

Abstract
Genome-wide association studies (GWAS) have prioritized a transcription factor, Nuclear Receptor 2 Family 2 (NR2F2), as being associated with essential hypertension in humans. Here we provide evidence that validates this association and indicates that Nr2f2 is a genetic determinant of blood pressure (BP). Using the zinc-finger nuclease technology, the generation of a targeted Nr2f2-edited rat model is reported. The resulting gene-edited rats have a 15bp deletion in exon 2 leading to a 5 amino acid deletion in the hinge region of the mutant Nr2f2 protein. Both systolic and diastolic blood pressures of the Nr2f2mutant rats are significantly lower than controls. Because the hinge region of Nr2f2 is required for interaction with Friend of Gata2 (Fog2), protein-protein interaction is examined. Interaction of Nr2f2mutant protein with Fog2 is greater than that with the wild type Nr2f2 indicating that the extent of interaction between these two transcription factors critically influences BP.
Introduction
BP is a complex polygenic trait influenced by multiple genetic elements, the number, magnitude of effect and identities of which are largely unknown. Genome-wide association studies have prioritized genes, based on their association with hypertension, as potential candidates for BP regulation. The Wellcome Trust Case-Control Consortium (WTCCC) was one of the first large scale GWAS to report a suggestive association on a region on human chromosome 15 to hypertension1. Subsequently, by a haplotype-based re-analysis of this data the gene nuclear receptor subfamily 2, group F, member 2 (NR2F2) also known as Chicken ovalbumin upstream promoter transcription factor (COUP-TFII) was highly prioritized as a candidate gene associated with hypertension2. Four other linkage studies in humans have reported the genomic region containing NR2F2 as being linked to essential hypertension3–6. Similar lines of evidence were obtained with multiple linkage and substitution mapping studies using rat models of hypertension7–11. Nr2f2 is a nuclear receptor transcription factor vital for angiogenesis and heart development12, but its role in BP regulation remains undefined.
The current study was conducted to directly test the involvement of Nr2f2 in BP regulation. For this purpose, a targeted gene-edited rat model was generated, in which the Nr2f2 protein was mutated in the hinge region. Both systolic and diastolic blood pressures (BP) of the Nr2f2mutant rats are significantly lower than controls, indicating that Nr2f2 is indeed a genetic determinant of BP. Further, the structural mechanism by which Nr2f2 influences BP is delineated to the hinge region, through which it interacts with another transcription factor, Friend of Gata2 (Fog2). The extent of interaction of Nr2f2 with Fog2 is enhanced when the Nr2f2 protein is mutated in the hinge region. These data provide evidence for the extent of interaction between two transcription factors, Nr2f2 and Fog2 as being important for the regulation of blood pressure.
Results and Discussion
Targeted-editing of the Nr2f2 locus
Gene deletion mouse models serve as genetic tools to validate observations documented through genetic linkage and association studies. Nr2f2 knockout mice are, however, embryonic lethal13. To examine the role of Nr2f2 in BP regulation, we therefore focused on constructing a Nr2f2mutant rat model without disrupting the two important domains of the Nr2f2 protein, the DNA-binding domain and the ligand-binding domain. Custom- zinc-finger nucleases (ZFNs) targeting exon 2 of Nr2f2 were designed. Exon 2 of Nr2f2 codes for amino acids in the hinge region of the Nr2f2 protein, which is between the DNA-binding and the ligand-binding domains. ZFNs targeting Nr2f2 were microinjected into single-cell embryos of the hypertensive rat strain- the Dahl Salt-sensitive (S) rat. Embryonic lethality was bypassed by the hypomorphic founder Nr2f2mutant rats. These rats had a shorter (345 bp) PCR-amplified genomic DNA fragment encompassing exon 2 of Nr2f2 compared with the wild-type Nr2f2 (360 bp) genomic fragment (Figure 1A). Sequencing of the PCR products confirmed the deletion to be 15bp from 1571 to 1556 bp of the mRNA of Nr2f2 (Fig. 1B). Two different antibodies against Nr2f2 were used to clarify the protein status of Nr2f2. Immunoblotting with an N-terminal antibody detected the presence of Nr2f2 protein in both the Nr2f2mutant rats and the wild type hypertensive rats, whereas, antibodies to the hinge region of Nr2f2 cross-reacted exclusively with the wild-type Nr2f2 protein but not the mutant Nr2f2 protein (Fig. 2, Supplementary Figure 1).
Figure 1. Screening animals for ZFN-targeted mutation at the Nr2f2 locus.

(A) Tail DNA samples from pups born post-microinjection of custom ZFNs targeting exon 2 of the Nr2f2 locus were screened by PCR amplification with primers designed to amplify rat genomic fragments encompassing the ZFN-targeted site. Deviations from the expected product size of 360bp were inferred as genomic DNA from rats with mutations at the Nr2f2 locus. (B) Representative sequencing results from the PCR products shown in panel A detected a 15 bp deletion in the mRNA from mutant rats. Also represented are the corresponding translated peptide sequences as a result of the nucleotide variations.
Figure 2. Targeted disruption of Nr2f2.

Top: Schematic diagram of Nr2f2 protein, highlighting the key domains: AB domain (aa 1-78); C domain or DNA binding domain (aa 79-144); D domain or Hinge region (aa 145-213); E domain or Ligand binding Domain (aa 214-414). Bottom: Western blot (WB) analysis of Nr2f2 protein expression in Nr2f2mutant and Wild-type hypertensive rats. Anti-Nr2f2 antibodies are labeled as Ab1 and Ab2. Ab1: Antibody from Abcam, raised against a synthetic peptide directed towards amino acids 43–64 (Abcam); Ab2: Antibody (Cell Signaling) raised against a synthetic peptide directed towards the hinge region (amino acids around 159).
Attenuation of hypertension in the Nr2f2mutant rats
Compared to the wild-type hypertensive rats, Nr2f2mutant rats maintained on a high salt (2% NaCl) diet had lower systolic BP (179±3 vs. 197±5 mmHg; t-test, p<0.05) as measured by the tail-cuff method. To further confirm these observations, rats were surgically implanted with radio transmitters and BP was continuously recorded by telemetry. Both the Nr2f2mutant rats and the wild type hypertensive rats maintained normal diurnal rhythms of systolic and diastolic BP. However, throughout the observation period, both systolic and diastolic BP of the Nr2f2mutant rats were consistently lower than that of the wild type hypertensive rats (Figs. 3A, B). Nr2f2mutant rats maintained on a low salt (0.3% NaCl) diet did not however demonstrate a similar BP lowering effect (Supplementary Fig. 2). Figures 3C and 3D demonstrate the blunted increase in salt-induced blood pressure in the mutant rats compared to the hypertensive rats. These data constitute strong evidence to indicate that the structural alterations within the hinge-region of Nr2f2 contribute importantly to salt-sensitive BP regulation.
Figure 3. Lower Blood Pressure of Nr2f2mutant rats.

Measurements of (A) Systolic and (B) Diastolic blood pressures of hypertensive rats (n = 7) and Nr2f2mutant rats (n = 8). Rats were monitored for BP after surgical implantation of radiotelemetry transmitters. Data plotted are the recordings obtained once every 5 min continuously for 24 h and averaged for 4 h intervals; (C) Comparison of low-salt versus high-salt diet regimens. Data presented from low-salt and high-salt fed rats was obtained from two independent experiments. (D) Delta BP calculated by subtracting the average BP of the hypertensive rat from the average BP of Nr2f2mutant rats within each of the dietary salt groups labeled as LS for low-salt diet and HS for high-salt diet. Levels of statistical significance for all data were analysed by independent sample t-test. *P<0.05, NS Not significant.
Improved cardiac and renal function of the Nr2f2mutant rats
Nr2f2mutant rats exhibited superior cardiac function compared with the wild type hypertensive rats. As measured by echocardiography, Nr2f2mutant exhibited superior left ventricular function as demonstrated by increased fractional shortening (FS) (56%+/-3 vs. 41%+/−2; t-test, p<0.05, Fig. 4A), increased velocity of circumferential shortening (Vcf) (8.6+/−0.3 vs. 6.1+/−0.4; t-test, p<0.05, Fig. 4B) and increased FS/MPI (Fractional shortening/Myocardial performance index) (1.48+/−0.22 vs. 0.78+/−0.08; t-test, p<0.05, Fig. 4C). Nr2f2mutant rats also exhibited superior renal function compared with the wild type hypertensive rats. Urinary protein excretion of Nr2f2mutant rats was lower compared with the wild type hypertensive rats (87.97±7.07 vs. 122.63±7.07; t-test, p<0.05) (Fig. 5). These additional physiological parameters that provide evidence to the overall improvement in cardiovascular and renal function that are associated with the observed lowering of BP in the Nr2f2mutant rats.
Figure 4. Nr2f2mutant rats exhibited superior left ventricular function.

Cardiac parameters of male hypertensive rats (n = 12) and Nr2f2mutant rats (n = 12) as determined by echocardiography. (A) Fractional Shortening (FS), (B) Velocity of Circumferential shortening (Vcf) (C) Fractional shortening/Myocardial performance index (FS/MPI). Levels of statistical significance for all data were analyzed by independent sample T-Test; *p< 0.05.
Figure 5. Lower Urinary Protein excretion in Nr2f2mutant rats compared with S rats.
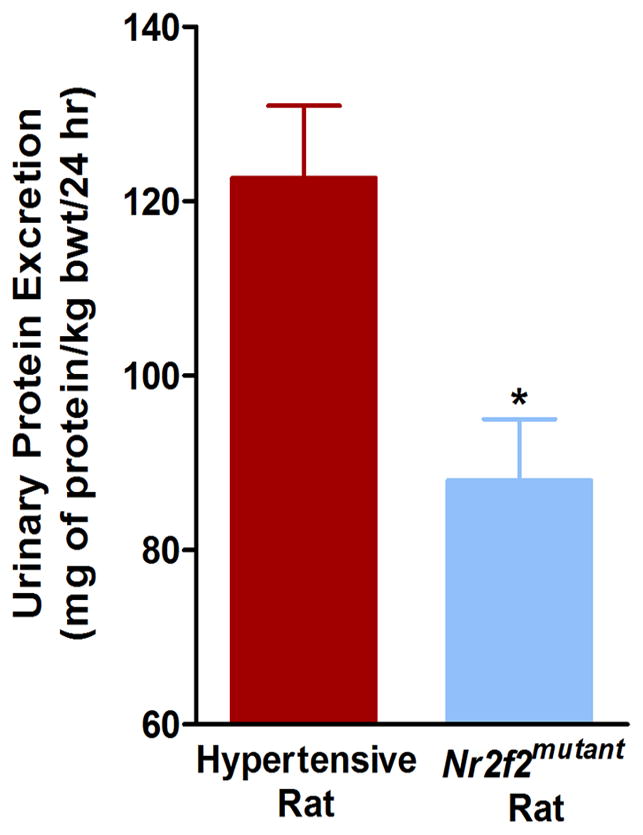
Total 24 h urine protein was assessed in male hypertensive rats (n = 18) and Nr2f2mutant rats (n =18) as described in the Materials and Methods section. Urinary protein excretion data are presented as milligrams of protein/kg of body weight over a 24h period; t-test, *p<0.05.
Interaction between Nr2f2 and Fog2 is linked to BP regulation
Next, we sought to explore the molecular mechanism by which Nr2f2 mediates the observed alterations in blood pressure, we focused on the sole difference between the two strains, which is in the hinge region of Nr2f2. Amino acids 117–414 of Nr2f2 are required for protein-protein interaction with another transcriptional regulator, Friend of Gata2 (Fog2)14. Fog2 is a multi-zinc finger transcriptional co-repressor protein required for cardiac development. Fog2 regulates cardiac morphogenesis by interaction with multiple cardiac specific transcriptional factors. Nr2f2 and Fog2 null mice have similar defects related to mesenchymal-epithelial and endothelial interaction, which are important for heart and blood vessel formation14. Of further interest, Ciullo et al., identified a new locus strongly linked to hypertension in human chromosome 8q22-23 region, within which, Fog2 is prioritized as one of the candidate genes for essential hypertension15. We therefore hypothesized that the protein-protein interaction between the hinge region of Nr2f2 and Fog2 is important for BP regulation. To test this hypothesis, wild type and mutant forms of Nr2f2 cDNA, with Histidine tags, were cloned into pCDNA 3.1(B) myc/His (−) expression vector and overexpressed in NRK52E or COS-7 cells (Fig. 6A, Supplementary Fig. 3). The overexpressed Nr2f2 proteins along with their binding partners were immunoprecipitated with the Nr2f2 N-terminal antibody or with histidine tags and probed with the antibody against Fog2. Compared with the wild-type Nr2f2 protein, the binding of the mutant form of Nr2f2 to Fog 2 was more pronounced (Figure 6B). To further assess the downstream effect of this interaction, we conducted a promoter chromatin immuno-precipitation experiment. As shown in Figure 7, an enrichment of Nr2f2 binding to the promoter of atrial natriuretic factor, Anf, was noted in the Nr2f2mutant rats. Anf is a direct target gene known to be influenced by the Nr2f2-Fog2 interaction14. Similar enrichments were not observed with promoters for renin and ApoB, which are other targets of Nr2f216,17 that are not described in the literature to require Fog2. Because Anf is a vasorelaxant18,19, rats were further tested for vasoreactivity. Vasorelaxation responses of the mesenteric arteries from Nr2f2mutant rats were significantly higher than that of the wild-type rats (Fig. 8). These data indicate that the extent of interaction between Nr2f2 and Fog2 through the hinge region of Nr2f2 is linked to the extent of blood pressure and that modulation of transcription of Anf, is perhaps, one of the potential underlying mechanisms.
Figure 6. Nr2f2 interaction with Fog2.
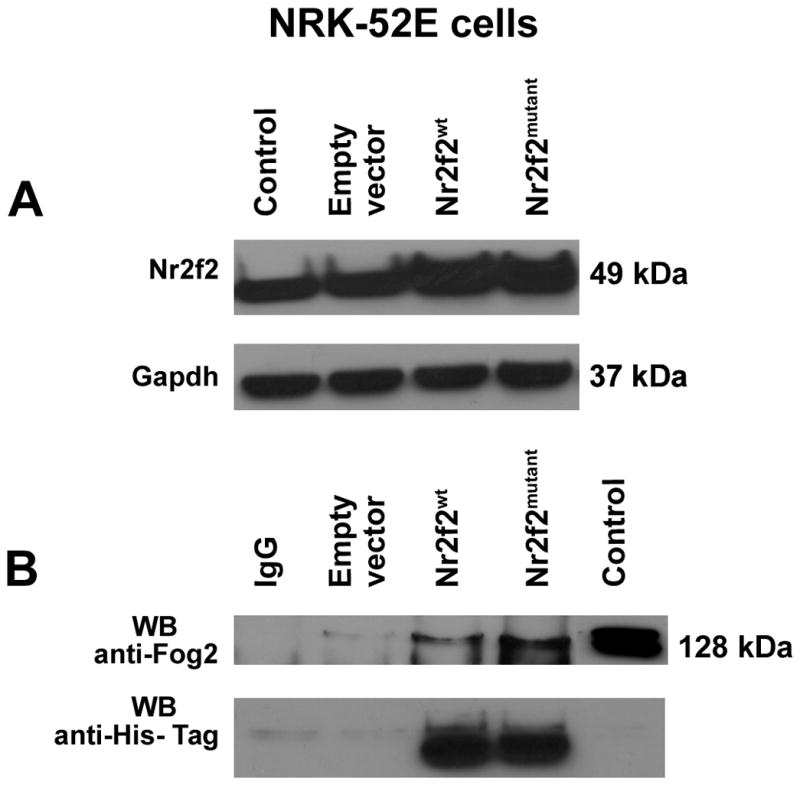
Co-Immunoprecipitation analysis of Nr2f2 and Fog2 association in NRK52E cells. Extracts from NRK52E cells were transfected with expression plasmid for Nr2f2mutant and Nr2f2wildtype. (A) Western blotting (WB) of cell lysates for Nr2f2 protein to confirm the over expression of both forms of Nr2f2. Lanes labeled from the left- Control: NRK52E cell lysate; Empty vector: NRK52E cells transfected with empty pcDNA 3.1(-B) myc/His vector; Nr2f2wt: NRK52E cells transfected with Nr2f2 wild type pcDNA 3.1(B) myc/His (−) construct; Nr2f2mutant: NRK52E cells transfected with Nr2f2 mutant pcDNA 3.1(B) myc/His (−) construct. (B) Cell extracts were subjected to immunoprecipitation (IP) with Nr2f2 antibody (Nr2f2 Ab-1). The immune complexes were captured using Agarose G beads. The immune-precipitated protein samples were denatured, resolved by SDS-Polyacrylamide gel electrophoresis and transferred onto a PVDF membrane. The membrane was probed by western blotting (WB) with anti-Fog2 antibody and anti-His-Tag antibody. A representative of four experiments is shown. Lanes labeled from the left- IgG: IgG antibody control; Empty vector: NRK52E cells transfected with empty pcDNA 3.1(-B) myc/His vector; Nr2f2wt: NRK52E cells transfected with Nr2f2 wild type pcDNA 3.1(B) myc/His (−) construct; Nr2f2mutant: NRK52E cells transfected with Nr2f2 mutant pcDNA 3.1(B) myc/His (−) construct; Control: NRK52E cell lysate. Corroborative data obtained from Histidine-tagged Nr2f2 overexpressed in COS-7 cells are presented in Supplementary Figure 3.
Figure 7. Enrichment of Nr2f2 in Anf promoter region in Nr2f2mutant rats.
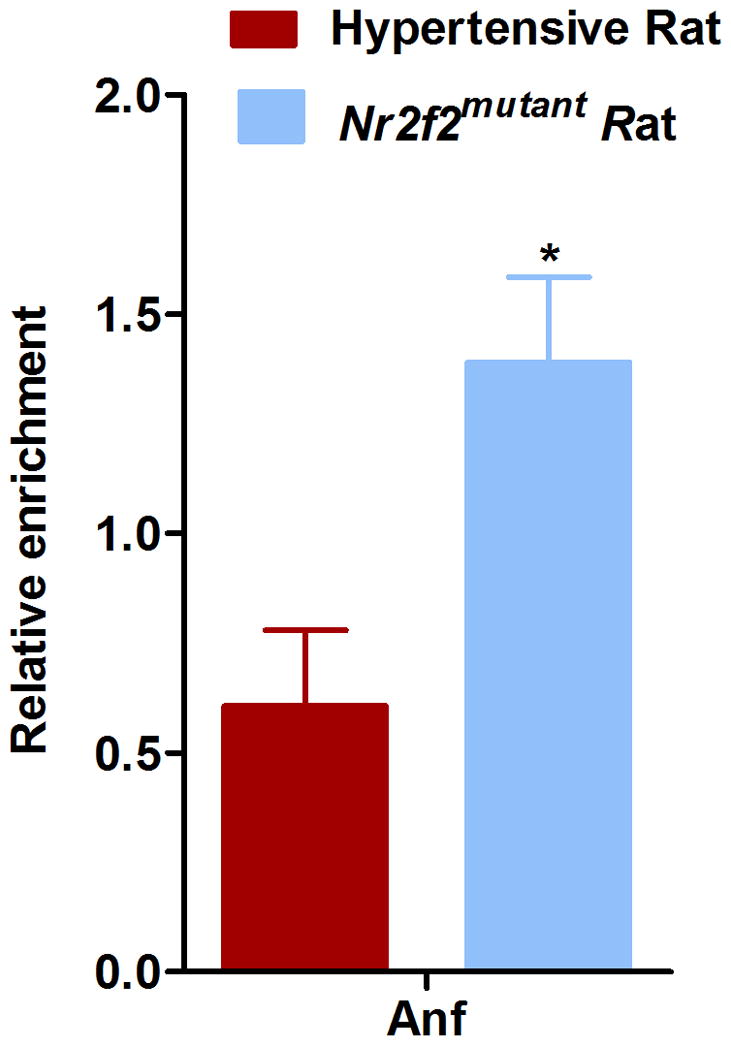
Chromatin immunoprecipitation (ChIP) was conducted using heart tissue lysate from wild-type hypertensive (n=3) and Nr2f2mutant rats (n=3) with an anti-Nr2f2 antibody (Nr2f2-Ab1, Abcam) or control IgG antibody (Cell signaling). ChIP enrichment at the Anf locus was quantified by qPCR and normalized to Hemoglobin (Hbb) promoter amplification, which was used as a negative control. *p< 0.05.
Figure 8. Nr2f2mutant rats demonstrated superior vascular function compared with hypertensive rats.

(A). Mesenteric arteries from male hypertensive rats (n=9) and Nr2f2mutant rats (n=4) were dissected and mounted as described under the methods section. Endothelium-dependent relaxation to Acetylcholine (Ach) was assessed by adding increasing concentrations of ACh (0.001–1μM) to the vessel preparation. Relaxation to ACh was expressed as a percentage of the level of pre-constriction induced by submaximal dose of phenylephrine. (B). Cumulative concentration-response curves (CCRC) to phenylephrine (0.1–100μM) of mesentery were recorded from wild-type hypertensive (n=9) and Nr2f2mutant rats (n=4). Results are expressed as mean±S.E. (C). Mesenteric arteries from male hypertensive (n=5) and Nr2f2mutant rats (n=5) were dissected and mounted as described under the methods section. Endothelium-independent relaxation to sodium nitroprusside (SNP) was assessed by adding increasing concentrations of SNP (1nM–100μM) to the vessel preparation. Relaxation to SNP was expressed as a percentage of the level of pre-constriction induced by submaximal dose of phenylephrine. Data were analyzed by t-test. *P<0.05, **P<0.01, ***P<0.001. NS, Not significant.
In summary, our study provides an important step forward in our current understanding of Nr2f2 as a genetic factor associated with essential hypertension in humans and in rodent models of hypertension. Specifically, combined with the available linkage studies in rat models, by utilizing a novel mutant model of Nr2f2, we have not only demonstrated that Nr2f2 is a genetic determinant of BP, but also documented the significance of the relatively less-explored hinge region of Nr2f2 as an important molecular feature, which through its interaction with Fog2, is linked to BP regulation.
METHODS
Animals
All animal procedures and protocols used in this report were approved by the University of Toledo Health Science Campus Institutional Animal Care and Use Committee.
Generation of a ZFN-Mediated Nr2f2mutant Rat
Zinc- finger nuclease (ZFN) construct specific for the rat Nr2f2 gene (NM_080778.2) were designed, assembled, and validated by Sigma-Aldrich, to target the first exon 2 of Nr2f2 (Target sequence: CAGGATGCCGCCCACCCAgcctacTCACGGGCAGTTTGCACTG; ZFN binds to each underlined sequence on opposite strands). mRNA encoding the Nr2f2 ZFN pairs at a concentration of 10 ng/uL were injected into one cell SS/McwiHsd (SS) rat embryos20. Out of a 147 embryos injected, 66 healthy embryos were transferred to pseudo-pregnant female SD:Crl rats (Charles River), of which five pups were born. At 14 days of age, pups were tagged, tail clipped and DNA was extracted and amplified using primers flanking the above target sequence Nr2f2-F (5′ GGGGAAGTGATTTGTGAGGA 3′) and Nr2f2-R (5′ GCCCATGATGTTGTTAGGCT 3′). PCR products were analyzed for ZFN-induced mutations using a Cel-I assay as described previously20. Among the 5 pups born, 2 positive founders (male/female) were identified. PCR products were TOPO cloned and sequenced to reveal the same 15-bp deletion ‘CCTACTCACGGGCAG’ in exon 2 of the Nr2f2 gene in both mutant animals. The founder male/female rats were backcrossed to the parental SS/JrHsdMcwi strain. Multiple separate pairs of mutation-carrying progeny were then intercrossed to generate an F2 population that was used for phenotyping and breeding to homozygosity. The DNA sequencing of the homozygous Nr2f2mutant rats confirmed the 15-bp deletion in F2 offspring, which results in a predicted loss of five amino acids PTHGQ.
Blood Pressure Measurements by the tail-cuff and radiotelemetry methods
Each set of wild-type hypertensive SS/JrHsd Mcwi rats, hitherto referred as hypertensive rats (n = 20 males) and Nr2f2mutant rats (n = 17 males), hypertensive rats (n = 6 females) and Nr2f2mutant rats (n = 6 females) were bred, housed, and studied concomitantly to minimize environmental effects. All rats were fed with 0.3% NaCl low-salt diet (Harlan Teklad), weaned at 30 days of age. To assess salt-sensitivity, at the age of six weeks rats were divided into two groups. The low salt group was continued to be fed with the 0.3% NaCl low-salt diet, whereas the high salt group was switched to a diet containing 2% NaCl. After 24 days on these diets, systolic blood pressure (BP) of the rats was measured by using the tail-cuff method21. Conscious restrained rats were warmed to 28°C. The BP of each rat was measured for 2 consecutive days by two blinded operators. BP values for each day were the mean of three or four consistent readings. The final BP value used was the mean of the two daily BP values. BP was also collected using a telemetry system (Data Sciences International, St. Paul, MN)22. Four days after the BP measurements by the tail cuff method, Nr2f2mutant (n = 8 males) and wild-type hypertensive rats (n = 7 males) rats were surgically implanted with C40 radio-telemetry transmitters such that the body of the transmitter is placed into the left flanks and the probe is inserted into their femoral arteries all the way into the lower abdominal aortae. Rats were allowed to recover from surgery for 4 days before the transmitters were turned on for recording BP.
Urinary Protein Excretion (UPE)
Within 5 days following BP measurements, Nr2f2mutant (n=17) and wild-type hypertensive (n=18) rats were housed individually in metabolic cages and urine samples were collected over a 24 hr period. The pyrogallol based QuanTtest Red Total Protein Assay from Quantimetrix (Redondo Beach, CA) was used to determine protein concentrations of the urine samples. A VERSAmax microplate reader from Molecular Devices (Sunnyvale, CA) was used to determine absorbance at 600nm. Protein concentrations were determined by reading against the absorbance of the QuanTtest human protein standards (25–200 mg/dL). UPE data is presented as milligrams of protein/kg of body weight over a 24h period (mg of protein/kg bwt/24h).
Echocardiography
Nr2f2mutant (n = 12) and wild-type hypertensive rats (n = 12) were evaluated by transthoracic echocardiography for determination of fractional shortening (FS), velocity of circumferential shortening (Vcf), and myocardial performance index(MPI) using a Sequoia C512 System (Siemens Medical) with a 15-MHz linear array transducer23,24. The rats were anesthetized with 1.5–2.0% isoflurane by mask, the chest was shaved, the animal was situated in the supine position on a warming pad and ECG limb electrodes were placed. Two-dimensional guided M-mode images and Doppler studies of aortic and trans-mitral flows were obtained from the parasternal long and foreshortened apical windows, respectively. Study duration was typically 15–20 min per animal.
All data were analyzed offline with software resident on the ultrasound system in an investigator-blinded fashion. LV end-diastolic (LVDd) and end-systolic (LVSd) dimensions were determined from M-mode tracings recorded at the mid-papillary level, and LV fractional shortening (FS) was calculated as [(LVDd-LVSd)/LVDd] × 100%. Ejection time and the isovolumic contraction and relaxation times were measured from color-flow directed Doppler pulsed-wave traces of mitral and aortic flow obtained at the level of the LV outflow tract from the apical four-chamber view. Aortic outflow and mitral inflow waveforms were recorded when the mitral and aortic wave forms were distinct and aortic and mitral valve clicks were clearly visible. Ejection, contraction, and relaxation times were determined from three consecutive beats and averaged to calculate MPI. MPI was calculated as the sum of the isovolumic contraction and relaxation times divided by the ejection time. The velocity of circumferential shortening (Vcf) was calculated as FS/ejection time. Values are means ± SE. Groups were compared with Student’s t-test.
Vascular Reactivity
After recording blood pressure by radiotelemetry the wild-type hypertensive (n=9) and Nr2f2mutant rats (n=4) were euthanized with CO2 inhalation method. The mesentery was immediately excised and placed in ice-cold physiological salt solution (PSS) consisting of mmoles/l of 118.5 NaCl, 4.7 KCl, 2.5 CaCl2, 1.2 MgSO4, 1.2 KH2PO4, 25.0 NaHCO3, and 5.5 D-glucose aerated with 95%O2-5%CO2. Using a dissecting microscope, the mesenteric arteries were gently cleaned of adipose and connective tissue and cut into segments of approximately 2 mm in length. The segments were then mounted in a Mulvany-Halpern style small vessel wire myograph chamber (Model 620M, Danish Myotechnology, Aarhus, Denmark) filled with 6 ml PSS, maintained at 37°C, and continuously aerated with 95% O2, 5% CO2 to measure vascular reactivity 25–27. After 30 min of equilibration period, a standardized computer-assisted normalization procedure was performed to set the pretension of the arteries using LabChart8 software (AD Instruments, Colorado Springs, CO). This involves setting a circumference of the arteries equivalent to 90% of that which they would have at a transmural pressure of 100mmHg. Segments were washed with PSS and left to equilibrate for another 30 min. Vessel contractility was then assessed with the high-K+ (120mmol/L) solution. After washing the segment 3–4 times with PSS, cumulative concentration-response curve (CCRC) to phenylephrine (0.1–100μM) was obtained. After another 30-min wash, the segments were precontracted with submaximal dose of phenylephrine until the plateau was achieved. Dose-dependent relaxation responses to ACh (0.001–1μM) and sodium nitroprusside (SNP) (1nM–100μM) were assessed by constructing CCRC. Relaxation in response to ACh and SNP were expressed as a percentage of the level of pre-constriction induced by submaximal dose of phenylephrine.
Western blotting (WB)
Antibodies were from the following sources: Anti-Nr2f2 mouse monoclonal (Nr2f2-Ab1, Abcam, 1:2000 dilution), anti-CoupTF II (Nr2f2-Ab2, Cell signaling, 1:500 dilution), anti-Fog2 (GeneTex, 1:1000 dilution), donkey anti-mouse IgG-HRP conjugate (SC-2020, 1:5000 dilution), anti-rabbit IgG-HRP conjugate (Cell Signaling, 1: 5000 dilution). Samples were homogenized in ice-cold RIPA lysis buffer with protease inhibitor cocktail (Pierce). Immuno-blotting for Nr2f2 was carried out using tissue lysates from Nr2f2mutant and wild-type hypertensive rat homogenates. In brief, forty microgram of proteins were boiled with Laemmli sample buffer (Bio-Rad) for 5 minutes at 95° C. Protein samples were resolved using 4–15% Criterion Tris-HCl Gel (Bio-Rad) at room temperature, and transferred on to PVDF membrane (Millipore), blocked with 5% fat free milk powder in PBST and incubated with anti-Nr2f2 and secondary antibodies conjugated with horseradish peroxidase. Immunoreactivity was detected by autoradiography using SuperSignal West Pico Chemiluminescent Substrate (Pierce). Full-length blots are shown in Supplementary Figure 1.
Chromatin Immunoprecipitation (ChIP)
ChIP16,28 was performed using heart tissue from wild-type male hypertensive (n=3) and Nr2f2mutant rats (n=3). Tissues was minced and fixed with 1% formaldehyde. After crosslinking, nuclei were isolated and the cross-linked chromatin was subjected to sonication to obtain DNA fragments. Chromatin (50μg) was immunoprecipitated with 8μg of Nr2f2 antibody (Nr2f2-Ab1). As a negative control, chromatin was precipitated with 8μg of mouse IgG (Cell signalling). The immune complexes were precipitated with protein G-Agarose beads. Precipitated chromatin was eluted from the beads and crosslinks were reversed overnight at 65°C. Chromatin was treated with RNase A, proteinase K, and the DNA was column purified (PCR Purification kit, Qiagen). Purified DNA was PCR amplified using primers (Supplementary Table 1) targeting the Anf, Renin and Apob promoter regions, and Hbb used as a negative control.
Cell culture, transfection and immunoprecipitation
NRK52E cells were obtained from ATCC (Catalog number CRL-1571) and cultured in DMEM supplemented with 5% fetal bovine serum, 100units/ml penicillin and 100μg/ml streptomycin in a humidified atmosphere at 37 °C with 5% CO2. Both wild type and mutant forms of Nr2f2 cDNA were cloned into the pCDNA 3.1(B) myc/His (−) expression plasmid. NRK52E cells were transfected with both wild-type and mutant forms of Nr2f2 plasmids using X-tremeGENE HP DNA transfection reagent (Roche Applied Science). Forty eight hours after transient transfection, cells were treated with the cell lysis buffer and used for subsequent experiments. For immunoprecipitation, lysates (500μg of protein) were pre-cleared with protein G agarose beads (Pierce) at 4°C for 2h. After removal of the beads by centrifugation, lysates were incubated overnight with anti-Nr2f2 antibody (5μg/IP experiment, Nr2f2-Ab1, Abcam) at 4°C. The immunoprecipitated complex was captured by incubating with 50 μl of protein G agarose beads. Western blotting was performed with Anti-Fog2 antibody (1:1000 dilution) as detailed above.
Statistical Analysis
All BP statistical analyses were done using the SPSS software (SPSS, Chicago, IL). Data were analyzed by independent sample t-test. Data are presented as the mean ± standard error. A p-value of ≤ 0.05 was used as a threshold for statistical significance. For BP and UPE experiments measurements the number of rats in each group ranged from 6 to 20.
Supplementary Material
Acknowledgments
This work was funded by National Institutes of Health Grants HL076709, HL112641 and HL020176 (to B.J.). Authors thank Drs. Howard Jacob and Aron Geurts at the Medical College of Wisconsin for assistance in developing the Nr2f2 mutant rat model; these models were developed, in part, through an American Reinvestment and Recovery Act (ARRA). Grand Opportunity award HL101681 to Dr. Jacob. Authors also thank Dr. Nitin Puri at the University of Toledo College of Medicine and Life Sciences for help with the vascular reactivity studies.
Footnotes
Author Contributions
B.J. conceived and designed the project. S.K., H.W., K.G., E. M., and B.M. performed the experiments. S.K., E.M., and H.W., analyzed the data. B.J., S.K., K.G., E.M., and H.W., analyzed the data. B.J., S.K., E.M., and H.W. prepared the manuscript.
Competing Financial Interest
The authors declare no competing financial interest.
References
- 1.Genome-wide association study of 14,000 cases of seven common diseases and 3, 000 shared controls. Nature. 2007;447:661–78. doi: 10.1038/nature05911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Browning BL, Browning SR. Haplotypic analysis of Wellcome Trust Case Control Consortium data. Hum Genet. 2008;123:273–80. doi: 10.1007/s00439-008-0472-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Krushkal J, et al. Genome-wide linkage analyses of systolic blood pressure using highly discordant siblings. Circulation. 1999;99:1407–10. doi: 10.1161/01.cir.99.11.1407. [DOI] [PubMed] [Google Scholar]
- 4.Weder AB, et al. Erythrocyte sodium-lithium countertransport and blood pressure: a genome-wide linkage study. Hypertension. 2003;41:842–6. doi: 10.1161/01.HYP.0000048703.16933.6D. [DOI] [PubMed] [Google Scholar]
- 5.Xu X, et al. An extreme-sib-pair genome scan for genes regulating blood pressure. Am J Hum Genet. 1999;64:1694–701. doi: 10.1086/302405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Xu X, et al. Mapping of a blood pressure quantitative trait locus to chromosome 15q in a Chinese population. Hum Mol Genet. 1999;8:2551–5. doi: 10.1093/hmg/8.13.2551. [DOI] [PubMed] [Google Scholar]
- 7.Joe B, et al. Transcriptional profiling with a blood pressure QTL interval-specific oligonucleotide array. Physiol Genomics. 2005;23:318–26. doi: 10.1152/physiolgenomics.00164.2004. [DOI] [PubMed] [Google Scholar]
- 8.Yagil C, Hubner N, Kreutz R, Ganten D, Yagil Y. Congenic strains confirm the presence of salt-sensitivity QTLs on chromosome 1 in the Sabra rat model of hypertension. Physiol Genomics. 2003;12:85–95. doi: 10.1152/physiolgenomics.00111.2002. [DOI] [PubMed] [Google Scholar]
- 9.Hubner N, et al. Integrated transcriptional profiling and linkage analysis for identification of genes underlying disease. Nat Genet. 2005;37:243–53. doi: 10.1038/ng1522. [DOI] [PubMed] [Google Scholar]
- 10.Kato N, et al. Isolation of a chromosome 1 region affecting blood pressure and vascular disease traits in the stroke-prone rat model. Hypertension. 2003;42:1191–7. doi: 10.1161/01.HYP.0000103161.27190.67. [DOI] [PubMed] [Google Scholar]
- 11.Cui ZH, et al. Exaggerated response to restraint stress in rats congenic for the chromosome 1 blood pressure quantitative trait locus. Clin Exp Pharmacol Physiol. 2003;30:464–9. doi: 10.1046/j.1440-1681.2003.03860.x. [DOI] [PubMed] [Google Scholar]
- 12.Pereira FA, Qiu Y, Zhou G, Tsai MJ, Tsai SY. The orphan nuclear receptor COUP-TFII is required for angiogenesis and heart development. Genes Dev. 1999;13:1037–49. doi: 10.1101/gad.13.8.1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lee CT, et al. The nuclear orphan receptor COUP-TFII is required for limb and skeletal muscle development. Mol Cell Biol. 2004;24:10835–43. doi: 10.1128/MCB.24.24.10835-10843.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huggins GS, Bacani CJ, Boltax J, Aikawa R, Leiden JM. Friend of GATA 2 physically interacts with chicken ovalbumin upstream promoter-TF2 (COUP-TF2) and COUP-TF3 and represses COUP-TF2-dependent activation of the atrial natriuretic factor promoter. J Biol Chem. 2001;276:28029–36. doi: 10.1074/jbc.M103577200. [DOI] [PubMed] [Google Scholar]
- 15.Ciullo M, et al. New susceptibility locus for hypertension on chromosome 8q by efficient pedigree-breaking in an Italian isolate. Hum Mol Genet. 2006;15:1735–43. doi: 10.1093/hmg/ddl097. [DOI] [PubMed] [Google Scholar]
- 16.Weatherford ET, Liu X, Sigmund CD. Regulation of renin expression by the orphan nuclear receptors Nr2f2 and Nr2f6. Am J Physiol Renal Physiol. 2012;302:F1025–33. doi: 10.1152/ajprenal.00362.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ladias JA, Karathanasis SK. Regulation of the apolipoprotein AI gene by ARP-1, a novel member of the steroid receptor superfamily. Science. 1991;251:561–5. doi: 10.1126/science.1899293. [DOI] [PubMed] [Google Scholar]
- 18.Ku DD, Guo L, Dai J, Acuff CG, Steinhelper ME. Coronary vascular and endothelial reactivity changes in transgenic mice overexpressing atrial natriuretic factor. Am J Physiol. 1996;271:H2368–76. doi: 10.1152/ajpheart.1996.271.6.H2368. [DOI] [PubMed] [Google Scholar]
- 19.Kim M, et al. GLP-1 receptor activation and Epac2 link atrial natriuretic peptide secretion to control of blood pressure. Nat Med. 2013;19:567–75. doi: 10.1038/nm.3128. [DOI] [PubMed] [Google Scholar]
- 20.Geurts AM, et al. Generation of gene-specific mutated rats using zinc-finger nucleases. Methods Mol Biol. 2010;597:211–25. doi: 10.1007/978-1-60327-389-3_15. [DOI] [PubMed] [Google Scholar]
- 21.Joe B, et al. Positional identification of variants of Adamts16 linked to inherited hypertension. Hum Mol Genet. 2009;18:2825–38. doi: 10.1093/hmg/ddp218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kumarasamy S, et al. Refined mapping of blood pressure quantitative trait loci using congenic strains developed from two genetically hypertensive rat models. Hypertens Res. 2011;34:1263–70. doi: 10.1038/hr.2011.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Morgan EE, et al. Validation of echocardiographic methods for assessing left ventricular dysfunction in rats with myocardial infarction. Am J Physiol Heart Circ Physiol. 2004;287:H2049–53. doi: 10.1152/ajpheart.00393.2004. [DOI] [PubMed] [Google Scholar]
- 24.Gopalakrishnan K, et al. Augmented rififylin is a risk factor linked to aberrant cardiomyocyte function, short-QT interval and hypertension. Hypertension. 2011;57:764–71. doi: 10.1161/HYPERTENSIONAHA.110.165803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pacurari M, et al. Endothelial cell transfusion ameliorates endothelial dysfunction in 5/6 nephrectomized rats. Am J Physiol Heart Circ Physiol. 2013;305:H1256–64. doi: 10.1152/ajpheart.00132.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hilgers RH, Todd J, Jr, Webb RC. Regional heterogeneity in acetylcholine-induced relaxation in rat vascular bed: role of calcium-activated K+ channels. Am J Physiol Heart Circ Physiol. 2006;291:H216–22. doi: 10.1152/ajpheart.01383.2005. [DOI] [PubMed] [Google Scholar]
- 27.Lu X, Kassab GS. Assessment of endothelial function of large, medium, and small vessels: a unified myograph. Am J Physiol Heart Circ Physiol. 2011;300:H94–H100. doi: 10.1152/ajpheart.00708.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mehrotra A, Joe B, de la Serna IL. SWI/SNF chromatin remodeling enzymes are associated with cardiac hypertrophy in a genetic rat model of hypertension. J Cell Physiol. 2013;228:2337–42. doi: 10.1002/jcp.24404. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.