

Abstract
The present trend of increasing paternal age is accompanied by concerns for the development of complex multigene diseases (e.g., autism and schizophrenia) in progeny. Recent studies have established strong correlations between male age, increased oxidative stress, decreased sperm quality, and structural aberrations of chromatin and DNA in spermatozoa. We tested the hypothesis that increasing age would result in altered gene expression relating to oxidative stress and DNA damage/repair in germ cells. To test this hypothesis, pachytene spermatocytes and round spermatids were isolated from Brown Norway (BN) rats at 4 (young) and 18 (aged) mo of age. Microarray analysis was used to compare gene expression between the groups. The probe sets with significantly altered expression were linked to DNA damage/repair and oxidative stress in pachytene spermatocytes but not in round spermatids. Further analysis of pachytene spermatocytes demonstrated that genes involved in the base excision repair (BER) and nucleotide excision repair (NER) pathways were specifically altered. Quantitative RT-PCR confirmed that NER genes were upregulated (>1.5-fold), whereas BER genes were downregulated (>1.5-fold). At the protein level the members of the BER pathway were also altered by up to 2.3-fold; levels of NER proteins remained unchanged. Furthermore, there was an increase in 8-oxo-2′-deoxyguanosine (8-oxodG) immunoreactivity in testes from aged males and in the number of spermatozoa positive for 8-oxodG. In conclusion, aging is associated with differential regulation of DNA repair pathways with a decrease in the BER pathway leading to deficient repair of 8-oxo-dG lesions in germ cells and spermatozoa.
Keywords: aging, gene expression, oxidative stress, spermatocyte
Testicular aging coincides with a decrease in the efficiency of the base excision repair pathway and increased DNA damage in germ cells.
INTRODUCTION
There is growing evidence that during aging, the functions of the male reproductive system decline and that, with increasing age in men, there is a decline in the production of functional spermatozoa that produce healthy offspring. It is now well documented that the age of the male partner is strongly correlated with a decrease in spermatozoal motility and normal morphology [1–4] as well as with decreased pregnancy rates and an increased time to pregnancy [5–7]. Paternal age has also been linked to chromosome damage and to genetic problems in children sired by older fathers. During spermatogenesis, germ cell DNA is replicated and exchanged by homologous recombination, and at this time the cells are vulnerable to the introduction of a range of errors. However, DNA damage can be introduced at all stages of spermatogenesis, and if these cells are not eliminated, then any errors incurred in their DNA could be passed on to the offspring.
DNA repair is required under normal circumstances for meiotic recombination and for the correction of DNA damage in developing germ cells [8]. Germ cell DNA is also continuously under attack from endogenous and exogenous factors that can induce a wide range of DNA abnormalities/lesions. These lesions can be formed under normal processes such as transcription, recombination, and replication; however, they can also be induced by irradiation, exposure to chemicals, and high levels of reactive oxygen species (ROS; reviewed in Aitken and De Iuliis [9]). Germ cells have the ability to produce high levels of ROS [10] that can lead to the formation of DNA lesions. It has been shown in many tissues that there is an increase in ROS with aging [11]. Although there is a complex antioxidant defense system present in the testis [12], the level of antioxidants in the aging testis declines [13, 14], thus causing an imbalance between ROS generation and the antioxidant defense system. This can have consequences on DNA integrity, and the fertilization capacity of spermatozoa can become compromised [15, 16]. We and others have shown that there is an increase in ROS in spermatozoa with age [13, 14]. The base excision repair (BER) pathway is the main operator in the removal of oxidative DNA damage in many tissues, including the testis; this mechanism allows for the removal of aberrant bases in DNA induced by hydrolysis, ROS, or other metabolites. One of the most common lesions caused by oxidative stress, 8-oxo-2′-deoxyguanosine (8-oxo-dG) [17], is strongly mutagenic and has the ability to block transcription.
Although a number of studies have investigated DNA damage in spermatozoa from the aging male, there are much fewer studies focused on the DNA damage that occurs during the process of spermatogenesis (e.g., in earlier germ cells, such as the spermatocytes and round spermatids). We hypothesize that germ cells from older males will have altered gene expression in comparison to younger males and that the genes that are altered will be associated primarily with oxidative stress and DNA damage recognition and repair. To test this hypothesis, we investigated the consequences of aging on gene expression in pachytene spermatocytes and round spermatids from the BN rat. BN rats were used, as they are a well-established model for the study of aging in the male reproductive tract; while remaining otherwise healthy, they exhibit the same reproductive aging phenotypes as found in man, including decreased testosterone without decreased gonadotropins [18, 19].
MATERIALS AND METHODS
Animals
Male Brown Norway rats of 4 and 18 mo of age (six animals per group) were maintained under standard conditions as described in A Guide to the Care and Use of Experimental Animals prepared by the Canadian Council on Animal Care. All animals were kept on a 12L:12D cycle with free access to food and water (McGill Animal Resources Centre protocol 4687). Rats in the “young” group (4 mo old) were purchased directly from Harlan, while rats in the “aged” group (18 mo old) were purchased from Harlan via the National Institute on Aging. Aged rats were chosen to be at 18 mo, as this is the time point just prior to the onset of germ cell loss and testicular atrophy.
Germ Cell Separation
Each rat was checked for the presence of regressed testes (<1.4 g); only those rats that did not have regressed testes were used in this study. Spermatogenic cells were obtained through cell separation using the STA-PUT velocity sedimentation technique as described by Bellve et al. [20] and modified by Aguilar-Mahecha et al. [21]. Briefly, the tunica albuginea was removed along with any large blood vessels; the parenchyma was subjected to enzymatic digestion at 34°C first with 0.5 mg/ml collagenase (Sigma Aldrich Canada) for 16 min followed by sedimentation and washing then with 0.5 mg/ml trypsin (Type I, T8003; Sigma) and DNase I (Type I, DN-25; Sigma) for 16 min. After dissociation, cells were filtered through a nylon mesh (70 μm) and washed with RPMI (RPMI medium 160; Invitrogen) containing 0.5% BSA. Cells were centrifuged and filtered, and 5.6 × 108 cells in 25 ml of 0.5% BSA (bovine serum albumin)/RPMI were loaded into the velocity sedimentation apparatus (STA-PUT; Proscience) and separated on a 2%–4% BSA gradient in RPMI for separation by sedimentation at unit gravity. Fractions of pachytene spermatocytes and round spermatids (steps 1–9) were identified by phase contrast microscopy. Fractions with an average purity of 85% (not less than 83%) were pooled, aliquoted at 5 × 106/ml, pelleted, and stored at −80°C until use.
RNA Extraction and Microarray
Total RNA was extracted from the six pachytene spermatocyte and the six round spermatid fractions (∼5 × 106 cells) using the RNeasy Mini Plus kit with on-column DNase digestion (Qiagen). RNA concentration was determined using the Nanodrop 2000 (Nanodrop Technologies) and quality assessed using a Bioanalyzer (Agilent Technologies). Gene expression analysis was done using Affymetrix Rat 230 2.0 microarrays (n = 6 per group) in collaboration with Genome Quebec. Three micrograms of RNA were reverse transcribed, and the cRNA was hybridized on the microarray according to the manufacturer's instructions. All chips had a background signal of less than 75 and Gapdh 3′/5′ hybridization rate from 0.91 to 1.6. The Robust Multiarray Algorithm (RMA) [22] was applied, and only those genes with an expression value higher than 50 were considered as “expressed.” The raw data obtained were further normalized per gene to the median (Genespring v11.0; Agilent Technologies). All GC-RMA data were placed in GEO (accession no. GSE29963; NCBI). Statistical significance between the two groups was tested by the Student t-test using a P-value <0.05, and those probe sets that were significantly altered were further filtered using a minimum 1.5-fold difference. Cellular pathways were obtained using Pathway Studio 7.1 (Ariadne Genomics) with the ResNet-7.0 database.
Real-Time Quantitative RT-PCR
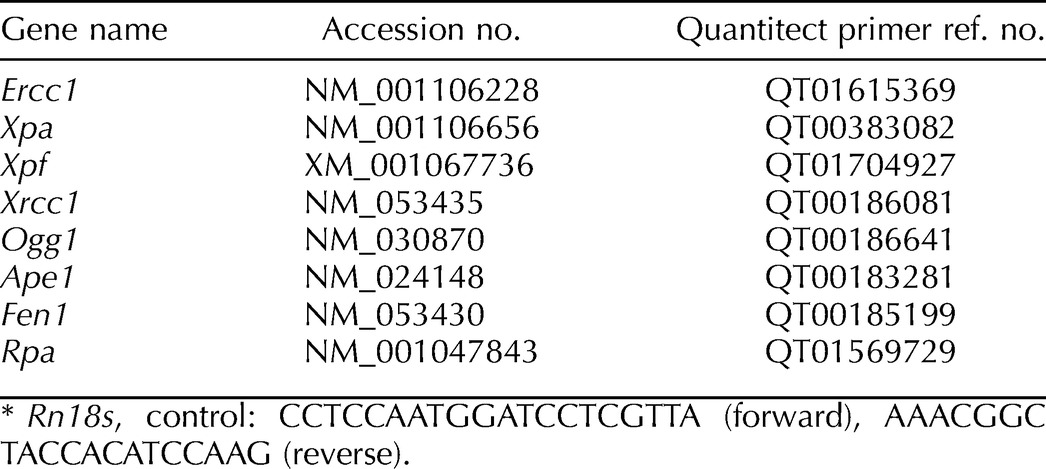
RNA was diluted to a working concentration of 2 ng/μl, and Quantitect One-Step SYBR Green quantitative RT-PCR (Qiagen) was done using the Roche Lightcycler (Roche Diagnostics) according to the manufacturer's instructions. PCR thermal cycling parameters were 95°C for 15 min (one cycle), 94°C for 15 sec, 55°C for 30 sec, and 72°C for 30 sec (50 cycles). Standard curves were generated using 0.1, 1, 10, and 100 ng/ml of RNA from control pachytene spermatocytes in each run for quantification. RT-PCR primers (Table 1) were either ready-made Quantitect Primer Assays (Qiagen) or designed using Primer3 software (http://frodo.wi.mit.edu) and provided by Alpha DNA. The expression levels of all genes of interest were corrected using an endogenous control (18S rRNA), and the fold difference in mRNA expression of the samples was determined. The results shown are the mean of at least six rats per group and done on two separate occasions, and each sample was analyzed in duplicate.
TABLE 1.
Primers used in quantitative RT-PCR.*
Immunohistochemistry
Rats were anaesthetized and the testes were perfused through the abdominal aorta first with 0.9% saline solution to clear the blood and then with Bouin fixative, as described previously [23]. Perfused testes were collected and further fixed in Bouin solution overnight followed by dehydration and embedding in paraffin. Testicular sections (5 μm) were cut and mounted on charged slides. Slides were dewaxed using xylene and rehydrated followed by blocking of endogenous peroxidase. Nonspecific binding sites were blocked using normal goat serum (NGS; Vector Laboratories) diluted 1:4 in BSA/TBS (Tris-buffered saline; 5%, w/v) for 30 min. Sections were incubated overnight at 4°C with the primary antibody specific for 8-oxodG (Oxis International) 1:100 in NGS/TBS/BSA; control sections were incubated with blocking serum alone. Bound antibodies were detected according to standard methods [24]. Images were captured using a Leica microscope DM LB2 (Leica) under a 63× lens fitted with an Infinity-3 video camera.
Western Blotting
Total protein was extracted from pachytene spermatocytes using RIPA lysis buffer, and protein concentration was determined using the BioRad protein assay kit according to manufacturer's instructions (BioRad). Samples (20 μg/lane) were resolved by SDS polyacrylamide (w/v) gradient (4%–12%) gels (Invitrogen) at 150 V for 1.5 h, then transferred onto PVDF membranes. Membranes were blocked with 5% nonfat milk in PBS containing 0.1% Tween-20. Proteins were detected using antibodies specific for ERCC1 (ab2356-500, 1:200), APE1 (ab194-50, 1:500), FEN1 (ab70815, 1:700), XRCC1 (ab1838-250, 1:500), OGG1 (ab204, 1:500; Abcam), XPA (sc853, 1:1000; Santa Cruz), RPA (2267s, 1:500, Cell Signaling), and XPF (NBP1-27881, 1:500, Novus Biologicals) all diluted in 3% nonfat milk in TBS-0.1% Tween followed by HRP-linked secondary antibodies and compared with the amount of β-actin (sc1616, 1:5000; Santa Cruz) to correct for loading.
Spermatozoa Collection
Cauda epididymides were excised, trimmed free of fat, and minced thoroughly in fresh PBS. The minced tissue was left for 5 min on ice with agitation to allow the spermatozoa to disperse and strained through a 70-μm nylon strainer (VWR International). The spermatozoa were washed twice in 0.45% saline to lyse contaminating red blood cells and finally washed twice in PBS before being centrifuged, and supernatant was removed. Spermatozoa were frozen at −80°C until further use. Frozen spermatozoa samples were resuspended in phosphate buffer (PB; 20 mM, pH 6.0 containing 1 mM EDTA) and sonicated twice on ice for 30 sec. Samples were centrifuged and resuspended in 50 mM Tris-HCl, pH 7.4, 1% SDS, and incubated at room temperature for 10 min. Samples were centrifuged for 1 min (10 000 × g), supernatant removed, and rewashed with 50 mM Tris-HCl, pH 7.4. Spermatozoa were resuspended in 25 mM Tris-HCl, 0.25 M (NH4)SO4, and 40 mM DTT and incubated at room temperature for 10 min. Thirty-microliter aliquots were placed on slides on ice and incubated for a further 40 min, washed three times in PBS (2 min), and fixed in 4% paraformaldehyde for 30 min at room temperature. Slides were washed in PBS and allowed to air dry before being stored at −20°C until further use. Positive controls were generated in the same way as above with the additional step of hydrogen peroxide (2 mM) treatment for 1 h at room temperature.
8-oxodG Immunofluorescence in Caudal Spermatozoa
Frozen slides were immersed in PBS for 5 min. Nonspecific binding sites were blocked using NGS (Vector Laboratories) diluted 1:4 in BSA/TBS (3%, w/v) for 30 min. Slides were incubated overnight at 4°C with primary antibody specific for 8-oxodG (Genox Corporation) 1:100 in NGS/TBS/BSA; control sections were incubated with blocking serum alone. Slides were washed in PBS and incubated with fluorescent anti-mouse secondary antibody (IgG) conjugated to Alexafluor 488 (Invitrogen) for 30 min at room temperature. Spermatozoa were counterstained with DAPI to stain nuclear DNA, washed, and mounted using Permafluor antifade mounting medium (Thermo Scientific). Staining was visualized on a Zeiss LSM 510 Axiovert confocal microscope using ×63/1.4 oil differential interface contrast objective. For quantitative analysis, at least 100 sperm per sample were counted as being positive or negative for 8-oxodG.
Statistical Analysis
Results expressed as means and standard errors of the mean were analyzed using Student t-test, using GraphPad Prism version 4 (Graph Pad Software Inc.).
RESULTS
Aging Alters Gene Expression in Pachytene Spermatocytes and Round Spermatids
Whole rat genome Affymetrix 230 2.0 microarrays were used to assess the impact of age on gene expression in pachytene spermatocytes and round spermatids. All RMA normalized data were deposited in GEO (accession no. GSE29963; NCBI). For the pachytene spermatocytes, of the 31 099 probe sets present on the array, 23 595 were considered expressed and 2837 had a fold change of at least 1.5 with respect to age; 1854 were known genes, and 983 had an unclassified name/function (Fig. 1A). For the round spermatids, 23 581 probe sets were considered expressed, and 817 had a fold change of at least 1.5 with respect to age; 565 were known genes, and almost half of these had an unclassified name/function (Fig. 1B). It is evident that aging results in altered expression of a high number of genes in both the pachytene spermatocytes and the round spermatids. In the pachytene spermatocytes, there were 3.5 times as many genes suppressed as those that were increased, suggesting an overall downregulation with age, whereas in the round spermatids, similar numbers were up- and downregulated (Fig. 1, A and B, respectively). The pachytene spermatocytes had 51 genes expressed only in the young and 98 expressed only in the aged (Fig. 1C). The round spermatids had 10 expressed only in the young and 261 expressed only in the aged (Fig. 1D). Those genes found to be uniquely expressed in only one age-group had a variety of different functions and are listed in the supplemental data (Supplemental Tables S1–S4, available online at www.biolreprod.org). Interestingly, a handful of genes were found to be commonly expressed in both aged cell types (spermatocytes and spermatids) but not in their young counterparts. These included a variety of different genes, such as Arg1 (arginase 1), Nup210 (Pom210, nuclear pore glycoprotein), Vsnl1 (visinin-like 1), and Runx1 (runt-related transcription factor).
FIG. 1.

Number of probe sets and known genes that are significantly altered (by at least 1.5-fold) in aged compared to young pachytene spermatocytes (A) and round spermatids (B). Venn diagrams of known genes expressed in both the young and old and those that are exclusively expressed in only one age-group in the pachytene spermatocytes (C) and the round spermatids (D). P < 0.05. n = 6.
Genes Involved in DNA Damage and Repair Are Specifically Affected in Aging Pachytene Spermatocytes
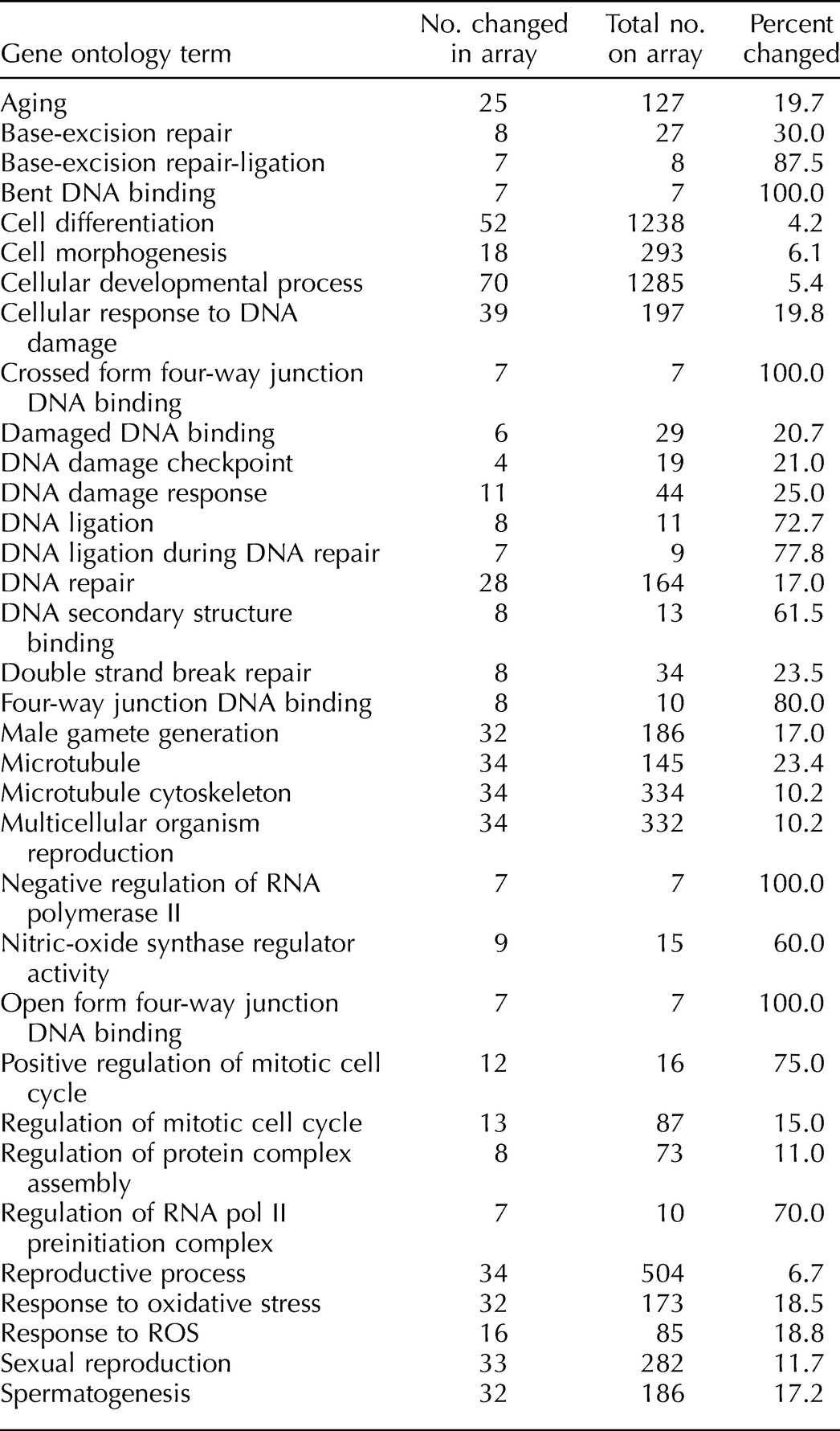
Probe sets that were expressed in both young and aged pachytene spermatocytes and round spermatids and were significantly changed were assessed further and divided according to their function using the Gene Ontology (GO) analysis tool in the Genespring software (Fig. 2). For the round spermatids, there was a wide variety of GO terms highlighted, including signaling, metabolism, development, protein metabolism, and biosynthesis (Fig. 2). There did not appear to be any distinct pathways changed in the round spermatids with age. For the pachytene spermatocytes, however, although a large number of different GO terms were highlighted, including cell differentiation, cell cycle regulation, reproductive processes, and spermatogenesis, there were many GO terms that showed large numbers of changed genes within specific categories; for example, response to stress and regulation of gene expression both had 175 members changed, and for the term response to oxidative stress, 32 out of 170 on the array were changed. In addition, regulation of cell death and apoptosis had 87 and 85 genes significantly altered, respectively. Some of the most predominant GO terms observed for the pachytene spermatocytes were those involved in DNA repair/DNA damage, including DNA ligation, DNA binding, DNA ligation during DNA repair, and BER (Fig. 2 and Table 2). This led us to further concentrate our analysis on DNA repair in the spermatocytes to determine if specific pathways were affected.
FIG. 2.

GO analysis of genes that are at least 1.5-fold changed in pachytene spermatocytes and round spermatids. Pie charts show the prevalence of different GO terms in relation to aging. The color-coded key under the figure describes each section of the pie with its GO subcategory.
TABLE 2.
Gene ontology terms that were observed predominantly in the altered genes (1.5-fold) in aged pachytene spermatocytes.
Of the four DNA repair pathways identified in the “changed genes,” the BER pathway showed the highest change (30%) in terms of percent of genes changed (Fig. 3). Mismatch repair (MMR) had the least number of genes changed, with only one gene having a significant difference in aged spermatocytes. Out of the 60 genes identified by the GO analysis to be involved in DNA damage recognition and repair, 30 were recognized by pathway analysis to have direct interactions (Fig. 4). These were separated into those involved in BER, NER, and double strand break (DSB) repair pathways and others that are involved in signaling (Fig. 4). The DSB pathway was not further studied due to many of the genes also being involved in homologous recombination that takes place in pachytene spermatocytes.
FIG. 3.

The number and percentage of genes changed in aged pachytene spermatocytes with GO terms related to the different DNA repair pathways: BER, MMR, NER, and DSB repair. At the top of each bar, the value indicates the percent of genes significantly changed out of the total number on the array that are related to each pathway.
FIG. 4.

Direct linkages between genes that are identified by GO analysis to be involved in the DNA repair pathways and significantly change in the aged spermatocytes compared with the young. Genes are clustered according to the repair pathway with which they are involved; the BER genes are highlighted in green, the NER are in pink, and the DSB repair genes are in blue.
Aging Causes Changes in NER Gene Expression but Not Protein Levels
Since approximately 20% of the genes in the NER pathway changed with aging (Fig. 3), the expression of four key NER genes was further analyzed. Using quantitative RT-PCR, Ercc1, Xpa, Rpa1, and Ercc4 (Xpf), were found to be significantly upregulated by 2-fold, 1.7-fold, 2.4-fold, and 1.9-fold, respectively (Fig. 5). These changes were consistent with those seen in the microarray. Western blot analysis was done for these NER members; however, the changes observed in mRNA level did not appear to be translated to the protein level for all four (Fig. 5, E–H). Only Ercc4 (Xpf) had a significant increase in protein in aged spermatocytes compared with young; although RPA was increased, this was not significant (Fig. 5, H and G, respectively).
FIG. 5.

Relative mRNA expression of Ercc1 (A), Xpa (B), Rpa1 (C), and Ercc4 (D) in young and aged pachytene spermatocytes. *P < 0.05 and ***P < 0.001. n = 6. Impact of age on NER protein levels: ERCC1 (E), XPA (F), RPA1 (G), and ERCC4 (H). *P < 0.05. n = 5.
The Impact of Age on the BER Pathway
The BER pathway had the highest percentage of genes changed. Therefore, the expression of the key BER genes Ogg1, Ape1, Fen1, and Xrcc1 was assessed by quantitative RT-PCR; the expression of all four was found to be significantly altered in all samples from aged animals (Fig. 6). Ogg1 mRNA was significantly upregulated (Fig. 6A); however, the other three members (Ape1, Fen1, and Xrcc1) in the pathway were significantly downregulated by at least 1.5-fold (Fig. 6, B–D). The protein levels (Fig. 6, E–H) of the aforementioned BER members were also altered: whereas OGG1 protein was not significantly changed (Fig. 6E), APE1, XRCC1, and FEN1 protein levels were significantly downregulated by 2-fold, 2.4-fold, and 1.6-fold, respectively (Fig. 6, F–H). Thus, there is an overall suppression of the key players in the BER pathway in pachytene spermatocytes. In contrast, in round spermatids none of the genes involved in this pathway were altered.
FIG. 6.

Relative mRNA expression of Ogg1 (A), Ape1 (B), Fen1 (C), and Xrcc1 (D) in young and aged pachytene spermatocytes. *P < 0.05 and **P < 0.01. n = 6. Impact of age on BER protein levels: OGG1 (E), APE1 (F), FEN1 (G), and XRCC1 (H). Each Western image shows two representative samples, each from young and aged pachytene spermatocytes. *P < 0.05, **P < 0.01, ***P < 0.001. n = 5.
Aging Causes DNA Damage in Pachytene Spermatocytes and Mature Spermatozoa
Due to the significant changes in expression, at both the mRNA and the protein levels of the components of the BER pathway, we examined DNA damage in the pachytene spermatocytes and spermatozoa. This was done by looking at the oxidized base 8-oxo-dG using immunohistochemistry on testes and immunofluorescence on spermatozoa (Fig. 7). Qualitative analysis of testis sections showed that there appeared to be an increase in both the number of 8-oxodG positive cells and also in the intensity of staining within these cells in the aged testes (Fig. 7A). 8-oxodG was localized to the spermatocytes (black arrows), elongating spermatids (black arrowhead), and also appeared to be present in some peritubular myoid cells (blue arrow). The spermatocytes and elongating spermatids exhibited stronger staining than the peritubular myoid cells. In addition, spermatozoa from the cauda epididymis of aged rats displayed a significant 7.5-fold increase in the number that were positive for 8-oxodG when compared to their younger counterparts (Fig. 7, B and C). This suggests that the BER deficiency results in the production of spermatozoa that have more chromatin damage and hence may be of lower quality.
FIG. 7.

A) 8-oxodG immunoreactivity in the testes from young and aged rats, showing strong staining in spermatocytes (black arrows) and elongating spermatids (black arrowheads) and some weaker staining in peritubular myoid cells (blue arrow) in the aged samples (inset shows negative control). Original magnification ×63. Bar = 50 μm. Qualitative analysis of 8-oxodG (green) immunofluorescence on caudal spermatozoa from young and aged rats counterstained with DAPI (B) and quantitative analysis showing an increase in the number of 8-oxodG-positive spermatozoa in the aged samples (C; P < 0.01, n = 4). Both positive (+ve) and negative (−ve) controls were included.
DISCUSSION
One of the main theories of aging states that aging results from an accumulation of unrepaired DNA lesions; such lesions have been routinely linked to aging in many somatic tissues, including the brain and the liver [25, 26]. The testis appears to be no exception. Once the DNA is damaged, a variety of responses can ensue, including cessation of transcription, cell cycle arrest, mutagenesis, and cell death [27]. Any one of these responses could result in the disruption of spermatogenesis, mutation in the germ line, and ultimately the passing on of genetic errors to offspring. There is an increase in ROS with aging (e.g., in spermatozoa), and this is concomitant with a decrease in antioxidant levels [13, 14], resulting in an imbalance in the antioxidant defence system. Thus, the hypothesis of our study was that the increased oxidative stress previously shown in Brown Norway rats would lead to the accumulation of DNA damage in germ cells and that this would cause gene expression changes with age. Microarray analysis did indeed show that a large selection of genes was differentially regulated in the aged rat spermatocytes (almost 2000) and round spermatids (approximately 500) in comparison to the young. After further analysis using Pathway Studio and Genespring to determine whether specific pathways were affected, it was decided to further investigate these changes specifically in the pachytene spermatocytes, as the analysis of round spermatids did not produce changes in specific pathways.
One of the predominant changes seen in the pachytene spermatocytes with age was in genes involved in the response to oxidative stress and DNA damage. In particular, the NER and BER pathways of DNA repair were highlighted, and so further analysis of these two pathways was undertaken. The BER is split into two subpathways with the involvement of each one depending on the number of bases to be repaired. The short patch BER pathway is involved when only one base is recognized and excised by the glycosylase OGG1 and Ape endonuclease (APE1) helped by XRCC1. This is followed with DNA synthesis by β-polymerase and ligation by DNA ligase III. In the long patch BER pathway several nucleotides are repaired; this results in the generation of a “flap” of DNA and is initiated in much the same way as the short patch but involves FEN1, which is responsible for the removal of the flap. Therefore, the genes/proteins chosen in this study cover both the short and long patch pathways (reviewed in Almeida and Sobol [28] and Zharkov [29]). At the levels of mRNA and protein, the BER pathway was overall downregulated, with the exception of OGG1 (uracil-DNA glycosylase), whereas the NER pathway was upregulated. The BER pathway is likely to be one of the main mechanisms involved in maintaining genomic stability in the male germ line, particularly in aging, as it is responsible for repairing spontaneous base damage caused by oxidative stress; several BER genes have been shown to be highly expressed in male germ cells [30]. Therefore, that changes were seen specifically in this pathway was not unexpected, though the increased Ogg1 mRNA expression in testes from the aged rats when the other BER members were significantly reduced was not anticipated. In the BER pathway, OGG1 encodes the enzyme responsible for the initiation of the excision of 8oxodG, a mutagenic base by-product that can result from exposure to ROS.
To our knowledge, this study is the first to look at the BER pathway specifically in spermatocytes from aged rats. However, previous studies in mice have shown that BER activity in young animals is limited by OGG1 and in the old animals is limited by AP endonuclease [31]. In our study, the AP endonuclease APE1 was significantly reduced both in mRNA and protein expressions. One previous study demonstrated in mice that there was a decline in levels of β-polymerase activity, protein, and RNA with age and that this corresponded to a decline in the BER pathway [32]. However, our study did not show alterations in this particular gene. In contrast, a study using immunohistochemical labelling of germ cells with proteins involved in the BER pathway showed that many of the proteins exhibited higher expression in the germ cells of older men [33]. It is worth noting, however, that these males were attending an andrology clinic for procedures such as inguinal hernia repair and orchidectomy for prostate cancer treatment; these conditions may have confounded the results obtained. In addition, it is extremely difficult to draw conclusions on protein expression based on immunohistochemistry scores.
The reduction of APE1 seen in our study was accompanied by reduced expression of downstream players in the BER pathway such as FEN1 and XRCC1. As the BER pathway is involved in removing the most common DNA lesions (8-oxodG) resulting from oxidative stress, it could be expected that there would also be changes in the occurrence of this lesion in our aged rats. Indeed, this is exactly what we saw both in the testis and in the sperm from aged rats. The 8-oxodG lesion has the ability to mimic the base T in the syn conformation and, accordingly, can pair with A, forming an 8-oxodG:A base pair that can allow its escape from proofreading and the bypass of DNA polymerases [34]. Failure to remove 8-oxodG prior to replication can result in G- to T-transversion mutations [35].
This reduction in the BER pathway may partly (in addition to the aforementioned increased ROS in spermatozoa) account for the increase in DNA damage observed in spermatozoa from older men. Animal studies have shown that transversion mutations can contribute to around 20% of mutations found in young mice; however, this is increased to 40% in aged mice [36, 37]. In addition, chromatin integrity in spermatozoa has also been shown to be altered in aging males using techniques such as the sperm chromatin structure assay, chromomycin A3 assay, and comet analysis [38–40].
In the NER pathway, Xeroderma pigmentosum complementation group C (XPC) initiates the process and acts as a DNA damage sensor and repair-recruitment factor [41] followed by strand separation where XPA verifies and binds the damage in an open DNA conformation and facilitates the assembly of the remainder of the repair machinery [42, 43]. Replication protein A (RPA) stabilizes the opened DNA complex and positions the XPG and ERCC1-ERCC4 endonucleases responsible for the DNA incisions around the lesion [44]. The final steps of NER involve DNA synthesis to fill the gap and ligation of the newly synthesized strand to the original sequence. Despite the increase seen in the mRNAs of NER genes, this was not translated to the protein level; this was unexpected and raises the question as to why there should be differential regulation of the BER and NER pathways at the transcriptional and translational levels. There have been many conflicting studies on the associations between aging and NER [45]. The fact that levels of three out of the four proteins tested were unchanged despite increases in their mRNAs could be accounted for by the fact that the NER pathway is regulated by the ubiquitin-proteasome pathway [46, 47], and in our study 26 ubiquitin-related genes were downregulated, including Ube2a (ubiquitin conjugating enzyme) mRNA, which was reduced by 50%. Ercc4 was the only protein of the four studied that was significantly upregulated in aged spermatocytes. This may be due to its role in pathways other than NER, such as the repair of DNA interstrand cross-links and DSB repair [48]; however, this would need further investigation. NER proteins are regulated by the nonproteolytic activities of the ubiquitin-proteasome pathway; thus, it is a possibility that ubiquitination leads to stabilization of the NER proteins rather than degradation [49]. Further study of this pathway in aging germ cells is warranted. In addition, NER has been shown to take place at a lower level of activity in germ cells than in somatic cells [50], and it has been suggested that, due to the large amount of germ cells in the testis, it may not be necessary to induce the NER pathway to save every cell and that apoptosis may be favoured over NER [50]. This agrees with the previous studies that have shown that aging in the testis of the Brown Norway rat is characterized by an increase in germ cell apoptosis [51]. In addition, it may be more indicative of the type of damage that is being induced in these cells, that is, that it is oxidative DNA damage that induces BER rather than damage that would induce the NER pathway.
The fact that the changes observed in DNA repair pathways in the pachytene spermatocytes were not carried through to the round spermatids may be due to the minimal levels of DNA repair in these cells [50]. In addition, there is evidence of transcriptional silencing and an uncoupling of transcription and translation in the haploid spermatids [52], which may also be a factor in the difference of DNA repair gene/protein expression in round spermatids.
In conclusion, we have utilized a genomewide approach to determine changes in gene expression associated with aging male germ cells; our data indicate that there is an age-related accumulation of DNA damage in the testis, particularly that caused by oxidative stress in the form of 8-oxodG lesions. Furthermore, aging seems to lower the capacity of germ cells to repair such DNA damage, resulting in the production of spermatozoa with increased DNA damage. This is likely to lead to a decline in genome integrity that may be passed on to future generations, specifically the offspring of older males.
ACKNOWLEDGMENTS
The authors wish to Trang Luu for her technical assistance and Genome Quebec for their help with the microarray and bioinformatics.
Footnotes
Supported by grant MOP-89767 from the Canadian Institutes of Health Research (CIHR). C.P. was funded by a CIHR postdoctoral fellowship. B.R. is a James McGill Professor.
REFERENCES
- Molina RI, Martini AC, Tissera A, Olmedo J, Senestrari D, Fiol de Cuneo M, Ruiz RD. Semen quality and aging: analysis of 9.168 samples in Cordoba. Argentina. Arch Esp Urol 2010; 63: 214 222. [PubMed] [Google Scholar]
- Cardona Maya W, Berdugo J, Cadavid Jaramillo A. The effects of male age on semen parameters: analysis of 1364 men attending an andrology center. Aging Male 2009; 12: 100 103. [DOI] [PubMed] [Google Scholar]
- Kidd SA, Eskenazi B, Wyrobek AJ. Effects of male age on semen quality and fertility: a review of the literature. Fertil Steril 2001; 75: 237 248. [DOI] [PubMed] [Google Scholar]
- Syntin P, Robaire B. Sperm structural and motility changes during aging in the Brown Norway rat. J Androl 2001; 22: 235 244. [PubMed] [Google Scholar]
- Ford WC, North K, Taylor H, Farrow A, Hull MG, Golding J. Increasing paternal age is associated with delayed conception in a large population of fertile couples: evidence for declining fecundity in older men. The ALSPAC Study Team (Avon Longitudinal Study of Pregnancy and Childhood). Hum Reprod 2000; 15: 1703 1708. [DOI] [PubMed] [Google Scholar]
- Hassan MA, Killick SR. Effect of male age on fertility: evidence for the decline in male fertility with increasing age. Fertil Steril 2003; 79 (suppl 3): 1520 1527. [DOI] [PubMed] [Google Scholar]
- Klonoff-Cohen HS, Natarajan L. The effect of advancing paternal age on pregnancy and live birth rates in couples undergoing in vitro fertilization or gamete intrafallopian transfer. Am J Obstet Gynecol 2004; 191: 507 514. [DOI] [PubMed] [Google Scholar]
- Baarends WM, van der Laan R, Grootegoed JA. DNA repair mechanisms and gametogenesis. Reproduction 2001; 121: 31 39. [DOI] [PubMed] [Google Scholar]
- Aitken RJ, De Iuliis GN. Origins and consequences of DNA damage in male germ cells. Reprod Biomed Online 2007; 14: 727 733. [DOI] [PubMed] [Google Scholar]
- Fisher HM, Aitken RJ. Comparative analysis of the ability of precursor germ cells and epididymal spermatozoa to generate reactive oxygen metabolites. J Exp Zool 1997; 277: 390 400. [DOI] [PubMed] [Google Scholar]
- Schoneich C. Reactive oxygen species and biological aging: a mechanistic approach. Exp Gerontol 1999; 34: 19 34. [DOI] [PubMed] [Google Scholar]
- Bauche F, Fouchard M, Jegou B. Antioxidant system in rat testicular cells. FEBS Lett 1994; 349: 392 396. [DOI] [PubMed] [Google Scholar]
- Vazquez-Memije ME, Capin R, Tolosa A, El-Hafidi M. Analysis of age-associated changes in mitochondrial free radical generation by rat testis. Mol Cell Biochem 2008; 307: 23 30. [DOI] [PubMed] [Google Scholar]
- Weir CP, Robaire B. Spermatozoa have decreased antioxidant enzymatic capacity and increased reactive oxygen species production during aging in the Brown Norway rat. J Androl 2007; 28: 229 240. [DOI] [PubMed] [Google Scholar]
- Aitken RJ, Clarkson JS. Cellular basis of defective sperm function and its association with the genesis of reactive oxygen species by human spermatozoa. J Reprod Fertil 1987; 81: 459 469. [DOI] [PubMed] [Google Scholar]
- Aitken RJ, Gordon E, Harkiss D, Twigg JP, Milne P, Jennings Z, Irvine DS. Relative impact of oxidative stress on the functional competence and genomic integrity of human spermatozoa. Biol Reprod 1998; 59: 1037 1046. [DOI] [PubMed] [Google Scholar]
- Lindahl T. Instability and decay of the primary structure of DNA. Nature 1993; 362: 709 715. [DOI] [PubMed] [Google Scholar]
- Wang C, Leung A, Sinha-Hikim AP. Reproductive aging in the male Brown Norway rat: a model for the human. Endocrinology 1993; 133: 2773 2781. [DOI] [PubMed] [Google Scholar]
- Wright WW, Fiore C, Zirkin BR. The effect of aging on the seminiferous epithelium of the brown Norway rat. J Androl 1993; 14: 110 117. [PubMed] [Google Scholar]
- Bellve AR, Cavicchia JC, Millette CA, O'Brien DA, Bhatnagar YM, Dym M. Spermatogenic cells of the prepubertal mouse, isolation and morphological characterization. J Cell Biol 1977; 74: 68 85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aguilar-Mahecha A, Hales BF, Robaire B. Expression of stress response genes in germ cells during spermatogenesis. Biol Reprod 2001; 65: 119 127. [DOI] [PubMed] [Google Scholar]
- Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U, Speed TP. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003; 4: 249 264. [DOI] [PubMed] [Google Scholar]
- Bieber AM, Marcon L, Hales BF, Robaire B. Effects of chemotherapeutic agents for testicular cancer on the male rat reproductive system, spermatozoa, and fertility. J Androl 2006; 27: 189 200. [DOI] [PubMed] [Google Scholar]
- Saunders PT, Sharpe RM, Williams K, Macpherson S, Urquart H, Irvine DS, Millar MR. Differential expression of oestrogen receptor alpha and beta proteins in the testes and male reproductive system of human and non-human primates. Mol Hum Reprod 2001; 7: 227 236. [DOI] [PubMed] [Google Scholar]
- Brasnjevic I, Hof PR, Steinbusch HW, Schmitz C. Accumulation of nuclear DNA damage or neuron loss: molecular basis for a new approach to understanding selective neuronal vulnerability in neurodegenerative diseases. DNA Repair (Amst) 2008; 7: 1087 1097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moller P, Lohr M, Folkmann JK, Mikkelsen L, Loft S. Aging and oxidatively damaged nuclear DNA in animal organs. Free Radic Biol Med 2010; 48: 1275 1285. [DOI] [PubMed] [Google Scholar]
- Essers J, Vermeulen W, Houtsmuller AB. DNA damage repair: anytime, anywhere? Curr Opin Cell Biol 2006; 18: 240 246. [DOI] [PubMed] [Google Scholar]
- Almeida KH, Sobol RW. A unified view of base excision repair: lesion-dependent protein complexes regulated by post-translational modification. DNA Repair (Amst) 2007; 6: 695 711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zharkov DO. Base excision DNA repair. Cell Mol Life Sci 2008; 65: 1544 1565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Intano GW, McMahan CA, Walter RB, McCarrey JR, Walter CA. Mixed spermatogenic germ cell nuclear extracts exhibit high base excision repair activity. Nucleic Acids Res 2001; 29: 1366 1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Intano GW, McMahan CA, McCarrey JR, Walter RB, McKenna AE, Matsumoto Y, MacInnes MA, Chen DJ, Walter CA. Base excision repair is limited by different proteins in male germ cell nuclear extracts prepared from young and old mice. Mol Cell Biol 2002; 22: 2410 2418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cabelof DC, Raffoul JJ, Yanamadala S, Ganir C, Guo Z, Heydari AR. Attenuation of DNA polymerase beta-dependent base excision repair and increased DMS-induced mutagenicity in aged mice. Mutat Res 2002; 500: 135 145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Domyati MM, Al-Din AB, Barakat MT, El-Fakahany HM, Xu J, Sakkas D. Deoxyribonucleic acid repair and apoptosis in testicular germ cells of aging fertile men: the role of the poly(adenosine diphosphate-ribosyl)ation pathway. Fertil Steril 2009; 91: 2221 2229. [DOI] [PubMed] [Google Scholar]
- Hsu GW, Ober M, Carell T, Beese LS. Error-prone replication of oxidatively damaged DNA by a high-fidelity DNA polymerase. Nature 2004; 431: 217 221. [DOI] [PubMed] [Google Scholar]
- Neeley WL, Essigmann JM. Mechanisms of formation, genotoxicity, and mutation of guanine oxidation products. Chem Res Toxicol 2006; 19: 491 505. [DOI] [PubMed] [Google Scholar]
- Walter CA, Intano GW, McCarrey JR, McMahan CA, Walter RB. Mutation frequency declines during spermatogenesis in young mice but increases in old mice. Proc Natl Acad Sci U S A 1998; 95: 10015 10019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter CA, Intano GW, McMahan CA, Kelner K, McCarrey JR, Walter RB. Mutation spectral changes in spermatogenic cells obtained from old mice. DNA Repair (Amst) 2004; 3: 495 504. [DOI] [PubMed] [Google Scholar]
- Schmid TE, Eskenazi B, Baumgartner A, Marchetti F, Young S, Weldon R, Anderson D, Wyrobek AJ. The effects of male age on sperm DNA damage in healthy non-smokers. Hum Reprod 2007; 22: 180 187. [DOI] [PubMed] [Google Scholar]
- Zubkova EV, Robaire B. Effects of ageing on spermatozoal chromatin and its sensitivity to in vivo and in vitro oxidative challenge in the Brown Norway rat. Hum Reprod 2006; 21: 2901 2910. [DOI] [PubMed] [Google Scholar]
- Wyrobek AJ, Eskenazi B, Young S, Arnheim N, Tiemann-Boege I, Jabs EW, Glaser RL, Pearson FS, Evenson D. Advancing age has differential effects on DNA damage, chromatin integrity, gene mutations, and aneuploidies in sperm. Proc Natl Acad Sci U S A 2006; 103: 9601 9606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugasawa K, Ng JM, Masutani C, Iwai S, van der Spek PJ, Eker AP, Hanaoka F, Bootsma D, Hoeijmakers JH. Xeroderma pigmentosum group C protein complex is the initiator of global genome nucleotide excision repair. Mol Cell 1998; 2: 223 232. [DOI] [PubMed] [Google Scholar]
- Tanaka K, Miura N, Satokata I, Miyamoto I, Yoshida MC, Satoh Y, Kondo S, Yasui A, Okayama H, Okada Y. Analysis of a human DNA excision repair gene involved in group A xeroderma pigmentosum and containing a zinc-finger domain. Nature 1990; 348: 73 76. [DOI] [PubMed] [Google Scholar]
- Miyamoto I, Miura N, Niwa H, Miyazaki J, Tanaka K. Mutational analysis of the structure and function of the xeroderma pigmentosum group A complementing protein: identification of essential domains for nuclear localization and DNA excision repair. J Biol Chem 1992; 267: 12182 12187. [PubMed] [Google Scholar]
- de Laat WL, Appeldoorn E, Sugasawa K, Weterings E, Jaspers NG, Hoeijmakers JH. DNA-binding polarity of human replication protein A positions nucleases in nucleotide excision repair. Genes Dev 1998; 12: 2598 2609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorbunova V, Seluanov A, Mao Z, Hine C. Changes in DNA repair during aging. Nucleic Acids Res 2007; 35: 7466 7474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillette TG, Yu S, Zhou Z, Waters R, Johnston SA, Reed SH. Distinct functions of the ubiquitin-proteasome pathway influence nucleotide excision repair. EMBO J 2006; 25: 2529 2538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed SH, Gillette TG. Nucleotide excision repair and the ubiquitin proteasome pathway—do all roads lead to Rome? DNA Repair (Amst) 2007; 6: 149 156. [DOI] [PubMed] [Google Scholar]
- Niedernhofer LJ, Odijk H, Budzowska M, van Drunen E, Maas A, Theil AF, de Wit J, Jaspers NG, Beverloo HB, Hoeijmakers JH, Kanaar R. The structure-specific endonuclease Ercc1-Xpf is required to resolve DNA interstrand cross-link-induced double-strand breaks. Mol Cell Biol 2004; 24: 5776 5787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang QE, Zhu Q, Wani G, El-Mahdy MA, Li J, Wani AA. DNA repair factor XPC is modified by SUMO-1 and ubiquitin following UV irradiation. Nucleic Acids Res 2005; 33: 4023 4034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu G, Spivak G, Mitchell DL, Mori T, McCarrey JR, McMahan CA, Walter RB, Hanawalt PC, Walter CA. Nucleotide excision repair activity varies among murine spermatogenic cell types. Biol Reprod 2005; 73: 123 130. [DOI] [PubMed] [Google Scholar]
- Wang C, Sinha Hikim AP, Lue YH, Leung A, Baravarian S, Swerdloff RS. Reproductive aging in the Brown Norway rat is characterized by accelerated germ cell apoptosis and is not altered by luteinizing hormone replacement. J Androl 1999; 20: 509 518. [PubMed] [Google Scholar]
- Steger K. Transcriptional and translational regulation of gene expression in haploid spermatids. Anat Embryol (Berl) 1999; 199: 471 487. [DOI] [PubMed] [Google Scholar]