

There is a growing interest in the role of reactive oxygen species (ROS) in chronic lymphocytic leukemia (CLL) in which tumor cells accumulate higher levels of such species than do normal B cells.1–3 As in other cells, mitochondria appear to be the main source of ROS and CLL cells have an increased mitochondrial mass compared to that of normal B cells.2,4 Higher levels of ROS confer increased sensitivity to induction of apoptosis by agents which further enhance ROS and it may be possible to exploit this as the basis for new treatments.1,2,5
Although ROS levels are generally higher in CLL cells than in normal B cells, the levels vary considerably between patients’ samples.1–3,5,6 Analyses have mainly been performed on samples from patients with more advanced disease and, in that setting, prior therapy appeared to be a major determinant of ROS levels.3,5,6 This may reflect a direct effect of drugs on ROS production, perhaps linked to accumulation of mitochondrial DNA damage.6 However, a recent study demonstrated variable ROS levels in cells from untreated patients2 indicating an influence of other factors.
The B-cell receptor (BCR) is now recognized as a key determinant of variable behavior of CLL7 and is a target for therapeutic attack. Antigen engagement appears to be iterative, with the outcome being either proliferation or anergy, a balance likely to influence disease outcome.7,8 Anergy is characterized by strong down-modulation of both expression and function of sIgM, and is evident in all CLL but predominates in cases expressing mutated IGHV genes (M-CLL) and is associated with a good prognosis. By contrast, growth-promoting BCR signaling is more evident in cases expressing unmutated IGHV genes (U-CLL) and is associated with progressive disease. In this work, we investigated potential links between ROS, anergy and clinical progression in CLL.
We used the ROS-sensitive fluorescent probe CM-H2DCFDA to quantify ROS in peripheral blood mononuclear cells from 33 patients (Figure 1Ai and Online Supplementary Table S1). ROS measurements were made on singlet, viable cells (based on forward and side scatter in flow cytometry analysis). We focused mainly (27/33) on untreated patients to avoid perturbing effects of chemotherapy, and the majority (27/33) had stage A disease. Samples generally contained a high proportion of CD19+CD5+ CLL cells (median 93%) to reduce contamination by non-malignant cells. Moreover, combined staining with CM-H2DCFDA and anti-CD19 and anti-CD5 antibodies in a subset of randomly selected samples demonstrated that quantification of ROS in total peripheral blood mononuclear cells and CD19+CD5+ cells yielded very similar results, including in sample 520A which contained ~80% CLL cells (Figure 1Aii). Repeat testing of samples on separate occasions confirmed high reproducibility of CM-H2DCFDA measurements (data not shown) and staining with a mitochondrial ROS-specific probe confirmed that CLL cells produced substantial levels of mitochondria-derived ROS (Online Supplementary Figure S1).2
Figure 1.
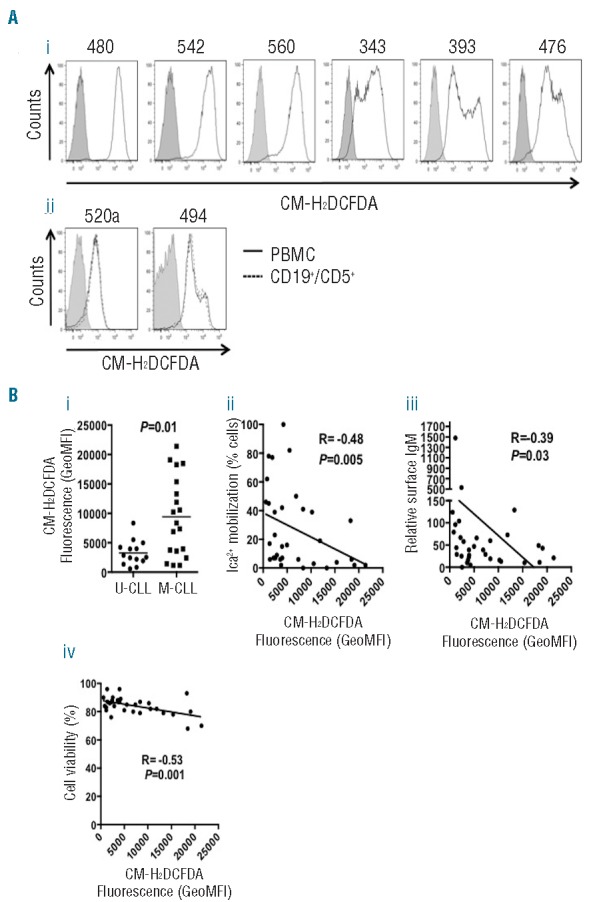
Inter-patient variability in ROS levels. (Ai) CLL peripheral blood monocuclear cells (PBMC) were recovered from cryopreservation, rested for 3 h and then stained with CM-H2DCFDA for 1 h at 37°C prior to flow cytometric analysis. Representative results for six of 33 samples analyzed. Shaded areas represents unlabeled, control cells. Open areas represents CM-H2DCFDA-labeled cells. (Aii) As (Ai), except a proportion of the sample was additionally incubated with anti-CD19 and anti-CD5 antibodies prior to flow cytometry. (B) Correlations between mean CM-H2DCFDA fluorescence (GeoMFI) in CLL PBMCs and (i) IGHV mutation status; (ii) anti-IgM-induced calcium; (iii) sIgM expression and (iv) cell viability. Results of statistical comparisons are shown. (i) Mann-Whitney test; (ii–iv) Spearman correlation.
ROS levels were highly variable between individual samples (Figure 1B and Online Supplementary Table S1). We first compared ROS levels with IGHV status and detected relatively low but consistent expression within U-CLL, but significantly higher and more heterogeneous expression in M-CLL (Figure 1Bi). There was also a significant negative association with sIgM signaling capacity, pointing to a correlation with anergy which predominates in M-CLL (Figure 1Bii).8 There was also an inverse correlation between ROS levels and sIgM expression (Figure 1Biii). Cell viability was generally high (median 84%) in the samples. However, there was a significant inverse correlation between ROS levels and cell viability (Figure 1Biv).
We also investigated potential correlations between ROS levels and disease behavior using Kaplan-Meier analysis. Receiver operating characteristic (ROC) analysis was used to segregate samples into two groups based on ROS levels and correlations with time-to-first treatment (TTFT). Using a ROC-derived cut-off of 2483, high ROS levels were associated with a significantly longer TTFT (Figure 2A). As expected, expression of mutated IGHV genes (M-CLL) was also associated with a significantly longer TTFT (Figure 2B).
Figure 2.
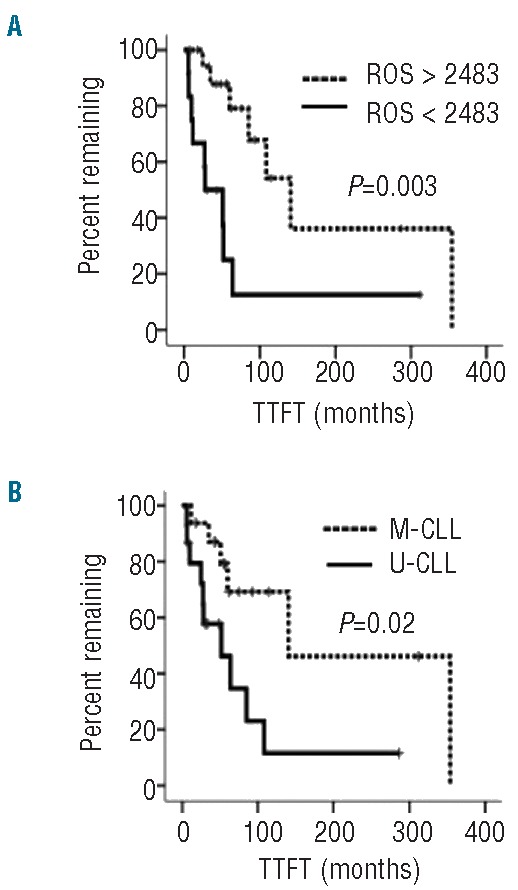
Association between elevated ROS and better prognosis. (A) Kaplan-Meier analysis of correlation between mean CM-H2DCFDA fluorescence (GeoMFI), as a measure of ROS, and time to first treatment (TTFT). (B) Kaplan-Meier analysis of correlation between IGHV mutation status and TTFT within the cohort of patients studied.
In addition to inter-sample variation, CM-H2DCFDA staining also revealed intra-sample heterogeneity in ROS levels (Figure 1Ai). Relatively discrete sub-populations were detected in samples from 16/33 patients. Similar results were obtained when we specifically quantified ROS in CD5+CD19+ cells confirming that this heterogeneity exists within the malignant clone (Figure 1Aii). Subpopulations were somewhat more common in U-CLL (9/14) than in M-CLL (7/19) although this difference was not statistically significant (data not shown).
We performed co-staining with CM-H2DCFDA and anti-CXCR4 to probe the basis for intraclonal ROS heterogeneity. CXCR4 is strongly down-modulated following CXCL12 engagement in tissues; reduced CXCR4 expression therefore marks cells that have recently emigrated from tissues to blood.9,10 In 4/4 randomly selected samples (3 M-CLL, 1 U-CLL), there was an inverse relationship between CXCR4 and ROS and, on average, CXCR4 expression was 2.1-fold higher in ROSlow cells than in ROShi cells (Figure 3A,B). Thus, ROS levels are higher within the most recently released cells, suggesting that increased ROS is a consequence of tissue-based stimulation.
Figure 3.
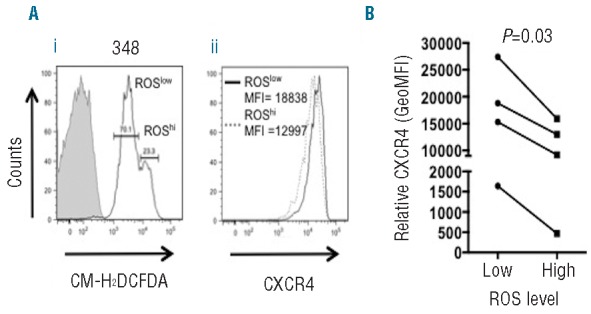
Heterogeneity of ROS in primary CLL. CLL samples were recovered from cryopreservation, rested for 3 h and then stained with CM-H2DCFDA and anti-CD19, anti-CD5 and anti-CXCR4 antibodies prior to flow cytometric analysis. (A) Representative flow analysis showing (i) CM-H2DCFDA fluorescence in CD19+CD5+ cells and (ii) CXCR4 expression in “ROSlow” and “ROShigh” sub-populations. (B) Quantitative data for CXCR4 expression in “ROSlow” and “ROShigh” sub-populations from four CLL samples analyzed.
Taken together, our results demonstrate that increased ROS may be associated with anergy in CLL since ROS levels were highest in M-CLL, and in samples with strong down-modulation of sIgM expression and function. CLL anergy appears to be a feature of more indolent forms of disease and high levels of ROS were also associated with longer TTFT. Consistent with the idea that CLL anergy is driven by antigen engagement within tissues,8 increased ROS may also be a consequence of tissue-based stimulation since the highest levels of ROS were found in cells with low CXCR4 that had been most recently released into the circulation.9,10 Interestingly, gene expression profiling revealed significant perturbation of ROS-related gene expression between CXCR4lowCD5hi and CXCR4hiCD5low sub-populations further supporting the idea that ROS production is a consequence of tissue-based stimulation in vivo.10
The mechanisms that mediate increased levels of ROS in anergic CLL cells remain unclear. One possibility is that increased ROS is a consequence of anergy-promoting, antigen interactions acting via the BCR. We attempted to measure effects of anti-IgM stimulation on ROS levels of CLL cells using CM-H2DCFDA (to quantify total ROS) or a sIgM-localizing ROS probe, to specifically quantify ROS in the vicinity of activated sIgM.11 However, we were unable to detect consistent changes in ROS levels using either approach (data not shown). Given the relatively low signaling capacity of CLL cells, more sensitive, regiospecific probes will be required for meaningful investigation of direct effects of sIgM of ROS production in CLL cells in vitro. Moreover, responses induced by anti-IgM in responsive samples in vitro most closely resemble positive, growth-promoting signal responses and may therefore not adequately model anergy-promoting antigen responses in vivo.
Increased ROS may be a consequence of increased production and/or decreased metabolism via cellular antioxidant protective mechanisms. Mitochondrial-derived ROS are mainly produced as a by-product of mitochondrial respiration and BCR signaling is known to modulate metabolism, including oxidative phosphorylation, with differences between naïve and anergic B cells.12 Thus, differences in metabolism, perhaps linked to changes in mitochondrial mass,2,4 between individual CLL samples may explain, at least in part, variation in ROS levels. It is also possible that ROS production may be linked to apoptosis. Interestingly, although the viability of the samples studied was generally high, we did identify a significant inverse correlation between ROS levels and the proportion of viable cells. One possibility is that increased ROS may be a consequence of activation of cell death pathways. Alternatively, high levels of ROS in CLL cells may promote susceptibility to apoptosis. Indeed, antioxidants can decrease CLL cell apoptosis.13,14
Although anergy is apparently benign, it is a reversible state and anergic CLL cells may act as a reservoir of cells which may be subject to subsequent growth-promoting antigen signaling and/or acquire secondary genetic alterations.15 Regardless of the underlying mechanisms, the observation that ROS are increased in anergic CLL cells suggests that targeting anergic CLL cells using agents that further elevate ROS levels may be an approach for selective induction of apoptosis and allow effective elimination of this important reservoir of malignant cells.
Acknowledgments
We are extremely grateful to the patients involved in this study for the kind gift of their samples. We are very grateful for the generous support of Drs Andrew Duncombe, Abraham Jacob and Henri Grech and their associated clinical teams. We are also very grateful for the support of Kathy Potter, Isla Henderson and Ian Tracy.
Footnotes
Funding: this work was supported by Leukaemia and Lymphoma Research, the CLL Global Research Foundation, Cancer Research UK, the Southampton Experimental Cancer Medicine Centre and the University of Southampton.
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Trachootham D, Zhang H, Zhang W, et al. Effective elimination of fludarabine-resistant CLL cells by PEITC through a redox-mediated mechanism. Blood. 2008;112(5):1912–1922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jitschin R, Hofmann AD, Bruns H, et al. Mitochondrial metabolism contributes to oxidative stress and reveals therapeutic targets in chronic lymphocytic leukemia. Blood. 2014;123(17):2663–2672. [DOI] [PubMed] [Google Scholar]
- 3.Tomic J, Lichty B, Spaner DE. Aberrant interferon-signaling is associated with aggressive chronic lymphocytic leukemia. Blood. 2011; 117(9):2668–2680. [DOI] [PubMed] [Google Scholar]
- 4.Carew JS, Nawrocki ST, Xu RH, et al. Increased mitochondrial bio-genesis in primary leukemia cells: the role of endogenous nitric oxide and impact on sensitivity to fludarabine. Leukemia. 2004; 18(12):1934–1940. [DOI] [PubMed] [Google Scholar]
- 5.Zhou Y, Hileman EO, Plunkett W, et al. Free radical stress in chronic lymphocytic leukemia cells and its role in cellular sensitivity to ROS-generating anticancer agents. Blood. 2003;101(10):4098–4104. [DOI] [PubMed] [Google Scholar]
- 6.Carew JS, Zhou Y, Albitar M, et al. Mitochondrial DNA mutations in primary leukemia cells after chemotherapy: clinical significance and therapeutic implications. Leukemia. 2003;17(8):1437–1447. [DOI] [PubMed] [Google Scholar]
- 7.Packham G, Krysov S, Allen A, et al. The outcome of B-cell receptor signaling in chronic lymphocytic leukemia: proliferation or anergy. Haematologica. 2014;99(7):1138–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mockridge CI, Potter KN, Wheatley I, et al. Reversible anergy of sIgM-mediated signaling in the two subsets of CLL defined by VH-gene mutational status. Blood. 2007;109(10):4424–4431. [DOI] [PubMed] [Google Scholar]
- 9.Coelho V, Krysov S, Steele A, et al. Identification in CLL of circulating intraclonal subgroups with varying B-cell receptor expression and function. Blood. 2013;122(15):2664–2672. [DOI] [PubMed] [Google Scholar]
- 10.Calissano C, Damle RN, Marsilio S, et al. Intraclonal complexity in chronic lymphocytic leukemia: fractions enriched in recently born/divided and older/quiescent cells. Mol Med. 2011;17(11–12):1374–1382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Capasso M, Bhamrah MK, Henley T, et al. HVCN1 modulates BCR signal strength via regulation of BCR-dependent generation of reactive oxygen species. Nat Immunol. 2010;11(3):265–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Caro-Maldonado A, Wang R, Nichols AG, et al. Metabolic repro gramming is required for antibody production that is suppressed in anergic but exaggerated in chronically BAFF-exposed B cells. J Immunol. 2014;192(8):3626–3636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Steele AJ, Jones DT, Ganeshaguru K, et al. The sesquiterpene lactone parthenolide induces selective apoptosis of B-chronic lymphocytic leukemia cells in vitro. Leukemia. 2006;20(6):1073–1079. [DOI] [PubMed] [Google Scholar]
- 14.Nilsson J, Soderberg O, Nilsson K, et al. Thioredoxin prolongs survival of B-type chronic lymphocytic leukemia cells. Blood. 2000; 95(4):1420–1426. [PubMed] [Google Scholar]
- 15.Caligaris-Cappio F. Anergy: the CLL cell limbo. Blood. 2014; 123(21):3214–3215. [DOI] [PubMed] [Google Scholar]