

Abstract
Hispolon has been demonstrated to possess analgesic, anti-inflammatory and anticancer activities. However, whether hispolon prevents the invasion of breast carcinoma cells and the underlying mechanisms of its action remain unknown. In the present study, various assays, including a matrigel-based Transwell invasion assay and electrophoretic mobility shift assay, were used to investigate the anti-invasion effect of hispolon and explore its mechanism of action. The results revealed that hispolon inhibited the 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced migration and invasion of MDA-MB-231 human breast cancer cells at non-toxic concentrations. Hispolon also prevented the TPA-induced secretion of matrix metalloproteinase-9 (MMP-9) and reduced its expression at the transcriptional and translational levels. Furthermore, the phosphorylation of IκBα was reduced by hispolon, which resulted in the suppression of nuclear factor-κB (NF-κB), and p65 phosphorylation and nuclear translocation. An electrophoretic mobility shift assay demonstrated that NF-κB DNA-binding activity was induced by TPA and inhibited by hispolon. In addition, Bay 11–7082, which is a specific inhibitor of NF-κB, functioned in a similar manner as hispolon and blocked the secretion and expression of MMP-9. In conclusion, the results of the present study indicated that hispolon inhibited TPA-induced migration and invasion of MDA-MB-231 cells by reducing the secretion and expression of MMP-9 through the NF-κB signaling pathway.
Keywords: hispolon, invasion, matrix metalloproteinase-9, nuclear factor-κB, MDA-MB-231 cancer cells
Introduction
Breast cancer is the most frequently-diagnosed type of cancer in women and results in over half a million mortalities worldwide each year (1). Advanced-stage breast cancer can metastasize to other tissues of the body, such as the lymph nodes, reducing the effectiveness of radiation therapy or chemotherapy. Metastatic disease is responsible for >90% of cancer-associated mortalities (2). Thus, controlling or even reversing the process of metastasis would be a significant breakthrough in the treatment of cancer.
Metastasis is a complex cascade process, in which tumor cell invasion through the extracellular matrix (ECM) is one of the key early events. Matrix metalloproteinases (MMPs) are zinc-dependent endopeptidases that are able to degrade numerous types of ECM proteins (3). At present, 24 types of MMP have been identified, and MMP-9 (also termed gelatinase-B, a 92-kDa type IV collagenase) is considered to be essential in tumor invasion and migration, particularly in breast cancer (4). The expression and secretion of MMP-9 is regulated by various stimuli, including inflammatory cytokines, growth factors, tumor necrosis factor-α and 12-O-tetradecanoylphorbol-13-acetate (TPA) (5,6). TPA is a well-known inflammatory stimulus that induces tumor invasion and metastasis by directly activating protein kinase C isoforms (7). The activity of MMP-9 is controlled primarily at the transcriptional level. Nuclear factor-κB (NF-κB) is a transcription factor that binds to the promoter region of MMP-9 to regulate the expression of MMP-9 (8).
Hispolon was initially isolated as a yellow pigment from Inonotus hispidus and then identified in a number of traditional medicinal mushrooms, including Phellinus linteus and Phellinus igniarius, which have been used to treat various cancer types in East Asia (9–11). Hispolon has been reported to possess analgesic, anti-inflammatory and anticancer activities (10,12–14). A previous study demonstrated that hispolon may induce apoptosis in breast and bladder cancer cells via the MDM2-recruited ERK1/2 activity, and downregulate the level of MDM2 through the chaperone-mediated autophagy pathway (15,16). Human gastric cancer cells, rather than normal gastric cells, were selectively killed by hispolon due to the abrogation of the glutathione antioxidant system and the resultant excessive accumulation of reactive oxygen species (10). However, only a limited number of studies have investigated the antimetastatic effects of hispolon, while the underlying mechanism of the process has not been elucidated. In the present study, TPA-treated MDA-MB-231 human breast cancer cells were used to evaluate the antimetastatic potential of hispolon. In addition, the effect of hispolon treatment on the secretion and expression of MMP-9 at the transcriptional and translational levels, as well as the effect on the NF-κB signaling pathway, were investigated. Finally, the study examined whether hispolon can be used as an antimetastatic drug.
Materials and methods
Reagents and chemicals
Hispolon was synthesized as previously described (10,17), and its purity was detected on the basis of the NMR and mass spectra. A stock solution of 40 mM hispolon was prepared in dimethyl sulfoxide (DMSO) and diluted to the appropriate concentrations prior to use. The final concentration of DMSO was kept below 0.1% (v/v) in all assays. Doxorubicin (Dox), MTT, DMSO and Matrigel were purchased from Sigma-Aldrich (St. Louis, MO, USA). Bay 11–7082 (an NF-κB inhibitor) and TPA were purchased from Beyotime Institute of Biotechnology (Haimen, Jiangsu, China). Primary antibodies [polyclonal rabbit anti-human MMP-9 (catalog no., #3852); monoclonal mouse IgG1 anti-human IκBα (catalog no., #4814); monoclonal rabbit IgG anti-human phospho-IκBα (catalog no., #2859); monoclonal rabbit IgG anti-human NF-κB p65 (catalog no., #8242); monoclonal rabbit IgG anti-human phospho-NF-κB p65 (catalog no., #3033); monoclonal rabbit IgG anti-human histone H3 (catalog no., #4499); and monoclonal rabbit IgG anti-human β-actin (catalog no., #4970)], and horseradish peroxidase-conjugated secondary antibodies [goat anti-rabbit IgG (catalog no., #7074) and horse anti-mouse IgG (catalog no., #7076)] were obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Cell culture and reagents
The human MDA-MB-231 breast cancer cell line was obtained from the Institute of Biochemistry and Cell Biology (Chinese Academy of Sciences, Shanghai, China). Cells were cultured in Dulbecco's modified Eagle's medium (Gibco Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Gibco Life Technologies), 100 units/ml penicillin and 100 units/ml streptomycin (Beyotime Institute of Biotechnology). All cells were cultured in a humidified cell incubator with an atmosphere of 5% CO2 at 37°C and subcultured with 0.25% trypsin and 0.02% EDTA (Beyotime Institute of Biotechnology).
Cell viability assay
Briefly, cells were seeded in 96-well microtiter plates at a density of 5×103 cells/well and left for 24 h to adhere. The cells were cultured with the indicated concentrations of hispolon (0, 5, 10, 20, 40 or 60 µM) in the absence or presence of 160 nM TPA for 24 h. Next, the cells were incubated with MTT (0.5 mg/ml) for an additional 4 h at 37°C. The resulting formazan precipitate was dissolved in 150 µl DMSO and the absorbance was measured at 490 nm with a Sunrise microplate reader (Tecan Group Ltd, Männedorf, Switzerland). Dox was used as a positive control for all the assays. Each experiment was repeated at least three times.
Matrigel-based Transwell invasion assay
Cell invasion assays were carried out as previously described (5). Briefly, 24-well Transwell chambers with 8 µm polycarbonate nucleopore filters (Corning Incorporated, Corning, NY, USA) were coated with 20 µg/well Matrigel (Sigma-Aldrich). A total of 2×105 cells were seeded onto the upper part of the Matrigel-coated filter, and serum-free medium supplemented with 0–40 µM hispolon was incubated in the lower part for 2 h prior to the addition of 160 nM TPA. Following incubation at 37°C for 24 h, the cells that had invaded on the lower side of the membrane were fixed with 4% paraformaldehyde for 15 min, and then stained with 0.1% crystal violet (Beyotime Institute of Biotechnology)for 10 min. Subsequently, the stained cells were counted under a light microscope (BX51; Olympus Corporation, Tokyo, Japan). The percentage inhibition of invasive cells was expressed relative to the TPA alone-treated group. Each experiment was repeated at least three times.
Wound-healing assay
MDA-MB-231 cells were seeded in a 24-well plate (10,000 cells/well) and grown to 80–90% confluence. The monolayer of cells was scratched with white pipette tips, washed twice with phosphate-buffered saline and replaced with serum-free medium. Next, the cells were treated with the indicated concentrations of hispolon (0, 10, 20 or 40 µM) for 2 h and incubated with TPA for another 24 h. Images of the cell cultures were captured using a light microscope (BX51, Olympus Corporation, Tokyo, Japan) at 0 and 24 h. The migration of cells was assessed by counting the cells that crossed the blank lines and the percentage inhibition of the migration of the cells was expressed relative to the TPA only-treated group. Each experiment was repeated at least three times.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis
A total of 5×105 MDA-MB-231 cells/well were seeded in a 6-well plate and pretreated with hispolon at the indicated concentrations (0, 10, 20 or 40 µM) for 2 h, followed by TPA stimulation for a further 24 h. Total RNA was isolated with TRIzol reagent (Sangon Biotech Co., Ltd., Shanghai, China) according to the manufacturer's instructions, and the concentration of RNA was measured using a UV-2550 spectrophotometer (Shimadzu Corporation, Kyoto, Japan). A total of 2 µg RNA was converted to complementary DNA (cDNA) using a PrimeScript 1st Strand cDNA Synthesis kit (Takara Bio, Inc., Otsu, Japan) and the cDNA was amplified with Takara rTaq (Takara Bio, Inc.). The amplification conditions were one cycle of denaturation at 95°C for 3 min, followed by 33 cycles of 94°C for 15 sec, 60°C for 30 sec and 72°C for 60 sec, with one extension cycle at 72°C for 10 min. The primer sequences used were as follows: MMP-9 forward, 5′-CAACATCACCTATTGGATCC-3′, and reverse, 5′-CTGTAGAGTCTCTCGCT-3′; GAPDH forward, 5′-CATGTAGGCCATGAGGTCCACCAC-3′, and reverse, 5′-TGAAGGTCGGTGTGAACGGATTTGGC-3′. The PCR products were subjected to electrophoresis on a 1% (w/v) agarose gel. The bands were visualized by GoldView staining (SBS Genetech Co., Ltd., Beijing, China).
Western blot analysis
Cell lysates were obtained and protein concentrations were determined using the BCA protein assay kit (Pierce Biotechnology, Inc., Rockford, IL, USA). Equal quantities of proteins (30–100 µg protein/lane) were subjected to 10–15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto nitrocellulose membranes (GE Healthcare Bio-Sciences, Pittsburgh, PA, USA). The membranes were blocked in 5% (w/v) nonfat dry milk for 1 h and incubated with polyclonal rabbit anti-human MMP-9 (1:1,000), monoclonal mouse anti-human IκBα (1:1,000), monoclonal rabbit anti-human p-IκBα (1:1,000), monoclonal rabbit anti-human p65 (1:2,000), monoclonal rabbit anti-human p-p65 (1:1,000), monoclonal rabbit anti-human histone H3 (1:2,000), monoclonal rabbit anti-human GAPDH (1:1,000) and monoclonal rabbit anti-human β-actin (1:1,000) primary antibodies at 4°C overnight. The membranes were then incubated with secondary goat anti-rabbit (1:2,000) or horse anti-mouse (1:8,000) IgG antibodies for 1 h. Bands were detected using the SuperSignal™ West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Waltham, MA, USA).
Gelatin zymography
MDA-MB-231 cells were incubated in serum-free medium pretreated with the indicated concentrations of hispolon or Bay 11–7082 for 2 h, followed by addition of 160 nM TPA to the medium and incubation for 24 h. Next, the conditioned medium was collected. Equal volumes were mixed with non-reducing sample buffer and subjected to 10% SDS-PAGE containing 0.1% (w/v) gelatin at 4°C. The gel was washed four times with denaturing buffer (2.5% Triton X-100, pH 7.6, 50 mM Tris-HCl, 5 mM CaCl2 and 1 µM ZnCl2) and equilibrated twice in developing buffer (pH 7.6, 50 mM Tris-HCl, 5 mM CaCl2 and 1 µM ZnCl2). Subsequently, the gel was incubated with the fresh developing buffer for 24 h at 37°C and stained with 0.05% coomassie blue R-250 (Sangon Biotech Co., Ltd.). The MMP-9 gelatinolytic activity was measured by clear bands against the blue background.
Electrophoretic mobility shift assay (EMSA)
MDA-MB-231 cells were pretreated with 0–40 µM hispolon for 2 h, followed by treatment with or without 160 nM TPA for 2 h. The cultured cells were collected and the nuclear proteins were prepared using a Nuclear Protein Extraction kit (Beyotime Institute of Biotechnology). The double-stranded oligonucleotides (Beyotime Institute of Biotechnology) containing the consensus sequence for NF-κB (5′-AGTTGAGGGGACTTTCCCAGGC-3′) were end labeled with biotin and used as probes for the EMSA. Nuclear extracts (10 µg) were incubated with the binding buffer (10 mM Tris-HCl, pH 7.5, 50 mM NaCl, 0.5 mM dithiothreitol, 0.5 mM EDTA, 2.5% (v/v) glycerol and 1 µg/µl poly dI-dC) for 10 min and then mixed with 0.5 pmol-labeled probes for 20 min at room temperature. The DNA-protein complex was separated on a 6% native polyacrylamide gel in 0.5X Tris/borate/EDTA buffer at 4°C and transferred onto positively charged nitrocellulose membranes (GE Healthcare Bio-Sciences). The competition assay was performed by adding 100-fold excess of an unlabeled oligonucleotide as a specific competitor. Bands were detected using the SuperSignal™ West Pico Chemiluminescent Substrate, as described by the manufacturer.
Statistical analysis
The results are expressed as the mean ± standard deviation. Analysis was performed with Excel 2007 software (Microsoft, Seattle, WA, USA) using a Student's two-tailed t-test or one-way analysis of variance, in which P<0.05 vs. control was considered to indicate a statistically significant difference.
Results
Effect of hispolon on the proliferation of MDA-MB-231 cells
Hispolon is an active polyphenolic compound (Fig. 1A). The effect of hispolon on the viability of MDA-MB-231 cells was evaluated. Hispolon demonstrated non-significant toxicity on TPA-treated and untreated MDA-MB-231 cells at concentrations between 0 and 40 µM for 24 h (Fig. 1B). To measure the anti-invasive effect of hispolon, non-toxic concentrations (0–40 µM) were used for the subsequent experiments, avoiding the interference resulting from the antiproliferative activity of hispolon.
Figure 1.
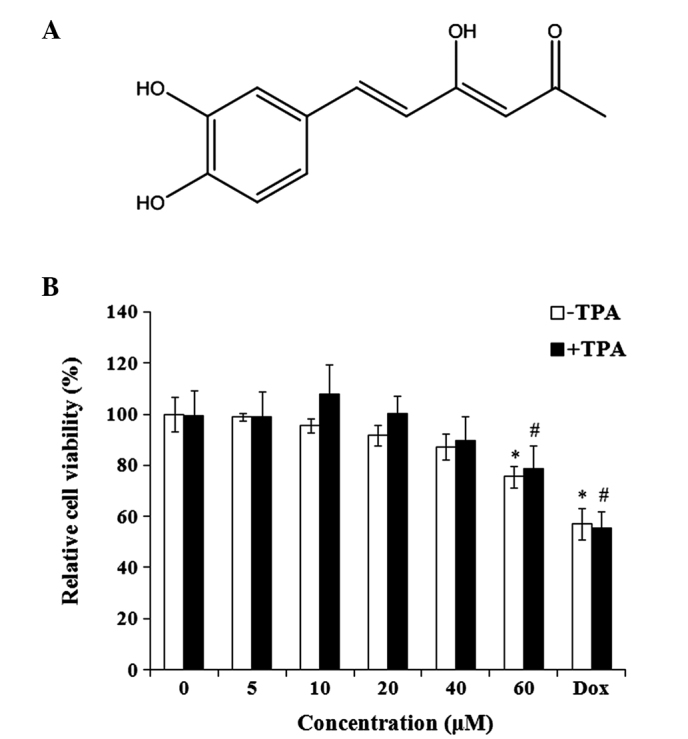
(A) Chemical structure of hispolon. (B) Effects of hispolon on cell viability. MDA-MB-231 cells were treated with 0–60 µM hispolon and/or TPA (160 nM) for 24 h, and the cell viability was measured by an MTT assay. Dox was used as a positive control. All the data are presented as the mean ± standard deviation of three independent experiments. *P<0.05 vs. DMSO control; #P<0.05 vs. TPA alone-treated group. TPA, 12-O-tetradecanoylphorbol-13-acetate; Dox, doxorubicin.
Hispolon inhibits TPA-induced migration and invasion of MDA-MB-231 cells
A wound-healing assay and a Matrigel-based Transwell invasion assay were performed in TPA-induced MDA-MB-231 cells. TPA treatment resulted in a marked increase in cell invasion and migration, while hispolon inhibited the TPA-induced cell invasion and migration in a dose-dependent manner (Fig. 2A and B). A concentration of 40 µM hispolon inhibited the cell invasion and migration significantly, and the levels of cell invasion and migration in the 40 µM hispolon-treated group were similar to the DMSO control group (Fig. 2C). Collectively, these results indicate that hispolon effectively prevented TPA-induced migration and invasion in the MDA-MB-231 cells.
Figure 2.

Hispolon inhibited TPA-induced migration and invasion of MDA-MB-231 cells. (A) MDA-MB-231 cells were treated with hispolon for 2 h followed by TPA (160 nM) treatment for 24 h. The anti-invasive properties of hispolon were detected by Matrigel-based Transwell invasion assay. (B) Cells were treated as described previously and the antimigration ability was detected by wound-healing assay. (C) Relative quantification of the cell invasion and migration. All the data are presented as the mean ± standard deviation of three independent experiments. *P<0.05 and #P<0.05, vs. TPA alone-treated group; **P<0.05 and ##P<0.05 vs. DMSO control. TPA, 12-O-tetradecanoylphorbol-13-acetate; DMSO, dimethyl sulfoxide; TPA+H-10, 160 nM TPA combined with 10 µM hispolon; TPA+H-20, 160 nM TPA combined with 20 µM hispolon; TPA+H-40, 160 nM TPA combined with 40 µM hispolon.
Hispolon inhibits TPA-induced MMP-9 expression and secretion
A previous study demonstrated that MMP-9 is an important ECM-degrading enzyme and that activation of MMP-9 may contribute to the process of cancer metastasis (18,19). In the present study, the effects of hispolon treatment on TPA-induced MMP-9 expression and secretion in MDA-MB-231 cells were investigated. Hispolon was found to significantly reduce TPA-induced MMP-9 gelatinolytic activity as assessed by gelatin zymography (Fig. 3A). In addition, western blot analysis of the conditioned medium demonstrated that hispolon inhibited the secretion of MMP-9 elicited by TPA in a dose-dependent manner. Furthermore, analysis of the whole cell lysates demonstrated that hispolon treatment resulted in a reduction in TPA-induced intracellular expression of MMP-9. It should be noted that the precursor (85 kDa, lower band) and mature forms (92 kDa, upper band) of MMP-9 were detected in cell lysates (Fig. 3A).
Figure 3.

Hispolon inhibited TPA-induced MMP-9 activation, as well as inhibited the secretion and expression in a dose-dependent manner. (A) Cells in serum-free medium were pretreated with hispolon at the indicated concentrations for 2 h prior to TPA incubation for another 24 h. Conditioned media were analyzed by gelatin zymography (upper panel) and western blot analysis (middle panel). Cell lysates were collected and subjected to western blot analysis (lower panel). β-actin was used as an internal control. (B) Hispolon inhibited TPA-induced MMP-9 gene expression. MMP-9 expression was detected by RT-PCR and GAPDH was used as an internal control. (C) Relative quantification of MMP-9 gene expression. All the data are presented as the mean ± standard deviation of three independent experiments. *P<0.05 vs. TPA alone-treated group; #P<0.05 vs. DMSO control. TPA, 12-O-tetradecanoylphorbol-13-acetate; MMP-9, matrix metalloproteinase-9; RT-PCR, reverse transcription-polymerase chain reaction; DMSO, dimethyl sulfoxide.
RT-PCR was then used to investigate the effect of hispolon treatment on the regulation of TPA-induced MMP-9 transcription. Hispolon treatment resulted in a reduction in the MMP-9 mRNA expression levels in a dose-dependent manner (Fig. 3B). The relative quantity of MMP-9 mRNA indicated that 40 µM hispolon suppressed TPA-induced MMP-9 gene expression by 90%. In addition, the mRNA expression of MMP-9 in the 40 µM hispolon-treated group was reduced compared with the DMSO control group (Fig. 3C). These results demonstrated that hispolon inhibited TPA-induced MMP-9 expression and secretion.
Hispolon inhibits the TPA-induced NF-κB signaling pathway
Previous studies have demonstrated that NF-κB is important in TPA-induced MMP-9 gene transcription and promotes breast cancer cell migration and metastasis (16,17). The effects of hispolon on the NF-κB signaling pathway were also assessed in the present study (Fig. 4). The activity of NF-κB is primarily regulated by the inhibitor of κBα (IκBα) protein. Western blot analysis demonstrated that TPA-induced phosphorylation of IκBα was prevented by hispolon treatment in a dose-dependent manner (Fig. 4A). Hispolon also resulted in a reduction in phosphorylated NF-κB p65 induced by TPA, while the level of total p65 remained the same in whole cell lysates. Phosphorylation of IκBα and p65 is critical for p65 nuclear translocation; thus, the effect of hispolon treatment on p65 nuclear translocation was assessed via western blot analysis of the nuclear protein fraction. TPA induced p65 translocation to the nucleus, and translocation was markedly reduced by 40 µM hispolon treatment (Fig. 4C).
Figure 4.

Hispolon inhibited the TPA-induced NF-κB signaling pathway. (A) Hispolon inhibited the phosphorylation of IκBα and p65. Cells were pretreated with hispolon for 2 h prior to TPA incubation and then for another 2 h. Cell lysates were collected and subjected to western blot analysis for total IκBα, total p65, phospho-IκBα and phospho-p65 proteins. β-actin was used as an internal control. (B) Relative quantity of phosphorylated IκBα and p65 expression levels. *P<0.05 and #P<0.05 vs. TPA alone-treated group, **P<0.05 and ##P<0.05 vs. DMSO control. All the data are presented as the mean ± standard deviation of three independent experiments. (C) Hispolon prevented NF-κB p65 nuclear translocation. Cells were preincubated with hispolon for 2 h prior to TPA treatment for another 2 h. Nuclear fractions were collected and histone H3 was used to verify equal loading. (D) Hispolon suppressed TPA-induced NF-κB DNA-binding activity. MDA-MB-231 cells were treated as described in part (A). Nuclear extracts (10 µg) were incubated with biotin-labeled oligonucleotides containing the NF-κB motif of the MMP-9 promoter. Bound mixtures were separated by 6% native-polyacrylamide electrophoresis. (E) Inhibitory effect of Bay 11–7082 on TPA-induced phosphorylations of IκBα and p65. Cells were pretreated with 5 µM Bay 11–7082 for 2 h and then stimulated with TPA for 2 h. (F) Inhibitory effect of Bay 11–7082 on TPA-induced MMP-9 expression. Cells in serum-free medium were pretreated with 2.5 or 5 µM Bay 11–7082 for 2 h and then stimulated with TPA for 24 h. Cell lysates were analyzed by western blot analysis. β-actin was used as an internal control. (G) Inhibitory effect of Bay 11–7082 on TPA-induced MMP-9 secretion. Cells in serum-free medium were preincubated with 5 µM Bay 11–7082 for 2 h, followed by treatment with TPA for 24 h. (H) Schematic diagram of the molecular mechanisms underlying the antimetastasis activity of hispolon in TPA-induced MDA-MB-231 cells. TPA, 12-O-tetradecanoylphorbol-13-acetate; DMSO, dimethyl sulfoxide; NF-κB, nuclear factor-κB; MMP-9, matrix metalloproteinase-9; Comp., competitor.
The EMSA assay results demonstrated that hispolon significantly reduced TPA-induced NF-κB DNA binding activity (Fig. 4D). The specificity of NF-κB binding ability was confirmed using a competition assay with excess unlabeled NF-κB probes. Bay 11–7082, an NF-κB-specific inhibitor, inhibited the activity of MMP-9 and the phosphorylation of IκBα and p65, which was in accordance with the results obtained following hispolon treatment (Fig. 4E-G). Therefore, these findings indicate that hispolon prevented TPA-induced MMP-9 gene expression by blocking the NF-κB signaling pathway in MDA-MB-231 cells.
Discussion
In China, traditional Chinese medicine has been used to treat patients for centuries. Hispolon is an active compound isolated from the fungi of Phellinus linteus (10), which is a traditional Chinese medicine and is also known as ‘Sanghuang’. Extracts of Phellinus linteus have previously been reported to have immunostimulatory, anti-inflammatory and anticancer activities (10,20–22). Numerous studies have demonstrated that hispolon induces apoptosis in human cancer cells (13–16,22). In the present study, a non-toxic concentration of hispolon that prevented TPA-induced migration and invasion of MDA-MB-231 cells was established. The anticancer effects of hispolon may, therefore, be attributed to its ability to induce apoptosis in cancer cells in addition to its antimetastatic activities.
Overexpression of MMP-9 in human breast cancer has been previously observed, and hispolon has been demonstrated to prevent the metastasis of SK-Hep1 hepatoma cells by inhibiting MMP-2/9 expression (14,23). The present study also indicated that hispolon suppressed the secretion and expression of MMP-9 in a dose-dependent manner in TPA-induced MDA-MB-231 cells. The mature form of MMP-9 (92 kDa, upper band) possessed gelatinolytic activity and was detected outside the cells, while the precursor form (85 kDa, lower band) remained inside the cells; these findings were consistent with the results of previous studies (24,25). Transcriptional regulation of MMP-9 has been proposed to be vital in its activation (26). The present study demonstrated that hispolon treatment reduced the MMP-9 mRNA expression levels in TPA-treated and untreated MDA-MB-231 cells.
NF-κB is a critical transcription factor in the regulation of MMP-9 gene expression and can be activated by TPA (25). The present study investigated whether hispolon treatment inhibited TPA-induced MMP-9 activity through the NF-κB signaling pathway. When IκBα is phosphorylated it is degraded by the proteasome, which is required for the activation of NF-κB; in the present study, hispolon treatment resulted in a reduction in this process. Phosphorylation of p65, which resulted in an optimal induction of NF-κB target genes, was also reduced by hispolon treatment. The nuclear fraction was isolated, while the nuclear translocation of p65 and NF-κB DNA-binding activity were demonstrated to be blocked by hispolon. A specific NF-κB inhibitor, Bay 11–7082, was then used to confirm the effects of the NF-κB signaling pathway in the metastasis of TPA-induced MDA-MB-231 cells. Similar to treatment with hispolon, the secretion and expression of MMP-9 were prevented by Bay 11–7082. These results indicate that NF-κB may be an upstream regulator of MMP-9 and that hispolon inhibited TPA-induced metastasis of MDA-MB-231 cells via the NF-κB signaling pathway.
In conclusion, the present study initially determined non-toxic concentrations of hispolon that inhibited TPA-induced migration and invasion in MDA-MB-231 human breast cancer cells. Hispolon treatment was found to suppress the secretion and expression of MMP-9 by blocking the activation of the transcription factor, NF-κB (summarized in Fig. 4H). Therefore, hispolon, which is regarded as a natural product, is notable for further development as a potential antimetastasis agent for clinical use.
Acknowledgements
The authors would like to thank Dr Yong-Ting Luo (Institute of Biophysics, Chinese Academy of Sciences, Beijing, China) for his helpful discussion and editing of this manuscript. This work was supported by a grant from the Natural Science Foundation of Zhejiang Province (no. LQ14H160016).
References
- 1.Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63:11–30. doi: 10.3322/caac.21166. [DOI] [PubMed] [Google Scholar]
- 2.Valastyan S, Weinberg RA. Tumor metastasis: molecular insights and evolving paradigms. Cell. 2011;147:275–292. doi: 10.1016/j.cell.2011.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lin CW, Hou WC, Shen SC, et al. Quercetin inhibition of tumor invasion via suppressing PKC delta/ERK/AP-1-dependent matrix metalloproteinase-9 activation in breast carcinoma cells. Carcinogenesis. 2008;29:1807–1815. doi: 10.1093/carcin/bgn162. [DOI] [PubMed] [Google Scholar]
- 4.Pellikainen JM, Ropponen KM, Kataja VV, et al. Expression of matrix metalloproteinase (MMP)-2 and MMP-9 in breast cancer with a special reference to activator protein-2, HER2, and prognosis. Clin Cancer Res. 2004;10:7621–7628. doi: 10.1158/1078-0432.CCR-04-1061. [DOI] [PubMed] [Google Scholar]
- 5.Laulan NB, St-Pierre Y. Bone morphogenetic protein 4 (BMP-4) and epidermal growth factor (EGF) inhibit metalloproteinase-9 (MMP-9) expression in cancer cells. Oncoscience. 2015;2:309–316. doi: 10.18632/oncoscience.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Roomi MW, Monterrey JC, Kalinovsky T, Rath M, Niedzwiecki A. Patterns of MMP-2 and MMP-9 expression in human cancer cell lines. Oncol Rep. 2009;21:1323–1333. doi: 10.3892/or_00000358. [DOI] [PubMed] [Google Scholar]
- 7.Han H, Du B, Pan X, et al. CADPE inhibits PMA-stimulated gastric carcinoma cell invasion and matrix metalloproteinase-9 expression by FAK/MEK/ERK-mediated AP-1 activation. Mol Cancer Res. 2010;8:1477–1488. doi: 10.1158/1541-7786.MCR-10-0114. [DOI] [PubMed] [Google Scholar]
- 8.Park SK, Hwang YS, Park KK, et al. Kalopanaxsaponin A inhibits PMA-induced invasion by reducing matrix metalloproteinase-9 via PI3K/Akt- and PKCdelta-mediated signaling in MCF-7 human breast cancer cells. Carcinogenesis. 2009;30:1225–1233. doi: 10.1093/carcin/bgp111. [DOI] [PubMed] [Google Scholar]
- 9.Ali NAA, Pilgrim H, Liberra K, Lindequist U, Jansen R. Hispolon, a yellow pigment from Inonotus hispidus. Phytochemistry. 1996;41:927–929. doi: 10.1016/0031-9422(95)00717-2. [DOI] [Google Scholar]
- 10.Chen W, Zhao Z, Li L, Wu B, Chen SF, Zhou H, Wang Y, Li YQ. Hispolon induces apoptosis in human gastric cancer cells through a ROS-mediated mitochondrial pathway. Free Radic Biol Med. 2008;45:60–72. doi: 10.1016/j.freeradbiomed.2008.03.013. [DOI] [PubMed] [Google Scholar]
- 11.Mo S, Wang S, Zhou G, et al. Phelligridins C-F: cytotoxic pyrano[4,3-c][2]benzopyran-1,6-dione and furo[3,2-c]pyran-4-one derivatives from the fungus Phellinus igniarius. J Nat Prod. 2004;67:823–828. doi: 10.1021/np030505d. [DOI] [PubMed] [Google Scholar]
- 12.Chang HY, Sheu MJ, Yang CH, Lu TC, Chang YS, Peng WH, Huang SS, Huang GJ. Analgesic effects and the mechanisms of anti-inflammation of hispolon in mice. Evid Based Complement Alternat Med. 2011;2011:478246. doi: 10.1093/ecam/nep027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huang GJ, Deng JS, Huang SS, et al. Hispolon induces apoptosis and cell cycle arrest of human hepatocellular carcinoma Hep3B cells by modulating ERK phosphorylation. J Agric Food Chem. 2011;59:7104–7113. doi: 10.1021/jf201289e. [DOI] [PubMed] [Google Scholar]
- 14.Huang GJ, Yang CM, Chang YS, et al. Hispolon suppresses SK-Hep1 human hepatoma cell metastasis by inhibiting matrix metalloproteinase-2/9 and urokinase-plasminogen activator through the PI3K/Akt and ERK signaling pathways. J Agric Food Chem. 2010;58:9468–9475. doi: 10.1021/jf101508r. [DOI] [PubMed] [Google Scholar]
- 15.Lu TL, Huang GJ, Lu TJ, et al. Hispolon from Phellinus linteus has antiproliferative effects via MDM2-recruited ERK1/2 activity in breast and bladder cancer cells. Food Chem Toxicol. 2009;47:2013–2021. doi: 10.1016/j.fct.2009.05.023. [DOI] [PubMed] [Google Scholar]
- 16.Lu TL, Huang GJ, Wang HJ, et al. Hispolon promotes MDM2 downregulation through chaperone-mediated autophagy. Biochem Biophys Res Commun. 2010;398:26–31. doi: 10.1016/j.bbrc.2010.06.004. [DOI] [PubMed] [Google Scholar]
- 17.Venkateswarlu S, Ramachandra MS, Sethuramu K, Subbaraju GV. Synthesis and antioxidant activity of hispolon, a yellow pigment from Inonotus hispidius. J Chem Sect B. 2002;41:875–877. [Google Scholar]
- 18.Helbig G, Christopherson KW, II, Bhat-Nakshatri P, et al. NF-kappaB promotes breast cancer cell migration and metastasis by inducing the expression of the chemokine receptor CXCR4. J Biol Chem. 2003;278:21631–21638. doi: 10.1074/jbc.M300609200. [DOI] [PubMed] [Google Scholar]
- 19.Shin Y, Yoon SH, Choe EY, et al. PMA-induced up-regulation of MMP-9 is regulated by a PKCalpha-NF-kappaB cascade in human lung epithelial cells. Exp Mol Med. 2007;39:97–105. doi: 10.1038/emm.2007.11. [DOI] [PubMed] [Google Scholar]
- 20.Kim GY, Oh YH, Park YM. Acidic polysaccharide isolated from Phellinus linteus induces nitric oxide-mediated tumoricidal activity of macrophages through protein tyrosine kinase and protein kinase C. Biochem Biophys Res Commun. 2003;309:399–407. doi: 10.1016/j.bbrc.2003.08.018. [DOI] [PubMed] [Google Scholar]
- 21.Sliva D, Jedinak A, Kawasaki J, Harvey K, Slivova V. Phellinus linteus suppresses growth, angiogenesis and invasive behaviour of breast cancer cells through the inhibition of AKT signalling. Br J Cancer. 2008;98:1348–1356. doi: 10.1038/sj.bjc.6604319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hsiao PC, Hsieh YH, Chow JM, et al. Hispolon induces apoptosis through JNK1/2-mediated activation of a caspase-8, -9, and -3-dependent pathway in acute myeloid leukemia (AML) cells and inhibits AML xenograft tumor growth in vivo. J Agric Food Chem. 2013;61:10063–10073. doi: 10.1021/jf402956m. [DOI] [PubMed] [Google Scholar]
- 23.Scorilas A, Karameris A, Arnogiannaki N, et al. Overexpression of matrix-metalloproteinase-9 in human breast cancer: a potential favourable indicator in node-negative patients. Br J Cancer. 2001;84:1488–1496. doi: 10.1054/bjoc.2001.1810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Toth M, Gervasi DC, Fridman R. Phorbol ester-induced cell surface association of matrix metalloproteinase-9 in human MCF10A breast epithelial cells. Cancer Res. 1997;57:3159–3167. [PubMed] [Google Scholar]
- 25.Ling H, Yang H, Tan SH, Chui WK, Chew EH. 6-Shogaol, an active constituent of ginger, inhibits breast cancer cell invasion by reducing matrix metalloproteinase-9 expression via blockade of nuclear factor-κB activation. Br J Pharmacol. 2010;161:1763–1777. doi: 10.1111/j.1476-5381.2010.00991.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jones CB, Sane DC, Herrington DM. Matrix metalloproteinases: a review of their structure and role in acute coronary syndrome. Cardiovasc Res. 2003;59:812–823. doi: 10.1016/S0008-6363(03)00516-9. [DOI] [PubMed] [Google Scholar]