

Abstract
This study examines the neural correlates of processing verbal entries with multiple argument structures using fMRI. We compared brain activation in response to ‘alternating transitivity’ verbs, corresponding to two different verbal alternates – one transitive and one intransitive - and simple verbs, with only one, intransitive, thematic grid. Fourteen young healthy participants performed a lexical decision task with the two verb types. Results showed significantly greater activation in the angular and supramarginal gyri (BAs 39 and 40) extending to the posterior superior and middle temporal gyri bilaterally, for alternating compared to simple verbs. Additional activation was detected in bilateral middle and superior frontal gyri (BAs 8 and 9). The opposite contrast, simple compared to alternating verbs, showed no significant differential activation in any regions of the brain. These findings are consistent with previous studies implicating a posterior network including the superior temporal, supramarginal and angular gyri for processing verbs with multiple thematic roles, as well as with those suggesting involvement of the middle and superior frontal gyri in lexical ambiguity processing. However, because ‘alternating transitivity’ verbs differ from simple intransitives with regard to both the number of thematic grids (two vs. one) and the number of thematic roles (two vs. one), our findings do not distinguish between activations associated with these two differences.
Introduction
A central part of knowing a language is knowing the argument structure of its predicates. The argument structure of a verb forms the interface between the conceptual/semantic properties of the event denoted by the verb (e.g. how many participants the event includes) and its syntactic properties (e.g. how many noun phrases accompany the verb in a sentence). Accordingly, verbs differ with regard to the number of arguments they select, their type, and the way they are mapped onto syntactic positions. Some verbs have extremely simple argument structures, whereas others have more complex ones.
Argument structure complexity can be instantiated in different ways. One of these is the number of thematic (θ) roles a verb has, i.e. verbs with fewer thematic roles (e.g. ‘sneeze’, an intransitive verb requiring only an agent) (1) are less complex than verbs with a greater number of thematic roles (e.g. ‘fix’, a transitive verb requiring an agent and a theme) (2).
- John sneezed.
- SNEEZE: θAGENT
- John fixed the car.
- FIX: θAGENT θTHEME
Another criterion for complexity is the number of different syntactic realization options of the thematic roles encoded within the verb's representation. For example, the verb ‘rely’ has only one syntactic realization option – it must take a PP complement (3). In contrast, ‘believe’ can appear with three different types of complements – an NP, a PP, or a clause (4).
- 3.
- John relied on Mary.
- *John relied Mary.
- *John relied that Mary liked him.
- 4.
- John believed Mary.
- John believed in Mary.
- John believed that Mary liked him.
The representations in (1)-(2) above assume that thematic role information is listed as part of the verbal entry in the mental lexicon. This is the traditional view of the lexicon-syntax interface, to which we will refer, following Ramchand (2008), as the lexical-thematic approach. According to this view (Chomsky, 1981; Reinhart, 2002; among many others) the lexicon includes, for each verb, its thematic grid (or different possible thematic grids). This information is projected from the lexicon when a sentence is constructed, and the thematic information of the verb determines the number of noun phrases that appear in the sentence, e.g. a transitive verb will necessitate two noun phrases.
In recent years, an opposing approach, the generative-constructivist approach, has emerged. Under this view, the lexicon does not include any grammatically relevant information, namely, it does not specify thematic grids. Thus, syntactic structures are not built based on the semantic/thematic information associated with verbs. Rather, syntactic building is free, restricted only by world knowledge (Borer, 2005).
In the last decades, many studies have established that argument structure complexity has neuropsychological consequences. Research across different languages, including Dutch, English, German, Italian, and Russian has shown a hierarchy of deficits in agrammatic aphasic verb production based on number of thematic roles, with 1-argument verbs presenting less difficulty than 2- or 3-argument verbs (Bastiaanse & Jonkers, 1998; De Bleser & Kauschke, 2003; Dragoy & Bastiaanse, 2010; Kim & Thompson, 2000, 2004; Luzzatti et al., 2002; Thompson, Lange, Schneider & Shapiro, 1997; Thompson, Shapiro, Li & Schendel, 1995).
Imaging studies provide further support for the mapping of argument structure complexity onto the brain. Thompson et al. (2007) and Thompson, Bonakdarpour & Fix (2010) examined processing of one-, two- and three-argument verbs using a lexical decision task, and found that greater complexity in terms of the number of thematic roles was associated with increased neural activation in the angular and supramarginal gyri (Brodmann areas 39 and 40) bilaterally. Similarly, Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky (2003) found activation in the left posterior superior temporal sulcus when comparing the processing of sentences with 3- and 2-argument verbs.1 These posterior temporal and inferior parietal activations in response to argument structure complexity are consistent with research indicating involvement of these regions in complex semantic processing (for a review, see Binder, Desai, Graves, & Conant, 2009). Examining verb processing in sentence contexts, Shetreet, Palti, Friedmann, & Hadar (2007) also found that the medial precuneus and the anterior cingulate, not considered to be ‘traditional’ language regions, were sensitive to the number of thematic roles a verb encodes. Finally, in a study examining naming of 2-argument compared to 1-argument verbs, Den Ouden, Fix, Parrish & Thompson (2009) found increased activation bilaterally in the superior parietal lobule (extending in the left hemisphere to the supramarginal gyrus) and middle temporal gyrus, as well as left hemisphere frontal regions. The authors attributed the frontal activation to the actual production process, whereas the posterior activation was considered to be associated with the processing of verbs with a greater number of thematic roles.
Complexity with respect to the number of syntactic realization options (as exemplified in (3)-(4) above) also has been shown to have neuropsychological consequences. In a cross-modal lexical decision experiment, Shapiro, Zurif, & Grimshaw (1987) found that healthy participants display longer reaction times when processing verbs with multiple syntactic realization options compared with simple verbs. In more recent fMRI studies with unimpaired listeners, Shetreet et al. (2007) and Shetreet, Friedmann, & Hadar (2010a) found increased neural activation in the left superior temporal gyrus, as well as in the mid-superior and inferior frontal gyri (BAs 9 and 47), in response to an increase in the number of the verb's syntactic realization options.
The present study examines the neural representation of yet another instantiation of argument structure complexity, not explicitly targeted in previous imaging studies: the argument structure ambiguity associated with verbs with alternating transitivity. In English, a large group of lexical verb entries, labeled ‘alternating transitivity’ verbs (Levin, 1993), correspond to two different verbal alternates: intransitive and transitive. These verbs present a basic semantic/conceptual ambiguity with regard to the type of event they denote, as they can denote both an event with one participant, and an event with two participants. Accordingly, as exemplified in (5)-(6), the same phonetic string, i.e. ‘opened’ or ‘rolled’, can be used as an intransitive verb taking only one NP argument (5a), (6a), or as a transitive verb, selecting for two NP arguments (5b), (6b).
- 5.
- The door opened.
- The teacher opened the door.
- 6.
- The dog rolled (off the bed).
- The children rolled the dog (off the bed).
The same alternation is found with verbs such as close, sink, break and many others. In all cases, one lexical entry is associated with two different types of events, and two thematic grids. That these verbs correspond to two different verbal alternates can be seen clearly in languages that have richer morphology than English. In such languages, e.g. Hebrew, Polish or Japanese, the transitive and intransitive alternates of these verbs are very often morphologically distinct (though naturally related).
The intransitive alternate of alternating transitivity verbs (5a), (6a) constitutes an unaccusative verb (Perlmutter 1978), selecting for a single (theme) argument, which in sentences undergoes syntactic NP-movement from object to subject position (see e.g. Burzio, 1986).2 In the generative linguistic literature, the alternation in (5)-(6) is thus referred to as the transitive-unaccusative (sometimes also the causative-inchoative) alternation.
In contrast to alternating transitivity verbs, other verbs in English do not display the alternation exemplified in (5)-(6). These verbs are exclusively intransitive (7a), rejecting a transitive use, as shown by the ungrammaticality of (7b).
- 7.
- The dog barked.
- *The vet barked the dog. (Intended meaning: The vet made the dog bark.)
According to the lexical-thematic approach, the difference between alternating transitivity verbs, e.g. break, and simple intransitive verbs, e.g. bark, is rooted in the different lexical information associated with them. While break is associated with two event types, or two thematic grids (transitive and intransitive), bark is associated with only one. Proponents of this approach can be further divided into two: 1) The transitive and intransitive alternates are related by a lexical operation reducing the transitive alternate's external thematic role to derive the unaccusative alternate (see e.g. Chierchia, 2004; Horvath & Siloni, 2011; Levin & Rappaport Hovav, 1995). Both verbal alternates are thus listed in the lexicon, with their respective thematic grids; 2) The lexicon consists solely of abstract roots, carrying some thematic information, and not of verbs. Actual verbs are built in the syntax, by merging the root with functional elements such as a verbal head, a causative head, etc. In the case of the transitive-unaccusative alternation, the root, e.g. break, encodes information such that it can give rise to both transitive and unaccusative verbs; the root bark encodes information such that it can only become an intransitive verb. This information guides the syntactic component: for the root break, both a transitive and an unaccusative verb can be syntactically constructed, whereas for the root bark, only an intransitive verb will be built (see e.g. Embick, 2004; Marantz, 1997; Ramchand, 2008).
In contrast to the lexical-thematic approach, under the generative-constructivist view, the verbal entry is devoid of any thematic information; the difference in verbal realization between e.g. break and bark is attributed solely to the syntactic structure in which they appear and to general interpretational principles (see e.g. Borer, 2004). According to this view, every verbal entry has the basic capacity to realize any argument structure (for further discussion of the different theoretical accounts of the alternation, see Horvath & Siloni, 2011; Schäfer, 2009).
The thematic properties of ‘alternating transitivity’ and simple intransitive verbs can be schematized as in (8).
- 8.
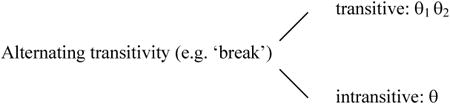
- Simple intransitives (e.g. ‘bark’) ------ intransitive: θ
As can be seen in (8), the two verb types differ on two dimensions: first, alternating transitivity verbs have two different thematic grids, whereas simple intransitive verbs have only one; second, on the transitive use of alternating transitivity verbs, they also differ from simple intransitives in the number of thematic roles they possess: two vs. one.
The present study examined the neural correlates of processing alternating transitivity and simple intransitive verbs.3 The verbs were presented in isolation, such that the sentential context would not restrict their argument structure.4 Based on our previous studies, investigating the effect of number of thematic roles of verbs presented in isolation in a lexical decision task (Thompson et al., 2007, 2010), we predicted differential activation in response to alternating transitivity verbs in the supramarginal or angular gyri bilaterally, due to the greater number of thematic roles they encode on their transitive reading, and their overall greater semantic/conceptual complexity, reflected also in the fact that they have two thematic grids. The existence of multiple thematic grids can be viewed as a type of lexical ambiguity. As such, we predicted that alternating transitivity verbs would also elicit activation in mid-superior frontal brain regions, associated in previous studies with the processing of words with multiple meanings (Chan, 2004; Mason & Just, 2007).
Along with the previous results mentioned above, such a result would reinforce lexical-thematic approaches to argument structure, since only under this view thematic information is listed in the lexicon and can differentiate verbs appearing in isolation. In contrast, a generative-constructivist account, stating that verbs do not carry any thematic information outside of a syntactic context, predicts no differential activation in response to alternating transitivity as opposed to simple intransitive verbs presented in isolation, since the lexical representation of the two verb types does not differ.5
Method
Participants
Participants included 14 young healthy volunteers (11 women), with ages ranging from 19 to 29 years (M = 23). All were right-handed, monolingual English speakers. None had a history of neurological or psychological disease or speech, language, or learning problems. Participants signed written informed consent approved by the Institutional Review Board at Northwestern University.
Stimuli
Stimuli consisted of 96 pseudowords and 42 verbs, including 20 ‘alternating transitivity’ verbs and 22 simple intransitive verbs. Verbs were classified as members of these groups based on Levin & Rappaport Hovav's (1995) identification of verb classes and their members. Specifically, the alternating verbs all belonged either to the class of non-agentive verbs of manner of motion, or to the class of externally caused verbs of change of state. Verb in both of these classes exhibit a transitive-intransitive alternation, and are shown by Levin & Rappaport Hovav (1995) to exhibit unaccusative properties on their intransitive use. Stimuli were presented visually in capital letter strings. All verbs were presented in the infinitive form, as in break and sing. The two verb classes were matched for number of syllables (t (40) = 0.15, p = 0.88) and for frequency of occurrence according to the CELEX database (Baayen, Piepenbrock & van Rijn, 1993) (M log alternating transitivity verb frequency = 1.46; M log simple verb frequency = 1.23, t (40) = 1.47, p = .15). Since verb-noun ambiguity is very widespread in English, many of the verbs in both groups (15 of the 20 alternating verbs and 16 of the 22 simple verbs) had an additional, nominal reading. However, the relative frequency of this reading compared to the verb frequency of the word was generally small, and did not differ between the two verb groups, such that the degree of lexical-class ambiguity was similar in the two verb classes (M nominal frequency / verbal frequency for alternating verbs: 0.32, M nominal frequency / verbal frequency for simple verbs: 0.19, t (40) = 1.33, p = .19) (see Appendix for lists of verbs used in the experiment).
Procedures
An event-related design was used in which words and pseudowords were visually displayed for 1000 ms, followed by a jittered interstimulus fixation cross lasting between 1900 ms and 8700 ms. The trial order and timing of the interstimulus intervals were calculated and optimized using the OPTSEQ program (http://surfer.nmr.mgh.harvard.edu/optseq). The experiment was comprised of two runs, each including the 42 verbs along with 48 pseudowords, in different orders. Participants were instructed to respond to the letter strings by pressing a button with their left hand when seeing a pseudoword.
Data Acquisition
Scanning was carried out on a Siemens Verio 3T scanner. A T1-weighted anatomical scan was made at the start of each protocol. During the experimental runs, functional volumes with BOLD contrast were obtained using gradient echo-planar imaging sequences (TR = 2000 ms; TE = 30 ms; flip angle = 90°; matrix size = 64 × 64; voxel size = 3.44 × 3.44 × 3 mm; 31 slices).
Data Analysis
Data analysis was performed with SPM8 (http://www.fil.ion.ucl.ac.uk/spm). Functional scans were realigned to a mean functional volume and corrected for slice-acquisition timing. The anatomical volume was coregistered to the mean image and normalized to the Montreal Neurological Institute (MNI) 152-subject template brain (ICBM, NIH P-20 project). The functional volumes were then normalized using the same transformation, resliced to a voxel size of 3 × 3 × 3 mm, and spatially smoothed using an 8 mm Gaussian kernel.
In the first-level statistical analysis, a high-pass filter of 128 seconds was used to eliminate scanner drift. For each run, six movement parameters obtained during preprocessing were entered as regressors. Additionally, an index of the degree of lexical class ambiguity of each verb (operationally defined as N freq / V freq) was incorporated as a parametric modulator into the two experimental conditions, to eliminate variability within conditions arising from this dimension of the stimuli. Individual participants' contrast maps for the alternating and simple verb conditions were then entered into second-level one-sample t-tests. These were evaluated at a threshold of p < .005, uncorrected, with a minimum cluster size (k) of 25 contiguous voxels (675 mm3).6
Results
Five clusters of differential activation were observed for the contrast of alternating transitivity over simple verbs, with the ambiguity index covaried out. Two large clusters were observed in the left hemisphere: one in the angular and supramarginal gyri (BAs 39 and 40), extending to the posterior superior and middle temporal gyri, and the other in the middle and superior frontal gyri (BAs 8 and 9). Two similar though smaller clusters were observed in the right hemisphere homologues of these regions. Finally, a small activation cluster was found in the anterior cingulate (see Figure 1 and Table 1). The opposite contrast, of simple over alternating verb processing, revealed no differential activation, even using the most lenient statistical threshold.
Figure 1.

Differential activation for the contrast of ‘alternating transitivity’ verbs over simple intransitive verbs (p < .005, uncorrected, k ≥ 25), with relative noun frequency co-varied out. Clusters of activation are observed bilaterally in the inferior parietal lobule (BAs 39 and 40) and prefrontal cortex (BAs 8 and 9), and in the anterior cingulate.
Table 1.
Peak MNI coordinates, cluster sizes (k), maximal t-values and cluster-corrected p-values for the clusters showing differential activation between ‘alternating transitivity’ verbs and simple intransitive verbs (p < .005, uncorrected, k ≥ 25)
| Contrast | Region | Peak coordinates | k | t | p | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Alternating > unergative | LH AG, SMG, pSTG, pMTG | -39 | -70 | 46 | 218 | 4.37 | .034 |
| LH MFG, SFG | -12 | 38 | 34 | 208 | 4.30 | .041 | |
| anterior cingulate | -6 | 29 | -2 | 32 | 4.42 | .918 | |
| RH AG, SMG, pSTG, pMTG | 51 | -61 | 25 | 109 | 4.32 | .269 | |
| RH MFG, SFG | -18 | 32 | 40 | 69 | 4.86 | .565 | |
|
| |||||||
| Unergative > alternating | None | ||||||
Abbreviations: LH = left hemisphere, RH = right hemisphere, AG = angular gyrus, SMG = supramarginal gyrus, pMTG = posterior middle temporal gyrus, pSTG = posterior superior temporal gyrus, MFG = middle frontal gyrus, SFG = superior frontal gyrus
Discussion
Results of the present study show differential clusters of activation in response to the contrast between alternating transitivity verbs and simple, intransitive verbs, presented in isolation. In line with lexical-thematic accounts of argument structure, these data indicate that verbs of alternating transitivity entail a more complex argument structure representation than simple intransitive verbs, thus requiring greater processing resources, which results in recruitment of greater posterior perisylvian and middle/superior frontal tissue. Let us discuss this point in more detail.
As noted in the introduction, alternating transitivity verbs and intransitive verbs differ on two dimensions. First, alternating verbs are associated with two thematic grids (transitive and intransitive), whereas intransitive verbs are associated with only one grid. Second, the grids themselves differ between the two verb types. In particular, the alternating verbs have a transitive grid as one of their options, whereas the simple verbs are invariably intransitive. Alternating transitivity verbs thus exhibit a greater number of thematic grids, as well as a greater number of thematic roles on one of these grids, than simple intransitive verbs.
Previous studies examining the effect of number of thematic roles (Ben Shachar et al., 2003; Den Ouden et al., 2009; Thompson et al., 2007, 2010) found increased posterior perisylvian activation associated with an increase in the number of thematic roles. Notably, Thompson et al. (2007, 2010), who used the same lexical decision task as the one used here, with 1-, 2-, and 3-argument verbs, found an effect of number of thematic roles only in bilateral posterior perisylvian regions, with activation peaks very close to the posterior peaks found in the current experiment. It is thus reasonable to suggest that the posterior activations identified in the present study reflect, at least in part, the fact that alternating verbs have a transitive thematic grid (as one of their thematic options), whereas simple verbs are intransitive. These results thus reinforce the role of the angular and supramarginal gyri in both hemispheres in processing verbs with multiple thematic roles. More generally, they are in line with the many findings showing involvement of the inferior parietal lobule in semantic tasks, as verbs with more thematic roles denote more complex events, involving more participants. Binder et al.'s (2009) meta-analysis shows that the angular gyrus is the region most consistently implicated in semantic processing tasks. While activations due to semantic processing were more common in the left hemisphere, there were many activation foci in the right angular gyrus, in line with the current results, showing right hemisphere posterior perisylvian recruitment associated with semantic complexity.
In addition to a greater number of thematic roles, however, alternating verbs also exhibit a greater number of thematic grids than simple intransitive verbs, as explained above. We suggest that this difference is reflected in the frontal activations we found in the middle and superior frontal gyri (BAs 8 and 9), which were associated with processing of alternating transitivity verbs. These regions were not implicated in the studies mentioned above investigating only number of thematic roles. They were, however, detected in a number of studies targeting the processing of lexical items associated with several interpretation options. Chan et al. (2004) found differential activation in this region in the left hemisphere in response to processing Chinese words with semantic ambiguity as opposed to words without such ambiguity. Similarly, Mason & Just (2007) found bilateral mid-superior frontal cortex activation during processing of ambiguous stimuli. These regions were also implicated in Ketteler, Kastrau, Vohn, & Huber's (2008) study of lexical ambiguity processing. In addition, Shetreet et al. (2007) detected activation in left BA 9 in response to processing verbs with multiple syntactic realization options. The current results are in line with these previous studies, as alternating transitivity verbs are likewise lexically ambiguous. Interestingly, unlike prototypical cases of polysemy, the difference between the two senses of these verbs is not captured in terms of semantic features (as in e.g. plant the biological entity vs. plant the physical location), or lexical category (as in e.g. the noun a nurse vs. the verb to nurse). Rather, it can be captured in terms of the number of participants in the event (one or two). This information is reflected in the fact that these verbs have two thematic grids. Taken together, these findings suggest that mid-superior frontal regions are involved in the processing of lexical ambiguity or the consideration of multiple lexical options. This is consistent with the commonly held assumption that dorsolateral prefrontal regions in the two hemispheres play a role in general working memory and maintenance processes (see D'esposito, 2001; Curtis & D'esposito, 2003 for reviews).
Interestingly, in their cross-modal lexical decision experiments, Shapiro & Levine (1990) and Shapiro, Gordon, Hack, & Killackey (1993) found that Broca's aphasic individuals behaved on a par with healthy participants, displaying longer reaction times to verbs with multiple syntactic realization options compared with simple verbs. This seems to contradict the claim made here, namely that frontal regions are involved in the processing of multiple lexical options. We note, however, that Shapiro et al. (1990, 1993) did not include anatomical descriptions of the lesion suffered by their aphasic participants. It is possible that the lesions involved left inferior frontal regions (as suggested by lesion analysis studies such as Dronkers, Wilkins, Van Valin, Redfern, & Jaeger, 2004; Kertesz, Harlock, & Coates, 1979), leaving intact the left and/or right middle superior frontal regions implicated in the processing of lexical ambiguity. Shetreet et al., (2007), who also found recruitment of left BA 9 in the processing of multiple lexical options, suggests that posterior regions may suffice in order to access lexical information (as suggested also below), accounting for the sensitivity of Broca's aphasic individuals to number of syntactic realization options, whereas frontal regions may be crucial only for sustaining activation of multiple lexical options.
A last remark with regard to the frontal activations found in the current study is in order. Thompson-Schill, D'esposito, Aguirre, & Farah (1997) claimed that left inferior prefrontal areas contribute to semantic search and selection. Interestingly, Chan and colleagues (2004) argue that their own findings are inconsistent with Thompson-Schill's hypothesis. The authors claim that although their task involved the identification of semantically ambiguous words, which hypothetically involves a high search and selection load, the activations they found were located in the left mid-superior frontal gyri, not in the left inferior frontal gyrus, as predicted by Thompson-Schill's hypothesis. However, Thompson-Schill's tasks manipulated the need to select a relevant feature of semantic knowledge from a set of competing alternatives. Our experimental task (as well as the tasks of Mason & Just, 2007, Shetreet et al., 2007, and possibly even Chan et al., 2004) entailed only activation of multiple options associated with a lexical item, and not selection from among activated semantic features. Thus, the fact that neural activity was found in more superior regions is not incompatible with the claim of Thompson-Schill and colleagues. The middle-superior frontal activations detected in our study thus possibly reflect general processing of multiple lexical options, which is independent of argument structure processing.
An interesting question that cannot be answered based on our study, is whether the increased number of thematic grids in the case of alternating verbs contributes also the posterior activations we found, or whether those were elicited only due to the increased number of thematic roles exhibited by alternating verbs. One possible piece of evidence to suggest that processing a greater number of thematic options involves superior temporal and inferior parietal regions comes from lesion data: several studies have shown that Wernicke's aphasic individuals do not perform normally on tasks involving verbs with multiple syntactic realization options. For example, In Shapiro & Levine (1990) and Shapiro et al.'s (1993) cross-modal lexical decision experiments, results showed that unlike healthy controls (and Broca's aphasic individuals), participants with Wernicke's aphasia did not show on-line sensitivity to the thematic properties of the presented verbs, performing in a comparable manner on verbs that differed in complexity in terms of thematic options. Although these studies do not provide information regarding the localization of the patients' lesions, given that Wernicke's aphasia often results from damage to posterior temporal and inferior parietal regions (e.g. Kertesz, 1977, Kertesz et al., 1979), including the angular gyrus (Dronkers et al., 2004), the insensitivity to the multiplicity of thematic options may be attributed to posterior perisylvian lesions. This would suggest that the posterior activations we found in our experiment reflect not only the processing of multiple thematic roles, but also the processing of multiple thematic options. In the future, it will be interesting to compare alternating transitivity verbs to simple transitive verbs, to further test this hypothesis.
A few words are in order about the anterior cingulate activation we found. Although the detected cluster was small, this result is consistent both with Shetreet et al. (2007), who found activation in the right anterior cingulate, very close to the midline, in response to verbs with greater as opposed to smaller number of thematic roles, and with Chan et al. (2004), where this region was implicated in the processing of ambiguous stimuli. Taken together, these results suggest that although the anterior cingulate is not considered a ‘classic’ language area, it might play a role in the processing of complex argument structures.
Finally, the findings from the present study are of interest with regard to linguistic theory. As mentioned in the introduction, there is an ongoing debate in linguistic literature about the nature of the lexicon-syntax interface. Lexical-thematic approaches hold that thematic information is present in the lexicon, whereas generative-constructivist approaches claim that verbal entries do not carry any grammatically relevant information, including argument structure specification. In our experiment, we found differential activation associated with alternating transitivity verbs and simple intransitives when these appeared in isolation. This is in line with the first approach, as it entails a lexical difference between the two verb types, which can only be attributed to argument structure. As explained in the introduction, a further split exists within the lexical-thematic view, with regard to alternating transitivity. According to one approach, both the transitive and the intransitive alternates are listed in the lexicon; the second approach argues that only a verbal root is listed, carrying the information that it can give rise to two syntactically-composed verbs: a transitive and an intransitive. At present, the results of the current experiment cannot shed light on this debate. Both lexical-thematic views hold that alternating transitivity verbs exhibit an inherent ambiguity, whether reflected in the actual listing of two verbal alternates or in the information that the root can give rise to two verbal structures, and this ambiguity arguably elicited in the frontal activation we found. The posterior activation too can be explained by both views. On the first account, upon encountering an alternating verb, e.g. break, the speaker accesses its lexical entry, which includes two verbal alternates, one of them with a transitive thematic grid. This complex lexical-semantic representation elicits posterior perisylvian activation. According to the second view, a speaker encountering the root break will engage in building two syntactic structures, one for the intransitive verb break and another for the transitive one. It has been suggested by various authors (Constable et al., 2004; Cooke et al., 2002; Hasson, Nusbaum, & Small, 2006; Schlesewsky & Bornkessel, 2004) that superior temporal / inferior parietal regions are involved in syntactic processing. Thus, the posterior activations found in the current study can be attributed to syntactic structure building.
While the results can be accounted for by both lexical-thematic approaches, it is clear that they undermine a strict constructionist account, which assumes that no argument structure information whatsoever is listed in the lexicon. If that were the case, than it would be unclear why the contrast between processing the two verbal entry types should invoke any differential activation, since both verb types have similar lexical representations and are complex to the same degree.
Conclusion
Alternating transitivity verbs in English present an interesting case of complex argument structure representation, in which the same verb can be used either transitively or intransitively. Contrasting such verbs with simple intransitive verbs, we found activation elicited by the alternating verbs in posterior perisylvian and mid-superior frontal regions bilaterally. Although the study cannot distinguish between activations associated with increased number of thematic grids and those associated with increased number of thematic roles, our findings are consistent with previous studies indicating that processing verbs with a greater number of thematic roles involves the angular and supramarginal gyri bilaterally. The frontal activation found in our study is conceivably associated with the processing of multiple lexical options, consistent with previous results implicating this region in ambiguity processing. Finally, the differential activation found for alternating transitivity versus simple verbs strongly suggests that some thematic information is included in the lexical representation of verbs.
Acknowledgments
This research was supported by NIH grant R01-DC01948-17 (C.K. Thompson). The authors wish to thank Sladjana Lukic, Alex Baria and Eisha Wali for their assistance with stimulus preparation and data collection, Jennifer Mack, Ph.D, for helpful comments and suggestions, and Aneta Kielar, Ph.D, for assistance with data analysis.
Appendix
Verbs used in the experiment with their verb and noun frequency according to the CELEX database (Baayen, Piepenbrock & van Rijn, 1993)
| Verb | V frequency | N freqency | N freq / V freq |
|---|---|---|---|
| Alternating | |||
|
| |||
| break | 4102 | 542 | 0.132 |
| drop | 2551 | 586 | 0.230 |
| roll | 1287 | 323 | 0.251 |
| bend | 1179 | 179 | 0.152 |
| tear | 1101 | 1093 | 0.993 |
| sink | 892 | 261 | 0.293 |
| boil | 768 | 172 | 0.224 |
| burst | 682 | 230 | 0.337 |
| fade | 643 | 0 | 0 |
| float | 585 | 183 | 0.313 |
| melt | 436 | 0 | 0 |
| shrink | 364 | 5 | 0.0137 |
| crack | 333 | 398 | 1.195 |
| crash | 335 | 277 | 0.827 |
| bounce | 289 | 39 | 0.135 |
| skid | 59 | 40 | 0.678 |
| expand | 832 | 0 | 0 |
| collapse | 518 | 284 | 0.548 |
| dissolve | 330 | 0 | 0 |
| implode | 0 | 0 | 0 |
|
| |||
| Mean (SD) | 0.316 (0.354) | ||
|
| |||
| Unergatives | |||
|
| |||
| gallop | 110 | 30 | 0.273 |
| laugh | 3058 | 453 | 0.148 |
| cry | 2158 | 526 | 0.244 |
| sing | 1407 | 0 | 0 |
| weep | 499 | 0 | 0 |
| swim | 837 | 165 | 0.197 |
| breathe | 791 | 0 | 0 |
| leap | 549 | 141 | 0.257 |
| pray | 505 | 0 | 0 |
| crawl | 454 | 14 | 0.031 |
| blink | 246 | 21 | 0.085 |
| sniff | 229 | 26 | 0.114 |
| cough | 223 | 211 | 0.946 |
| bark | 186 | 126 | 0.677 |
| skip | 168 | 23 | 0.137 |
| wink | 142 | 59 | 0.415 |
| snore | 69 | 20 | 0.290 |
| wander | 661 | 0 | 0 |
| hobble | 98 | 16 | 0.163 |
| scurry | 87 | 3 | 0.034 |
| slither | 96 | 0 | 0 |
| scuttle | 67 | 16 | 0.239 |
|
| |||
| Mean (SD) | 0.193 (0.236) | ||
Footnotes
Palti, Ben-Shachar, Hendler, & Hadar (2007) found a similar activation when contrasting the processing of verbs to that of nouns. The authors hypothesized that this activation might be due to the fact that verbs have thematic roles, whereas (concrete) nouns do not.
Shetreet, Friedmann, & Hadar (2010b) examined the neural correlates of processing sentences with unaccusative verbs, and found that it was associated with activation in the left inferior frontal gyrus and middle temporal gyrus. The verbs in Shetreet et al.'s study, unlike the alternating transitivity verbs discussed here, were exclusively unaccusative, and not associated with two argument structures.
It may seem preferable to compare activations caused by alternating transitivity verbs to those caused by so-called non-alternating unaccusative verbs, e.g. appear, fall, selecting for a theme argument. However, in addition to the fact that the latter verbs are very rare in English, several authors have argued that verbs such as fall in fact do have a transitive alternate, existing as a ‘frozen’ form in the mental lexicon (see Horvath & Siloni, 2008 for theoretical arguments, and Fadlon, to appear, for experimental evidence for the psychological reality of such ‘frozen’ transitive alternates in Hebrew). If this were the case, then these verbs also would be considered alternating and the contrast between them and alternating transitivity verbs would, hence, predict no differential activation. Another option would be to compare alternating transitivity verbs to simple transitive verbs, e.g. lock, not displaying a transitive-intransitive alternation. As in the current design, these two verb types would differ on two dimensions: number of thematic grids (two for alternating verbs, one for transitive verbs), and, under the intransitive use of alternating transitivity verbs, also number of thematic roles (one vs. two).
Although, as noted by an LCP reviewer, there is evidence to suggest that all argument structure options are activated with the verb, even when it appears in a sentence, where naturally only one option is realized. Such evidence was presented with regard to verbs with multiple complementation options in Shapiro et al. (1987), Shetreet et al. (2007, 2010a). In a study investigating the processing of sentences with alternating unaccusatives, Friedmann et al. (2008) attribute the inconsistent priming at the trace position to the activation of the transitive, in addition to the unaccusative, alternate of the verb.
The generative-constructivist approach, for that matter, would not predict differential brain activations in response to any argument structure contrast (e.g. simple transitive vs. simple intransitive verbs) when verbs are presented in isolation, since lexical representations of verbs do not contain any thematic information.
This threshold was reported in several papers investigating similar linguistic contrasts (e.g. Shetreet et al.'s 2010 study of unaccusative vs. unergative verbs; Chan et al.'s 2004 study of ambiguous vs. unambiguous words in Chinese; Mason & Just's 2007 study of lexical ambiguity vs. nonambiguity in sentence contexts) as well as other subtle linguistic phenomena (e.g. Altmann, Bledowski, Wibral, & Kaiser, 2007; Santi & Grodzinsky, 2007).
References
- Altmann CF, Bledowski C, Wibral M, Kaiser J. Processing of location and pattern changes of natural sounds in the human auditory cortex. Neuroimage. 2007;35:1192–1200. doi: 10.1016/j.neuroimage.2007.01.007. [DOI] [PubMed] [Google Scholar]
- Baayen RH, Piepenbrock R, van Rijn H. The CELEX lexical database (Release 1) [CD-ROM] Philadelphia, PA: Linguistic Data Consortium, University of Pennsylvania; 1993. [Google Scholar]
- Bastiaanse R, Jonkers R. Verb retrieval in action naming and spontaneous speech in agrammatic and anomic aphasia. Aphasiology. 1998;12:951–969. [Google Scholar]
- Ben-Shachar M, Hendler T, Kahn I, Ben-Bashat D, Grodzinsky Y. The neural reality of syntactic transformations. Psychological Science. 2003;14:433–440. doi: 10.1111/1467-9280.01459. [DOI] [PubMed] [Google Scholar]
- Binder JR, Desai RH, Graves WW, Conant LL. Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cerebral Cortex. 2009;19:2767–2796. doi: 10.1093/cercor/bhp055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borer H. The grammar machine. In: Artemis A, Anagnostopoulou E, Everaert M, editors. The Unaccusativity Puzzle: Explorations of the Syntax-Lexicon Interface. Oxford: Oxford University Press; 2004. pp. 288–331. [Google Scholar]
- Borer H. The Normal Course of Events: Structuring Sense. II. Oxford: Oxford University Press; 2005. [Google Scholar]
- Burzio L. Italian Syntax. Dordrecht: Reidel; 1986. [Google Scholar]
- Chan AHD, Liu H, Yip V, Fox PT, Gao J, Tan LH. Neural systems for word meaning modulated by semantic ambiguity. Neuroimage. 2004;22:1128–1133. doi: 10.1016/j.neuroimage.2004.02.034. [DOI] [PubMed] [Google Scholar]
- Chierchia G. A semantics for unaccusatives and its syntactic consequences. In: Alexiadou A, Anagnostopoulou E, Everaert M, editors. The unaccusativity puzzle: Studies on the syntax-lexicon interface. Oxford: Oxford University Press; 2004. pp. 60–84. [Google Scholar]
- Chomsky N. Lectures on Government and Binding. Hague: Mouton de Gruyter; 1981. [Google Scholar]
- Constable RT, Pugh KR, Berroya E, Mencl WE, Westerveld M, Ni W, et al. Sentence complexity and input modality effects in sentence comprehension: an fMRI study. Neuroimage. 2004;22:11–21. doi: 10.1016/j.neuroimage.2004.01.001. [DOI] [PubMed] [Google Scholar]
- Cooke A, Zurif EB, DeVita C, Alsop D, Koenig P, Detre J, et al. Neural basis for sentence comprehension: Grammatical and short-term memory components. Human Brain Mapping. 2002;15:80–94. doi: 10.1002/hbm.10006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtis CE, D'esposito M. Persistent activity in the prefrontal cortex during working memory. Trends in Cognitive Sciences. 2003;7:415–423. doi: 10.1016/s1364-6613(03)00197-9. [DOI] [PubMed] [Google Scholar]
- De Bleser R, Kauschke C. Acquisition and loss of nouns and verbs: Parallel or divergent patterns? Journal of Neurolinguistics. 2003;16:213–229. [Google Scholar]
- Den Ouden D, Fix S, Parrish T, Thompson C. Argument structure effects in action verb naming in static and dynamic conditions. Journal of Neurolinguistics. 2009;22:196–215. doi: 10.1016/j.jneuroling.2008.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Esposito M. Functional neuroimaging of working memory. In: Cabeza R, Kingstone A, editors. Handbook of Functional Neuroimaging of Cognition. Cambridge, MA: MIT Press; 2001. pp. 293–328. [Google Scholar]
- Dragoy O, Bastiaanse R. Verb production and word order in Russian agrammatic speakers. Aphasiology. 2010;24:28–55. [Google Scholar]
- Dronkers NF, Wilkins DP, Van Valin RD, Jr, Redfern BB, Jaeger JJ. Lesion analysis of the brain areas involved in language comprehension. Cognition. 2004;92:145–177. doi: 10.1016/j.cognition.2003.11.002. [DOI] [PubMed] [Google Scholar]
- Embick D. Unaccusative syntax and verbal alternations. In: Artemis A, Anagnostopoulou E, Everaert M, editors. The Unaccusativity Puzzle: Explorations of the Syntax-Lexicon Interface. Oxford: Oxford University Press; 2004. pp. 137–158. [Google Scholar]
- Fadlon J. The hidden existence of derivational gaps: a psycholinguistic study. In: Everaert M, Marelj M, Siloni T, editors. The Theta System: Argument Structure at the Interface. Oxford: Oxford University Press; to appear. [Google Scholar]
- Friedmann N, Taranto G, Shapiro LP, Swinney D. The leaf fell (the leaf): The online processing of unaccusatives. Linguistic Inquiry. 2008;39:355–377. doi: 10.1162/ling.2008.39.3.355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasson U, Nusbaum HC, Small SL. Repetition suppression for spoken sentences and the effect of task demands. Journal of Cognitive Neuroscience. 2006;18:2013–2029. doi: 10.1162/jocn.2006.18.12.2013. [DOI] [PubMed] [Google Scholar]
- Horvath J, Siloni T. Adjectival Passives: Active Lexicon. In: Armon-Lotem S, Danon G, Rothstein S, editors. Current Issues in Generative Hebrew Linguistics. Amsterdam: John Benjamins; 2008. pp. 105–134. [Google Scholar]
- Horvath J, Siloni T. Causatives across components. Natural Language and Linguistic Theory. 2011;29:657–704. [Google Scholar]
- Kertesz A. Isotope localization of infarcts in aphasia. Archives of Neurology. 1977;34:590–601. doi: 10.1001/archneur.1977.00500220024004. [DOI] [PubMed] [Google Scholar]
- Kertesz A, Harlock W, Coates R. Computer tomographic localization, lesion size, and prognosis in aphasia and nonverbal impairment. Brain and Language. 1979;8:34–50. doi: 10.1016/0093-934x(79)90038-5. [DOI] [PubMed] [Google Scholar]
- Ketteler D, Kastrau F, Vohn R, Huber W. The subcortical role of language processing. High level linguistic features such as ambiguity-resolution and the human brain: An fMRI study. Neuroimage. 2008;39:2002–2009. doi: 10.1016/j.neuroimage.2007.10.023. [DOI] [PubMed] [Google Scholar]
- Kim M, Thompson CK. Patterns of comprehension and production of nouns and verbs in agrammatism: Implications for lexical organization. Brain and Language. 2000;74:1–25. doi: 10.1006/brln.2000.2315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim M, Thompson CK. Verb deficits in Alzheimer's disease and agrammatism: Implications for lexical organization. Brain and Language. 2004;88:1–20. doi: 10.1016/s0093-934x(03)00147-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin B. English Verb Classes and Alternations: A Preliminary Investigation. Chicago, IL: University of Chicago Press; 1993. [Google Scholar]
- Levin B, Rappaport Hovav M. Unaccusativity: At the Syntax-Lexical Semantics Interface. Cambridge, MA: MIT Press; 1995. [Google Scholar]
- Luzzatti C, Raggi R, Zonca G, Pistarini C, Contardi A, Pinna GD. Verb-noun double dissociation in aphasic lexical impairments: The role of word frequency and imageability. Brain and Language. 2002;81:432–444. doi: 10.1006/brln.2001.2536. [DOI] [PubMed] [Google Scholar]
- Marantz A. No escape from syntax: Don't try morphological analysis in the privacy or your own lexicon. University of Pennsylvania Working Papers in Linguistics. 1997;4:201–225. [Google Scholar]
- Mason R, Just M. Lexical ambiguity in sentence comprehension. Brain Research. 2007;1146:115–127. doi: 10.1016/j.brainres.2007.02.076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palti D, Ben-Shachar M, Hendler T, Hadar U. Neural correlates of semantic and morphological processing of Hebrew nouns and verbs. Human Brain Mapping. 2007;28:303–314. doi: 10.1002/hbm.20280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perlmutter DM. Impersonal passives and the unaccusative hypothesis. Proceedings of the Annual Meeting of the Berkeley Linguistic Society. 1978;4:159–189. [Google Scholar]
- Ramchand G. Verb Meaning and the Lexicon: A First-Phase Syntax. Cambridge, UK: Cambridge University Press; 2008. [Google Scholar]
- Reinhart T. The Theta System: An overview. Theoretical Linguistics. 2002;28:229–290. [Google Scholar]
- Santi A, Grodzinsky Y. Working memory and syntax interact in Broca's area. Neuroimage. 2007;37:8–17. doi: 10.1016/j.neuroimage.2007.04.047. [DOI] [PubMed] [Google Scholar]
- Schäfer F. The causative alternation. Language and Linguistics Compass. 2009;3:641–681. [Google Scholar]
- Schlesewsky M, Bornkessel I. On incremental interpretation: Degrees of meaning accessed during sentence comprehension. Lingua. 2004;114:1213–1234. [Google Scholar]
- Shapiro L, Levine B. Verb processing during sentence comprehension in aphasia. Brain and Language. 1990;38:21–47. doi: 10.1016/0093-934x(90)90100-u. [DOI] [PubMed] [Google Scholar]
- Shapiro L, Zurif E, Grimshaw J. Sentence processing and the mental representation of verbs. Cognition. 1987;27:219–246. doi: 10.1016/s0010-0277(87)80010-0. [DOI] [PubMed] [Google Scholar]
- Shapiro L, Gordon B, Hack N, Killackey J. Verb argument processing in complex sentences in Broca and Wernicke's aphasia. Brain and Language. 1993;45:423–447. doi: 10.1006/brln.1993.1053. [DOI] [PubMed] [Google Scholar]
- Shetreet E, Friedmann N, Hadar U. The cortical representation of verbs with optional complements: The theoretical contribution of fMRI. Human Brain Mapping. 2010a;31:770–785. doi: 10.1002/hbm.20904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shetreet E, Friedmann N, Hadar U. The neural correlates of linguistic distinctions: Unaccusative and unergative verbs. Journal of Cognitive Neuroscience. 2010b;22:2306–2315. doi: 10.1162/jocn.2009.21371. [DOI] [PubMed] [Google Scholar]
- Shetreet E, Palti D, Friedmann N, Hadar U. Cortical representation of verb processing in sentence comprehension: Number of complements, subcategorization, and thematic frames. Cerebral Cortex. 2007;17:1958–1969. doi: 10.1093/cercor/bhl105. [DOI] [PubMed] [Google Scholar]
- Thompson CK, Bonakdarpour B, Fix S. Neural mechanisms of verb argument structure processing in agrammatic aphasic and healthy age-matched listeners. Journal of Cognitive Neuroscience. 2010;22:1993–2011. doi: 10.1162/jocn.2009.21334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson CK, Bonakdarpour B, Fix S, Blumenfeld H, Parrish T, Gitelman D, Mesulam M. Neural correlates of verb argument structure processing. Journal of Cognitive Neuroscience. 2007;19:1753–1767. doi: 10.1162/jocn.2007.19.11.1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson CK, Lange K, Schneider S, Shapiro L. Agrammatic and non-brain-damaged subjects' verb and verb argument structure production. Aphasiology. 1997;11:473–490. [Google Scholar]
- Thompson CK, Shapiro L, Li L, Schendel L. Analysis of verbs and verb-argument structure: A method for quantification of aphasic language production. Clinical Aphasiology. 1995;23:121–140. [Google Scholar]
- Thompson-Schill SL, D'Esposito M, Aguirre GK, Farah MJ. Role of left prefrontal cortex in retrieval of semantic knowledge: A re-evaluation. Proceedings of the National Academy of Science. 1997;94:14792–14797. doi: 10.1073/pnas.94.26.14792. [DOI] [PMC free article] [PubMed] [Google Scholar]