

Abstract
The anemia of sickle cell disease is associated with a severe inflammatory vasculopathy and endothelial dysfunction, which leads to painful and life-threatening clinical complications. Growing evidence supports the anti-inflammatory properties of ω-3 fatty acids in clinical models of endothelial dysfunction. Promising but limited studies show potential therapeutic effects of ω-3 fatty acid supplementation in sickle cell disease. Here, we treated humanized healthy and sickle cell mice for 6 weeks with ω-3 fatty acid diet (fish-oil diet). We found that a ω-3 fatty acid diet: (i) normalizes red cell membrane ω-6/ω-3 ratio; (ii) reduces neutrophil count; (iii) decreases endothelial activation by targeting endothelin-1 and (iv) improves left ventricular outflow tract dimensions. In a hypoxia-reoxygenation model of acute vaso-occlusive crisis, a ω-3 fatty acid diet reduced systemic and local inflammation and protected against sickle cell-related end-organ injury. Using isolated aortas from sickle cell mice exposed to hypoxia-reoxygenation, we demonstrated a direct impact of a ω-3 fatty acid diet on vascular activation, inflammation, and anti-oxidant systems. Our data provide the rationale for ω-3 dietary supplementation as a therapeutic intervention to reduce vascular dysfunction in sickle cell disease.
Introduction
Sickle cell disease (SCD) is a worldwide hereditary red cell disorder caused by a point mutation in the β-globin gene resulting in the synthesis of pathological hemoglobin S (HbS), and abnormal HbS polymerization in hypoxic conditions.1 The two main clinical manifestations of SCD are chronic hemolytic anemia and acute vaso-occlusive crises (VOC), which are the principal causes of hospitalization of SCD patients. Recent studies have underscored the central role of sickle vasculopathy in the generation of sickle cell-related acute events and chronic organ complications.2–4 The pathophysiology of these complications is based on intravascular sickling in capillaries and small vessels leading to vaso-occlusion, impaired blood flow, vascular inflammation, and thrombosis with ischemic cell damage.2–4 Studies in various models of vasculopathy, including those with ischemia and inflammation, have shown protective effects of ω-3 polyunsaturated fatty acid (PUFA) supplementation.5 This is supported by several mechanisms: (i) favorable changes in cell membrane lipid composition;6 (ii) modulation of soluble and cellular inflammatory responses;7 (iii) modulation of the coagulation cascade,8 and (iv) production of nitric oxide.9
The fatty acid profile of the red cell membrane from SCD patients is intrinsically different from that of healthy controls, with a relative increase in the ratio of ω-6 to ω-3 PUFA.10,11 Sickle erythrocytes have increased ω-6-derived arachidonic acid (20:4n6) and decreased eicosapentaenoic acid (20:5n3) and docosahexaenoic acid (22:6n3).10,11 Recently, human studies have demonstrated that supplementation of SCD subjects with fish oil containing ω-3 fatty acids reduces VOC, pain episodes, and blood transfusions.12–14 To date, however, there are insufficient clinical and molecular data to support routine dietary supplementation with fish oil for SCD patients.
In the present study, we studied the effects of an ω-3 diet (fish oil diet, FD) on the vasculopathy of a humanized mouse model for SCD. The evaluation of the impact of FD supplementation on SCD is relevant since the typical Western diet has a 15:1 ratio of ω-6 to ω-3 PUFA.15
We found that in SCD mice, the FD modified sickle red cell membrane lipid composition and was associated with a reduction in vascular activation and an improvement of cardiovascular dysfunction. In a model of acute VOC induced by exposure to hypoxia-reoxygenation (H/R) stress, we showed that FD decreases vascular activation, modulates the inflammatory response, and reduces sickle cell-related organ damage. Thus, our findings shed new light on the significance of PUFA in SCD and VOC and support a therapeutic role of ω-3 PUFA supplementation in SCD.
Methods
Mice and study design
Experiments were performed on 4- to 6-week old healthy control (Hbatm1(HBA)Tow Hbbtm3(HBG1,HBB)Tow) and SCD (Hbatm1(HBA)Tow Hbbtm2(HBG1,HBB*)Tow) mice.16,17 The animal protocol was approved by the Animal Care and Use Committee of the University of Verona (CIRSAL). Two-month old animals were fed for 6 weeks with either the standard AIN-93M purified rodent diet (soy-diet with n6/n3 ratio of 8:1-Dyets Inc., Bethlehem, PA, USA), containing 140 g/kg casein, 1.8 g/kg L-cystine, 100 g/kg sucrose, 465.9 g/kg cornstarch, 155 g/kg dextrose, 40 g/kg soybean oil, 0.8 mg/kg t-butylhydroquinone, 50 g/kg cellulose, 35 g/kg mineral mix, 10 g/kg vitamin mix, and 2.5 g/kg choline bitartrate18 or the ω-3 FD, an AIN-93M-based purified rodent diet in which all of the calories provided by fat (10%) are replaced by 7.9% from HCO and 2.1% from ω-3 oil (Dyets Inc., Bethlehem, PA, USA). At the end of the 6 weeks of FD supplementation, animals were anesthetized with isofluorane, and whole blood was collected from each mouse via retro-orbital venipuncture by heparinized microcapillaries. Mice were euthanized and organs removed immediately. The organs were divided into two and either frozen immediately in liquid nitrogen or fixed in 10% formalin and embedded in paraffin for histology. Whenever indicated, 6-week old mice were exposed to hypoxia (8% oxygen for 10 h) followed by 3 h of reoxygenation (21% oxygen) (H/R stress) to mimic an acute VOC, as previously described.19
Hematologic parameters and red cell indices
Hematologic parameters and red cell indices were measured as previously described.19–21 Details are reported in the Online Supplementary Materials and Methods.
Red cell fatty acid composition
Red cell membrane total lipids were extracted as previously reported22 and determined by gas liquid chromatography using a Hewlett Packard 6890 equipped with a flame ionization detector. Peaks were identified by comparison of retention times with external fatty acid methyl ester standard mixtures from NuCheck Prep (Elysian, MN, USA). The fatty acid profiles were expressed as percentage of the total fatty acid (weight percent).
Blood pressure and echocardiographic measurements
Heart rate, systolic, diastolic, and mean blood pressures were measured in conscious mice with a non-invasive computerized tail cuff system (CODA, Kent Scientific Corp) as previously reported.16 Transthoracic echocardiography was performed with a Vevo 2100 echocardiograph (Visual Sonics, Toronto, Canada) equipped with a 22–55 MHz transducer (MicroScan Transducers, MS500D) as previously described.16 Details are reported in the Online Supplementary Materials and Methods.
Histological analysis
Multiple (at least five) three-micron whole mount sections were obtained for each paraffin-embedded liver and stained with hematoxylin & eosin, Masson trichome, and May-Grünwald-Giemsa. The lung and liver pathology were analyzed and the liver pathological score, inflammatory cell infiltrate and presence of thrombi were determined by blinded pathologists as previously described.19,23 Details on the histological analysis are provided in the Online Supplementary Materials and Methods.
Tissue molecular analysis
Total RNA was extracted from mouse tissues using Trizol reagent (Life Technologies). cDNA was synthesized from total RNA (2 μg) using Super Script II First Strand kits (Life Technologies). Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) was performed using the SYBR-green method as previously reported.24 Primers used are reported in Online Supplementary Table S1. For the immunoblot analysis, frozen lungs, livers, and aortas from each studied group were homogenized and lysed with iced lysis buffer (LB containing: 150 mM NaCl, 25 mM bicine, 0.1% SDS, Triton 2%, EDTA 1 mM, protease inhibitor cocktail tablets (Roche), 1 mM Na3VO4 final concentration) then centrifuged at 12,000 for 10 min at 4°C g.25 Proteins were quantified and analyzed by one-dimensional sodium dodecylsulfate polyacrylamide gel electrophoresis. Gels were transferred to nitrocellulose membranes for immunoblot analysis with specific antibodies. Detailed information on the antibodies used is provided in the Online Supplementary Materials and Methods.
Bronchoalveolar lavage measurements
Bronchoalveolar lavage (BAL) fluids were collected and cellular contents were recovered by centrifugation and counted by microcytometry as previously reported.19,26 The percentage of neutrophils was determined on cytospin centrifugation. The remaining BAL samples were centrifuged at 1,500 × g for 10 min at 4°C. The supernatant fluids were used for determination of total protein content.19,26
Statistical analysis
The two-way ANOVA algorithm for repeated measures combined with Bonferroni correction was used for data analysis. Differences with P<0.005 were considered statistically significant.
Results
ω-3 polyunsaturated fatty acids modify red cell membrane composition and reduce neutrophil count in mice with sickle cell disease
We analyzed the lipid composition of red cell membranes from healthy (AA) and sickle cell (SS) mice fed with isocaloric diets containing either soy (SD) or fish-oil (FD) for 6 weeks. As shown in Figure 1A and Online Supplementary Table S2, red cell membrane ω-6 content was higher in SS-SD mice than in AA-SD mice, similar to that observed in human SCD patients.13 FD supplementation markedly increased ω-3 membrane content in both AA-FD and SS-FD mice, resulting in a dramatically lower ω-6/ω-3 ratio compared to that in both mouse strains fed SD (Figure 1A, Online Supplementary Table S2). In addition, in SS mice, FD resulted in significantly increased mean corpuscular volume and decreased mean cell hemoglobin concentration compared to control mice fed with SD, without significant changes in hemoglobin levels. No differences in hematologic parameters or red cell indices were evident in AA mice based on which of the two dietary interventions they received (Table 1). Interestingly, SS-SD mice showed increased neutrophil counts compared to AA-SD mice, as reported in other mouse models of SCD, a likely indication of elevated chronic systemic inflammation.27 As shown in Table 1, FD reduced the neutrophil count by 30.4±7.2% (n=6) and 64.6±15.1% (n=6) in AA mice and SS mice, respectively, compared to the counts in SD groups. Since inflammation plays an important role in the pathogenesis of vasculopathy in SCD,4 we evaluated the impact of FD on cardiovascular function in SCD mice.
Figure 1.

FD modified red cell membrane fatty acid composition and beneficially affects cardiovascular system in sickle cell mice. (A) Total ω-6 fatty acids (n-6) and ω-3 fatty acid/ω-6 fatty acid ratio (n3/n6 ratio) red cell membrane content as determined by mass spectroscopy in AA and SS mice under a soy-diet (SD) or fish-oil diet (FD). (B) Measurements of diastolic and systolic blood pressure (upper panel) in AA and SS mice under a SD or FD. (C) Immunoblot analysis with specific antibodies against endothelin-1 (ET-1), vascular adhesion molecule-1 (VCAM-1) and heme oxygenase-1 (HO-1) of isolated aortas from AA and SS mice under a SD or FD. One representative gel from six with similar results is presented. Densitometric analysis of anti- ET-1, VCAM-1 and HO-1 immunoblots is shown in Online Supplementary Figure S1A. (D) Upper panel. Left ventricular outflow tract (LVOT) diameter and left ventricular hypertrophy expressed as heart weight/body weight (HW/BW61) in AA and SS mice under a SD or FD. Lower panel. Immunoblot analysis with specific antibodies against ET-1, VCAM-1, and HO-1 of hearts from AA and SS mice under a SD or FD. One representative gel from six with similar results is presented. Densitometric analysis of immunoblots is shown in Online Supplementary Figure S1B.
Table 1.
Hematologic parameters in wild-type (AA) and sickle cell mice (SS) treated with ω−3 fatty acid supplementation.
ω-3 polyunsaturated fatty acids beneficially affect the cardiovascular system and endothelial activation in sickle cell mice
Vascular system
As shown in Figure 1B, FD treatment of SS mice resulted in a significantly lower diastolic blood pressure compared to that in either SS mice or AA mice fed SD. In order to understand the effects of FD on the vascular system in SCD better, we isolated aortas from AA and SS mice and evaluated the expression of endothelin-1 (ET-1) as a key vasoactive and pro-inflammatory cytokine,28 vascular adhesion molecule 1 (VCAM-1) as a marker of vascular endothelial activation,29 and heme-oxygenase-1 (HO-1) as an efficient anti-oxidant system interfacing heme overload and vascular inflammation in SCD.16,30 As expected, ET-1, VCAM-1 and HO-1 expression was increased in aortas from SS mice compared to that in AA mice fed the SD (Figure 1C, lanes 1 and 3). In SS mice, FD significantly reduced the expression of all three markers compared to their levels in SS control mice, reaching values similar to those observed in AA mice (Figure 1C, see also Online Supplementary Figure S1A for densitometric analysis).
Cardiac system
Echocardiographic measurements were carried out in both mouse strains fed either SD or FD diet. In SS-SD mice, we observed left ventricular hypertrophy, as measured by the ratio of heart weight to body weight, as well as a significant increase in left ventricular outflow tract diameter compared to that in AA-SD mice (Figure 1D). FD significantly reduced left ventricular outflow tract diameter in SS mice, without causing changes in the heart weight to body weight ratio (Figure 1D). No major differences were present in AA mice. Heart VCAM-1 and HO-1 expression was significantly higher in SS mice than in AA mice fed the SD (Figure 1D, lower panel). FD supplementation significantly reduced VCAM-1 and HO-1 in SS mice compared to levels in both SS and AA-SD groups (Figure 1D lower panel, Online Supplementary Figure S1B for densitometric analysis). No major differences were observed in ET-1 expression in either AA or SS mouse strains (Figure 1D, see also Online Supplementary Figure S1B for densitometric analysis).
These data suggest that in SS mice, FD reduces biochemical markers of vascular/oxidant damage and decreases evidence of ventricular loading.
ω-3 polyunsaturated fatty acids reduce pulmonary vascular permeability and endothelin-1 expression in mice with sickle cell disease
Abnormal vascular activation plays an important role in pulmonary complications of SCD.4,19 ω-3 PUFA has previously been shown to be protective in several models of lung injury.31,32 We evaluated pulmonary vascular leakage by measuring protein content and leukocyte counts in BAL fluid from AA and SS mice. As shown in Figure 2A, SS mice fed the SD had higher BAL protein and leukocyte levels compared to healthy mice, indicating accumulation of proteins and inflammatory cells in the alveolar space. FD significantly reduced both the leukocyte and protein content in the BAL fluid of the SS mice, suggesting an amelioration of vascular permeability compared to that in animals fed the SD (Figure 2A).
Figure 2.

FD ameliorates vascular dysfunction in lung and liver from SS mice. (A) Left panel. Bronchoalveolar (BAL) fluid protein content from AA and SS mice under a soy-diet (SD) or fish-oil diet (FD). Right panel. BAL fluid leukocyte content from AA and SS mice under a SD or FD. (B) Immunoblot analysis with specific antibodies against endothelin-1 (ET-1); vascular adhesion molecule-1 (VCAM-1) and heme-oxygenase-1 (HO-1) of lungs from AA and SS mice under a SD or FD. One representative gel from six with similar results is presented. Densitometric analysis of ET-1, VCAM-1 and HO-1 immunoblots is shown in Online Supplementary Figure S2A. (C) Immunoblot analysis with specific antibodies against ET-1, VCAM-1, and HO-1 of livers from AA and SS mice under a SD or FD. One representative gel from six with similar results is presented. Densitometric analysis of ET-1, VCAM-1 and HO-1 immunoblots is shown in Online Supplementary Figure S2B.
Since ET-1 plays a key role in clearance of alveolar fluid, local inflammatory response33 and in SCD vasculopathy,28,34 we evaluated the expression of ET-1 in lungs from SS and AA mice. Pulmonary ET-1 expression at both molecular and protein levels was significantly increased in SS-SD mice compared to that in AA-SD (Figure 2B, Online Supplementary Figure S2A for densitometric analysis). FD significantly reduced lung ET-1 expression in SS mice (Figure 2B, Online Supplementary Figures S2A and S3), in agreement with the observed improvement of pulmonary vascular leakage (Figure 2A). VCAM-1 levels were similar in both AA and SS mice, without modulation by ω-3 PUFA supplementation (Figure 2B, Online Supplementary Figure S2A). The increased pulmonary inflammatory state was associated with higher expression of HO-1 in SS mice than in AA mice, without significant changes in FD-treated groups (Figure 2B, Online Supplementary Figure S2B). These data suggest that ω-3 PUFA supplementation significantly reduces lung ET-1 expression, thereby resolving pulmonary vascular permeability and local inflammation in SCD mice.
ω-3 polyunsaturated fatty acids modulate hepatic inflammation and endothelin-1 expression in mice with sickle cell disease
Studies in models of liver injury have shown an important role of ET-1 on hepatic microvasculature, specifically related to local ischemic/reperfusion injury and inflammatory response.35,36 Although the liver is not a target organ for SCD, we have previously demonstrated that its functional anatomy, its high metabolic rate and the fine regulation of its microcirculation make the liver an interesting window-organ to explore the pathogenesis of SCD vasculopathy and the effects of new therapeutic tools.23 Here, we found increased inflammatory cell infiltrates in the liver portal space of SS mice compared to AA mice fed SD (data not shown). This was associated with increased expression of ET-1 at both the molecular and protein levels in livers from SS mice (Figure 2C, see also Online Supplementary Figure S2B for densitometric analysis and Online Supplementary Figure S3 for molecular analysis), suggesting a role of ET-1 as both a pro-inflammatory and a potent vasoconstrictor cytokine in the portal vascular system. The levels of VCAM-1 were significantly higher in livers from SS mice than AA mice, which argues for the presence of abnormal vascular activation and local inflammation.37 This is in agreement with the increased expression of HO-1 as a vaso-protective anti-oxidant.16,38 FD significantly reduced hepatic expression of ET-1 and HO-1 in SS mice compared to SS-SD mice, but it did not affect expression of VCAM-1 in the liver (Figure 2C, Online Supplementary Figure S2B).
Hypoxia/reoxygenation stress mimics acute sickle cell-related lung and liver damage
To evaluate the impact of FD on acute sickle cell-related events, we developed a model of acute VOC based on the exposure of mice to H/R stress.19,23,28 The H/R conditions were chosen based on previous models of acute VOC with related tissue injury and preliminary experiments to identify the time course of H/R that would recapitulate sickle cell acute organ damage in the present mouse model of SCD.19,23,28 We selected an experimental protocol of H/R in which AA and SS mice were exposed to hypoxia (8% oxygen) for 10 h followed by 3 h of re-oxygenation (21% oxygen, room air).19,23,28 In SS mice, H/R induced significant reductions in hematocrit and hemoglobin and a marked increase in hemoglobin distribution width, an index of the amount of dense red cells39 (Figure 3A,B). In H/R mice fed the SD we observed a significant increase in neutrophil count in both AA (SD: 257±23.2%; n=6) and SS mice (176.8±12%; n=6) compared to counts in normoxic mice, indicating a sustained systemic inflammatory response to H/R stress (Figure 3C). No major changes were observed in reticulocyte count comparing SS mice subjected to H/R stress or normoxic conditions (data not shown).
Figure 3.

FD prevents H/R-induced dense cell formation and increased neutrophil counts in SS mice. (A) Hematocrit (Hct), hemoglobin (Hb), and red cell distribution width (HDW) of AA and SS mice under a soy-diet (SD) or fish-oil diet (FD) under normoxia (white bars) and exposed to hypoxia (black bars) (8% oxygen; 10 h) followed by reoxygenation (21% oxygen; 3 h) (H/R). (B) Red cell distribution histograms generated for erythrocyte cell volume (RBC Volume) and cell hemoglobin concentration (RBC-HC). Red cell morphology is shown for each condition. (C) Neutrophil counts in AA and SS mice treated as in (A) under normoxia (white bars) and exposed to hypoxia (black bars) (8% oxygen; 10 h) followed by reoxygenation (21% oxygen; 3 h) (H/R). All statistical data are presented as means ± standard deviation (n=6; *P<0.005; **P<0.002).
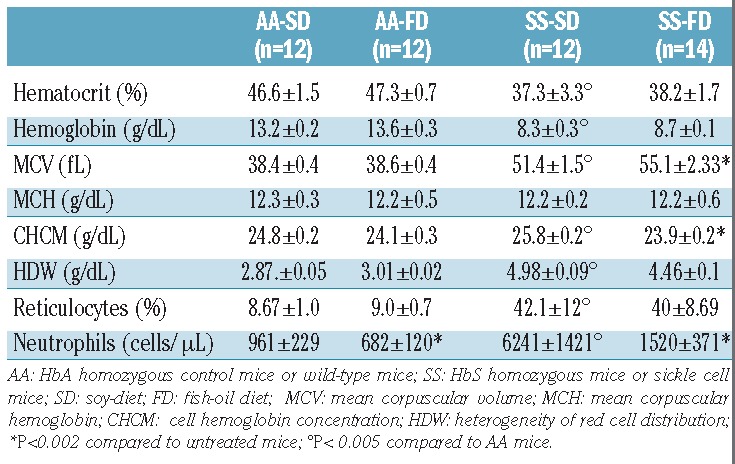
Histological analysis revealed that H/R stress induced a severe sickle cell-related tissue injury in both the lungs and livers of SS mice (Table 2; Figures 4 and 5). In the lung, H/R induced inflammatory cell infiltration, as well as bronchial mucus secretion and thrombi in 80% of the SS mice (Table 2). In AA mice, H/R induced modest inflammatory cell infiltration with mucus production, but without thrombi formation (Table 2). Livers from SS mice exposed to H/R demonstrated severe hepatic injury characterized by the loss of hepatic architecture, coagulative necrosis, inflammatory cell infiltrates and thrombi (Table 2). Livers from AA mice exposed to H/R demonstrated a modest inflammatory cell infiltrate with mild liver damage (Table 2).
Table 2.
Effects of ω−3 supplementation on lung and liver pathology of wild-type (AA) and sickle cell (SS) mice under normoxia and exposed to hypoxia/reoxygenation stress
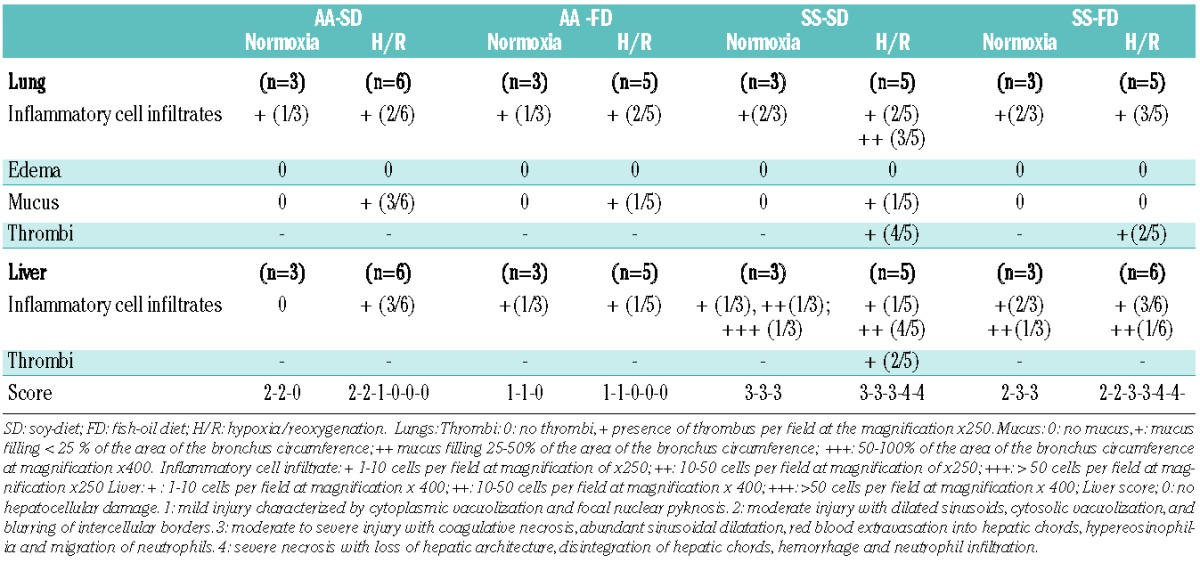
Figure 4.

FD reduces H/R-induced lung damage and prevents H/R-induced ET-1 up-regulation. (AD) Hematoxylin and eosin-stained sections of lung tissue at 400x magnification from SS mice under a SD (A and C) or supplemented with FD (B and D) exposed to hypoxia (8% oxygen; 10 h) followed by reoxygenation (21% oxygen; 3 h) (H/R). Lungs from SS mice given a FD have less inflammatory cellular infiltrate (B) and thrombi (D) than SS mice not fed FD (A and C). (E) Upper panel. BAL protein content from AA and SS mice treated as in (A–E). Lower panel. BAL leukocyte content from AA and SS mice treated as in (A–E). White bars show data from mice under normoxia and black bars show data from mice under H/R. (F) Immunoblot analysis with specific antibodies against ET-1, VCAM-1, and HO-1 of lung from AA and SS mice treated as in (A–E). One representative gel from six with similar results is presented. Densitometric analysis of ET-1, VCAM-1 and HO-1 immunoblots is shown in Online Supplementary Figure S4B.
Figure 5.
FD reduces H/R-induced liver damage and vascular activation. (A–D) Hematoxylin and eosin-stained sections of liver tissue at 400x magnification from SS mice under a SD (A and C) or FD supplementation (B and D) exposed to H/R. Livers from mice given FD have less inflammatory cellular infiltrate (B) and thrombi (D) than livers from SS mice fed a SD. The infiltrate shown best in (B) is composed mostly of lymphocytes. Scattered hemosiderin deposits and areas of necrosis are also present. (E) Immunoblot analysis with specific antibodies against ET-1, VCAM-1, HO-1, and heat shock protein-27 (HSP27) of liver from AA and SS mice treated as in (A–D). One representative gel from six with similar results is presented. Densitometric analysis of ET-1, VCAM-1, HO-1 and HSP27 immunoblots is shown in Online Supplementary Figure S5.
ω-3 polyunsaturated fatty acids prevent hypoxia/reoxygenation-induced formation of dense red cells and neutrophilic response
In SS mice, FD: (i) significantly mitigated the H/R-induced reductions in hematocrit and hemoglobin Hb levels; and (ii) significantly reduced the fraction of H/R-generated dense red cells, as supported by the decrease in high distribution width of red cells in these animals compared with SS mice (Figure 3A,B). In addition, we observed a significant reduction in the H/R-induced increased neutrophil count in both AA-FD (41.9±3.7%; n=6) and SS-FD (46.6%±23%; n=6) mice compared to the SD groups of mice (Figure 3C). These findings suggest that ω-3 PUFA supplementation causes a shift towards an anti-inflammatory phenotype in SS mice during acute VOC.
Hypoxia/reoxygenation-induced sickle cell lung injury is attenuated by ω-3 polyunsaturated fatty acids via the endothelin-1 pathway
The lungs of SS-FD mice exposed to H/R stress showed reduced inflammatory cell infiltrates (Figure 4B, Table 2), no mucus filling the bronchi, and fewer thrombotic events (Figure 4D, Table 2) compared to those of SS-SD mice (Table 2). Both the protein content and leukocyte counts of BAL fluid were significantly reduced in SS-FD mice compared to SS-SD animals (Figure 4E). A reduction in BAL fluid protein content was also observed in AA-FD mice exposed to H/R stress (Figure 4E). These findings indicate a protective role of ω-3 PUFA against the H/R-induced pulmonary vascular permeability and local inflammatory cellular response, suggesting a possible modulation of the H/R-induced ET-1 pathway.28
In SS mice, FD promoted a significant reduction in H/R-induced ET-1 molecular and protein expression compared to levels in SS-SD mice (Figure 4F, see also Online Supplementary Figure S4A,B for densitometric analysis), which was paralleled by a decrease in endothelin-1B receptor (ETBR) expression (Online Supplementary Figure S5A). The ability of FD to blunt the cytokine storm in H/R stress was also supported by significant reductions in interleukin-1β and interleukin-10 expression in the lungs of SS-FD mice compared to SS-SD ones (Online Supplementary Figure S5B). VCAM-1 expression was increased in H/R SS mice compared with AA mice, with no significant differences between FD- and SD-fed SS mice (Figure 4F, see also Online Supplementary Figure S4B for densitometric analysis). The absence of effects of FD on H/R-induced VCAM-1 lung expression in SS mice is not surprising given that H/R stress promotes a redistribution of extracellular VCAM-1 towards the alveolar compartment, as shown in asthma models.40 This is also supported by the significant down-regulation of lung VCAM-1 gene expression in SS-FD and AA-FD mice compared to SD groups (Online Supplementary Figure S4A). Thus, VCAM-1 lung expression reflects only one of the functional components of VCAM-1 involved in H/R lung injury. H/R induced a significant up-regulation of HO-1 in SS mice compared to AA mice, which was prevented in SS-FD mice (Figure 4F, Online Supplementary Figure S4B). These data suggest that FD efficiently protects SS mice from acute H/R lung injury through the reduction of ET-1 expression and the modulation of the inflammatory response, as supported by the reduction in inflammatory cell infiltrates, BAL fluid leukocyte content, and the decreased expression of HO-1.
ω-3 polyunsaturated fatty acids blunt the hypoxia/reoxygenation-induced inflammatory response and decrease vascular activation in the livers of sickle cell mice
The histological analysis of livers from SCD mice exposed to H/R stress showed that FD markedly reduced inflammatory cellular infiltrates and thrombi formation compared to the SD (Figure 5B,D; Table 2). This was in agreement with a reduced pathologic severity score in H/R-exposed SS-FD mice compared with SS-SD mice (Table 2). Serum levels of alananine and aspartate transaminases were significantly lower in both AA-FD and SS-FD mice than in the groups fed the SD (Online Supplementary Table S3). No significant changes in liver ET-1 expression were observed in SS-SD mice compared to SS-FD mice (Figure 5E). We found a marked increase in VCAM-1 expression in SS mice, compared to AA mice, in response to H/R stress (Figure 5E, see also Online Supplementary Figure S6A for densitometric analysis). FD reduced the H/R-related VCAM-1 up-regulation compared to that in SS-SD mice (Figure 5). HO-1 expression in response to H/R was higher in livers of SS mice than in AA mice, but it was unaffected by ω-3 PUFA supplementation (SS-FD and AA-FD; Figure 5). Interestingly, HO-1 mRNA levels were down-regulated in SS-FD mice compared to SS-SD mice (Online Supplementary Figure S6B). Since HO-1 also plays a role as a heat shock protein (HSP32) in other animal models, such as the rat,41 we evaluated the hepatic expression of HSP27, which is known to be important in H/R liver injury.23,42 FD attenuated the H/R-induced HSP27 expression in SS mice compared to SS-SD animals (Figure 5E, Online Supplementary Figure S6B), supporting the anti-inflammatory effects of ω-3 PUFA following H/R stress. These findings align with data from other models of H/R liver injury35,43 and confirm that VCAM-1 mediates adhesion events in the hepatic microvasculature and contributes to amplified inflammatory liver disease in SCD mice. In livers from SS mice, FD prevented vascular activation and reduced inflammation, improving endothelial dysfunction and reducing organ damage.
Vascular protective effects of ω-3 polyunsaturated fatty acids during acute vaso-occlusive crises in sickle cell mice
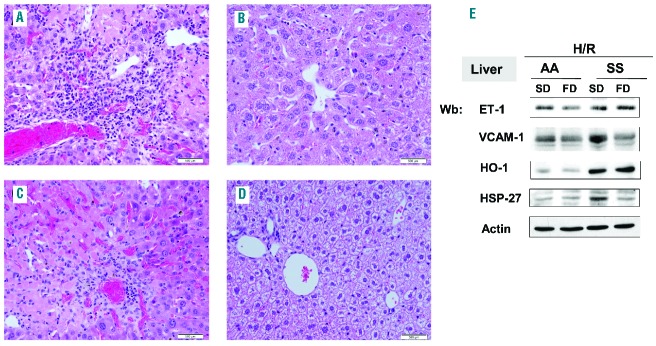
We isolated aortic tissue from AA and SS mice exposed to H/R stress. As shown in Figure 6A, in SS mice, FD reduced H/R-induced ET-1 and VCAM-1 expression compared to SS-SD. This finding was associated with reduced expression of acute inflammatory response-related proteins such as HO-1, SOD-1 and peroxiredoxin-2 (Prx2) (Figure 6A, see also Online Supplementary Figure S7 for densitometric analysis). These effects were also evident in AA-FD mice exposed to H/R compared to AA-SD mice (Figure 6A). These results support the hypothesis that FD attenuates endothelial dysfunction in SCD by reducing vascular activation, vasoconstriction, inflammation and oxidative stress.
Figure 6.
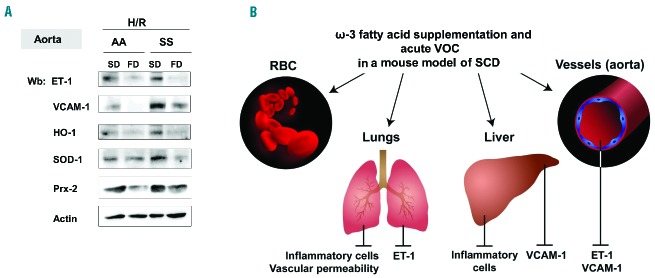
FD prevents H/R-induced ET-1 expression, vascular activation and modulates anti-oxidant systems. (A) Immunoblot analysis with specific antibodies against ET-1, VCAM-1, HO-1, SOD-1, and peroxiredoxin-2 (Prx-2) of isolated aortas from AA and SS under a SD or FD and exposed to H/R. One representative gel from six with similar results is presented. Densitometric analysis of ET-1, VCAM-1 and HO-1 immunoblots is shown in Online Supplementary Figure S7. (B) Schematic diagram of effects of ω-3 fatty supplementation on acute vaso-occlusive crises (VOC) in a mouse model of SCD. ω-3 fatty supplementation (i) beneficially affects sickle red cell membrane composition, with a shift towards an anti-inflammatory substrate (ii) has potent anti-inflammatory action in target organs for SCD such as lung and liver; (iii) improves vascular dysfunction through either a reduction of ET-1 (lung) or VCAM-1 (liver) or the synergizing decrease of both molecules in aorta.
Discussion
The effects of ω-3 fatty acid supplementation have been widely studied in cardiovascular diseases and in other disorders characterized by acute or chronic vascular damage and an amplified inflammatory response.44,45 In SCD, vasculopathy, inflammation and intravascular oxidative stress are crucial in the pathogenesis of acute and chronic clinical complications. Recent reports on the beneficial effects of ω-3 fatty acid supplementation on the human hematologic phenotype of SCD suggest a possible therapeutic effect of ω-3 fatty acids in SCD.12–14 The present work provides evidence for several relevant mechanisms underlying this potential benefit. We propose that the beneficial role of ω-3 PUFA on SCD vascular dysfunction might be exerted: (i) directly through the reduction of ET-1 expression, which is involved in adaptive inflammation, expression of adhesion molecules such as VCAM-1, and neutrophil chemotaxis;46 and (ii) indirectly through reduced inflammation and oxidative stress. Importantly, the ω-3 PUFA supplementation in our study is comparable to that achieved in recent human studies.47
In SS mice, ω-3 PUFA consistently modulate ET-1, a potent vasoconstrictor, in agreement with recent meta-analyses demonstrating a significant improvement in flow-mediated dilation in subjects supplemented with FD.45,48 This is of particular interest in the context of SCD, in which ET-1 has been implicated in severe clinical complications of SCD.28,49 The positive impact of FD on ET-1 is not limited to the aorta but also involves the pulmonary vascular bed, where we observed a decrease in the abnormal pulmonary vascular leakage and a decrease in the accumulation of protein and inflammatory cells in the alveolar space (Figure 2A,B).
To explore the critical balance between vascular tone and endothelial activation in SCD, we used a H/R stress model, which mimics the acute organ damage observed in SCD VOC (Figures 4 and 5; Table 2).19,50 Compared to SD, FD significantly reduced H/R-related ET-1 expression in the aorta (Figure 6). This was accompanied by a significant reduction in both ET-1 and ET-1BR expression in lungs from SS-FD mice (Fig. 4F, Online Supplementary 5SA), suggesting that the beneficial effects of FD treatment on H/R-induced pulmonary vascular permeability might be related to the combination of both ligand and target reduction in the ET-1 system.33,51 No significant changes in ET-1 expression were observed between livers from H/R-exposed SS-FD and SS-SD mice (Figure 5), most likely because of the peculiar organization of liver vasculature or because of the organ’s sensitivity in the context of H/R stress used in the present study.
We then evaluated the effects of FD on VCAM-1 expression, since elevated VCAM-1 has been associated with a severe SCD phenotype and increased risk of early death.49,52,53 In conditions of normoxia, FD significantly reduced VCAM-1 expression in aortas and hearts of SS mice compared to SS-SD mice (Figure 1B,C). This was associated with improved left ventricular outflow tract dimensions, suggesting a possible contribution of FD in delaying the progression of SCD cardiovascular disease.54–56 We demonstrated a synergistic effect of ω-3 PUFA on both ET-1-mediated endothelial dysfunction and VCAM-1-mediated endothelial activation on aortas following H/R (Figure 6A). Notably, FD also significantly reduced VCAM-1 expression in livers of SS mice exposed to H/R stress compared to the expression in SS-SD mice (Figure 5). These data support a multimodal therapeutic action of FD on both arterial and microvascular sites, preventing vascular activation and possible amplification of local pro-inflammatory events during acute VOC.
HO-1 is a key cytoprotector interfacing between vascular dysfunction and oxidative stress in SCD.23,30 Under normoxia, ω-3 PUFA significantly reduced HO-1 expression in the aorta, heart and liver of SS mice (Figure 1B and 2C). Interestingly, FD did not affect lung HO-1 expression in SS mice, even though a pulmonary anti-inflammatory effect for FD supplementation was observed (i.e. reduced neutrophil counts in BAL fluid and reduced pulmonary cell infiltrates) (Figures 2 and 4). This suggests that, under steady state conditions, the anti-inflammatory effects of FD on lungs of SCD mice may be more related to down-regulation of ET-1. During acute VOC mimicked by H/R stress, FD reduced HO-1 expression, indicating a stronger role of ω-3 PUFA in acute events involving the lung than in the steady state condition. This is in agreement with recent studies on the protective role of ω-3-derived pro-resolving mediators in models of acute lung injury.57,58 The pro-resolving effects of FD supplementation during H/R stress were also evident in aortas of SS-FD mice, in which we observed a marked reduction in H/R-induced HO-1 expression and prevention of the H/R-induced expression of antioxidant systems such as SOD-1 and Prx2 (Figure 6A).
In livers from SCD mice exposed to H/R stress, FD was associated with HO-1 mRNA down-regulation and a marked decrease in the expression of HSP27, which is a crucial liver cytoprotector against H/R stress.42 This finding supports the anti-inflammatory activity of FD in reducing liver injury during H/R stress in SCD mice.
Finally, we observed a reduction in dense red cell formation in SS-FD mice exposed to H/R compared to SS-SD mice, which may contribute to the multimodal action of ω-3 PUFA on SCD vasculopathy. Changes in membrane lipid composition might affect red cell deformability, as recently shown in a mouse model supplemented with ω-3 PUFA (docosahexaenoic acid),59 as well as variations in pro-inflammatory arachidonic acid incorporated into sickle erythrocytes.60 These red cell changes may play a role in reducing the systemic pro-inflammatory environment, which contributes to the pathogenesis of vasculopathy in SCD.6
In summary, we propose that ω-3 PUFA may limit sickle cell-related acute damage by: (i) shifting the sickle red cell membrane towards an anti-inflammatory substrate; (ii) exerting local anti-inflammatory effects in target organs for SCD (lung, liver); (iii) improving vascular dysfunction through the reduction of VCAM-1 or ET-1 or the synergistic decrease of both molecules. This functional model supports the preliminary results of a clinical trial with ω-3 PUFA supplementation in SCD patients, which showed a reduction in the rate and severity of painful crises.13 Taken together, our data add significant new elements to the mechanism of action of ω-3 PUFA in SCD, and generate a rationale for future studies focused on the bioactive metabolites of ω-3 PUFA.
Footnotes
The online version of this article has a Supplementary Appendix.
Funding
This work was supported by PRIN (LDF: 201228PNX83_003) and FUR_UNIVR (LDF). The authors would like to thank Amy Pan for her help in isolating RBC fatty acids for membrane analysis. The authors would also like to thank Marykate O’Malley and Kristin Johnson for their help with the figures.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Bunn HF. Pathogenesis and treatment of sickle cell disease. N Engl J Med. 1997;337(11):762–769. [DOI] [PubMed] [Google Scholar]
- 2.Sparkenbaugh E, Pawlinski R. Interplay between coagulation and vascular inflammation in sickle cell disease. Br J Haematol. 2013;162(1):3–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.De Franceschi L, Cappellini MD, Olivieri O. Thrombosis and sickle cell disease. Semin Thromb Hemost. 2011;37(3):226–236. [DOI] [PubMed] [Google Scholar]
- 4.Hebbel RP, Vercellotti G, Nath KA. A systems biology consideration of the vasculopathy of sickle cell anemia: the need for multi-modality chemo-prophylaxsis. Cardiovasc Hematol Disord Drug Targets. 2009;9(4):271–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Massaro M, Scoditti E, Carluccio MA, De Caterina R. Basic mechanisms behind the effects of n-3 fatty acids on cardiovascular disease. Prostaglandins Leukot Essent Fatty Acids. 2008;79(3–5):109–115. [DOI] [PubMed] [Google Scholar]
- 6.Calder PC. Omega-3 fatty acids and inflammatory processes. Nutrients. 2010;2(3):355–374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rangel-Huerta OD, Aguilera CM, Mesa MD, Gil A. Omega-3 long-chain polyunsaturated fatty acids supplementation on inflammatory biomakers: a systematic review of randomised clinical trials. Br J Nutr. 2012;107(Suppl 2):S159–170. [DOI] [PubMed] [Google Scholar]
- 8.Wachira JK, Larson MK, Harris WS. n-3 Fatty acids affect haemostasis but do not increase the risk of bleeding: clinical observations and mechanistic insights. Br J Nutr. 2014;111(9):1652–1662. [DOI] [PubMed] [Google Scholar]
- 9.Chen J, Shearer GC, Chen Q, et al. Omega-3 fatty acids prevent pressure overload-induced cardiac fibrosis through activation of cyclic GMP/protein kinase G signaling in cardiac fibroblasts. Circulation. 2011;123(6):584–593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ren H, Ghebremeskel K, Okpala I, et al. Abnormality of erythrocyte membrane n-3 long chain polyunsaturated fatty acids in sickle cell haemoglobin C (HbSC) disease is not as remarkable as in sickle cell anaemia (HbSS). Prostaglandins Leukot Essent Fatty Acids. 2006;74(1):1–6. [DOI] [PubMed] [Google Scholar]
- 11.Ren H, Obike I, Okpala I, et al. Steady-state haemoglobin level in sickle cell anaemia increases with an increase in erythrocyte membrane n-3 fatty acids. Prostaglandins Leukot Essent Fatty Acids. 2005;72(6):415–421. [DOI] [PubMed] [Google Scholar]
- 12.Okpala I, Ibegbulam O, Duru A, et al. Pilot study of omega-3 fatty acid supplements in sickle cell disease. APMIS. 2011;119(7):442–448. [DOI] [PubMed] [Google Scholar]
- 13.Daak AA, Ghebremeskel K, Hassan Z, et al. Effect of omega-3 (n-3) fatty acid supplementation in patients with sickle cell anemia: randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2013;97(1):37–44. [DOI] [PubMed] [Google Scholar]
- 14.Tomer A, Kasey S, Connor WE, et al. Reduction of pain episodes and prothrombotic activity in sickle cell disease by dietary n-3 fatty acids. Thromb Haemost. 2001; 85(6):966–974. [PubMed] [Google Scholar]
- 15.Simopoulos AP. Omega-3 fatty acids in health and disease and in growth and development. Am J Clin Nutr. 1991;54(3):438–463. [DOI] [PubMed] [Google Scholar]
- 16.Vinchi F, De Franceschi L, Ghigo A, et al. Hemopexin therapy improves cardiovascular function by preventing heme-induced endothelial toxicity in mouse models of hemolytic diseases. Circulation. 2013;127(12):1317–1329. [DOI] [PubMed] [Google Scholar]
- 17.Wu LC, Sun CW, Ryan TM, et al. Correction of sickle cell disease by homologous recombination in embryonic stem cells. Blood. 2006;108(4):1183–1188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Reeves PG, Nielsen FH, Fahey GC., Jr AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr. 1993;123(11):1939–1951. [DOI] [PubMed] [Google Scholar]
- 19.de Franceschi L, Baron A, Scarpa A, et al. Inhaled nitric oxide protects transgenic SAD mice from sickle cell disease-specific lung injury induced by hypoxia/reoxygenation. Blood. 2003;102(3):1087–1096. [DOI] [PubMed] [Google Scholar]
- 20.Stocker JW, De Franceschi L, McNaughton-Smith GA, et al. ICA-17043, a novel Gardos channel blocker, prevents sickled red blood cell dehydration in vitro and in vivo in SAD mice. Blood. 2003;101(6):2412–2418. [DOI] [PubMed] [Google Scholar]
- 21.De Franceschi L, Saadane N, Trudel M, et al. Treatment with oral clotrimazole blocks Ca(2+)-activated K+ transport and reverses erythrocyte dehydration in transgenic SAD mice. A model for therapy of sickle cell disease. J Clin Invest. 1994;93(4):1670–1676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 1957;226(1):497–509. [PubMed] [Google Scholar]
- 23.Siciliano A, Malpeli G, Platt OS, et al. Abnormal modulation of cell protective systems in response to ischemic/reperfusion injury is important in the development of mouse sickle cell hepatopathy. Haematologica. 2011;96(1):24–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Andolfo I, Alper SL, De Franceschi L, et al. Multiple clinical forms of dehydrated hereditary stomatocytosis arise from mutations in PIEZO1. Blood. 2013;121(19):3925–3935. [DOI] [PubMed] [Google Scholar]
- 25.De Franceschi L, Platt OS, Malpeli G, et al. Protective effects of phosphodiesterase-4 (PDE-4) inhibition in the early phase of pulmonary arterial hypertension in transgenic sickle cell mice. FASEB J. 2008;22(6):1849–1860. [DOI] [PubMed] [Google Scholar]
- 26.de Franceschi L, Malpeli G, Scarpa A, et al. Protective effects of S-nitrosoalbumin on lung injury induced by hypoxia-reoxygenation in mouse model of sickle cell disease. Am J Physiol Lung Cell Mol Physiol. 2006;291(3):457–465. [DOI] [PubMed] [Google Scholar]
- 27.Kaul DK, Hebbel RP. Hypoxia/reoxygenation causes inflammatory response in transgenic sickle mice but not in normal mice. J Clin Invest. 2000;106(3):411–420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sabaa N, de Franceschi L, Bonnin P, et al. Endothelin receptor antagonism prevents hypoxia-induced mortality and morbidity in a mouse model of sickle-cell disease. J Clin Invest. 2008;118(5):1924–1933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Osborn L, Hession C, Tizard R, et al. Direct expression cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lymphocytes. Cell. 1989;59(6):1203–1211. [DOI] [PubMed] [Google Scholar]
- 30.Belcher JD, Mahaseth H, Welch TE, et al. Heme oxygenase-1 is a modulator of inflammation and vaso-occlusion in transgenic sickle mice. J Clin Invest. 2006;116(3):808–816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sharma S, Chhibber S, Mohan H, Sharma S. Dietary supplementation with omega-3 polyunsaturated fatty acids ameliorates acute pneumonia induced by Klebsiella pneumoniae in BALB/c mice. Can J Microbiol. 2013;59(7):503–510. [DOI] [PubMed] [Google Scholar]
- 32.Umar S, Nadadur RD, Li J, et al. Intralipid prevents and rescues fatal pulmonary arterial hypertension and right ventricular failure in rats. Hypertension. 2011;58(3):512–518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Berger MM, Rozendal CS, Schieber C, et al. The effect of endothelin-1 on alveolar fluid clearance and pulmonary edema formation in the rat. Anest Analg. 2009;108(1):225–231. [DOI] [PubMed] [Google Scholar]
- 34.Rybicki AC, Benjamin LJ. Increased levels of endothelin-1 in plasma of sickle cell anemia patients. Blood. 1998;92(7):2594–2596. [PubMed] [Google Scholar]
- 35.Kitagawa T, Yokoyama Y, Kokuryo T, Nagino M. Protective effects of branched-chain amino acids on hepatic ischemiareperfusion-induced liver injury in rats: a direct attenuation of Küpffer cell activation. Am J Physiol Gastrointest Liver Physiol. 2013;304(4):346–355. [DOI] [PubMed] [Google Scholar]
- 36.Shetty S, Lalor PF, Adams DH. Lymphocyte recruitment to the liver: molecular insights into the pathogenesis of liver injury and hepatitis. Toxicology. 2008;254(3):136–146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Volpes R, Van Den Oord JJ, Desmet VJ. Vascular adhesion molecules in acute and chronic liver inflammation. Hepatology. 1992;15(2):269–275. [DOI] [PubMed] [Google Scholar]
- 38.Vercellotti GM, Khan FB, Nguyen J, et al. H-ferritin ferroxidase induces cytoprotective pathways and inhibits microvascular stasis in transgenic sickle mice. Front Pharmacol. 2014;5:79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Billett HH, Fabry ME, Nagel RL. Hemoglobin distribution width: a rapid assessment of dense red cells in the steady state and during painful crisis in sickle cell anemia. J Lab Clin Med. 1988;112(3):339–344. [PubMed] [Google Scholar]
- 40.Chihara J, Yamamoto T, Kurachi D, Nakajima S. Soluble ICAM-1 in sputum of patients with bronchial asthma. Lancet. 1994;343(8905):1108. [DOI] [PubMed] [Google Scholar]
- 41.Shibahara S, Muller RM, Taguchi H. Transcriptional control of rat heme oxygenase by heat shock. J Biol Chem. 1987;262(27):12889–12892. [PubMed] [Google Scholar]
- 42.Chen SW, Park SW, Kim M, et al. Human heat shock protein 27 overexpressing mice are protected against hepatic ischemia and reperfusion injury. Transplantation. 2009;87(10):1478–1487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hafez T, Moussa M, Nesim I, et al. The effect of intraportal prostaglandin E1 on adhesion molecule expression, inflammatory modulator function, and histology in canine hepatic ischemia/reperfusion injury. J Surg Res. 2007;138(1):88–99. [DOI] [PubMed] [Google Scholar]
- 44.De Caterina R. n-3 fatty acids in cardiovascular disease. N Engl J Med. 2011;364(25): 2439–2450. [DOI] [PubMed] [Google Scholar]
- 45.Wang Q, Liang X, Wang L, et al. Effect of omega-3 fatty acids supplementation on endothelial function: a meta-analysis of randomized controlled trials. Atherosclerosis. 2012;221(2):536–543. [DOI] [PubMed] [Google Scholar]
- 46.Sampaio AL, Rae GA, Henriques MG. Participation of endogenous endothelins in delayed eosinophil and neutrophil recruitment in mouse pleurisy. Inflamm Res. 2000;49(4):170–176. [DOI] [PubMed] [Google Scholar]
- 47.Scorletti E, Bhatia L, McCormick KG, et al. Effects of purified eicosapentaenoic and docosahexaenoic acids in non-alcoholic fatty liver disease: results from the *WELCOME study. Hepatology. 2014;60(4):1211–1221. [DOI] [PubMed] [Google Scholar]
- 48.Xin W, Wei W, Li X. Effect of fish oil supplementation on fasting vascular endothelial function in humans: a meta-analysis of randomized controlled trials. PloS One. 2012;7(9):e46028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hatzipantelis ES, Pana ZD, Gombakis N, et al. Endothelial activation and inflammation biomarkers in children and adolescents with sickle cell disease. Int J Hematol. 2013;98(2):158–163. [DOI] [PubMed] [Google Scholar]
- 50.Belcher JD, Mahaseth H, Welch TE, et al. Critical role of endothelial cell activation in hypoxia-induced vasoocclusion in transgenic sickle mice. Am J Physiol Heart Circ Physiol. 2005;288(6):H2715–2725. [DOI] [PubMed] [Google Scholar]
- 51.Ott J, Hiesgen C, Mayer K. Lipids in critical care medicine. Prostaglandins Leukot Essent Fatty Acids. 2011;85(5):267–273. [DOI] [PubMed] [Google Scholar]
- 52.Kato GJ, Martyr S, Blackwelder WC, et al. Levels of soluble endothelium-derived adhesion molecules in patients with sickle cell disease are associated with pulmonary hypertension, organ dysfunction, and mortality. Br J Haematol. 2005;130(6):943–953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Elmariah H, Garrett ME, De Castro LM, et al. Factors associated with survival in a contemporary adult sickle cell disease cohort. Am J Hematol. 2014;89(5):530–535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Desai AA, Patel AR, Ahmad H, et al. Mechanistic insights and characterization of sickle cell disease-associated cardiomyopathy. Circ Cardiovasc Imaging. 2014;7(3):430–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Junqueira FP, Fernandes JL, Cunha GM, et al. Right and left ventricular function and myocardial scarring in adult patients with sickle cell disease: a comprehensive magnetic resonance assessment of hepatic and myocardial iron overload. J Cardiovasc Magn Reson. 2013;15:83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dimitrow PP, Undas A, Wolkow P, Tracz W, Dubiel JS. Enhanced oxidative stress in hypertrophic cardiomyopathy. Pharmacol Rep. 2009;61(3):491–495. [DOI] [PubMed] [Google Scholar]
- 57.Wang Q, Zheng X, Cheng Y, et al. Resolvin D1 stimulates alveolar fluid clearance through alveolar epithelial sodium channel, Na,K-ATPase via ALX/cAMP/PI3K pathway in lipopolysaccharide-induced acute lung injury. J Immunol. 2014;192(8):3765–3777. [DOI] [PubMed] [Google Scholar]
- 58.El Kebir D, Gjorstrup P, Filep JG. Resolvin E1 promotes phagocytosis-induced neutrophil apoptosis and accelerates resolution of pulmonary inflammation. Proc Nat Acad Sci USA. 2012;109(37):14983–14988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wandersee NJ HM,, Maciaszek JL, Larson MC, et al. Dietary supplememntation with Docosahexanoic acid (DHA) improves RBC flexibility and reduces cold hypersensitivity in mice with sickle cell disease. ASH; 2012; 2012:2116. [Google Scholar]
- 60.Setty BN, Chen D, Stuart MJ. Sickle red blood cells stimulate endothelial cell production of eicosanoids and diacylglycerol. J Lab Clin Med. 1996;128(3):313–321. [DOI] [PubMed] [Google Scholar]
- 61.Ha T, Li Y, Hua F, Ma J, Gao X, Kelley J, et al. Reduced cardiac hypertrophy in toll-like receptor 4-deficient mice following pressure overload. Cardiovasc Res. 2005;68(2):224–234. [DOI] [PubMed] [Google Scholar]