

Abstract
A new tick species belonging to the genus Dermacentor Koch, 1844, namely, Dermacentor limbooliati n. sp., is described. Adults of this species are similar to those of Dermacentor auratus Supino, 1897 and Dermacentor compactus Neumann, 1901, with which it was previously confused. Males of D. limbooliati may be distinguished from those of D. auratus and D. compactus by the following suite of characters: relatively broad conscutum with slightly straightened lateral sides, conscutum widest approximately at mid-length, oval shape of pseudoscutum, central brown patch in the center of pseudoscutum broad and diffused and not continuous with central patch in posteromedian area, conscutum posterior to pseudoscutum rugose, wide and blunt internal spur on coxa I, relatively long, narrow, and pointed external spur on coxa I, numerous internal spurs on coxa IV and trochanter I with moderate and broadly triangular spur with tapering apex. Females of D. limbooliati may be distinguished from those of D. auratus and D. compactus by the following suite of characters: rounded shape of scutum, central brown patch broad and diffused in the center of scutum, relatively long alloscutum setae, genital aperture moderately narrow V-shaped with preatrial fold bulging, wide and blunt internal spur on coxa I, relatively long, narrow, and pointed external spur on coxae I and trochanter I with moderate and broadly triangular spur with tapering apex. D. limbooliati is known from Malaysia and Vietnam where the adults were collected from vegetation, Sus scrofa resting beds, a human, and clothing. The immature stages remain unknown.
Keywords: Dermacentor limbooliati n. sp., description, Malaysia, Vietnam
Introduction
The systematics of species in the genus Dermacentor Koch, 1844 is still debatable in many aspects, and there are many gaps in our knowledge of this genus. All currently valid Oriental species of the genus, such as D. atrosignatus Neumann, 1906, D. auratus Supino, 1897, D. compactus Neumann, 1901, D. confragus (Schulze, 1933), D. steini (Schulze, 1933), and D. taiwanensis Sugimoto, 1935 have common distinct morphological features and were assigned to the subgenus Indocentor Schulze, 1933. Systematics of this subgenus was critically revised, and adults of most of the species were redescribed in a series of publications by Wassef and Hoogstraal (1983, 1984a, 1984b, 1986a, 1986b). Recently Sun and Xu (2013) provided short diagnoses to Chinese Indocentor species that are partially based on previously mentioned works of Wassef and Hoogstraal. Larvae and nymphs for most of these species remain unknown.
Recent re-examination of extensive holdings of Oriental Dermacentor ticks in the United States National Tick Collection (USNTC) revealed a morphologically distinct new species of this genus, originating from the northern part of continental Malaysia and Central Vietnam. Additional specimens belonging to this species from Vietnam were lately found in the collection of the Zoological Museum of Moscow State University. Male and female of this species are described below.
Materials and Methods
The material examined for the description is summarized in Table 1 . Only field-collected ticks were available for study. The specimens that were examined are deposited in the USNTC (The James H. Oliver, Jr. Institute for Coastal Plain Science, Georgia Southern University, Statesboro, Georgia, USA) and the Zoological Museum of Moscow State University (Moscow, Russia). For comparative purposes near 1,200 males and 770 females of D. auratus originating from Cambodia, China, India, Indonesia, Laos, Malaysia, Myanmar, Nepal, Sri Lanka, Thailand, and Vietnam and near 4,800 males and 5,900 females of D. compactus collected in Malaysia and Indonesia were examined.
Table 1.
Dermacentor limbooliati n. sp., material examined
| No. ticks |
Host | Locality; altitude (m) | Date | Collectora | Accession no. | |
|---|---|---|---|---|---|---|
| ♂ | ♀ | |||||
| Malaysia | ||||||
| 1 | Sus scrofa bed | Kelantan, 5 mi S of Kuala Krai, Sungei Durian Forest Reserve | 17.XI.1967 | BLL | USNMENT 00714038 | |
| 1 | Vegetation | Perlis, Padang Besar, Mata Ayer Forest Reserve | 25.X.1967 | BLL | USNMENT 00714998 | |
| 1 | Vegetation | Perlis, Padang Besar, Mata Ayer Forest Reserve | 26.XI.1967 | BLL | USNMENT 00714772 | |
| 1 | Vegetation | Perlis, Padang Besar, Mata Ayer Forest Reserve | 26.XI.1967 | BLL | USNMENT 00714900 | |
| 1 | 1 | Vegetation | Perlis, Padang Besar, Mata Ayer Forest Reserve | 27.XI.1967 | BLL | USNMENT 00714484 |
| 1 | Vegetation | Perlis, Kangar, Kaki Bukit Forest Reserve | 29.XI.1968 | BLL | USNMENT 00714802 | |
| 1 | Vegetation | Perlis, Kangar, Kaki Bukit Forest Reserve | 28.XI.1968 | BLL | USNMENT 00714170 | |
| Vietnam | ||||||
| 1 | Human | Central Highlands, near Cambodian border | 17.XI.1966 | USNMENT 00714664 | ||
| 1 | 1 | Vegetation | Gia Lai, An Khe, Buon Luoi | 20.X.1986 | Tdi-4725 | |
| 1 | Clothing | Binh Phuoc, 13 km NE of Bu Gia Map village, Bu Gia Map National Park; 540 | IV.2009 | AVA | Tdi-5801 | |
| 6 | 6 | Total | ||||
aAVA, A.V. Abramov; BLL, B.L. Lim.
The adults were studied by means of a stereoscopic microscope (Olympus SZX16, Olympus Corporation, Tokyo, Japan). Because of a very limited number of specimens available, many of them damaged or too fragile due to inadequate storage conditions in the past, we avoided using the scanning electron microscopy in this study. Measurements for the male and female are given in millimeters. The measurements are arranged as follows: minimum–maximum (mean ± SD, n = number of specimens measured). All illustrations have been drawn by D.A. Apanaskevich and colored and edited by M.A. Apanaskevich.
Dermacentor limbooliati n. sp.
Male (Figs. 1A and 2)
Fig. 1.
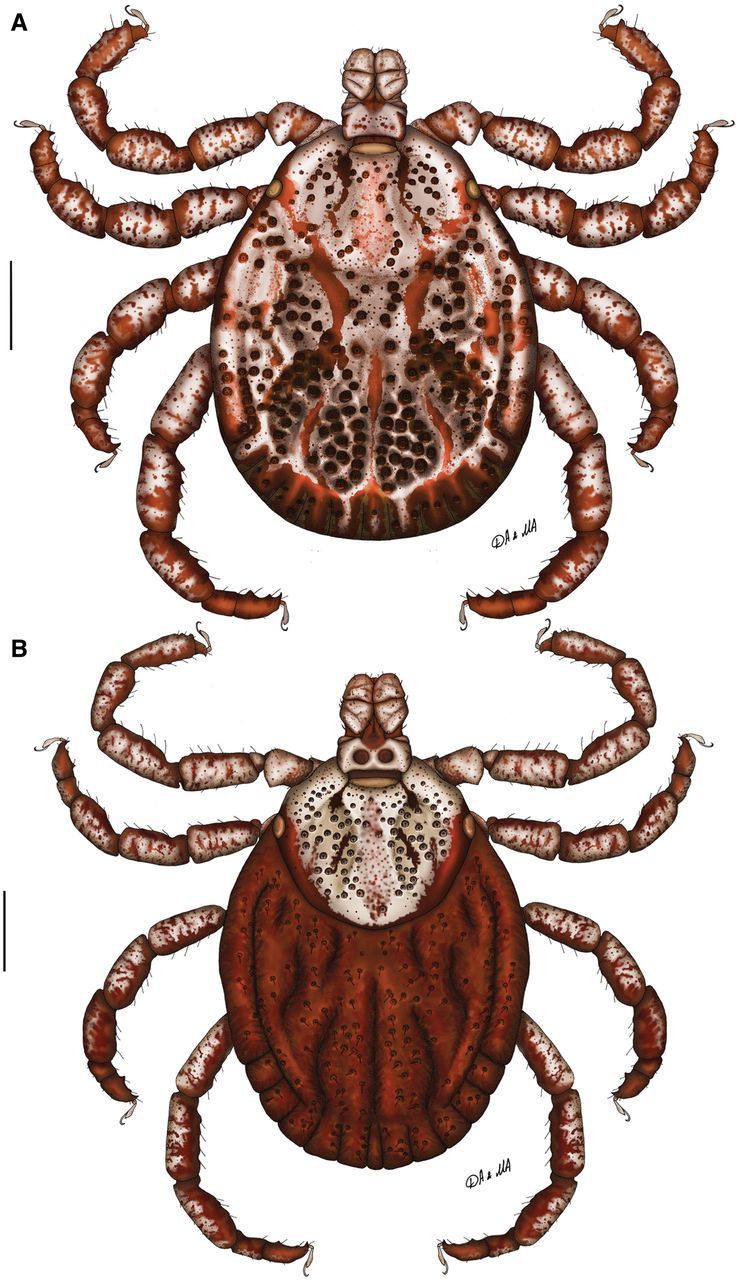
Dermacentor limbooliati n. sp., dorsally. (A) Male. (Malaysia, Kelantan Province, 5 mi S of Kuala Krai, Sungei Durian Forest Reserve, USNMENT 00714038) (B) Female. (Vietnam, Central Highlands, near Cambodian border, USNMENT 00714664). Scale bar = 1 mm.
Conscutum (Fig. 1A): broadly oval with slightly straitened lateral margins, widest approximately at mid-length; distance from scapular apices to posterior margin of conscutum 3.69–4.62 (4.26 ± 0.37; n = 6), maximum width 2.81–3.56 (3.25 ± 0.28; n = 6), ratio length to width 1.26–1.38 (1.31 ± 0.04; n = 6). Coloration as illustrated: ornamentation moderate to intensive, light- to dark-brown background forms several patches often with indistinct hazy margins; a pair of narrow cervical patches extending from cervical pits to margin of pseudoscutum; a pair of large narrow arcuate patches extending from eyes posteromedially to midlength of conscutum and diverging posterolaterally to the level of first festoons; posteromedian margin of pseudoscutum clearly indicated by narrow hazy strip; central patch on pseudoscutum broad, hazy; lateral field with two brown patches: anterior narrow patch extending along the most lateral margin from eye to midlength where it extends medially to lateral groove and posterior roughly rectangular patch; hazy indistinct patches medially to lateral grooves at conscutal midlength; narrow brown stripes in posteromedian and paramedian regions; first festoon mostly ivory colored, median and paramedian festoons have clear ivory colored spot, other festoons may have some hazy ivory colored spots; all punctations light- to dark-brown. Central field posterior to pseudoscutum is flat giving a concave appearance to conscutum; combination of elevations in this field with large deep punctations gives it the appearance of a rugose and uneven surface. Cervical grooves shallow; a pair of central depressions, and a second posterior pair that correspond to paramedian grooves; lateral grooves superficial, but distinct posteriorly where they are aligned with large punctations; 11 distinct festoons; median festoon slightly narrower than paramedian. Very large and deep punctations moderately dense, distributed over entire conscutum but denser laterally and posteriorly; large punctations on pseudoscutum moderately dense to fairly sparse; fine punctations dense, evenly distributed. Eyes oval, very slightly convex, at anterior one-fifth of conscutal length. Setae relatively short and inconspicuous. Spiracular plates (Fig. 2A): suboval; dorsal prolongation moderately long and broad, with obvious broad not-perforated widening anteriorly; perforations very small and fairly numerous. Sclerotized plaques on festoons ventrally intensively ornated. Gnathosoma (Figs. 1A, 2B and C): length from palpal apices to cornual apices dorsally 0.89–1.06 (0.99 ± 0.06; n = 6), width of basis capituli 0.62–0.70 (0.67 ± 0.04; n = 6), ratio length to width 1.43–1.52 (1.47 ± 0.04; n = 6). Basis capituli (Figs. 1A, 2B and C): dorsally subrectangular; posterior margin nearly straight; length 0.38–0.46 (0.41 ± 0.03; n = 6), ratio width to length 1.52–1.69 (1.63 ± 0.07; n = 6); cornua moderately broad, short, equivalent to a proportion of 8.00–8.67 (8.44 ± 0.28; n = 6) of length of basis capituli; dorsally with extensive whitish enameling. Basis capituli ventrally subrectangular; posterior margin convex. Palpi (Figs. 1A, 2B and C): short, broad; length dorsally (I–III segments) 0.52–0.60 (0.57 ± 0.03; n = 6), width 0.29–0.35 (0.32 ± 0.02; n = 6), ratio length to width 1.71–1.85 (1.78 ± 0.05; n = 6), length of segments in descending order: 2, 3, 1, 4; segment I well-developed ventrally; segment II narrower at base and thereafter widening, without clear denticle at posterior margin dorsally; segment III subrectangular with broadly rounded apex; segments II and III with extensive whitish enameling on dorsal surfaces. Hypostome (Fig. 2C): club-shaped; dental formula 3/3. Legs (Fig. 1A): of medium length, moderately robust; with extensive whitish enameling mostly on dorsal and lateral aspects of leg segments. Coxae (Fig. 2D): coxa I with relatively short widely spaced internal and external spurs; internal spur broadly triangular with narrowly rounded apex and slightly shorter than external spur; external spur narrowly triangular with tapering apex, slightly curved medially; both spurs of coxa I generally directed posteriorly; coxae II and III each with triangular external and internal spurs with tapering apex, external spur slightly larger than internal; coxa IV with moderate in size triangular external spur with tapering apex and with several (ca. 3) moderate in size triangular internal spurs with tapering apex; coxa IV enlarged, ratio length to width 0.83–0.97 (0.91 ± 0.04; n = 6); coxae, especially coxae I, with small spots of whitish enameling. Trochanter I (Figs. 1A and 2E) with moderate, broadly triangular dorsal spur with tapering apex. Genu IV and Tibia IV (Fig. 1A) with two rows of short projections ventrally. Genu IV length 0.91–1.14 (1.03 ± 0.10; n = 5), width 0.38–0.48 (0.44 ± 0.04; n = 5), ratio length to width 2.31–2.40 (2.37 ± 0.04; n = 5).
Fig. 2.

Dermacentor limbooliati n. sp., male (Malaysia, Kelantan Province, 5 mi S of Kuala Krai, Sungei Durian Forest Reserve, USNMENT 00714038). (A) Spiracular plate. Scale bar = 1 mm. Arrows show orientation of spiracular plate (a—anterior; d—dorsal). (B) Gnathosoma dorsally. Scale bar = 1 mm. (C) Gnathosoma ventrally. Scale bar = 1 mm. (D) Coxae. Scale bar = 1 mm. (E) Trochanter I. Scale bar = 1 mm.
Female (Figs. 1B and 3)
Fig. 3.

Dermacentor limbooliati n. sp., female (Vietnam, Central Highlands, near Cambodian border, USNMENT 00714664). (A) Genital aperture. Scale bar = 0.5 mm. (B) Spiracular plate. Scale bar = 1 mm. Arrows show orientation of spiracular plate (a—anterior; d—dorsal). (C) Gnathosoma dorsally. Scale bar = 1 mm. (D) Gnathosoma ventrally. Scale bar = 1 mm. (E) Coxae. Scale bar = 1 mm. (F) Trochanter I. Scale bar = 1 mm.
Idiosoma (Fig. 1B): broadly oval, widest approximately at mid-length. Scutum (Fig. 1B): length 1.90–2.28 (2.13 ± 0.12; n = 6), width 2.15–2.56 (2.41 ± 0.15; n = 6), ratio length to width 0.85–0.90 (0.88 ± 0.02; n = 6), margins in anterior half of scutum diverge posteriorly, thereafter gradually converging to broadly rounded posterior margin, slight posterolateral angular projections. Coloration of scutum: ornamentation very intensive, brown colored patches arranged as follows: two pairs of small patches in cervical pits, two narrow patches in cervical fields, large and broad central patch extending from anterior margin to posterior margin of scutum hazy with speckles of darker little patches, two narrow patches bordering posterior margin of scutum extending from just anterior to eyes and connecting at the posterior apex of scutum. Cervical grooves distinct, moderately deep. Surface of scutum slightly rugose; very large and deep punctations moderately dense to fairly sparse and situated mostly in cervical grooves; fine punctations dense, evenly distributed over scutum. Eyes oval, very slightly convex, positioned at mid-length. Setae relatively sparse and short. Alloscutum (Fig. 1B): as illustrated; 11 festoons. Setae of alloscutum relatively long (ca. 0.18), moderately dense. Genital aperture (Fig. 3A): at level of coxae II, moderately narrow V-shaped, semioval sclerites bordering genital aperture laterally distinct; preatrial fold clearly bulging. Spiracular plates (Fig. 3B): suboval; dorsal prolongation moderately long and broad, broadly rounded to its apex, with obvious broad not-perforated widening anteriorly; perforations very small and fairly numerous. Gnathosoma (Figs. 1B, 3C and D): length from palpal apices to posterior margin of basis capituli dorsally 1.07–1.24 (1.18 ± 0.06; n = 6), width of basis capituli 0.80–0.91 (0.87 ± 0.04; n = 6), ratio length to width 1.32–1.42 (1.36 ± 0.04; n = 6). Basis capituli (Figs. 1B, 3C and D): dorsally subrectangular; posterior margin nearly straight; length 0.42–0.48 (0.46 ± 0.02; n = 6), ratio width to length 1.83–1.96 (1.90 ± 0.05; n = 6); cornua very short, broad, comprising a 13.00–24.00 (19.61 ± 4.57; n = 6) proportion of length of basis capituli; dorsally intensively ornate with whitish enameling. Porose areas moderate, circular, deeply sunken with clearly circumscribed borders, separated by space less than their width. Basis capituli ventrally subrectangular, with convex posterior margin. Palpi (Figs. 1B, 3C and D): short and broad; length dorsally (I–III segments) 0.69–0.78 (0.75 ± 0.04; n = 6), width 0.34–0.42 (0.36 ± 0.03; n = 6), ratio length to width 1.81–2.29 (2.07 ± 0.17; n = 6), length of segments in descending order: 2, 3, 1, 4; segment I well developed ventrally; segment II narrower at base and thereafter parallel-sided, without clear denticle at posterior margin dorsally; segment III broad, subrectangular with broadly rounded apex; segments II and III with intensive whitish enameling on dorsal surfaces. Hypostome (Fig. 3D): club-shaped; dental formula 3/3. Legs (Fig. 1B): of medium length, slender; intensively ornate with whitish enameling on dorsal and lateral aspects of leg segments. Coxae (Fig. 3E): coxa I with relatively short widely spaced internal and external spurs; internal spur broadly triangular with narrowly rounded apex and slightly shorter than external spur; external spur narrowly triangular with tapering apex, slightly curved medially; both spurs of coxa I generally directed posteriorly; coxae II and III each with triangular external and internal spurs with tapering apex, external spur slightly larger than internal; coxa IV with moderate in size triangular external and internal spurs with tapering apex; coxae, especially coxae I, with small spots of ivory enameling. Trochanter I (Fig. 3F) with moderate, broadly triangular dorsal spur with tapering apex. Genu IV length 0.96–1.18 (1.09 ± 0.09; n = 4), width 0.40–0.48 (0.43 ± 0.04; n = 4), ratio length to width 2.40–2.80 (2.54 ± 0.18; n = 4).
Nymph and Larva remain unknown
HOLOTYPE
Male, from vegetation, Kaki Bukit Forest Reserve (6° 39′ N; 100° 12′ E), Kangar, Perlis Province, Malaysia, 29.XI.1968, B.L. Lim; deposited in the USNTC (USNMENT 00714802).
ALLOTYPE
Female, from human, near Cambodian border, Central Highlands, Vietnam, 17.XI.1966; deposited in the USNTC (USNMENT 00714664).
PARATYPES
Malaysia: one male and one female, from vegetation, Mata Ayer Forest Reserve (6° 40′ N; 100° 15′ E), Padang Besar, Perlis Province, 27.XI.1967, B.L. Lim; deposited in the USNTC (USNMENT 00714484); one female, from vegetation, Mata Ayer Forest Reserve, Padang Besar, Perlis Province, 26.XI.1967, B.L. Lim; deposited in the USNTC (USNMENT 00714772); one female, from vegetation, Mata Ayer Forest Reserve, Padang Besar, Perlis Province, 26.XI.1967, B.L. Lim; deposited in the USNTC (USNMENT 00714900); one male, from vegetation, Mata Ayer Forest Reserve, Padang Besar, Perlis Province, 25.X.1967, B.L. Lim; deposited in the USNTC (USNMENT 00714998); one male, from vegetation, Kaki Bukit Forest Reserve, Kangar, Perlis Province, 28.XI.1968, B.L. Lim; deposited in the USNTC (USNMENT 00714170); one male, from Sus scrofa resting bed, Sungei Durian Forest Reserve (5° 35′ N; 102° 20′ E), 5 mi S of Kuala Krai, Kelantan Province, 17.XI.1967, B.L. Lim; deposited in the USNTC (USNMENT 00714038). Vietnam: one male and one female, from vegetation, Buon Luoi, An Khe District, Gia Lai Province, 20.X.1986; deposited in the Zoological Museum of Moscow State University (Tdi-4725); one female, from clothing, Bu Gia Map National Park (12° 11′ N; 107° 12′ E), 13 km NE of Bu Gia Map Village, Binh Phuoc Province, Vietnam, 540 m alt., IV.2009, A.V. Abramov; deposited in the Zoological Museum of Moscow State University (Tdi-5801).
Distribution and Hosts
The collection data for D. limbooliati are those of the type series (Table 1). This species is confined to Malaysia (Kelantan and Perlis Provinces) and Vietnam (Bing Phuoc and Gia Lai Provinces; Fig. 4). Visible disjunction in the distribution of D. limbooliati is most probably artificial and can be explained by the absence or extreme scarcity of available Dermacentor collections from Cambodia, Laos, and Thailand. We hope that our work will promote tick studies in the region and this new species will also be found in these countries. Single females were collected from a human (an unengorged female without the indication whether it was attached to the host or not) and clothing. All other adults were collected from vegetation and Sus scrofa resting bed. Most probably as in the case of other Oriental Dermacentor species the major host for the adults is wild boar, Sus scrofa L.
Fig. 4.

Dermacentor limbooliati n. sp., map of geographical distribution. Star shows type locality, filled circles show confirmed localities.
Most of the adult ticks were collected during October and November while one female was found in April.
Etymology
The species is named after Dr. Lim Boo Liat, Malaysian zoologist who made invaluable Dermacentor collections in Malaysia during 1960s–1970s.
Related Species
Morphologically the adults of D. limbooliati are most similar to those of D. auratus and D. compactus. Adults of these three species have widely spaced internal and external spurs on coxa I.
Both males and females of D. limbooliati can be readily distinguished from those of D. auratus and D. compactus by a relatively short and blunt triangular internal spur and a long, narrow and tapering external spur on coxa I (shorter, broader, and blunt external spur in D. auratus and narrow, pointed internal spur in D. compactus). Females of D. limbooliati can easily be distinguished from those of D. auratus and D. compactus by the moderately narrow V-shaped genital aperture with bulging preatrial fold and distinct semioval sclerites bordering the genital aperture (broadly U-shaped with slightly bulging preatrial fold in D. auratus and very small and narrow U-shaped with flat preatrial fold in D. compactus; in both species sclerites not clearly visible).
Additionally males of D. limbooliati can be distinguished from those of D. auratus and D. compactus by the following suite of characters: conscutum with slightly straightened lateral sides and it widest approximately at mid-length (lateral sides convex and it widest posterior to mid-length in D. auratus), oval shape of pseudoscutum (broadly trapezium-like in D. auratus), central brown patch on conscutum wide and diffused in the center of pseudoscutum, separate from center patch in posteromedian area (narrow distinct line through the center of pseudoscutum often continuous to posteromedian area in D. auratus; not distinguished in D. compactus); conscutum posterior to pseudoscutum rugose (smooth in D. compactus); numerous internal spurs on coxa IV (only 1 in D. compactus) and trochanter I with moderate, broadly triangular spur with tapering apex (very short, broad spur with blunt apex in D. compactus).
Additionally females of D. limbooliati can be distinguished from those of D. auratus and D. compactus by the following suite characters: more rounded shape of conscutum (transversally oval in D. compactus), central patch of scutum wide and diffused in the center of scutum (narrow distinct line through the center of scutum in D. auratus; not distinguished in D. compactus), long and distinct alloscutum setae (very short and indistinct setae in D. compactus) and trochanter I with moderate, broadly triangular spur with tapering apex (very short, broad spur with blunt apex in D. compactus).
Acknowledgments
We are most grateful to Dr. Olga V. Voltzit (Zoological Museum, Moscow State University, Moscow, Russia) for making her specimens available for examination. We are in debt to Dr. Gerrit Uilenberg for his careful editing of the manuscript. Dmitry A. Apanaskevich’s part of this project was supported by Grant R15AI096317 from the National Institute of Allergy and Infectious Diseases. The content is the sole responsibility of the authors and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases or the National Institutes of Health.
References Cited
- Sun Y., Xu R. 2013. The genus Dermacentor and the subgenus Indocentor (Acari: Ixodidae) from China. Oriental Insects 47: 155–168. [Google Scholar]
- Wassef H. Y., Hoogstraal H. 1983. Dermacentor (Indocentor) compactus (Acari: Ixodoidea: Ixodidae): identity of male and female. J. Med. Entomol. 20: 648–652. [DOI] [PubMed] [Google Scholar]
- Wassef H. Y., Hoogstraal H. 1984a. Dermacentor (Indocentor) auratus (Acari: Ixodoidea: Ixodidae): identity of male and female. J. Med. Entomol. 21: 169–173. [DOI] [PubMed] [Google Scholar]
- Wassef H. Y., Hoogstraal H. 1984b. Dermacentor (Indocentor) atrosignatus (Acari: Ixodoidea: Ixodidae): identity of male and female. J. Med. Entomol. 21: 586–591. [DOI] [PubMed] [Google Scholar]
- Wassef H. Y., Hoogstraal H. 1986a. Dermacentor (Indocentor) taiwanensis (Acari: Ixodoidea: Ixodidae): identity of male and female. J. Med. Entomol. 23: 173–177. [DOI] [PubMed] [Google Scholar]
- Wassef H. Y., Hoogstraal H. 1986b. Dermacentor (Indocentor) steini (Acari: Ixodoidea: Ixodidae): identity of male and female. J. Med. Entomol. 23: 532–537. [DOI] [PubMed] [Google Scholar]