

Abstract
Reactions to individuals who possess features associated with multiple racial groups may be particularly susceptible to external contextual influences, leading to meaningfully different racial perceptions and judgments in different situations. In the present study, we found that an extrinsic race-label cue not only changed evaluative associations activated by a racially ambiguous face, but also changed quickly occurring neural responses sensitive to racial perception. Behaviorally, prototypical Black faces and racially ambiguous faces labeled as Black activated more negative implicit associations than prototypical White faces and racially ambiguous faces labeled as White. Neurally, prototypical faces and racially ambiguous faces cued with the same race elicited similar responses. Specifically, prototypical Black and racially ambiguous faces labeled as Black elicited larger P200s but smaller N200s than prototypical White and racially ambiguous faces labeled as White. These results show that racial perception can be changed by an external cue and this, in turn, influences subsequent evaluative reactions.
Keywords: racial categorization, multiracial faces, racial bias, ERPs
INTRODUCTION
There is a robust tendency to think in more categorical than individuating terms when encountering racial outgroup than ingroup members (Brewer, 1988; Fiske and Neuberg, 1990). Consequences of this categorical thinking include more quickly classifying outgroup than ingroup members in terms of race (e.g. Levin, 1996; Kubota and Ito, 2007), implicitly activating stereotypes and negative evaluations of outgroup members (e.g. Fazio et al., 1995; Wittenbrink et al., 2001), and poorer memory for outgroup members (e.g. Malpass and Kravitz, 1969; Levin, 2000). Differences in perceptual expertise may contribute to this categorical processing. Perceivers are assumed to typically have more experience interacting with ingroup members, producing qualitative differences in how ingroup and outgroup members are processed (Rhodes et al., 1989; Valentine, 1991; Tanaka et al., 2004).
At the same time, research on social cognitive mechanisms suggests that differential perceptual expertise cannot fully account for the divergent behavioral responses to ingroup and outgroup members (e.g. Hugenberg et al., 2010). Instead, this research shows the influence of motivational and situational factors on responses to outgroup members, as when racial outgroup members portrayed in positive contexts or roles elicit relatively less negative implicit evaluations (Wittenbrink et al., 2001; Barden et al., 2004). These kinds of contextual effects provide a useful framework for examining responses to individuals who cannot be easily categorized into a single racial category because they possess features strongly associated with more than one group. We propose that such extrinsic factors might be particularly influential in determining responses to multiracial and other individuals with racially ambiguous physical features. In this article, we examine how such cues can affect racial perceptions and implicit evaluations of individuals with racially ambiguous facial features.
Extant research supports the influence of extrinsic features on reactions to racially ambiguous individuals. Ito et al. (2011) found that manipulating the racial context in which racially ambiguous faces were encountered affected both perceptions of racial prototypicality and implicit evaluative associations. The contextual manipulation consisted of showing either Black or White faces in conjunction with the racially ambiguous faces. A contrast effect was observed, with racially ambiguous faces perceived as more prototypical of Blacks and eliciting more negative implicit evaluative associations when viewed within a context of White faces. When viewed within a context of Black faces, however, the same racially ambiguous faces were perceived as more prototypical of Whites and elicited more positive associations (see also Rhodes et al., 2010). Other research shows that racially ambiguous faces labeled as ingroup members are assimilated to the ingroup, remembered better and recalled as possessing more ingroup prototypical features (Eberhardt et al., 2003; Pauker et al., 2009).1 Similar context and labeling manipulations also influence configural processing, with racially ambiguous faces shown in contrast to outgroup members or labeled as ingroup members eliciting more configural processing (Michel et al., 2007, 2010). Since configural processing is more likely with ingroup members, these results demonstrate the influence of context on the way in which racially ambiguous faces are encoded.
To date, these studies have focused either on showing how extrinsic factors influence the encoding of racially ambiguous individuals (Levin and Banaji, 2006; Michel et al., 2007, 2010) or on subsequent behavioral responses (Eberhardt et al., 2003; Pauker et al., 2009; Ito et al., 2011), but not both. Here, we attempt to directly link the two by having participants complete a sequential priming task in which faces of Black, White and racially ambiguous Black-White men were shown before positive and negative words. Before each face prime, an extrinsic cue consisting of the label Black or White was shown (cf. Eberhardt et al., 2003; Pauker et al., 2009). Event-related potentials (ERPs) were recorded to the face primes and used to examine on-line neural changes in racial perception in response to the extrinsic cue. Numerous past ERP studies show that two components occurring within the first 300 ms of face perception—the P200 and N200—consistently differentiate between Black and White faces (Ito and Urland, 2003, 2005; Willadsen-Jensen and Ito, 2006, 2008; Dickter and Bartholow, 2007; Kubota and Ito, 2007). In the context of racial categorization, the P200 may reflect selective attention to novel, outgroup, faces whereas the N200 may reflect individuating processes associated with ingroup faces [see Ito and Bartholow (2009) for a review]. If contextual information succeeds in changing the perception of the racially ambiguous faces, effects should be seen in ERPs and behavioral responses. Specifically, racially ambiguous faces labeled as Black should elicit the same neural and behavioral responses as prototypical Black (i.e. unambiguous) faces, whereas racially ambiguous faces labeled as White should elicit the same neural and behavioral responses as prototypical White faces. Such an examination meets two goals. The first is to establish a direct link between the influence of extrinsic cues on racial perceptions and race-related behavioral responses. The second is to better understand factors that influence responses to individuals with less prototypical racial features. If extrinsic cues can fundamentally change racial perceptions and subsequent behavioral responses, this implies the potential for a good deal of variability in responses that are elicited by individuals with less racially prototypical features depending on local context.
METHOD
Participants
Twenty-three introductory psychology students (16 males) participated in this study in partial fulfillment of course credit. One participant was dropped for excessive movement that obscured the ERP data. Analyses are, therefore, based on 22 participants. All self-identified as White.
Materials
Faces
Face primes consisted of 16 pictures each of Black, White and racially ambiguous Black–White male faces. Black and White faces were obtained from yearbook photos, edited to have a uniform blue background and black clothing. The racially ambiguous faces consisted of 16 digitally morphed faces from Willadsen-Jensen and Ito (2006) created using Gryphon Morph 2.5 by mathematically averaging pairs of Black and White faces in a 50–50% proportion. The parent faces used to create the racially ambiguous ones were different from the faces used in the current study and were chosen to be racially prototypical and suitable for the morphing procedure (e.g. no facial hair or jewelry, frontal view). Both racially ambiguous and prototypical faces were pre-tested within a larger set of faces by a separate set of participants (n = 15). Racial ambiguity was assessed in three different ways. Pre-test participants were first asked to make a dichotomous choice between ‘Black’ and ‘White’ for each face. Chosen prototypical Black and White faces were correctly categorized 100% of the time. Morphed faces categorized as Black or White more than 70% of the time were rejected. Participants were then asked to rate the faces on a scale from 1 to 7 (1 = Black, 7 = White). Chosen prototypical Black and White faces had mean ratings of 1 and 7, respectively. Every chosen racially ambiguous face was rated between 3 and 5 by every pre-test participant. Finally, these participants were asked to write down whatever racial category they felt best described each face. Answers were examined to ensure they did not converge on a single answer for the ambiguous faces, such as Latino. Participants were also asked to rate the attractiveness of the faces on a scale from 1 to 7 (1 = Not at all attractive, 7 = Very attractive). Selected faces all had attractiveness Z-scores within ±1; attractiveness did not differ significantly as a function of face type (MBlack = 0.16, MWhites = 0.12, MAmbiguous = 0.16, F(2,47) = 0.11, ns. Finally, to ensure that the morphed faces looked realistic, all pre-test participants were asked if any faces looked unusual or were particularly memorable. Any faces indicated as such were omitted from further consideration.
Target words
Positive and negative nouns were beauty, joy, love, paradise, romance, smile, success, vacation, cockroach, despair, disgust, garbage, pest, poison, sewage, and vomit (Livingston and Brewer, 2002).
Electrophysiological recording
ERPs were recorded from 26 scalp locations using tin electrodes. Miniature tin electrodes were also placed above and below the left eye and next to the outer canthus of each eye to monitor vertical and horizontal eye movements. Scalp electrodes were referenced online to the left mastoid site. All impedances were below 5 KΩ. ERP data were referenced on-line to the left mastoid. Off-line, ERP data were re-referenced to an average of the left and right mastoids and an eye blink algorithm was used to remove the effects of vertical eye movements from the ERP data, which can distort measurements at scalp sites (Semlitsch et al., 1986). Data were epoched into segments beginning 100 ms before face prime onset to 1000 ms post-prime onset. The data were then corrected to the mean voltage of the pre-stimulus recording period. Trials were visually inspected for remaining ocular or other artifact (e.g. due to movement); data for that trial were eliminated from further analysis if an artifact was detected (11% of all trials).2 Finally, data were filtered with a 30-Hz low pass filter then ensemble averages were constructed by aggregating the electrical activity associated with each of the eight Race Cue × Face Ambiguity × Word Valence conditions for each participant at each scalp site. Because the components of interest have been largest at frontal (Fz), central (Cz) and parietal (Pz) midline scalp sites in past research (Ito and Urland, 2003, 2005; Willadsen-Jensen and Ito, 2006; Kubota and Ito, 2007) analyses focused on these scalp sites. Within each participants’ ensemble averages, a positive-going potential between 150 and 250 ms (the P200) and a negative-going potential between 200 and 300 ms (the N200) were scored by locating within each of the condition averages the largest positive-going potential between 150 ms and 250 ms after stimulus onset (P200) and the largest negative-going potential between 200 ms and 350 ms after stimulus onset (N200) (Ito and Urland, 2003). The morphology of the entire waveform at the Fz, Cz, and Pz scalp sites is shown in Figure 1. As can be seen in the figure, there is a negative-going component occurring between 70 and 120 ms after face onset (the N100). This component was scored and analyzed but did not yield any significant results of theoretical interest and so is not discussed further (cf., Willadsen-Jensen and Ito, 2006, 2008).
Fig. 1.

Grand average ERP waveforms to Black-cued and White-cued racially ambiguous and prototypical faces at Fz, Cz and Pz, plus the voltage topographies across all participants and all conditions.
Procedure
In order to examine how categorization into one category vs another affects subsequent perception and evaluative reactions, participants completed the sequential priming task from Fazio et al. (1995) while neural and behavioral data were collected. Participants were presented with Black, White and racially ambiguous Black–White faces as primes. Before each prime, a contextual manipulation was presented in the form of a race label (Black or White) (Eberhardt et al., 2003; Pauker et al., 2009). This resulted in a 2 (Race Cue: Black, White) × 2 (Face Ambiguity: Prototypical, Ambiguous) × 2 (Word Valence: Positive, Negative) within-subject design.
The experimenter first placed an elastic cap into which electrodes were sewn on the participants’ heads (Electro-Cap International, Eaton, OH, USA). Participants were told they would be completing several computer tasks aimed at understanding the automaticity of word comprehension. Participants completed four phases, following Fazio et al. (1995). In the first, participants categorized each target word as good or bad, with all target words presented twice in a random order. This phase was designed to collect baseline, unprimed word categorization latencies. In the next phase, participants viewed four Black and four White faces. The instructions asked participants to attend to and memorize the faces. In the third phase, participants completed a memory task consisting of four previously presented faces (two Black, two White) and four foils (two Black, two White). Participants judged whether or not the face was part of the set presented in the previous phase. Only a subset of previously seen faces was shown because the purpose of this phase was simply to maintain the cover story. All were racially prototypical to avoid sensitizing participants to our interest in racially ambiguous faces. None of the faces used in Phases 2 and 3 were used in subsequent phases.
In the fourth, and crucial, phase participants were told that the experiment would now assess the automaticity of word comprehension by combining prior phases. Participants were told that if word comprehension was automatic, they would react as quickly during this fourth phase, when they were distracted by faces, as they did in the first phase, when there were no faces to distract them. Participants were asked to remember the faces and were told that they would be tested on the faces later.3 They were further told:
In order to help you remember the faces, a racial category will appear before each face. Prior research has shown that attending to racial information can improve memory. Therefore, in order to help you, we will provide the information before each picture.
The race cue appeared on the screen for 315 ms followed by a white screen for 600 ms. The face prime appeared next for 315 ms, followed by the target word 135 ms later, which remained on the screen until participants responded by pressing a button labeled GOOD or BAD (Fazio et al., 1995). The next trial began after a 2500-ms intertrial interval. The stimulus onset asynchrony (SOA) between the race cue and to-be-categorized word was 1365 ms. This relatively long duration was chosen based on previous examinations of SOAs in affective priming (Fazio et al., 1986; Hermans et al., 1994, 2001, 2003; De Houwer et al., 1998, 2001) to minimize any direct effect of the race label on word categorization and make it unlikely that the race label itself produced priming effects on the target words.
The racially ambiguous faces shown in Phase 4 were randomly divided into two sets. For each participant, the racially ambiguous faces in one set were cued by the label Black and the other set as White.4 This varied between subjects, so that the labeling of a particular racially ambiguous face differed across participants.5 The prototypical Black and White faces were always accurately cued as Black and White, respectively. Each face was shown three times with a positive word and three times with a negative word for a total 288 trials.
RESULTS
Analyses focused on three main questions. First, we determined whether the race cue affects racial perception, as reflected in differences in neural responses to Black- and White-cued racially ambiguous faces. Second, we evaluated whether the race cue affects evaluative associations activated by the racially ambiguous faces, as seen in word categorization response latencies. Finally, we examined the relation between the two types of measures to see whether individual differences in the way the faces were processed is predictive of differences in activated evaluative associations. Preliminary analyses showed no effect of participant gender, so that factor was dropped from the reported analyses.
Race perception
P200 and N200 amplitudes were first examined to determine whether cues extrinsic to a target influence early racial perception. Analyses used SAS PROC MIXED to run separate 2 (Race Cue: Black, White) × 2 (Face Ambiguity: Prototypical, Ambiguous) × 3 (Scalp Site: Fz, Cz, Pz) Multilevel Models (MLMs). Because psychophysiological data are naturally clustered within subject, our analyses used multilevel modeling to nest scalp site within subject; we also included a random intercept of subject to model a-theoretical between-subject psychophysiological variance (Raudenbush and Bryk, 2002). This approach has several advantages for analyzing psychophysiological data compared with repeated-measures ANOVA (Vasey and Thayer, 1987; Handy, 2005; Gratton, 2007) including estimation of random subject and item effects simultaneously, separate error terms at each level of nesting, less strict assumptions regarding variance differences between factor levels (sphericity), and increased statistical power (Jeager, 2008). Scalp site did not alter the pattern of effects found at any of the ERP components and so its effects are not reported.
P200 amplitude
P200 amplitude is typically larger to racial outgroup than ingroup faces (Ito and Urland, 2003, 2005; Willadsen-Jensen and Ito, 2006, 2008; Dickter and Barthlow, 2007; Kubota and Ito, 2007). As all participants in the present study were White, we expected larger P200s to Black faces. If extrinsic cues influence the racial perceptions of ambiguous faces, we should also find larger P200s to Black-cued than White-cued ambiguous faces. This was obtained, as shown in the Race Cue main effect; P200s were larger to Black-cued (M = 7.18 µV, SE = 0.68) than White-cued faces (M = 5.77 µV, SE = 0.73), F(1, 217) = 59.33, P < 0.0001, R2 = 0.21.
There were two additional significant effects, but neither of them indicates a lack of racial differentiation for either the prototypical or ambiguous faces. First, the Face Ambiguity main effect showed larger P200s to prototypical (M = 6.78 µV, SE = 0.67) than ambiguous faces (M = 6.18 µV, SE = 0.73), F(1, 217) = 10.54, P < 0.001, R2 = 0.05. There was also an unpredicted interaction between Face Ambiguity and Race Cue, F(1, 217) = 8.70, P = 0.004, R2 = 0.04 (Figure 2). Because it is particularly important to determine whether the race labels changed perceptions of the ambiguous faces, we assessed the simple race effect within each level of face ambiguity. For prototypical faces, P200s were larger to Black (M = 7.76 µV, SE = 0.69) than White faces (M = 5.80 µV, SE = 0.70), F(1, 217) = 56.74, P < 0.0001, R2 = 0.21. Of particular importance, P200s to the racially ambiguous faces showed a similar effect, with larger P200s to Black-cued racially ambiguous (M = 6.62 µV, SE = 0.73) than White-cued racially ambiguous faces (M = 5.74 µV, SE = 0.79), F(1, 217) = 11.30, P < 0.001, R2 = 0.05. Black-cued racially ambiguous faces also elicited larger P200s than prototypical White faces, F(1, 217) = 9.92, P = 0.002, R2 = 0.04). The interaction appears attributable to differences in the Black-cued faces, with larger P200s to the prototypical (Black) when compared with ambiguous Black-cued faces, F(1, 217) = 19.19, P < 0.0001, R2 = 0.08. For the White-cued faces, P200s did not differ between the prototypical (i.e. White) and ambiguous faces, F(1, 217) = 0.04, ns.
Fig. 2.
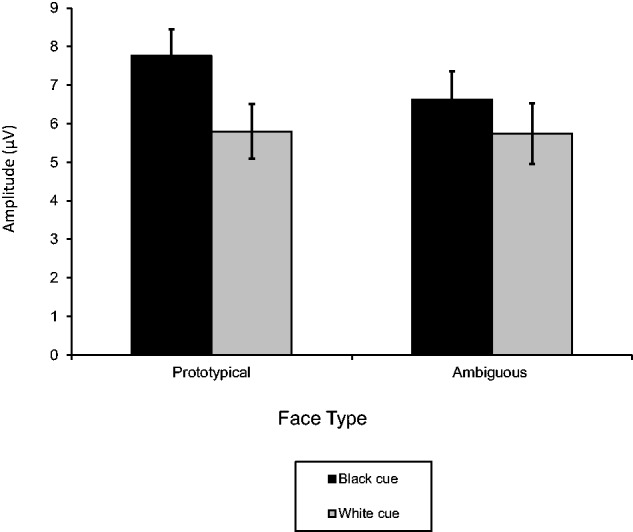
Mean P200 amplitudes to Black-cued and White-cued racially ambiguous and prototypical faces. Errors bars show standard errors.
N200 amplitude
Although P200 amplitude is typically larger to racial outgroup than ingroup faces, N200 amplitude is typically larger to racial ingroup than outgroup faces (Ito and Urland, 2003, 2005; Ito et al., 2004; Willadsen-Jensen and Ito, 2006; Dickter and Bartholow, 2007; Kubota and Ito, 2007). These N200 effects were replicated here and, again, show the effects of extrinsic cues on the reactions to the racially ambiguous faces. The Race Cue main effect was significant, showing larger N200s to White-cued (M = −7.38 µV, SE = 0.64) than Black-cued faces (M = −4.75 µV, SE = 0.64), F(1, 217) = 152.01, P < 0.0001, R2 = 0.41.
As with the P200, there was also a Face Ambiguity main effect, F(1, 217) = 11.38, P < 0.001, R2 = 0.05, showing larger responses to the ambiguous (M = −6.43 µV, SE = 0.68) than prototypical faces (M = −5.71 µV, SE = 0.59). Also significant was the Race Cue × Face Ambiguity interaction (Figure 3), F(1, 217) = 8.12, P = 0.005, R2 = 0.04. We again were particularly interested in whether perceptions of the ambiguous faces were affected by the labels. As expected, N200s were larger to prototypical White (M = −7.33 µV, SE = 0.63) than Black faces (M = −4.09 µV, SE = 0.60), F(1, 217) = 115.19, P < 0.0001, R2 = 0.35. Of importance, this same difference occurred for the racially ambiguous faces. N200s were larger for White-cued racially ambiguous (M = −7.44 µV, SE = 0.66) than Black-cued racially ambiguous faces (M = −5.42 µV, SE = 0.77), F(1, 217) = 44.94, P < 0.0001, R2 = 0.17). Prototypical White faces also elicited larger N200s than the Black-cued racially ambiguous faces, F(1, 217) = 40.07, P < 0.0001, R2 = 0.16. As in the P200, the interaction was due to differences between the Black-cued faces. N200s were bigger (i.e. more negative) to ambiguous Black-cued faces than prototypical (i.e. Black) faces F(1, 217) = 19.36, P < 0.0001, R2 = 0.08. For the White-cued faces, N200s did not differ between the prototypical (i.e. White) and ambiguous faces, F(1, 217) = 0.14, ns.6
Fig. 3.
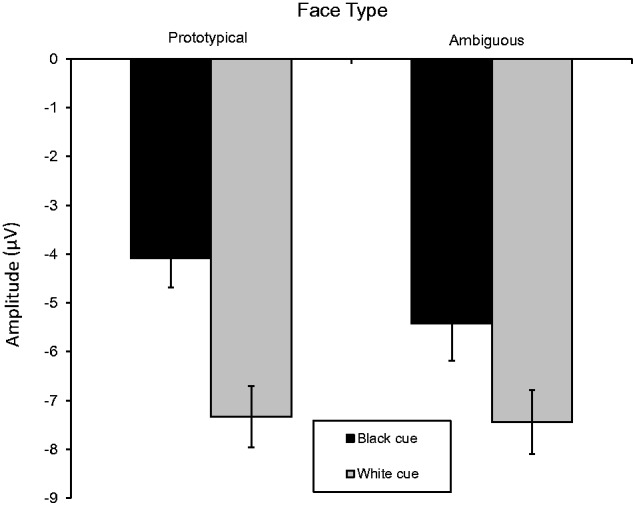
Mean N200 amplitudes to Black-cued and White-cued racially ambiguous and prototypical faces. Errors bars show standard errors.
Response latencies
We next examined cue effects on implicit evaluative associations. Reaction times were scored by first dropping trials on which words were categorized incorrectly (M = 3.58%), then dropping trials that were greater than ±3 standard deviations relative to each participants’ mean reaction time (M = 1.68%). The remaining responses were log transformed. We then computed facilitation scores by subtracting response latencies during the fourth phase of the experiment, when the words were primed with faces, from response latencies in the first phase of the experiment, when the words were unprimed, following Fazio et al. (1995). Positive scores indicate greater facilitation when the words were primed with a given type of face relative to when they were unprimed. All analyses were performed using the log-transformed scores although values are reported in milliseconds for ease of interpretation.
A 2 (Race Cue: Black, White) × 2 (Face Ambiguity: Prototypical, Ambiguous) × 2 (Word Valence: Positive, Negative) within subjects ANOVA revealed a main effect of Race Cue, F(1, 21) = 11.40, P = 0.003, = 0.35. Participants showed greater facilitation after White-cued (M = 61.34 ms, SE = 12.48) than Black-cued faces (M = 49.83 ms, SE = 11.94). The main effect of Face Ambiguity was also significant, F(1, 21) = 5.13, P = 0.034, = 0.20, with greater facilitation following prototypical (M = 61.01 ms, SE = 12.73) than ambiguous faces (M = 50.15 ms, SE = 11.91).
Of more theoretical interest, the predicted interaction of Race Cue and Word Valence was significant, with evaluative associations following the extrinsic cue, F(1, 21) = 11.11, P = 0.003, = 0.35. As shown in Figure 4, Black-cued face primes resulted in significantly greater facilitation for negative (M = 63.23 ms, SE = 14.19) than positive words (M = 36.42 ms, SE = 12.78), F(1, 21) = 4.58, P = 0.044, = 0.18. Directionally, White-cued faces facilitated faster responses to positive (M = 64.96 ms, SE = 12.90) than negative words (M = 57.71 ms, SE = 14.27), but this difference was not significant, F(1, 21) = 0.53, ns. The stronger priming effect following Black than White primes is consistent with past research (e.g. Payne, 2001; Ito et al., 2011) and may indicate a stronger difference between negative and positive associations for outgroup than ingroup members. This effect was not moderated by face ambiguity.7
Fig. 4.
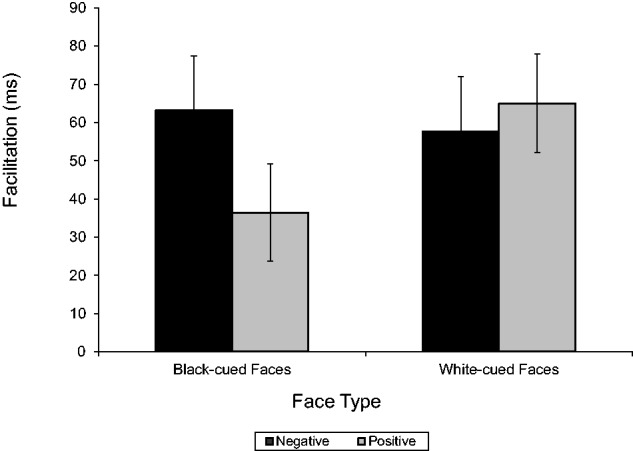
Facilitation scores for positive and negative words primed by Black-cued and White-cued faces. Errors bars show standard errors.
Relation between neural responses and response latencies
Previous research shows that greater neural differentiation between prototypical Black and White faces is positively correlated with implicit evaluative bias; the more participants differentiate Black and White faces as shown in P200 and N200 amplitudes, the greater the implicit evaluative bias against Blacks (Correll et al., 2006; see also He et al., 2009). We expected to replicate that finding in the present study, while also extending it to the racially ambiguous faces. If differences in attention to Black and White faces are associated with greater evaluative bias, and if racially ambiguous faces are perceived as Black or White in accordance with a racial label, then differences in how perceivers attend to faces should also predict the evaluative bias the faces elicit.
This hypothesis was evaluated by computing contrast scores for both the ERP and response latency data. Separate scores were computed for the P200 and N200 reflecting the difference in responses to Black-cued (i.e. both the racially prototypical Black and racially ambiguous Black-cued faces) vs White-cued faces. The contrast score for response latency reflected the degree of racial bias, as shown in relatively greater facilitation to negative than positive words following Black-cued primes but relatively greater facilitation to positive than negative words following White-cued primes.8
As predicted, individuals who showed relatively larger P200s to Black-cued faces also showed relatively more negative implicit associations following Black-cued than White-cued primes, r(20) = 0.52, P = 0.013. Similarly, individuals who showed relatively larger N200s to White-cued than Black-cued faces also showed relatively more negative implicit associations following Black-cued than White-cued primes, r(20) = 0.55, P = 0.008.9
DISCUSSION
Our results demonstrate the profound effect contextual information can have on reactions to faces that possess features associated with two racial groups. When external cues lead a perceiver to view a racially ambiguous individual as Black, more negative associations are activated. If that same individual is instead perceived as White, relatively more positive associations are activated. Of importance, this study demonstrates that the contextual information changes on-line racial perceptions within the first 200 ms of perception, which in turn underlie the behavioral responses. These findings are consistent with other studies showing contextual influences on the perceptions of racially ambiguous faces (Eberhardt et al., 2003; Peery and Bodenhausen, 2008; Pauker et al., 2009; Ito et al., 2011; Dickter and Kittel, 2012). The present data, however, are the first to our knowledge to directly link a contextual cue to a change in the racial perception of the face, and then link that change in perception to a change in implicit evaluative bias.
In addition to elucidating mechanisms that affect reactions to multiracial and other individuals who possess features associated with more than one racial group, the present results speak to the dynamic relation between social categorization and judgment. Models of person perception hypothesize a direct link between categorization and the activation of group-based associations (e.g. Brewer, 1988; Fiske and Neuberg, 1990; Macrae and Bodenhausen, 2000). In this view, changing the way in which an individual is categorized should change the activated associations, exactly the pattern obtained here.
Although we obtained the predicted Race Cue main effects in both the P200 and N200 analyses, we also obtained unexpected Race Cue × Face Ambiguity interactions in both analyses. Of primary importance, simple effects tests within each component revealed significant differences in responses to Black-cued and White-cued racially ambiguous faces. This provides clear evidence that the way in which racially ambiguous individuals are perceived can be quickly moderated by contextual information. In both components, the interaction was attributable to differences in responses to the Black-cued faces, with more positive P200s and less negative N200s to prototypical Black than Black-cued ambiguous faces. Together, the results indicate that waveforms were relatively more positive to prototypical Black than Black-cued ambiguous faces. Such a pattern suggests that while Black-cued racially ambiguous faces are clearly differentiated from both prototypical White faces and White-cued racially ambiguous faces, they do not elicit responses as extreme as prototypical Black faces.
This pattern may reflect differences in perceptual expertise in encoding ingroup and outgroup members that paradoxically decreases distinctions between racially ambiguous faces and the ingroup when compared with the outgroup. Our pattern of results may specifically be consistent with Valentine and Endo’s (1992) multidimensional space model in which faces are argued to be encoded along dimensions acquired through experience to best differentiate among the faces an individual most frequently encounters. Because perceivers often have more experience interacting with racial ingroup members, the dimensions representing the face space will typically be tuned to differentiate among ingroup faces. The model further assumes that individual ingroup faces are clustered around the intersection of the n-dimensions, but because ingroup faces are well-differentiated by the dimensions used to encoded them, they are relatively loosely clustered around this point of origin. By contrast, outgroup faces are assumed to be clustered distant from the point of origin of the dimensions, and because they are encoded with respect to dimensions that were derived from experience with faces of a different race, they tend to be more densely clustered together than ingroup faces (i.e. the dimensions used to encode them are not well-suited for differentiating among them).
Since the racially ambiguous faces we used here contained a blend of ingroup and outgroup features, they are likely to be represented in the face space at a location equidistant from the center of the ingroup and outgroup face clusters. However, because ingroup faces are more loosely dispersed around their center, racially ambiguous faces are likely to be closer to individual ingroup than outgroup faces. This would produce greater perceptual overlap between the racially ambiguous faces and ingroup than outgroup faces, at least at very early stages of perception. In this way, racially ambiguous faces may be initially perceptually pulled toward the ingroup (cf. Willadsen-Jensen and Ito, 2006). This pattern has in fact been observed before with racially ambiguous faces viewed without preceding race cues. In these studies, White participants’ P200s and N200 to racially ambiguous White–Black and White-Asian faces were indistinguishable to responses to prototypical White faces but were significantly different from responses to prototypical Black and Asian faces (Willadsen-Jensen and Ito, 2006).
The difference in P200 and N200 responses to prototypical Black and racially ambiguous Black-cued faces is also interesting to consider from the perspective of ingroup overexclusion, which argues that social identity concerns motivate individuals to maintain positive ingroup distinctiveness (Leyens and Yzerbyt, 1992). This can translate into being particularly conservative in accepting people as members of the ingroup (Blascovich et al., 1997; Castano et al., 2002; Peery and Bodenhausen, 2008; Dickter and Kittel, 2012). Our effects were the opposite, with seemingly less differentiation of racially ambiguous individuals from ingroup members. The resolution may lie in the type of response being assessed. Studies finding evidence of ingroup overexclusion have typically measured processes that occur subsequent to initial perception (e.g. explicit racial categorization) whereas our ERP results seem to capture the earlier perceptual processes. Our pattern of results in conjunction with ingroup overexclusion effects may indicate that even in the face of early perceptual processes that facilitate assimilation of racially ambiguous individual to the ingroup (as reflected here in the ERP responses), motivational forces subsequently override this tendency, and in fact can push racially ambiguous faces away from the ingroup. Consistent with this, the evaluative responses we obtained, which occurred subsequent to the initial perceptual responses, reflected in the ERPs, showed no differentiation between prototypical Black and Black-cued racially ambiguous faces.
Although these data demonstrate the effects of extrinsic cues on perceptions and evaluations, we do not think external factors are the sole determinant of reactions to multiracial individuals. The growing body of research on perceptions of and reactions to racially ambiguous faces supports the role of several other factors including features of the target individual (MacLin and Malpass, 2001; Hugenberg and Bodenhausen, 2004; Ito et al., 2011), motivations to protect the ingroup (Blascovich et al., 1997; Castano et al., 2002; Pauker et al., 2009), differential experience with racial groups (Halberstadt et al., 2011), and historical considerations (Banks and Eberhardt, 1998; Peery and Bodenhausen, 2008). Perceptions of multiracial individuals are clearly a multifaceted process. Nevertheless, the present results highlight the importance of considering how greatly reactions to the same individual can vary based on simple external cues.
Conflict of Interest
None declared.
Acknowledgments
The authors are grateful to dissertation committee members Irene Blair, Bernadette Park, Lewis O. Harvey, Jr, and Hillary Potter, to Christine Bennett and Nathan Hand for assistance with data collection, and to Keith Senholzi and Sarah Grover for assistance with analyses. This work was supported by National Institute of Mental Health Grant R01MH071257 to Tiffany Ito and was completed as part of Eve Willadsen-Jensen’s dissertation.
Footnotes
1 In Eberhardt et al. (2003), labeling racially ambiguous faces as ingroup members lead to assimilation to the ingroup only among participants who possessed a more entity view in which traits are perceived as immutable, essential properties of an individual. The pattern actually reversed for individuals who held a more incremental view in which traits are perceived as more malleable and possessing less predictive validity, resulting in a contrast away from the ingroup for these participants.
2 Criteria for rejection included horizontal eye movement, amplitudes greater than ±100 µV, or remaining eyeblink artifact.
3 A memory test did follow the critical priming block in order to maintain the cover story. Due to a programming error, unequal numbers of faces of different types were shown across participants, leaving us unable to assess race effects on face memory.
4 We had intended each participant to see eight racially ambiguous faces cued as Black and eight racially ambiguous faces cued as White. However, due to a programming error, 7 participants saw seven Black-cued racially ambiguous and nine White-cued racially ambiguous faces whereas the other 12 participants saw nine Black-cued racially ambiguous and seven White-cued racially ambiguous faces.
5 There was no effect of which subset of faces was labeled as Black or White, so that factor was dropped from further analyses.
6 Given the inverse relation between the P200 and N200 race effects, we conducted two additional analyses to determine that the more negative N200 to Whites was independent from the less positive P200s to Whites. In the first, we quantified N200 amplitude as the peak-to-peak value relative to the amplitude of the preceding P200 peak. If larger N200s to Whites were simply a function of there being less positive responses to Whites in the preceding P200, then the race effect in the N200 should be eliminated in the peak-to-peak measure. By contrast, the Race Cue main effect remained in this analysis, indicating that N200s are larger to White-cued faces, even when racial differences in the P200 are considered, F(1, 217) = 25.41, P < 0.001, R2 = 0.11 (a Site main effect was the only other significant effect in this analysis). Similarly, when the P200 amplitude was included as a covariate in the MLM analysis of N200 amplitude, the pattern of results was unaffected. The N200 Race Cue main effect remained significant (F(1, 234) = 92.74, P < 0.0001, R2 = 0.28) as did the main effect of Face Ambiguity (F(1, 220) = 6.35, P = 0.013, R2 = 0.03), and the interaction of Race Cue × Face Ambiguity (F(1, 220) = 4.32, P = 0.04, R2 = 0.02). Based on these ancillary analyses, we are confident that the greater negativity to Whites observed in the N200 is independent of voltages in the prior P200 time range.
7 Although none of the interactions involving face ambiguity were significant (all Ps > 0.15), we separately examined facilitation following just the racially ambiguous faces to verify that they specifically elicited differential responses as a function of Race Cue and Word Valence. Mirroring the pattern reported in the main text, racially ambiguous primes cued as Black lead to significantly greater facilitation of negative (M = 62.92 ms, SE = 14.73) than positive words (M = 22.97 ms, SE = 14.34), F(1, 21) = 4.88, P = 0.039, = 0.19. For racially ambiguous primes cued as White, there was no significant difference in facilitation of negative (M = 52.03 ms, SE = 14.56) vs positive (M = 62.68 ms, SE = 14.73) words, F(1,21) = 0.55, ns.
8 The response latency contrast score was computed as (facilitation of negative words primed by Black-cued, Black faces − facilitation of positive words primed by Black-cued, Black faces) + (facilitation of negative words primed by Black-cued, ambiguous faces − facilitation of positive words primed by Black-cued, ambiguous faces) + (facilitation of positive words primed by White-cued, White faces − facilitation of negative words primed by White-cued, White faces) + (facilitation of positive word primed by White-cued, ambiguous faces − facilitation of negative word primed by White-cued, ambiguous faces).
9 We examined the correlations separately for the racially ambiguous faces to verify that the relation between racial perceptions, as reflected in the ERPs, and implicit evaluations was obtained for these faces in particular. Replicating the main results, racial differences in both the P200 (r(20) = 0.50, P = 0.018) and N200 (r(20) = 0.40, P = 0.065) predicted implicit racial bias, with the N200 relationship being marginally significant.
REFERENCES
- Banks R, Eberhardt JL. Social psychological processes and the legal bases of racial categorization. In: Eberhardt JL, Fiske ST, editors. Confronting Racism: The Problem and the Response. CA: Sage Publications; 1998. pp. 54–75. [Google Scholar]
- Barden J, Maddux WW, Petty RE, Brewer MB. Contextual moderation of racial bias: the impact of social roles on controlled and automatically activated attitudes. Journal of Personality and Social Psychology. 2004;87:5–22. doi: 10.1037/0022-3514.87.1.5. [DOI] [PubMed] [Google Scholar]
- Blascovich J, Wyer NA, Swart LA, Kibler JL. Racism and racial categorization. Journal of Personality and Social Psychology. 1997;72:1364–72. [Google Scholar]
- Brewer MB. A dual process model of impression formation. Advances in Social Cognition. 1988;1:1–36. [Google Scholar]
- Castano E, Yzerbyt V, Bourguignon D, Seron E. Who may enter? The impact of in-group identification on in-group/out-group categorization. Journal of Experimental Social Psychology. 2002;38:315–322. [Google Scholar]
- Correll J, Urland GR, Ito TA. Event-related potentials and the decision to shoot: the role of threat perception and cognitive control. Journal of Experimental Social Psychology. 2006;42:120–8. [Google Scholar]
- De Houwer J, Hermans D, Eelen P. Affective and identity priming with episodically associated stimuli. Cognition and Emotion. 1998;12:145–169. [Google Scholar]
- De Houwer J, Hermans D, Spruyt A. Affective priming of pronunciation responses: effects of target-degradation. Journal of Experimental Social Psychology. 2001;37:85–91. [Google Scholar]
- Dickter CL, Bartholow BD. Racial ingroup and outgroup attention biases revealed by event-related brain potentials. Social Cognitive and Affective Neuroscience. 2007;2:189–98. doi: 10.1093/scan/nsm012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickter CL, Kittel JA. The effect of stereotypical primes on the neural processing of racially ambiguous faces. Social Neuroscience. 2012;7(6):622–31. doi: 10.1080/17470919.2012.690345. [DOI] [PubMed] [Google Scholar]
- Eberhardt JL, Dasgupta N, Banaszynski TL. Believing is seeing: the effects of racial labels and implicit beliefs on face perception. Personality and Social Psychology Bulletin. 2003;29:360–70. doi: 10.1177/0146167202250215. [DOI] [PubMed] [Google Scholar]
- Fazio RH, Jackson JR, Dunton BC, Williams CJ. Variability in automatic activation as an unobtrusive measure of racial attitudes: a bona fide pipeline? Journal of Personality and Social Psychology. 1995;69:1013–27. doi: 10.1037//0022-3514.69.6.1013. [DOI] [PubMed] [Google Scholar]
- Fazio RH, Sanbonmatsu DM, Powell MC, Kardes FR. On the automatic activation of attitudes. Journal of Personality and Social Psychology. 1986;50:229–38. doi: 10.1037//0022-3514.50.2.229. [DOI] [PubMed] [Google Scholar]
- Fiske ST, Neuberg SL. A continuum of impression formation, from category-based to individuating processes: influences of information and motivation on attention and interpretation. In: Zanna MP, editor. Advances in Experimental Social Psychology. Vol. 23. New York: Academic Press; 1990. pp. 1–74. [Google Scholar]
- Gratton G. Biosignal processing. In: Cacioppo J, Tassinary L, Berntson G, editors. Handbook of Psychophysiology. New York, NY: Cambridge University Press; 2007. pp. 900–23. [Google Scholar]
- Halberstadt J, Sherman S, Sherman J. Why Barack Obama is black: a cognitive account of hypodescent. Psychological Science. 2011;22:29−33. doi: 10.1177/0956797610390383. [DOI] [PubMed] [Google Scholar]
- Handy TC. Event-Related Potentials: A Methods Handbook. Cambridge: MIT press; 2005. [Google Scholar]
- He Y, Johnson MK, Dovidio JF, McCarthy G. The relation between race-related implicit associations and scalp-recorded neural activity evoked by faces from different races. Social Neuroscience. 2009;4(5):426–42. doi: 10.1080/17470910902949184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermans D, De Houwer J, Eelen P. The affective priming effect: automatic activation of evaluative information in memory. Cognition and Emotion. 1994;8:515–33. [Google Scholar]
- Hermans D, De Houwer J, Eelen P. A time course analysis of the affective priming effect. Cognition and Emotion. 2001;15:143–65. [Google Scholar]
- Hermans D, Spruyt A, Eelen P. Automatic affective priming of recently acquired stimulus valence: priming at SOA 300 but not at SOA 1000. Cognition and Emotion. 2003;17:83–99. doi: 10.1080/02699930302276. [DOI] [PubMed] [Google Scholar]
- Hugenberg K, Bodenhausen GV. Ambiguity in social categorization: the role of prejudice and facial affect in racial categorization. Psychological Science. 2004;15:342–5. doi: 10.1111/j.0956-7976.2004.00680.x. [DOI] [PubMed] [Google Scholar]
- Hugenberg K, Young SG, Bernstein KJ, Sacco DF. The categorization-individuation model: an integrative account of the other-race recognition deficit. Psychological Review. 2010;117:1168–1187. doi: 10.1037/a0020463. [DOI] [PubMed] [Google Scholar]
- Ito TA, Bartholow BD. The neural correlates of race. Trends in Cognitive Sciences. 2009;13:524–31. doi: 10.1016/j.tics.2009.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito TA, Thompson E, Cacioppo JT. Tracking the timecourse of social perception: the effects of racial cues on event-related brain potentials. Personality and Social Psychology Bulletin. 2004;30:1267–80. doi: 10.1177/0146167204264335. [DOI] [PubMed] [Google Scholar]
- Ito TA, Urland GR. Race and gender on the brain: electrocortical measures of attention to race and gender of multiply categorizable individuals. Journal of Personality and Social Psychology. 2003;85:616–26. doi: 10.1037/0022-3514.85.4.616. [DOI] [PubMed] [Google Scholar]
- Ito TA, Urland GR. The influence of processing objectives on the perception of faces: an ERP study of race and gender perception. Cognitive, Affective, and Behavioral Neuroscience. 2005;5:21–36. doi: 10.3758/cabn.5.1.21. [DOI] [PubMed] [Google Scholar]
- Ito TA, Willadsen-Jensen EC, Kaye J, Park B. Contextual variation in automatic evaluative bias to racially ambiguous faces. Journal of Experimental Social Psychology. 2011;47:818–23. doi: 10.1016/j.jesp.2011.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeager TF. Categorical data analysis: away from ANOVAs (transformation or not) and towards logit mixed models. Journal of Memory and Language. 2008;59:434–46. doi: 10.1016/j.jml.2007.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubota JT, Ito TA. Multiple cues in social perception: the time course of processing race and facial expression. Journal of Experimental Social Psychology. 2007;43:738–52. doi: 10.1016/j.jesp.2006.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin DT. Classifying faces by race: the structure of face categories. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1996;22:1364–82. [Google Scholar]
- Levin D. Race as a visual feature: using visual search and perceptual discrimination tasks to understand face categories and the cross-race recognition deficit. Journal of Experimental Psychology: General. 2000;129:559–74. doi: 10.1037//0096-3445.129.4.559. [DOI] [PubMed] [Google Scholar]
- Leyens JP, Yzerbyt VY. The ingroup overexclusion effect: impact of valence and confirmation on stereotypical information search. European Journal of Social Psychology. 1992;22:549–69. [Google Scholar]
- Levin DT, Banaji MR. Distortions in the perceived lightness of faces: the role of race categories. Journal of Experimental Psychology: General. 2006;135:501–12. doi: 10.1037/0096-3445.135.4.501. [DOI] [PubMed] [Google Scholar]
- Livingston RW, Brewer MB. What are we really priming? Cue-based versus category-based processing of facial stimuli. Journal of Personality and Social Psychology. 2002;82:5–18. doi: 10.1037//0022-3514.82.1.5. [DOI] [PubMed] [Google Scholar]
- Maclin OH, Malpass RS. Racial categorization of faces: the ambiguous-race face effect. Psychology, Public Policy, and Law. 2001;7:98–118. [Google Scholar]
- Macrae CN, Bodenhausen GV. Social cognition: thinking categorically about others. Annual Review of Psychology. 2000;51:93–120. doi: 10.1146/annurev.psych.51.1.93. [DOI] [PubMed] [Google Scholar]
- Malpass R, Kravitz J. Recognition for faces of own and other race. Journal of Personality and Social Psychology. 1969;13:330–4. doi: 10.1037/h0028434. [DOI] [PubMed] [Google Scholar]
- Michel C, Corneille O, Rossion B. Race categorization modulates holistic face encoding. Cognitive Science. 2007;31:911–24. doi: 10.1080/03640210701530805. [DOI] [PubMed] [Google Scholar]
- Michel C, Corneille O, Rossion B. Holistic face encoding is modulated by perceived face race: evidence from perceptual adaptation. Visual Cognition. 2010;18:434–55. [Google Scholar]
- Payne KB. Prejudice and perception: the role of automatic and controlled processes in misperceiving a weapon. Journal of Personality and Social Psychology. 2001;81:181–92. doi: 10.1037//0022-3514.81.2.181. [DOI] [PubMed] [Google Scholar]
- Pauker K, Weisbuch M, Ambady N, Sommers SR, Adams RB, Jr, Ivcevic Z. Not so black and white: memory for ambiguous group members. Journal of Personality and Social Psychology. 2009;96:795–810. doi: 10.1037/a0013265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peery D, Bodenhausen GV. Black + White = Black: hypodescent in reflexive categorization of racially ambiguous faces. Psychological Science. 2008;19:973–77. doi: 10.1111/j.1467-9280.2008.02185.x. [DOI] [PubMed] [Google Scholar]
- Raudenbush SW, Bryk AS. Hierarchical Linear Models. 2nd edn. Thousand Oaks, CA: Sage; 2002. [Google Scholar]
- Rhodes G, Brake S, Taylor K, Tan S. Expertise and configural coding in face recognition. British Journal of Psychology. 1989;80:313–31. doi: 10.1111/j.2044-8295.1989.tb02323.x. [DOI] [PubMed] [Google Scholar]
- Rhodes G, Lie HC, Ewing L, Evangelista E, Tanaka JW. Does perceived race affect discrimination and recognition of ambiguous-race faces? A test of the sociocognitive hypothesis. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2010;36:217–23. doi: 10.1037/a0017680. [DOI] [PubMed] [Google Scholar]
- Semlitsch HV, Anderer P, Schuster P, Presslich P. A solution for reliable and valid reduction of ocular artifacts, applied to the P300 ERP. Psychophysiology. 1986;23:695–703. doi: 10.1111/j.1469-8986.1986.tb00696.x. [DOI] [PubMed] [Google Scholar]
- Tanaka JW, Kiefer M, Bukach CM. A holistic account of the own-race effect in face recognition: evidence from a cross-cultural study. Cognition. 2004;93:B1–B9. doi: 10.1016/j.cognition.2003.09.011. [DOI] [PubMed] [Google Scholar]
- Valentine T. A unified acount of the effects of distinctiveness, invasion, and race in face recognition. The Quaterly Journal of Experimental Psychology. 1991;43:161–204. doi: 10.1080/14640749108400966. [DOI] [PubMed] [Google Scholar]
- Valentine T, Endo M. Towards an exemplar model of face processing: the effects of race and distinctiveness. The Quarterly Journal of Experimental Psychology. 1992;44:671–703. doi: 10.1080/14640749208401305. [DOI] [PubMed] [Google Scholar]
- Vasey MW, Thayer JF. The continuing problem of false positives in repeated measures ANOVA in psychophysiology: a multivariate solution. Psychophysiology. 1987;24:479–86. doi: 10.1111/j.1469-8986.1987.tb00324.x. [DOI] [PubMed] [Google Scholar]
- Willadsen-Jensen EC, Ito TA. Ambiguity and the timecourse of racial perception. Social Cognition. 2006;24:580–606. [Google Scholar]
- Willadsen-Jensen EC, Ito TA. A foot in both worlds: Asian Americans’ perceptions of Asian, White, and racially ambiguous faces. Group Processes and Interpersonal Relations. 2008;11:182–200. [Google Scholar]
- Wittenbrink B, Judd CM, Park B. Spontaneous prejudice in context: variability in automatically activated attitudes. Journal of Personality and Social Psychology. 2001;81:815–27. doi: 10.1037//0022-3514.81.5.815. [DOI] [PubMed] [Google Scholar]