

Abstract
AIM: To study the protective effects and mechanisms of Se-enriched lactobacillus on liver injury caused by carbon tetrachloride (CCl4) in mice.
METHODS: Seventy-two ICR mice were randomly divided into four groups: normal group, CCl4-induced model group, low Se-enriched lactobacillus treatment group (L-Se group), and high Se-enriched lactobacillus treatment group (H-Se group). During a 3-wk experimental period, the common complete diet was orally provided daily for normal group and model group, and the mice in L-Se and H-Se groups were given a diet with 2 and 4 mg of organoselenium from Se-enriched lactobacillus per kg feed, respectively. From the 2nd wk of experiment, the model group, L-Se group, and H-Se group received abdominal cavity injection of olive oil solution containing 500 mL/L CCl4 (0.07 mL/100 g body mass) to induce liver injury, and the normal group was given olive oil on every other day for over 2 wk. In the first 2 wk post injection with CCl4, mice in each group were killed. The specimens of blood, liver tissue, and macrophages in abdominal cavity fluid were taken. Then the activities of the following liver tissue injury-associated enzymes including glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), alanine aminotransferase (ALT) and aspartate aminotransferase (AST) as well as malondialdehyde (MDA) content were assayed. Changes of phagocytic rate and phagocytic index in macrophages were observed with Wright-Giemsa stain. Plasma TNF-α level was measured by radioimmunoassay. The level of intracellular free Ca2+ ([Ca2+]i) in hepatocytes was detected under a laser scanning confocal microscope.
RESULTS: During the entire experimental period, the AST and ALT activities in liver were greatly enhanced by CCl4 and completely blunted by both low and high doses of Se-enriched lactobacillus. The Se-enriched lactobacillus-protected liver homogenate GSH-Px and SOD activities were higher or significantly higher than those in model group and were close to those in normal group. CCl4 significantly increased MDA content in liver homogenates, while administration of Se-enriched lactobacillus prevented MDA elevation. Phagocytic rate and phagocytic index of macro-phages decreased after CCl4 treatment compared to those in normal control, but they were dramatically rescued by Se-enriched lactobacillus, showing a greatly higher phagocytic function compared to model group. CCl4 could significantly elevate plasma TNF-α and hepatocyte [Ca2+]ilevel, which were also obviously prevented by Se-enriched lactobacillus.
CONCLUSION: Se-enriched lactobacillus can intervene in CCl4-induced liver injury in mice by enhancing macrophage function activity to keep normal and beneficial effects, elevating antioxidant-enzyme activities and reducing lipid peroxidation reaction, inhibiting excessive release of TNF-α, preventing the dramatic elevation of [Ca2+]i in hepatocytes.
Keywords: Se-enriched lactobacillus, Liver injury, Carbon tetrachloride, Macrophage, Ca2+
INTRODUCTION
Hepatic fibrosis represents the response of the liver to diverse chronic insults such as parasitic disease, chronic viral infection (hepatitis B and C), immunologic attack (autoimmune hepatitis), hereditary metal overload, toxic damage, etc. Because of the worldwide prevalence of these insults, liver fibrosis is a common pathological process of hepatic disease, leading to the development of irreversible cirrhosis[1,2]. If treated properly at fibrosis stage, cirrhosis could be prevented[3]. However, there are no effective anti-fibrosis drugs till date. Selenium (Se), an essential nutrient element, can prevent the malignant transformation of normal cells and the activation of oncogenes with anticarcinogenic effects within a physiological dosage range[4,5]. It has been found in clinical investigations that the blood selenium of hepatocarcinoma patients is low, and the progress of liver injury and carcinomatous change are related to the degree of selenium deficiency[6]. In addition, some studies indicate that selenium is closely related to the inhibition of hepatic fibrosis[7,8]. However, the supplement of selenium is relevant to selenium style and dosage[5]. Se-enriched lactobacillus is an organoselenium agent. We have demonstrated that concomitant Se-enriched lactobacillus administration to mice and rats subjected to CCl4-induced liver injury results in a reliable hepatoprotection against hepatic damage, as well as an effective action on enhancing peripheral blood lymphocyte proliferation activity and RBC immune function[9,10]. Therefore, the aim of this study was to further evaluate the beneficial action of Se-enriched lactobacillus on reversing a well-established liver injury and the possible mechanisms by analyzing the activities of several liver tissue injury-associated enzymes, phagocytic function activities in macrophages, plasma TNF-α level and change of hepatocyte intracellular free Ca2+ [Ca2+]i homeostasis.
MATERIALS AND METHODS
Preparation of a complete diet with Se-enriched lactobacillus
Lactobacillus was grown to a density of 5.88107/mL in a medium containing sodium selenite to obtain Se-enriched lactobacillus. The Se-enriched lactobacillus was then processed into an organoselenium agent with 11.45 mg of selenium/g, then mixed into the common complete feed to produce the palletized complete diet with 2 and 4 mg of organoselenium/kg feed according to compound feed processing method in a feed processing plant.
Animals
Seventy-two ICR mice, equal number of males and females, weighing 27.740.37 g, were obtained from Experimental Animal Center of Nanjing Medical University, China. The mice were housed at room temperature (20-25 °C) in a 12-h light and dark cycle. Free access to water and food was allowed during the experimental period. All mice were randomly divided into four groups: normal control group (n = 18), model control group (n = 18), low Se-enriched lactobacillus treatment group (L-Se group, n = 18), and high Se-enriched lactobacillus treatment group (H-Se group, n = 18). During a 3-wk experimental period, the common complete diet was orally provided daily for normal group and model group, and the mice in L-Se and H-Se groups were given a diet with 2 and 4 mg of organoselenium from Se-enriched lactobacillus/kg feed, respectively. In the 2nd wk of experiment, hepatic injury was induced in mice by carbon tetrachloride (CCl4). The mice in model group, L-Se and H-Se groups were given abdominal cavity injection of olive oil solution containing 500 mL/L CCl4 (0.07 mL/100 g body mass) every other day for over 2 wk. The mice in normal group received abdominal injection of olive oil at the same dose and frequency.
Phagocytic function detection of macrophage
The macrophage activities were observed in six mice of each group in the first 2 wk post injection with CCl4. The mice received abdominal cavity injection of 1 mL saline suspension containing 5% chicken’s erythrocytes, followed by 1 mL saline after 1 h. Abdominal cavity fluid was collected from the killed mice and incubated for 30 min at 37 °C, then centrifuged at 1 000 r/min for 10 min to obtain the remaining deposit for smearing and staining with Wright-Giemsa stain. The phagocytic rate and phagocytic index of macrophages were counted under a light microscope.
Collection of specimens
In the first 2 wk post injection with CCl4, six animals were chosen from each group and anesthetized with ether before they were killed to collect blood samples by enucleating eyeballs, which were put into test tubes containing anticoagulant heparin sodium (1:500). Plasma was obtained by centrifugation at 3 000 r/min for 20 min and stored for analysis. Then, the animals were killed and the liver was quickly washed in situ with ice-cold isotonic saline, followed by stripping the capsules. Parenchyma (0.5 g) was sheared into broken bits and put into test tubes with 4 mL physiological saline for tissue homogenates. Tubes were centrifuged at 4 000 r/min for 30 min at 4 °C to separate upper fluid, which was stored at -20 °C for analysis.
Laser scanning confocal microscopic analysis of intracellular free Ca2+ in hepatocytes
Hepatocyte isolation In the 1st wk post injection with CCl4, six mice from each group were exsanguinated and liver was minced with dissecting scissors into approximately 1 mm3 pieces. The pieces of liver were placed into 3 mL D-Hank’s solution containing 0.25% trypsin (1:250, Amresco Co. USA) for 1 h digestion at 37 °C, and then added into small 1640 medium (pH 7.2) containing 10% neonatal bovine serum (Sijiqing Co., Hangzhou, China). Streptomycin and penicillin (1105 U/L for each) were used to terminate the digestion. The released cells were filtered through sterilized nylon membranes and washed in D-Hank’s solution via two centrifugations at 1 000 r/min for 5 min after erythrocytes were dissolved with 0.85% amchlor solution, and then resuspended in 1640 medium containing 10% neonatal bovine serum.
Fluo-3/AM loading of hepatocytes Twenty microliters of suspension samples containing freshly isolated hepatocytes from each group was placed in 13 mm culture dishes. The glass bottom of the culture dishes were covered with a layer of 0.1 mg/mL ConA (Fluka Co., USA) and dried at 37 °C in order to allow cells to adhere. The cells were loaded with 15 mL of 0.1 mg/mL Fluo-3/AM (Calbiochem Co., USA) for 30 min at 37 °C, and washed thrice with Hank抯 solution (pH 7.2) to remove the extracellular Fluo-3/AM.
The fluorescence intensity of [Ca2+]i in hepatocytes was determined by ACAS Ultima-312 laser scanning confocal microscopy. Total images were scanned in each experiment and the data were stored in disks for analysis.
Parameters of plasma and liver homogenates in mice with hepatic inju ry
The activities of glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), alanine aminotransferase (ALT) and aspartate aminotransferase (AST) as well as malondialdehyde (MDA) content in liver tissue homogenates were assayed by spectrophotometry using commercial kits (Nanjing Jiancheng Biotechnology Institute, China). Plasma TNF-α level was measured by 125I-based radioimmunoassay using commercial reagents (Radio-Immunity Institute of General Hospital of Chinese Liberation Army). In general, the procedures indicated by the kits were performed strictly according to themanufacturer’s protocol.
Statistical analysis
Results were expressed as mean±SE. Statistical analysis was performed by Student’s t-test (STATISTICA, Statsoft Inc., Tulsa, USA) on a conventional personal computer. P < 0.05 was considered statistically significant.
RESULTS
Effect of Se-enriched lactobacillus on AST and ALT activities of liver homogenates in mice with CCl4-induced hepatic injury
As shown in Table 1, during the entire experimental period, CCl4 significantly increased AST activity in liver homogenates compared to normal control (P < 0.05). This effect was completely prevented by both low and high doses of Se-enriched lactobacillus (P < 0.05). Upon exposure to CCl4, ALT activity in liver homogenates was not affected in the 1st wk of treatment, but significantly enhanced in the 2nd wk (P < 0.05). This effect was also blunted by a high dose of Se-enriched lactobacillus (P < 0.05).
Table 1.
Changes of AST and ALT activities in mice with hepatic injury after Se-enriched lactobacillus treatment (mean±SE)
| Index | Group |
Post-injection with CCl4 |
|
| 1 wk | 2 wk | ||
| AST (U/mg tissue) | Normal | 20.43 ± 0.54a | 17.68 ± 1.21a |
| Model | 28.05 ± 1.03c | 26.81 ± 0.46c | |
| L-Se | 23.60 ± 0.93a | 17.86 ± 1.75a | |
| H-Se | 18.90 ± 0.94a | 18.90 ± 2.26a | |
| ALT (U/mg tissue) | Normal | 31.64 ± 0.23 | 42.93 ± 0.42a |
| Model | 31.15 ± 0.32 | 50.61 ± 1.05c | |
| L-Se | 30.71 ± 0.39 | 49.47 ± 0.26c | |
| H-Se | 28.99 ± 0.13ac | 43.64 ± 1.61a | |
P<0.05 vs model group;
P<0.05 vs normal group.
Effect of Se-enriched lactobacillus on GSH-Px and SOD activities of liver homogenates in mice with CCl4-induced hepatic injury
As demonstrated in Table 2, GSH-Px activity in the liver homogenates slightly decreased in the 1st wk, and significantly decreased in the 2nd wk after CCl4 treatment. This effect was blunted by a high dose of Se-enriched lactobacillus (P < 0.05). Similarly, during the entire period of treatment, SOD activity dramatically decreased after CCl4 treatment (P < 0.05), which was rescued by both low and high doses of Se-enriched lactobacillus.
Table 2.
Changes of GSH-Px and SOD activities in mice with hepatic injury after Se-enriched lactobacillus treatment (mean±E)
| Index | Group |
Post-injection with CCl4 |
|
| 1 wk | 2 wk | ||
| GSH-Px (U/mg tissue) | Normal | 6.43 ± 0.67 | 6.40 ± 0.38a |
| Model | 6.24 ± 0.36 | 5.21 ± 0.24c | |
| L-Se | 7.68 ± 0.33 | 5.67 ± 0.25 | |
| H-Se | 8.10 ± 0.21a | 6.45 ± 0.31a | |
| SOD (U/mg tissue) | Normal | 60.57 ± 2.25a | 61.34 ± 0.83a |
| Model | 47.88 ± 2.15c | 48.95 ± 1.19c | |
| L-Se | 54.43 ± 2.43 | 52.70 ± 2.14c | |
| H-Se | 52.61 ± 3.46 | 51.50 ± 2.20c | |
P<0.05 vs model group;
P<0.05 vs normal group.
Effect of Se-enriched lactobacillus on MDA content in liver homogenates of mice with CCl4-induced hepatic injury
Figure 1 shows that CCl4 significantly increased MDA content in liver homogenates compared to normal control during the entire experimental period (P < 0.05). However, administr-ation of Se-enriched lactobacillus prevented MDA elevation.
Figure 1.
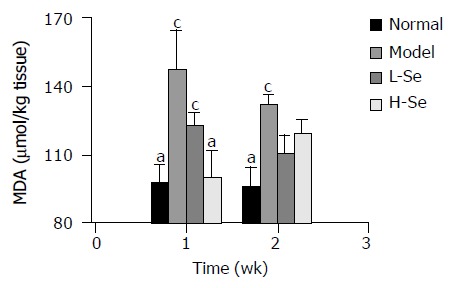
Change of MDA content in liver homogenates of mice with hepatic injury after Se-enriched lactobacillus treatment. aP < 0.05 vs model group; cP < 0.05 vs normal group.
Effect of Se-enriched lactobacillus on macrophage activities in mice with CCl4-induced liver injury
During the entire experimental period, the phagocytic rate of macrophages significantly decreased after CCl4 treatment compared to normal control (P < 0.05), and was dramatically rescued by both low and high doses of Se-enriched lactobacillus, showing an obviously higher phagocytic rate than that in model group (P < 0.05). The phagocytic index of macrophages slightly decreased in the 1st wk, and significantly decreased in the 2nd wk after CCl4 treatment. This effect was protected by high dose of Se-enriched lactobacillus (P < 0.05, Table 3).
Table 3.
Changes of phagocytic function in mice with hepatic injury after Se-enriched lactobacillus treatment (mean±SE)
| Index | Group |
Post-injection with CCl4 |
|
| 1 wk | 2 wk | ||
| Phagocytic rate (%) | Normal | 17.25 ± 0.81a | 15.25 ± 1.18a |
| Model | 11.67 ± 1.00c | 10.64 ± 0.56c | |
| L-Se | 14.92 ± 0.33ac | 12.67 ± 0.49a | |
| H-Se | 14.92 ± 0.33ac | 12.67 ± 0.49a | |
| Phagocytic index (RBC/macrophage) | Normal | 1.75 ± 0.11 | 1.69 ± 0.08a |
| Model | 1.46 ± 0.08 | 1.17 ± 0.06c | |
| L-Se | 1.52 ± 0.12 | 1.36 ± 0.07 | |
| H-Se | 1.72 ± 0.04a | 1.55 ± 0.09a | |
P<0.05 vs model group;
P<0.05 vs normal group.
Effect of Se-enriched lactobacillus on plasma TNF-α in mice with CCl4-induced liver injury
During the entire experimental period, CCl4 significantly elevated plasma TNF-α level compared to normal control (P < 0.05). This effect was completely prevented by both low and high doses of Se-enriched lactobacillus, showing an obviously lower value than that in model group (P < 0.05) and no significant difference as compared to normal group (Figure 2).
Figure 2.
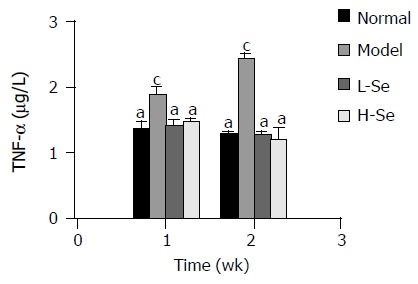
Change of plasma TNF-α level in mice with hepatic injury after Se-enriched lactobacillus treatment. aP < 0.05 vsmodel group; cP < 0.05 vs normal group.
Effect of Se-enriched lactobacillus on hepatocytes [Ca2+]i in mice with CCl4-induced liver injury
Figure 3A shows changes of hepatocyte [Ca2+]i fluorescent visualizations in mice with hepatic injury after Se-enriched lactobacillus treatment. In model group, the hepatocyte [Ca2+]i content significantly raised and was 5.5-fold higher than that in normal group (P < 0.05), but hepatocytes [Ca2+]i in mice with liver injury after Se-enriched lactobacillus treatment was significantly downregulated (P < 0.05) and close to that in normal control (Figure 3B).
Figure 3.
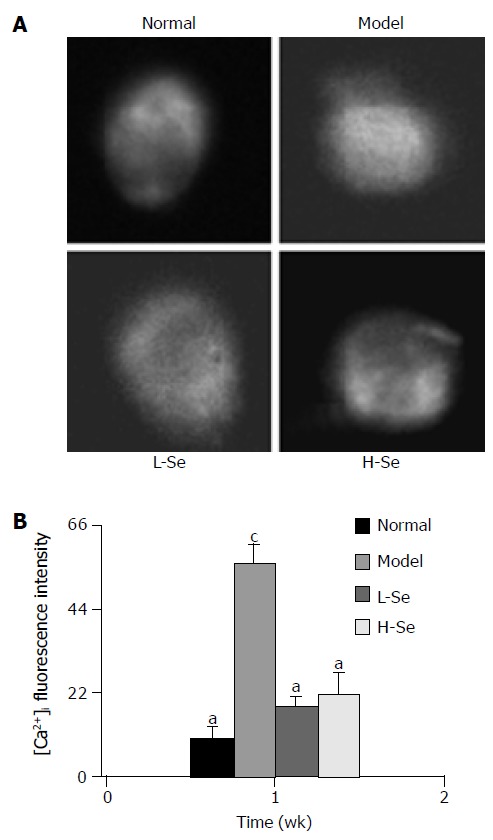
[Ca2+]i fluorescence visualizations (A) and intensities (B) in mice with hepatic injury after Se-enriched lactobacillus treatment. aP < 0.05 vs model group; cP < 0.05 vs normal group.
DISCUSSION
Liver fibrosis is a common pathological process of hepatic disease, leading to the development of irreversible cirrhosis in patients. In recent years, the mechanism of hepatic fibrosis has been partly disclosed[11]. If treated properly at fibrosis stage, cirrhosis could be prevented[3]. Several drugs, including cytokines, antioxidants, chemical drugs, soluble type II receptor for TGF-b1, and antibodies directed against TGF-b1 have been used to block liver fibrosis, but their effects are not as good as expected[12]. The present study has demonstrated that Se-enriched lactobacillus is effective in treating mice hepatic injury and fibrosis based on both lipid peroxidation reaction and functional analysis. The underlying protective mechanism may involve enhanced immunity, and downregulation of TNF-α release and equilibration of hepatocyte [Ca2+]i homeostasis.
There are various kinds of liver injuries all over the world, causing great affliction to patients. Searches for effective ways to inhibit fibrogenesis and to prevent the development of cirrhosis are of great significance. Selenium can prevent the malignant transformation of normal cells and the activation of oncogenes with anticarcinogenic effects within a physiological dosage range[4,5]. Selenium is closely related to the inhibition of hepatic fibrosis[7,8]. Se-enriched lactobacillus is an organoselenium agent. The present study showed that ALT, AST and MDA in mice after Se-enriched lactobacillus treatment dropped significantly when compared to those in model group, indicating that Se-enriched lactobacillus reduces the violent lipid peroxidation reaction induced by CCl4 and prevents hepatic injury.
GSH-Px, a selenium-containing enzyme, is a primary selenium-dependent, glutathione-utilizing peroxidase in the liver[13-15]. The decreased GSH-Px activity has been implicated in animals fed with a selenium-deficient diet[16,17]. Conversely, selenium supplement can also impact GSH-Px activity directly[18,19]. In this study, CCl4 caused an obvious decrease of GSH-Px activity in liver homogenates, but the enzyme activity was higher after Se-enriched lactobacillus treatment, suggesting that Se-enriched lactobacillus can ameliorate GSH-Px activity or enhance its activity by participating in GSH-Px biosynthesis in mice with CCl4-induced liver injury. SOD is one of the potent antioxidant enzymes in cells and catalyzes the conversion of superoxide ions into oxygen and hydrogen peroxide. In our study, SOD activity was dramatically decreased by CCl4, but the effect was rescued by both low and high doses of Se-enriched lactobacillus. GSH-Px and SOD are the important antioxidant enzymes of antioxidant defense systems for stabilizing oxidative reactions. Decreased activity and content of these enzymes cause the descent of antioxidant ability in the body[20]. Therefore, Se-enriched lactobacillus may prevent experimental liver injury by modulating or enhancing GSH-Px and SOD activities.
Macrophages are important immune cells of the immune system and the first line of defense to infection and inflammation in the body, and play a very important role in regulating the body immune status. Many experiments have demonstrated that the macrophage phagocytosis in mice with CCl4-induced liver injury can be reduced[16,21,22]. In our study, Se-enriched lactobacillus was found to be able to enhance the immunity of the body by increasing the phagocytic rate and phagocytic index in macrophages in L-Se group and H-Se group. This fact may be relevant to the protection of its biomembrane against oxidative destruction. Mayanski et al[22] showed that structural and functional integrity of the cell membrane system including macrophages in mice with CCl4-induced liver injury is damaged, thus depressing the activity of macrophages.
To understand the mechanism, we evaluated the effect of Se-enriched lactobacillus treatment on blood TNF-α concentration in mice with liver injury. There is evidence that TNF-α is a primary and key endogenous mediator inducing acute inflammatory conditions[23-25]. When an inflammatory reaction occurs, TNF-α is expressed by both infiltrating inflammatory cells such as macrophages in the blood and hepatocytes in liver injuries. Excessive TNF-α can indirectly induce liver damage by further enhancing the release of mass inflammatory mediators including free radicals, prostaglandin, leukotriene, thromboxane, interleukin-1, and TNF-α from macrophages, and also promote liver injury by increasing nitric oxide production from liver cells[26,27]. The present study showed that plasma TNF-α level due to CCl4 increased obviously, but the effect was significantly blunted by Se-enriched lactobacillus, indicating that Se-enriched lactobacillus downregulates excessive release of TNF-α.
It was reported that changes of intracellular cation homeostasis are closely related to the mechanism of hepatic cell injury[28,29]. Elevation of [Ca2+]i concentration is associated with the development of cell damage[30,31]. In the present study, CCl4 caused the increase of hepatocyte [Ca2+]i, suggesting that the dramatic elevation of hepatocyte [Ca2+]i due to CCl4 is closely related to CCl4-induced hepatocellular injury. However, the hepatocyte [Ca2+]i level in mice with liver injury after Se-enriched lactobacillus treatment was significantly lower than that in model group, indicating that Se-enriched lactobacillus may prevent disturbance of [Ca2+]i homeostasis. Other investigations showed that hepatocyte [Ca2+]i elevation is caused by the decreased activities of Na+/K+- and Ca2+-ATPases in hepatocyte membranes[32,33]. In addition, it is possible that CCl4 may release Ca2+ by inhibiting the endoplasmic reticulum Ca2+ pump so that a passive leak of Ca2+ from the endoplasmic reticulum results in the increase of [Ca2+]i and induces cell death[30,34,35]. Thus, Se-enriched lactobacillus may play an important role in preventing liver injury. The exact mechanisms remain to be explored.
In conclusion, Se-enriched lactobacillus can significantly reduce CCl4-induced liver injury in mice by enhancing macrophage function activity to keep normal and beneficial effects, elevating antioxidant-enzyme activities and reducing lipid-peroxidation reaction, inhibiting excessive release of TNF-α and preventing dramatic elevation of hepatocyte [Ca2+]i concentration.
ACKNOWLEDGMENTS
We thank Dr. Chao-Ying Zhang at the College of Life Sciences, Nanjing Normal University for assistance with laser scanning confocal microscopic analysis of [Ca2+]i in hepatocytes.
Footnotes
Science Editor Wang XL and Guo SY Language Editor Elsevier HK
Supported by the Special Programs of State Science and Technology Ministry of China During the 10th 5-Year Plan Period, No. 2002BA518A12, and Open Foundation from Key Laboratory of Resource Biotechnology of Jiangsu Province, China, No. KJS00033
References
- 1.Shen L, Fan JG, Shao Y, Zeng MD, Wang JR, Luo GH, Li JQ, Chen SY. Prevalence of nonalcoholic fatty liver among administrative officers in Shanghai: an epidemiological survey. World J Gastroenterol. 2003;9:1106–1110. doi: 10.3748/wjg.v9.i5.1106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wang FS, Wu ZZ. Current situation in studies of gene therapy for liver cirrhosis and liver fibrosis. Shijie Huaren Xiaohua Zazhi. 2000;8:371–373. [Google Scholar]
- 3.Riley TR, Bhatti AM. Preventive strategies in chronic liver disease: part II. Cirrhosis. Am Fam Physician. 2001;64:1735–1740. [PubMed] [Google Scholar]
- 4.Schrauzer GN. Anticarcinogenic effects of selenium. Cell Mol Life Sci. 2000;57:1864–1873. doi: 10.1007/PL00000668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schrauzer GN. Nutritional selenium supplements: product types, quality, and safety. J Am Coll Nutr. 2001;20:1–4. doi: 10.1080/07315724.2001.10719007. [DOI] [PubMed] [Google Scholar]
- 6.Shamberger RJ, Frost DV. Possible protective effect of selenium against human cancer. Can Med Assoc J. 1969;100:682. [PMC free article] [PubMed] [Google Scholar]
- 7.Zhang M, Song G, Minuk GY. Effects of hepatic stimulator substance, herbal medicine, selenium/vitamin E, and ciprofloxacin on cirrhosis in the rat. Gastroenterology. 1996;110:1150–1155. doi: 10.1053/gast.1996.v110.pm8613004. [DOI] [PubMed] [Google Scholar]
- 8.Ianăş O, Olinescu R, Bădescu I, Simionescu L, Popovici D. The influence of "selenium organicum" upon the hepatic function of carbon tetrachloride poisoned rats. Rom J Intern Med. 1995;33:113–120. [PubMed] [Google Scholar]
- 9.Sun YY, Chen L, Jiang YZ. Protection effect of lactobacillus with selenium enrichment on growth and lymphocyte transformation of rats with liver injuries. Zhongguo YingYong ShengLiXue ZaZhi. 2003;19:366–37, 397. [PubMed] [Google Scholar]
- 10.Zhou J, Chen L, Chen LJ, Pan DD, Jiang YZ. Effects of lactobacillus rich in selenium on RBC lipid peroxidation and immune function in mice with CCl4–injected liver injury. Yingyang Xuebao. 2003;25:405–408. [Google Scholar]
- 11.Zhu YH, Hu DR, Nie QH, Liu GD, Tan ZX. Study on activation and c-fos, c-jun expression of in vitro cultured human hepatic stellate cells. Shijie Huaren Xiaohua Zazhi. 2000;8:299–302. [Google Scholar]
- 12.Bissell DM, Roulot D, George J. Transforming growth factor beta and the liver. Hepatology. 2001;34:859–867. doi: 10.1053/jhep.2001.28457. [DOI] [PubMed] [Google Scholar]
- 13.Ho YS, Magnenat JL, Bronson RT, Cao J, Gargano M, Sugawara M, Funk CD. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J Biol Chem. 1997;272:16644–16651. doi: 10.1074/jbc.272.26.16644. [DOI] [PubMed] [Google Scholar]
- 14.Esposito LA, Kokoszka JE, Waymire KG, Cottrell B, MacGregor GR, Wallace DC. Mitochondrial oxidative stress in mice lacking the glutathione peroxidase-1 gene. Free Radic Biol Med. 2000;28:754–766. doi: 10.1016/s0891-5849(00)00161-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Esposito LA, Melov S, Panov A, Cottrell BA, Wallace DC. Mitochondrial disease in mouse results in increased oxidative stress. Proc Natl Acad Sci USA. 1999;96:4820–4825. doi: 10.1073/pnas.96.9.4820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hamazaki K, Sato S, Yunoki M, Noda T, Moreira LF, Mimura H, Orita K. Kupffer cell function in chronic liver injury and after partial hepatectomy. Res Exp Med (Berl) 1994;194:237–246. doi: 10.1007/BF02576385. [DOI] [PubMed] [Google Scholar]
- 17.Noguchi T, Cantor AH, Scott ML. Mode of action of selenium and vitamin E in prevention of exudative diathesis in chicks. J Nutr. 1973;103:1502–1511. doi: 10.1093/jn/103.10.1502. [DOI] [PubMed] [Google Scholar]
- 18.Kvícala J. [Selenium and the organism] Cas Lek Cesk. 1999;138:99–106. [PubMed] [Google Scholar]
- 19.Tolando R, Jovanovic A, Brigelius-Flohé R, Ursini F, Maiorino M. Reactive oxygen species and proinflammatory cytokine signaling in endothelial cells: effect of selenium supplementation. Free Radic Biol Med. 2000;28:979–986. doi: 10.1016/s0891-5849(00)00183-0. [DOI] [PubMed] [Google Scholar]
- 20.Menegola E, Broccia ML, Prati M, Ricolfi R, Giavini E. Glutathione status in diabetes-induced embryopathies. Biol Neonate. 1996;69:293–297. doi: 10.1159/000244323. [DOI] [PubMed] [Google Scholar]
- 21.Noda T, Mimura H, Orita K. Assessment of Kupffer cell function in rats with chronic liver injury caused by CCl4. Hepatogastroenterology. 1990;37:319–323. [PubMed] [Google Scholar]
- 22.Mayanski DN, Schwartz YS, Kutina SN, Zubakhin AA, Mayanskaya NN, Tsyrendorjiev DD. Mononuclear phagocyte system responsiveness in CCl4-induced liver cirrhosis. Int J Exp Pathol. 1993;74:229–236. [PMC free article] [PubMed] [Google Scholar]
- 23.Chen L, Zhou J, Gao W, Jiang YZ. Action of NO and TNF-alpha release of rats with cadmium loading in malfunctiion of multiple system organ. Sheng Li Xue Bao. 2003;55:535–540. [PubMed] [Google Scholar]
- 24.Kimura K, Bowen C, Spiegel S, Gelmann EP. Tumor necrosis factor-alpha sensitizes prostate cancer cells to gamma-irradiation-induced apoptosis. Cancer Res. 1999;59:1606–1614. [PubMed] [Google Scholar]
- 25.Karck U, Peters T, Decker K. The release of tumor necrosis factor from endotoxin-stimulated rat Kupffer cells is regulated by prostaglandin E2 and dexamethasone. J Hepatol. 1988;7:352–361. doi: 10.1016/s0168-8278(88)80008-4. [DOI] [PubMed] [Google Scholar]
- 26.Al-Shabanah OA, Alam K, Nagi MN, Al-Rikabi AC, Al-Bekairi AM. Protective effect of aminoguanidine, a nitric oxide synthase inhibitor, against carbon tetrachloride induced hepatotoxicity in mice. Life Sci. 2000;66:265–270. doi: 10.1016/s0024-3205(99)00589-5. [DOI] [PubMed] [Google Scholar]
- 27.Hoebe KH, Witkamp RF, Fink-Gremmels J, Van Miert AS, Monshouwer M. Direct cell-to-cell contact between Kupffer cells and hepatocytes augments endotoxin-induced hepatic injury. Am J Physiol Gastrointest Liver Physiol. 2001;280:G720–G728. doi: 10.1152/ajpgi.2001.280.4.G720. [DOI] [PubMed] [Google Scholar]
- 28.Gasbarrini A, Borle AB, Farghali H, Bender C, Francavilla A, Van Thiel D. Effect of anoxia on intracellular ATP, Na+i, Ca2+i, Mg2+i, and cytotoxicity in rat hepatocytes. J Biol Chem. 1992;267:6654–6663. [PubMed] [Google Scholar]
- 29.Carini R, Bellomo G, Benedetti A, Fulceri R, Gamberucci A, Parola M, Dianzani MU, Albano E. Alteration of Na+ homeostasis as a critical step in the development of irreversible hepatocyte injury after adenosine triphosphate depletion. Hepatology. 1995;21:1089–1098. [PubMed] [Google Scholar]
- 30.Ueda T, Takeyama Y, Hori Y, Takase K, Goshima M, Kuroda Y. Pancreatitis-associated ascitic fluid increases intracellular Ca(2+) concentration on hepatocytes. J Surg Res. 2000;93:171–176. doi: 10.1006/jsre.2000.5959. [DOI] [PubMed] [Google Scholar]
- 31.Orrenius S, McCabe MJ, Nicotera P. Ca(2+)-dependent mechanisms of cytotoxicity and programmed cell death. Toxicol Lett. 1992;64-65 Spec No:357–364. doi: 10.1016/0378-4274(92)90208-2. [DOI] [PubMed] [Google Scholar]
- 32.Muriel P, Favari L, Soto C. Erythrocyte alterations correlate with CCl4 and biliary obstruction-induced liver damage in the rat. Life Sci. 1993;52:647–655. doi: 10.1016/0024-3205(93)90456-d. [DOI] [PubMed] [Google Scholar]
- 33.Srivastava SP, Chen NQ, Holtzman JL. The in vitro NADPH-dependent inhibition by CCl4 of the ATP-dependent calcium uptake of hepatic microsomes from male rats. Studies on the mechanism of the inactivation of the hepatic microsomal calcium pump by the CCl3.radical. J Biol Chem. 1990;265:8392–8399. [PubMed] [Google Scholar]
- 34.Long RM, Moore L. Cytosolic calcium after carbon tetrachloride, 1,1-dichloroethylene, and phenylephrine exposure. Studies in rat hepatocytes with phosphorylase a and quin2. Biochem Pharmacol. 1987;36:1215–1221. doi: 10.1016/0006-2952(87)90073-6. [DOI] [PubMed] [Google Scholar]
- 35.Cheng JS, Jan CR. Effect of nordihydroguaiaretic acid on intracellular Ca2 concentrations in hepatocytes. Toxicol In Vitro. 2002;16:485–490. doi: 10.1016/S0887-2333(02)00051-6. [DOI] [PubMed] [Google Scholar]