952(97)00631-x. [DOI] [PubMed] [Google Scholar]
- 3.Stone LS, Broberger C, Vulchanova L, Wilcox GL, Hokfelt T, Riedl MS, Elde R. Differential distribution of alpha2A and alpha2C adrenergic receptor immunoreactivity in the rat spinal cord. J. Neurosci. 1998;18:5928–5937. doi: 10.1523/JNEUROSCI.18-15-05928.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stone LS, MacMillan LB, Kitto KF, Limbird LE, Wilcox GL. The alpha2a adrenergic receptor subtype mediates spinal analgesia evoked by alpha2 agonists and is necessary for spinal adrenergic-opioid synergy. J Neurosci. 1997;17:7157–7165. doi: 10.1523/JNEUROSCI.17-18-07157.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fairbanks CA, Wilcox GL. Moxonidine, a selective alpha2-adrenergic and imidazoline receptor agonist, produces spinal antinociception in mice. J Pharmacol Exp Ther. 1999;290:403–412. [PubMed] [Google Scholar]
- 6.Fairbanks CA, Stone LS, Kitto KF, Nguyen HO, Posthumus IJ, Wilcox GL. alpha(2C)-Adrenergic receptors mediate spinal analgesia and adrenergic-opioid synergy. J Pharmacol Exp Ther. 2002;300:282–290. doi: 10.1124/jpet.300.1.282. [DOI] [PubMed] [Google Scholar]
- 7.Graham BA, Hammond DL, Proudfit HK. Synergistic interactions between two alpha(2)-adrenoceptor agonists, dexmedetomidine and ST-91, in two substrains of Sprague-Dawley rats. Pain. 2000;85:135–143. doi: 10.1016/s0304-3959(99)00261-4. [DOI] [PubMed] [Google Scholar]
- 8.MacMillan LB, Hein L, Smith MS, Piascik MT, Limbird LE. Central hypotensive effects of the alpha2a-adrenergic receptor subtype. Science. 1996;273:801–803. doi: 10.1126/science.273.5276.801. [DOI] [PubMed] [Google Scholar]
- 9.Link RE, Stevens MS, Kulatunga M, Scheinin M, Barsh GS, Kobilka BK. Targeted inactivation of the gene encoding the mouse alpha 2c-adrenoceptor homolog. Mol Pharmacol. 1995;48:48–55. [PubMed] [Google Scholar]
- 10.Kable JW, Murrin LC, Bylund DB. In vivo gene modification elucidates subtype-specific functions of alpha(2)-adrenergic receptors. J Pharmacol Exp Ther. 2000;293:1–7. [PubMed] [Google Scholar]
- 11.Hylden JLK, Wilcox GL. Intrathecal morphine in mice: a new technique. Eur. J. Pharmacol. 1980;67:313–316. doi: 10.1016/0014-2999(80)90515-4. [DOI] [PubMed] [Google Scholar]
- 12.Hylden JLK, Wilcox GL. Intrathecal substance P elicits a caudally-directed biting and scratching behavior in mice. Brain Res. 1981;217:212–215. doi: 10.1016/0006-8993(81)90203-1. [DOI] [PubMed] [Google Scholar]
- 13.Roerig SC, Lei S, Kitto K, Hylden JK, Wilcox GL. Spinal interactions between opioid and noradrenergic agonists in mice: multiplicativity involves delta and alpha-2 receptors. J Pharmacol Exp Ther. 1992;262:365–374. [PubMed] [Google Scholar]
- 14.Tallarida RJ, Murray RB. Manual of pharmacological calculations with computer programs. New York: Springer Verlag; 1987. pp. 26–231. [Google Scholar]
- 15.Tallarida RJ. Statistical analysis of drug combinations for synergism. Pain. 1992;49:93–97. doi: 10.1016/0304-3959(92)90193-F. [DOI] [PubMed] [Google Scholar]
- 16.Xie JY, Herman DS, Stiller CO, Gardell LR, Ossipov MH, Lai J, Porreca F, Vanderah TW. Cholecystokinin in the rostral ventromedial medulla mediates opioid-induced hyperalgesia and antinociceptive tolerance. J Neurosci. 2005;25:409–416. doi: 10.1523/JNEUROSCI.4054-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Quartilho A, Mata HP, Ibrahim MM, Vanderah TW, Ossipov MH, Lai J, Porreca F, Malan TP., Jr Production of paradoxical sensory hypersensitivity by alpha 2-adrenoreceptor agonists. Anesthesiology. 2004;100:1538–1544. doi: 10.1097/00000542-200406000-00029. [DOI] [PubMed] [Google Scholar]
- 18.Moulin DE, Ling GSf, Pasternak GW. Unidirectional analgesic cross-tolerance between morphine and levorphanol in the rat. PAIN. 1988;33:233–239. doi: 10.1016/0304-3959(88)90095-4. [DOI] [PubMed] [Google Scholar]
- 19.Stevens CW, Yaksh TL. Studies of morphine and D-ala2-D-leu5-enkephalin (DADLE) cross-tolerance after continuous intrathecal infusion in the rat. ANSTHESIOLOGY. 1992;76:596–603. doi: 10.1097/00000542-199204000-00017. [DOI] [PubMed] [Google Scholar]
- 20.Kalso EA, Sullivan AF, McQuay HJ, Dickenson AH, Roques BP. Cross-tolerance between mu opioid and alpha-2 adrenergic receptors, but not between mu and delta opioid recetpros in the spinal cord of the rat. J. Pharmacol. Exp. Ther. 1993;265:551–558. [PubMed] [Google Scholar]
- 21.Stevens CW, Monasky MS, Yaksh TL. Spinal infusion of opiate and alpha-2 agonists in rats: tolerance and cross-tolerance studies. J. Pharmacol. Exp. Ther. 1988;244:63–70. [PubMed] [Google Scholar]
- 22.Paul D, Tran JG. Differential cross-tolerance between analgesia produced by alpha2-adrenocceptor agonists and receptor subtype selective opioid treatements. Eur. J. Pharmacol. 1995;272:111–114. doi: 10.1016/0014-2999(94)00695-4. [DOI] [PubMed] [Google Scholar]
- 23.Aley KO, Levine JD. Multiple receptors involved in peripheral alpha 2, mu, and A1 antinociception, tolerance, and withdrawal. J Neurosci. 1997;17:735–744. doi: 10.1523/JNEUROSCI.17-02-00735.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Van der Laan JW, Van Veenendaal W, Voorthuis P, Weick G, Hillen FC. The effects of centrally acting adrenergic agonists on temperature and on explorative and motor behaviour. Relation with effects on quasi-morphine withdrawal behaviour. Eur J Pharmacol. 1985;107:367–373. doi: 10.1016/0014-2999(85)90264-x. [DOI] [PubMed] [Google Scholar]
- 25.Guyenet PG. Is the hypotensive effect of clonidine and related drugs due to imidazoline binding sites? Am J Physiol. 1997;273:R1580–R1584. doi: 10.1152/ajpregu.1997.273.5.R1580. [DOI] [PubMed] [Google Scholar]
- 26.Miaskowski C, Sutters KA, Taiwo YO, Levine JD. Antinociceptive and motor effects of delta/mu and kappa/mu combinations of intrathecal opioid agonists. Pain. 1992;49:137–144. doi: 10.1016/0304-3959(92)90200-U. [DOI] [PubMed] [Google Scholar]
- 27.Ossipov MH, Harris S, Lloyd P, Messineo E. An isobolographic analysis of the antinociceptive effect of systemically and intrathecally administered combinations of clonidine and opiates. J Pharmacol Exp Ther. 1990;255:1107–1116. [PubMed] [Google Scholar]
- 28.Ossipov MH, Harris S, Lloyd P, Messineo E, Lin BS, Bagley J. Antinociceptive interaction between opioids and medetomidine: systemic additivity and spinal synergy. Anesthesiology. 1990;73:227–235. doi: 10.1097/00000542-199012000-00022. [DOI] [PubMed] [Google Scholar]
- 29.Pavlovic ZW, Bodnar RJ. Opioid supraspinal analgesic synergy between the amygdala and periaqueductal gray in rats. Brain Research. 1998;779:158–169. doi: 10.1016/s0006-8993(97)01115-3. [DOI] [PubMed] [Google Scholar]
- 30.Takano Y, Yaksh TL. Characterization of the pharmacology of intrathecally administered alpha-2 agonists and antagonists in rats. J Pharmacol Exp Ther. 1992;261:764–772. [PubMed] [Google Scholar]
- 31.Takano Y, Yaksh TL. Chronic spinal infusion of dexmedetomidine, ST-91 and clonidine: spinal alpha 2 adrenoceptor subtypes and intrinsic activity. J Pharmacol Exp Ther. 1993;264:327–335. [PubMed] [Google Scholar]
- 32.Stone LS, Kitto KF, Eisenach JC, Fairbanks CA, Wilcox GL. ST91 [2-(2,6-diethylphenylamino)-2-imidazoline hydrochloride]-mediated spinal antinociception and synergy with opioids persists in the absence of functional alpha-2A- or alpha-2C-adrenergic receptors. J Pharmacol Exp Ther. 2007;323:899–906. doi: 10.1124/jpet.107.125526. [DOI] [PubMed] [Google Scholar]
- 33.Piletz JE, Zhu H, Chikkala DN. Comparison of ligand binding affinities at human I-1-Imidazoline binding sites and the high affinity state of alpha-2 adrenoceptor subtypes. J Pharmacol Exp Ther. 1996;279:694–702. [PubMed] [Google Scholar]
- 34.Jansson CC, Pohjanoksa K, Lang J, Wurster S, Savola JM, Scheinin M. Alpha2-adrenoceptor agonists stimulate high-affinity GTPase activity in a receptor subtype-selective manner. Eur J Pharmacol. 1999;374:137–146. doi: 10.1016/s0014-2999(99)00306-4. [DOI] [PubMed] [Google Scholar]
- 35.Tan CM, Wilson MH, MacMillan LB, Kobilka BK, Limbird LE. Heterozygous alpha(2A)-adrenergic receptor mice unveil unique therapeutic benefits of partial agonists. Proceedings of the National Academy of Sciences of the United States of America. 2002;99:12471–12476. doi: 10.1073/pnas.122368499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Eisenach JC, De Kock M, Klimscha W. alpha(2)-adrenergic agonists for regional anesthesia. A clinical review of clonidine (1984–1995) Anesthesiology. 1996;85:655–674. doi: 10.1097/00000542-199609000-00026. [DOI] [PubMed] [Google Scholar]
- 37.Eisenach JC, DuPen S, Dubios M, Miguel R, Allin D. Epidural clonidine analgesia for intractable cancer pain. Pain. 1995;61:391–399. doi: 10.1016/0304-3959(94)00209-W. [DOI] [PubMed] [Google Scholar]
- 38.Eisenach JC, Dewan DM. Intrathecal clonidine in obstetrics: sheep studies. Anesthesiology. 1990;72:663–668. doi: 10.1097/00000542-199004000-00015. [DOI] [PubMed] [Google Scholar]
- 39.Mendez R, Eisenach JC, Kashtan K. Epidural clonidine analgesia after cesarean section. Anesthesiology. 1990;73:848–852. doi: 10.1097/00000542-199011000-00009. [DOI] [PubMed] [Google Scholar]
- 40.Eisenach JC, Lysak SZ, Viscomi CM. Epidural clonidine analgesia following surgery: phase I. Anesthesiology. 1989;71:640–646. doi: 10.1097/00000542-198911000-00003. [DOI] [PubMed] [Google Scholar]
- 41.Eisenach JC, Rauck RL, Buzzanell C, Lysak SZ. Epidural clonidine analgesia for intractable cancer pain: phase I. Anesthesiology. 1989;71:647–652. doi: 10.1097/00000542-198911000-00004. [DOI] [PubMed] [Google Scholar]
- 42.Eisenach JC, D'Angelo R, Taylor C, Hood DD. An isobolographic study of epidural clonidine and fentanyl after cesarean section. Anesth Analg. 1994;79:285–290. doi: 10.1213/00000539-199408000-00014. [DOI] [PubMed] [Google Scholar]
- 43.Siddall PJ, Molloy AR, Walker S, Mather LE, Rutkowski SB, Cousins MJ. The efficacy of intrathecal morphine and clonidine in the treatment of pain after spinal cord injury. Anesth Analg. 2000;91:1493–1498. doi: 10.1097/00000539-200012000-00037. [DOI] [PubMed] [Google Scholar]
- 44.Sites BD, Beach M, Biggs R, Rohan C, Wiley C, Rassias A, Gregory J, Fanciullo G. Intrathecal clonidine added to a bupivacaine-morphine spinal anesthetic improves postoperative analgesia for total knee arthroplasty. Anesth Analg. 2003;96:1083–1088. doi: 10.1213/01.ANE.0000055651.24073.59. [DOI] [PubMed] [Google Scholar]
- 45.Walker SM, Goudas LC, Cousins MJ, Carr DB. Combination spinal analgesic chemotherapy: a systematic review. Anesth Analg. 2002;95:674–715. doi: 10.1097/00000539-200209000-00033. [DOI] [PubMed] [Google Scholar]
- 46.Belleville JP, Ward DS, Bloor BC, Maze M. Effects of intravenous dexmedetomidine in humans. I. Sedation, ventilation, and metabolic rate. Anesthesiology. 1992;77:1125–1133. doi: 10.1097/00000542-199212000-00013. [DOI] [PubMed] [Google Scholar]
- 47.Kamibayashi T, Maze M. Clinical uses of alpha2-adrenergic agonists. Anesthesiology. 2000;93:1345–1349. doi: 10.1097/00000542-200011000-00030. [DOI] [PubMed] [Google Scholar]
- 48.Kanazi GE, Aouad MT, Jabbour-Khoury SI, Al Jazzar MD, Alameddine MM, Al-Yaman R, Bulbul M, Baraka AS. Effect of low-dose dexmedetomidine or clonidine on the characteristics of bupivacaine spinal block. Acta Anaesthesiol Scand. 2006;50:222–227. doi: 10.1111/j.1399-6576.2006.00919.x. [DOI] [PubMed] [Google Scholar]
- 49.Ugur F, Gulcu N, Boyaci A. Intrathecal infusion therapy with dexmedetomidine-supplemented morphine in cancer pain. Acta Anaesthesiol Scand. 2007;51:388. doi: 10.1111/j.1399-6576.2006.01202.x. [DOI] [PubMed] [Google Scholar]
- 50.Kalso EA, Poyhia R, Rosenberg PH. Spinal antinociception by dexmedetomidine, a highly selective alpha 2-adrenergic agonist. Pharmacol Toxicol. 1991;68:140–143. doi: 10.1111/j.1600-0773.1991.tb02052.x. [DOI] [PubMed] [Google Scholar]
- 51.Post C, Gordh T, Jr, Minor BG, Archer T, Freedman J. Antinociceptive effects and spinal cord tissue concentrations after intrathecal injection of guanfacine or clonidine into rats. Anesth Analg. 1987;66:317–324. [PubMed] [Google Scholar]
- 52.Takano Y, Yaksh TL. Relative efficacy of spinal alpha-2 agonists, dexmedetomidine, clonidine and ST-91, determined in vivo by using N-ethoxycarbonyl-2-ethoxy-1,2-dihydroquinoline, an irreversible antagonist. J Pharmacol Exp Ther. 1991;258:438–446. [PubMed] [Google Scholar]
- 53.Yaksh TL, Collins JG. Studies in animals should precede human use of spinally administered drugs. Anesthesiology. 1989;70:4–6. [PubMed] [Google Scholar]
- 54.Yaksh TL, Allen JW. The use of intrathecal midazolam in humans: a case study of process. Anesth Analg. 2004;98:1536–1545. doi: 10.1213/01.ANE.0000122638.41130.BF. [DOI] [PubMed] [Google Scholar]
- 55.Hood DD, Eisenach JC, Tong C, Tommasi E, Yaksh TL. Cardiorespiratory and spinal cord blood flow effects of intrathecal neostigmine methylsulfate, clonidine, and their combination in sheep. Anesthesiology. 1995;82:428–435. doi: 10.1097/00000542-199502000-00013. [DOI] [PubMed] [Google Scholar]
- 56.Chiari A, Eisenach JC. Spinal anesthesia: mechanisms, agents, methods, and safety. Reg Anesth Pain Med. 1998;23:357–362. doi: 10.1016/s1098-7339(98)90006-2. [DOI] [PubMed] [Google Scholar]
- 57.Eisenach JC, Yaksh TL. Safety in numbers: How do we study toxicity of spinal analgesics? Anesthesiology. 2002;97:1047–1049. doi: 10.1097/00000542-200211000-00003. [DOI] [PubMed] [Google Scholar]
- 58.Eisenach JC, James FM, 3rd, Gordh T, Jr, Yaksh TL. New epidural drugs: primum non nocere. Anesth Analg. 1998;87:1211–1212. doi: 10.1097/00000539-199811000-00048. [DOI] [PubMed] [Google Scholar]
- 59.Konakci S, Adanir T, Yilmaz G, Rezanko T. The efficacy and neurotoxicity of dexmedetomidine administered via the epidural route. Eur J Anaesthesiol. 2008;25:403–409. doi: 10.1017/S0265021507003079. [DOI] [PubMed] [Google Scholar]
- 60.Tallarida RJ, Porreca F, Cowan A. Statistical analysis of drug-drug and site-site interactions with isobolograms. Life Sci. 1989;45:947–961. doi: 10.1016/0024-3205(89)90148-3. [DOI] [PubMed] [Google Scholar]