

Abstract
In meiosis, homologous chromosomes face the obstacle of finding, holding onto and segregating away from their partner chromosome. There is increasing evidence, in a diverse range of organisms, that centromere–centromere interactions that occur in late prophase are an important mechanism in ensuring segregation fidelity. Centromere pairing appears to initiate when homologous chromosomes synapse in meiotic prophase. Structural proteins of the synaptonemal complex have been shown to help mediate centromere pairing, but how the structure that maintains centromere pairing differs from the structure of the synaptonemal complex along the chromosomal arms remains unknown. When the synaptonemal complex proteins disassemble from the chromosome arms in late prophase, some of these synaptonemal complex components persist at the centromeres. In yeast and Drosophila these centromere-pairing behaviors promote the proper segregation of chromosome partners that have failed to become linked by chiasmata. Recent studies of mouse spermatocytes have described centromere pairing behaviors that are similar in several respects to what has been described in the fly and yeast systems. In humans, chromosomes that fail to experience crossovers in meiosis are error-prone and are a major source of aneuploidy. The finding that centromere pairing is a conserved phenomenon raises the possibility that it may play a role in promoting the segregation fidelity of non-exchange chromosome pairs in humans.
Keywords: achiasmate segregation, bi-orientation, centromere pairing, DNA threads, heterochromatin, meiosis, synaptonemal complex
Introduction
In meiosis, diploid cells duplicate their DNA and then undergo two rounds of chromosome segregation. This results in the production of haploid meiotic products. In most organisms studied, proper movement of the chromosomes away from their partners at anaphase I is dependent on the homologs first becoming tethered to one another in meiotic prophase. Homologs can become tethered in a number of ways. In prophase of most organisms, each homolog pair becomes linked by one or more chiasmata, connections between the axes of the chromosomes that occur at sites of genetic exchange, or crossovers (see accompanying minireview by Sansam and Pezza 1; reviewed in 2) (Fig. 1).
Figure 1.
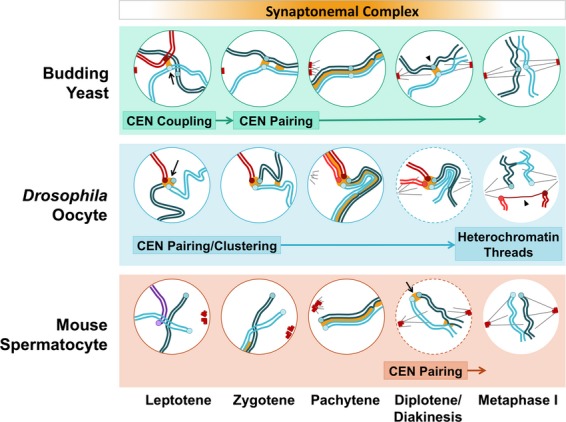
Centromere behaviors in meiotic prophase in yeast, female Drosophila and male mouse. The synaptonemal complex (SC) proteins are implicated in the many interactions between centromeres; see text for details. The orange gradient at the top indicates the period through which SC proteins load onto and disassemble from the chromosomes, with exceptions noted in the text.
The linkages provided by chiasmata are critical for the segregation of chromosomes in meiosis I for three reasons. First, the tethers provided by chiasmata in conjunction with sister chromatid cohesion help partners to remain associated from prophase I, when partners are identified, until the segregation process begins (see accompanying minireview by Rankin 3). This is particularly important in mammalian oocytes in which the pairing (prophase) and segregation (anaphase) stages of meiosis I can be separated by years 4. Second, tethers help prevent precocious sister chromatid separation. When there is no crossover, in fission yeast and mice oocytes, the cell reacts by splitting sister chromatids to presumably satisfy the spindle checkpoint in the absence of a partner chromosome 5–7 (see the accompanying minireview by Gorbsky 8). Third, chiasmata contribute to the process by which the kinetochores attach to microtubules in such a way that the homologs will be pulled to the opposite rather than the same side of the spindle at anaphase I. Correct attachment of the homologous kinetochores to microtubules from opposite poles creates tension at the microtubule–kinetochore interface (Fig. 2), which in turn stabilizes the connections 9. This tension is counteracted by the chiasmata.
Figure 2.
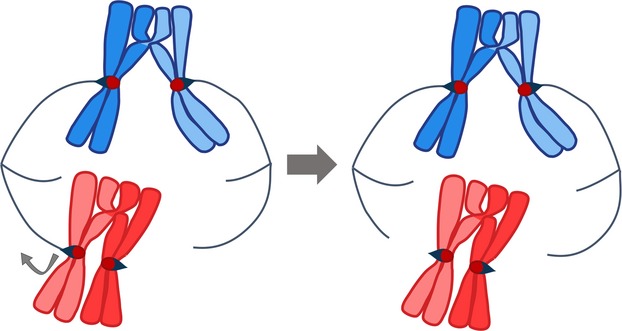
Kinetochore–microtubule attachments. Bi-oriented attachment to the spindle creates tension at the kinetochore–microtubule interface and stabilizes the kinetochore–microtubule interaction (blue chromosomes). When chromosomes are only attached to one pole, or when both kinetochores are attached to the same pole (not shown), the kinetochore–microtubule attachments are not stabilized by tension. These attachments are released, which allows for a second try at bi-orientation (red chromosomes).
These critical roles for chiasmata would predict dire consequences in meiosis for mutants that fail to form proper chiasmata. Indeed, in budding yeast and other fungi, nematodes, Drosophila, mice and Arabidopsis mutations that block the initiation of crossover formation result in sterility or greatly reduced fertility 10–20. This infertility can be due either to the production of aneuploid gametes or to checkpoint mediated arrests in gametogenesis (see accompanying minireviews by Gorbsky 8 and Sansam and Pezza 1).
Despite the clear importance of chiasmata in mediating pairing and segregation of homologous chromosomes in meiosis I, there are many recognized examples of organisms that partition achiasmate chromosome pairs in meiosis. How is this accomplished? In nearly all instances the achiasmate pair becomes tethered, but by a mechanism that does not involve conventional crossover and chiasma formation. There is emerging evidence (see below) that some of these mechanisms might work in conjunction with chiasma to accomplish high fidelity chromosome segregation.
Achiasmate mechanisms sometimes involve the entire chromosome set 21–23, while in some organisms only particular chromosome partners are achiasmate 24–32. The tethering of achiasmate chromosomes can extend along entire chromosomes at designated pairing sites, as in male Drosophila 33, or at unique sites on a chromosome 31,34,35. The focus of this review is the pairing of chromosomes at their centromeres in ways that promote the segregation of achiasmate partners in meiosis I and could potentially act to improve the segregation of chiasmate chromosomes as well.
Early observations of CEN–CEN interactions
The initial observation that centromeres can actively become associated with one another was in cells at earlier stages of meiosis, before synapsis 36–45 (for review see 46,47). Early meiotic centromere–centromere (CEN–CEN) associations were first documented in the onion Allium fistulosum using electron microscopy 36. These CEN–CEN interactions were apparent to investigators because they were occurring in early stages of prophase before other types of chromosomal associations were evident. The studies showed that centromeres clustered in groups of two or a few centromeres prior to synapsis. In addition, it was found that these pairs and clusters included associations between non-homologous centromeres. Subsequent studies in wheat, yeast, rice, maize and mouse described similar associations of centromeres or associated heterochromatin 37–45; however, in some of these organisms the CEN–CEN interactions appeared to be restricted to pairs rather than small groups. In most of these studies it was possible to determine whether the CEN–CEN interactions were between homologous partners; indeed, in every case that homology was tested, it appeared that pairing partner choice was homology independent.
The discovery of homology-independent interactions of early prophase in yeast 39 made it possible to more specifically probe the timing of CEN–CEN associations and identify pairing partners. CEN–CEN interactions were shown to occur in a homology-independent fashion that initiated in early prophase just after DNA replication (Fig. 1, budding yeast, arrow). These associations gave way to alignment of homologous centromeres later in prophase 39,40,48. These early CEN–CEN associations, termed CEN-coupling, were found to be dependent on the synaptonemal complex (SC) protein Zip1 (Fig. 3), which was shown to localize to the coupled centromeres prior to SC assembly 39. Hoffman, Hochwagen and colleagues have used a zip1-S75E coupling-deficient mutant to demonstrate that the formation of crossovers and synapsis do not depend on coupling 48. While investigations of CEN-coupling have not revealed its function, they have demonstrated that there are mechanisms that allow the formation of persistent associations of centromeres. Furthermore, the fact that the associations are homology independent demonstrates that they depend upon centromeric chromatin rather than DNA.
Figure 3.
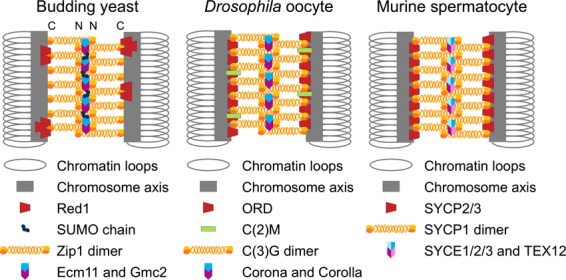
Synaptonemal complex in yeast, Drosophila and mice. Models of budding yeast, Drosophila oocytes and murine spermatocyte synaptonemal complexes. The synaptonemal complex (SC) forms a bridge between the axes of the two homologous chromosomes (gray bars and loops). The lateral elements (LEs), containing the proteins indicated in red, extend along the chromosome axes (gray bars). Recent high resolution imaging suggests that in yeast Red1 is in distinct foci distributed along the axes 120. In budding yeast SUMOylation of the Ecm11 central element protein appears to promote Zip1 assembly 68. Drosophila oocyte LEs contain an additional protein, C(2)M (green), that is discontinuous along the SC. The transverse filament is depicted as an orange/yellow dimer in each species. It forms an anti-parallel tetramer zipper structure that bridges the two LEs (see N and C designation in the budding yeast diagram, indicating the N terminus and the C terminus). The central elements of each species are depicted in blue/purple. The transverse filament and central element, together, form the central region (CR). For a more detailed overview of SC structure see 63.
Discovery of CEN-pairing
A form of CEN–CEN association that occurs later in meiotic prophase, which we shall refer to as CEN-pairing, was identified by efforts to understand the basis of achiasmate chromosome segregation in Drosophila females. The evidence that, in an organism with meiotic crossing-over, individual chromosome pairs could be correctly segregated from one another without chiasmata was provided in 1936 from genetic studies of meiosis in Drosophila females 24. This study found that in oocytes homologous X chromosomes often failed to recombine, but segregated well regardless. The same was found to be true for the fourth chromosome pair 26,27. Nearly 50 years after the recognition of achiasmate segregation, a key genetic study suggested that the pairing of centromere proximal regions might direct the process 49. Investigators tested the ability of an additional chromosome in the cell, a duplication chromosome composed of regions of chromosome 4, to disrupt the normally accurate segregation of the natural chromosome 4 pair. The ability of a duplication chromosome to interfere with the chromosome 4 pair correlated directly with whether it carried regions normally found in chromosome 4 pericentric heterochromatin. This and other experiments suggested to the authors that ‘this chromatin may well be or contain a meiotic pairing site’ 49. This notion was confirmed by further work showing that a portion of centric heterochromatin, about 420 kb in length, conferred high fidelity segregation to mini-chromosomes 50,51. Further, cytological experiments 34 demonstrated that achiasmate X and fourth chromosome pairs establish associations in prophase between large blocks of heterochromatin that extend outward from the centromeres. This heterochromatic pairing was shown to persist until metaphase when the chromosome pairs begin to attach to the meiotic spindle.
Studies in yeast and mice have suggested that CEN-pairing may be a conserved phenomenon. Work done in budding yeast demonstrated that chromosomes lacking chiasmata were still able to segregate away from each other in anaphase I, though not nearly with the fidelity of exchange partners or achiasmate partners in Drosophila females 28–30. These studies used achiasmate centromere plasmids, homeologous chromosomes with a crossover frequency of 1 in 2500 meioses 52,53, artificial mini-chromosomes and natural non-homologous chromosome pairs to show unexpectedly high levels (∼ 75–90%) of proper segregation 35,53–57. Some of these studies went on to show that this pairing was mediated by a persistence of the SC (including Zip1) at the centromeres after SC disassembly, and this pairing was essential for achiasmate chromosomes to segregate properly 54,55. It is important to note in the yeast experiments using Saccharomyces carlsbergensis or Saccharomyces paradoxus homeologous chromosomes that pairing partners were shown to be obligate non-exchange chromosomes; and, in fact, increasing the rate of crossovers by impairing the mismatch repair system actually increased segregation errors 52,53,56. Whereas in Drosophila females the centromere pairing occurs preferentially between homologous blocks of peri-centric heterochromatin 49, in yeast CEN-pairing does not require homology of the underlying DNA sequences. Whether this difference between Drosophila and budding yeast reflects a mechanistic difference in the pairing mechanism is not clear at this point.
Studies with yeast have begun to explore the role of CEN-pairing in the segregation of exchange chromosomes. Zip1 persists at the centromeres of exchange chromosomes after SC disassembly and appears to mediate the pairing of homologous centromeres until prometaphase when chromosomes begin attaching to microtubules 54,55. Elimination of Zip1 or the spindle checkpoint (mad2) led to modest levels of segregation errors for exchange chromosomes, but removal of both elements led to random segregation 54,55. Although it should be noted that deletion of ZIP1 also disrupts wild-type levels of recombination and chiasma formation 58, this result suggested that CEN-pairing may help chiasmate partners attach in a bi-oriented configuration to the spindle. By this model, in the presence of a functional spindle checkpoint, CEN-pairing might not be important as repeated cycles of attachment and detachment ultimately result in a correct attachment in most cases; but when the spindle checkpoint is non-functional, CEN-pairing is critical to promote bi-orientation. Analogous experiments with achiasmate partners suggest that Zip1 and a prophase delay caused by Mad3 act in parallel pathways to enhance the segregation of non-exchange chromosomes 54,55.
In mouse and human, centromere associations in the form of clustering were first described by Scherthan and colleagues 44. Most of the associations described in this work appeared to be driven by attachment of the telomeres to the nuclear envelope and alignment of the chromosome arms. Subsequent studies revealed CEN-pairing in late prophase in mouse spermatocytes 59,60. In these reports, systematic evaluation of the SC disassembly process using fluorescence microscopy revealed that after SC proteins were removed from the chromosome arms (in diplotene) they remained at the centromeres (Fig. 1, mouse, arrow, and Fig. 4). Further, the chromosomal arms could freely separate after SC removal, but the centromeres – with their associated SC proteins – remained paired. In these studies using mouse spermatocytes CEN-pairing was shown to be occurring between chiasmate chromosomes. Interestingly, studies in mouse oocytes show no detectable SC components (SYCP3) at the centromeres following exit from pachytene 61 whereas SYCP3 does persist at centromeres through meiosis I and meiosis II in human oocytes, where it has been speculated to play a role in centromere orientation 62. While these studies reveal that CEN-pairing occurs in mammals in a manner with many parallels to what is seen in budding yeast, a number of questions persist. How widespread is this phenomenon? What are the implications for differences in CEN-pairing between spermatocytes and oocytes? Where it does exist is CEN-pairing promoting segregation of exchange chromosomes? Is CEN-pairing critical for the segregation of achiasmate partners in mammals as it is in yeast? Finally, how can SC components persist at the centromeres when it is disassembled from the arms?
Figure 4.

CEN-pairing in mouse spermatocytes. (A) Chromosome spreads of wild-type mouse spermatocytes in the diplotene stage of meiotic prophase (see Fig. 1) (image provided by R. Pezza). SYCP3 antibody was used to label axial elements. CREST antibody was used to label centromeres. SYCP1 antibody was used to demonstrate its presence at the paired centromeres and sites of chiasmata during this stage of meiosis. Blue arrowheads indicate examples of paired centromeres on chromosomes that have disassembled most synaptonemal complex proteins from the chromosome arms. (B) A cartoon of two homologous chromosomes engaged in centromere pairing (blue arrowhead). Light purple circles represent centromeres. Green lines represent the lateral elements. Persisting SC proteins are indicated in red.
What mediates CEN-pairing?
The precise structure that pairs centromeres remains to be elucidated, but genetic and cytological studies have shown that many of the same components involved in the formation of the SC are involved in CEN-pairing 54,55,59,60. SC components were initially implicated in mediating centromere associations by the finding, described above, that Zip1 is required for efficient centromere coupling in budding yeast 39. Subsequent experiments showed that CEN-pairing of achiasmate partners in late prophase also requires Zip1 54,55.
To consider the roles played by Zip1 and other SC proteins in CEN-pairing, it is useful to understand the basics of SC assembly (Fig. 3). The SC is a proteinaceous structure, common to most eukaryotes, that assembles during meiotic prophase and holds together the aligned homologs (reviewed in 63). The mature SC is a tripartite structure composed of two lateral elements and a unifying central region (Fig. 3) 64–69 (reviewed in 70). Although the major proteins of the SC from different organisms often share little sequence homology, they share structural homology and a common function: holding together homologous chromosome axes along their entire length. The exact events that trigger SC assembly are unknown. In many organisms SC assembly initiates near sites of recombination initiation, but SC assembly does not require recombination initiation events to trigger assembly and can often be observed in recombination mutants (reviewed in 71). In fact, in organisms such as Drosophila and Caenorhabditis elegans, SC formation precedes recombination 72–76. In budding yeast, SC formation may initiate from both sites of recombination initiation and paired centromeres 77. In these situations, alignment and pairing of certain chromosomal sites may promote SC assembly.
As cells exit pachytene the SC disassembles and the axes of the homologous partners are free to separate. It is at this stage that CEN-pairing can be clearly visualized. In both budding yeast and mouse spermatocytes, the homologous centromeres remain joined at this stage (Fig. 1, yeast, arrowhead, and mouse, arrow) 54,55,59,60. Thus in mouse chromosome spreads the axes appear tacked together at the centromeres and chiasmata (Fig. 1, mouse, arrow, and Fig. 4). In both budding yeast and mouse spermatocytes, the major transverse filament protein of the SC persists at the paired centromeres (Zip1 and SYCP1 respectively). More extended studies in mice have shown that every SC component tested (SYCP1, SYCP3, SYCE1 and TEX12; see Fig. 3) persists at the centromere 59,60 consistent with the notion that the structure holding the centromeres together, at least in mice, could be structurally similar to the rest of the SC.
In Drosophila females, the centromeres do not appear as discrete pairs in prophase; instead they group in one to a few clusters (Fig. 1, Drosophila, arrow) 78. Upon SC disassembly, as in budding yeast and mouse spermatocytes, the Zip1/SYCP1 homolog C(3)G and the central element protein Corona (Fig. 3) persist at the clustered centromeres (Fig. 1, Drosophila, Diplotene/Diakinesis) 79. Recent studies have shown that the clustering of the centromeres into one or two groups occurs in the cell divisions before meiotic entry and that the homologous centromeres pair prior to this clustering 80,81. Notably, the centromere clusters, but not the homologous centromere pairs, are highly diminished in SC mutants of Drosophila 76,79,80. CEN-pairing (and clustering) are dependent on the conventional cohesin subunit SMC1 and the Drosophila-specific cohesin components ORD, SOLO and SUNN 76,82–84. Whether these cohesin proteins participate directly in CEN-pairing or provide an environment in which pairing can occur remains to be determined.
How is it that in mice, human oocytes, yeast and Drosophila, SC components at the centromere (and at the chiasmata in mouse spermatocytes 60) are able to persist when the vast majority of SC proteins leave the chromosomes? Is the structure at the centromeres somehow fundamentally different, or is it protected from the SC removal mechanism? In both yeast and rodents there is evidence that phosphorylation of the SC by polo-like kinase homologs could trigger its disassembly 85–87. Furthermore, both SYCP1 and SYCP3 have potential PLK1 phosphorylation sites and can be phosphorylated in meiotic cell extracts 88.
Could protection of the centromeric SC from phosphorylation explain its persistence? Notably, centromeric cohesins are removed from chromosome arms in a process that includes phosphorylation, and in meiosis I centromeric cohesins are protected from removal by the protein Shugoshin and its associated phosphatases 89–92 (see accompanying minireview by Rankin 3). Could this same or a similar mechanism protect centromeric SC components from the phosphorylation that triggers SC disassembly? Future experiments that clarify the structure, components and disassembly regulation of the CEN-pairing apparatus will help to address these questions.
What drives CEN–CEN partner choice?
In budding yeast, centromere plasmids, artificial chromosomes and homeologous chromosomes appear to pair with one another in late prophase regardless of their degree of homology at the centromere. Experiments with these model chromosomes suggest they pair by an ‘exclusion mechanism’ where the last two unpaired chromosomes pair after the chiasmate chromosomes have identified and paired with their partners 35. This CEN-pairing has been suggested to be a continuation of the homology-independent CEN-coupling that occurs in early prophase 39,48. By this model, centromeres couple, uncouple and couple with another partner until recombination-based homology search processes identify the correct partners 48. This would leave the last two centromeres to pair by default, regardless of their homology 54,55,93.
Although budding yeast chromosomes may be dependent on an ‘exclusion mechanism’, a different mechanism is at work in female Drosophila. Once termed ‘distributive segregation’, the homologous achiasmate system in Drosophila oocytes was once hypothesized to function under a similar mechanism in which homologs or non-homologs would pair by default if they were not engaged with a homologous partner forming a chiasma 94. But now it is understood that X and fourth chromosome partners in Drosophila oocytes pair by a mechanism that depends on homologous regions of peri-centric heterochromatin. Such regions are notoriously difficult to analyze at the DNA sequence level. Despite this, a study of one peri-centromeric region, sufficient for mediating CEN-pairing, found that it contained many fragments of multiple transposable element families in addition to repeats of simple satellite sequences 95. How CEN-pairing choice is biased by this constellation of repetitious sequences, copies of some of which are scattered on more than one chromosome, is a mystery. An interesting possibility is that it is a pattern of epigenetic marks 96 rather than the DNA sequence per se that confers pairing choice.
Unlike budding yeast, fission yeast has complex centromere regions that bear multiple repetitive elements. In the absence of recombination, the chromosomes’ arms no longer align in meiosis, but the centromeres are still able to pair with their homologous partners 42. The achiasmate partners are able to segregate properly but with low fidelity 97. Here, too, the mechanism by which CEN-pairing is mediated is not known. Studies in a number of organisms including fission yeast 98–100 have implicated centromeric-encoded RNAs, or transcription, in centromere functions such as heterochromatin formation and kinetochore protein recruitment (reviewed in 101). Recent work revealed that RNA transcripts play a key role at a specific chromosome arm pairing site in fission yeast 100, raising the possibility that this might be a general pairing mechanism 102 that could also work at centromeres.
How could CEN-pairing promote disjunction?
How could CEN-pairing help partner chromosomes to become properly oriented on the meiotic spindle? Studies in yeast and mice suggest that initial chromosome attachments to the spindle are usually incorrect, often biased towards one pole, and usually undergo two or more bi-orientation attempts before stabilizing their attachment to the spindle 103,104. Three models have been suggested to explain how CEN-pairing might optimize this process: the Janus model 46,54,105, the connector model, and the elastic thread model 106.
The Janus model (Fig. 5A) 54,105, named after the two-faced god of beginnings and transitions from Roman mythology, suggests that CEN-pairing locks the kinetochores in a back-to-back configuration with their microtubule attachment ‘faces’ looking in opposite directions. This model assumes that, in the absence of CEN-pairing, the kinetochores would have rotational freedom and their orientations relative to the spindle poles would be uncoordinated (Fig. 5A). Chromosome pairs often begin the bi-orientation process by moving towards one pole 103,104, presumably due to an initial attachment to microtubules to that pole. In mice, the pole that chromosomes are pulled toward is random, whereas in yeast the pole is usually the ‘mother’ spindle pole body. This pulling towards one pole would serve to rotate the chromosome so that the opposite kinetochore face would be oriented towards the opposite pole. Observations of centromere pairs in many organisms show that, once they are actively moving on the spindle during the bi-orientation process, the centromeres are well separated and not back-to-back. This is true in both mouse spermatocytes 59,60 and yeast (E.L.K. unpublished and 104). Thus, this mechanism would seem to mainly optimize the chance that the initial microtubule attachments are correct.
Figure 5.
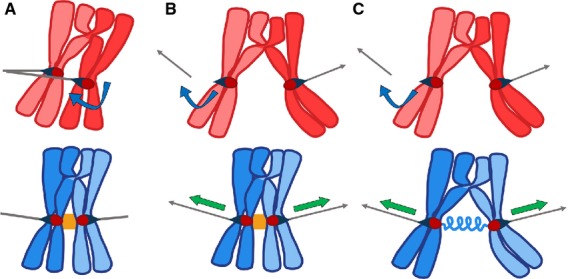
Three models for CEN-pairing. (A) The Janus model. By this model, in the absence of CEN-pairing (red chromosomes), kinetochores have rotational freedom (blue arrow). This would allow both the kinetochores to attach to the same pole. CEN-pairing (orange block between centromeres of blue chromosomes) would lock the kinetochores together such that their microtubule attachment ‘faces’ are oriented away from one another, optimizing the chance that, if one kinetochore attaches to one pole, its partner kinetochore will be facing towards microtubules emanating from the opposite pole. (B) The centromere connector model. By this model, in the absence of CEN-pairing (red chromosomes), the chromosomes with a distal crossover would not be able to generate tension at the kinetochore–microtubule interface necessary to stabilize kinetochore–microtubule connections. CEN-pairing (orange block between centromeres of blue chromosomes) would hold the centromeres together, allowing opposing poleward forces to generate tension (green arrows) when a bipolar attachment is made. (C) The elastic thread model. In this model, similar to the centromere connector model, CEN-pairing promotes bi-orientation by providing a means for bi-oriented centromeres to generate tension (green arrows) when opposing kinetochore–microtubule connections are formed. Here, CEN-pairing does not hold centromeres together at the time of bi-orientation, but instead provides an environment in which chromatin from the two chromosomes becomes connected (symbolized by the blue spring between centromeres). By this model, direct CEN-pairing can be lost prior to the microtubule attachment process. Instead, the partner centromeres remain joined by a chromatin bridge – even in the absence of a crossover.
The connector model (Fig. 5B) suggests that CEN-pairing acts as a connection between the centromeres over which tension can be transmitted (like chiasmata do at chromosome arms). Kinetochore–microtubule attachments that are under tension – because they are pulling against a kinetochore that is attached to a microtubule from the opposite pole – are more stable than attachments without tension (Fig. 2; reviewed in 9,107). In meiosis, that tension must be provided across a chiasma or another connection. By this model, CEN pairing would provide that bridge. As in the previous model, the fact that homologous centromeres are often well separated in the bi-orientation process suggests that this mechanism could only work early on, prior to disengagement of the paired centromeres.
The elastic thread model has been suggested by recent work in female Drosophila which indicates that achiasmate (and perhaps chiasmate) partners are connected during the bi-orientation process by some sort of chromatin thread (Fig. 1, Drosophila, arrowhead, and Fig. 5C) 106. By this model, CEN-pairing does not act directly to mediate bi-orientation in prometaphase. Instead, CEN-pairing in prophase acts to provide an environment in which chromatin connections can be established between the partner centromeres. It is this chromatin bridge, not the direct CEN-pairing, which then promotes bi-orientation. An elastic bridge could promote bi-orientation by transmitting tension signals or by keeping the pair in physical proximity.
In Drosophila females the achiasmate chromosome partners do not immediately become bi-oriented when the centromeres separate. Instead the chromosomes oscillate on the spindle before becoming bi-oriented 106. A significant portion of the separated partners could be shown to be connected by nearly imperceptible chromatin threads (technical limitations made it impossible to determine whether every pair is joined by a thread). The notion of connections between separated meiotic partners is not a new one (see 108 and references within), and in fact these mysterious connections have been shown to have elastic properties that pull the partners towards one another. A significant development is that the most recent report 106 identifies the threads as containing peri-centromeric DNA sequences. These meiotic threads of peri-centric heterochromatin should not be confused with the ultrafine anaphase bridges (UFBs) observed in mitotic cell lines 109–112. The mechanism of UFB formation, though still unclear, has been hypothesized to be from collapsed replication forks or DNA repair mechanisms and the UFBs are marked by proteins implicated in these processes 109–112. In meiotic cells, the connections are between homologous partners and not sister chromatids 106. Some of the unanswered questions regarding these threads surround their formation and components. How are the threads formed? Meiotic centromeres are rich in topoisomerase II and cohesin proteins (see review by Rankin 3; 113–115). Could topoisomerase, cohesins or some form of non-crossover recombination or DNA repair mechanism mediate the formation of inter-homolog connections? What mechanisms are required to dissolve the connections without causing unintended DNA damage? A recent study of Drosophila oocytes has shown that in the absence of topoisomerase II heterochromatin threads persist between separating homologous partners in meiosis I, blocking their segregation, suggesting the model that the threads between homologous centromeres are due, at least in part, to catenation 106.
The behavior of Zip1/SYCP1 at centromeres is consistent with a role in providing an environment hospitable to the formation of CEN–CEN connections, while not directly acting in the bi-orientation process. In budding yeast Zip1 is necessary for CEN-pairing between achiasmate partners and their subsequent disjunction. But the majority of Zip1 has left the paired centromeres before the bi-orientation process begins 54,55. Similarly, in mouse spermatocytes, SYCP1 is present at paired centromeres in early diplotene and is necessary for CEN-pairing, but it is not detectable at paired centromeres in late diplotene 59,60. Further, once the mouse centromeres begin to separate, they seem to be joined by bridges containing the axis component SYCP3 60. SYCP3 also persists at the centromeres into meiosis I in human oocytes 62, but studies that might detect SYCP3 between the centromeres of separating homologous partners have yet to be reported.
Conclusion
There is growing evidence that CEN–CEN interactions are not only a conserved phenomenon but serve an important role in promoting the fidelity of meiotic chromosome segregation. Drosophila, yeast and mouse have proved to be valuable model organisms to study the way in which CEN-pairing occurs and its impacts on the behavior of chromosomes. This is especially true for achiasmate partners. In humans, the mis-segregation of chromosomes that failed to establish chiasmata is a major cause of spontaneous abortion and aneuploidy-based syndromes 4. The smallest chromosomes (21 and 22) fail to experience crossovers in about 5% of meioses 116–118 yet are estimated to non-disjoin in fewer than 1% of meioses 117–119, suggesting that in there may be factors beyond chiasmata that promote the segregation of homologous partners in human cells. Future studies of CEN-pairing will probably reveal new general principles for meiotic chromosome behavior and may well have implications for human health.
Acknowledgments
We thank Roberto Pezza for comments on the manuscript. DD is supported by grant R01GM087377 from the National Institutes of General Medical Sciences.
Glossary
- CEN–CEN
centromere–centromere
- SC
synaptonemal complex
- UFB
ultrafine anaphase bridges
Author contributions
Emily Kurdzo prepared the first draft of this review. Emily Kurdzo and Dean Dawson worked together to edit, and revise the document to produce the final manuscript.
References
- Sansam CL. Pezza RJ. Connecting by breaking and repairing: mechanisms of DNA strand exchange in meiotic recombination. FEBS J. 2015;282:2431–2444. doi: 10.1111/febs.13317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohl KP. Sekelsky J. Meiotic and mitotic recombination in meiosis. Genetics. 2013;194:327–334. doi: 10.1534/genetics.113.150581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rankin S. Complex elaboration: making sense of meiotic cohesin dynamics. FEBS J. 2015;282:2413–2430. doi: 10.1111/febs.13301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassold T. Hunt P. To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet. 2001;2:280–291. doi: 10.1038/35066065. [DOI] [PubMed] [Google Scholar]
- Sakuno T, Tanaka K, Hauf S. Watanabe Y. Repositioning of aurora B promoted by chiasmata ensures sister chromatid mono-orientation in meiosis I. Dev Cell. 2011;21:534–545. doi: 10.1016/j.devcel.2011.08.012. [DOI] [PubMed] [Google Scholar]
- Kouznetsova A, Lister L, Nordenskjold M, Herbert M. Hoog C. Bi-orientation of achiasmatic chromosomes in meiosis I oocytes contributes to aneuploidy in mice. Nat Genet. 2007;39:966–968. doi: 10.1038/ng2065. [DOI] [PubMed] [Google Scholar]
- LeMaire-Adkins R. Hunt PA. Nonrandom segregation of the mouse univalent X chromosome: evidence of spindle-mediated meiotic drive. Genetics. 2000;156:775–783. doi: 10.1093/genetics/156.2.775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorbsky G. The spindle checkpoint and chromosome segregation in meiosis. FEBS J. 2015 doi: 10.1111/febs.13166. , doi: 10.1111/febs.13166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicklas RB. Koch CA. Chromosome micromanipulation. 3. Spindle fiber tension and the reorientation of mal-oriented chromosomes. J Cell Biol. 1969;43:40–50. doi: 10.1083/jcb.43.1.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keeney S, Giroux CN. Kleckner N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell. 1997;88:375–384. doi: 10.1016/s0092-8674(00)81876-0. [DOI] [PubMed] [Google Scholar]
- Celerin M, Merino ST, Stone JE, Menzie AM. Zolan ME. Multiple roles of Spo11 in meiotic chromosome behavior. EMBO J. 2000;19:2739–2750. doi: 10.1093/emboj/19.11.2739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowring FJ, Yeadon PJ, Stainer RG. Catcheside DE. Chromosome pairing and meiotic recombination in Neurospora crassa spo11 mutants. Curr Genet. 2006;50:115–123. doi: 10.1007/s00294-006-0066-1. [DOI] [PubMed] [Google Scholar]
- Storlazzi A, Tesse S, Gargano S, James F, Kleckner N. Zickler D. Meiotic double-strand breaks at the interface of chromosome movement, chromosome remodeling, and reductional division. Genes Dev. 2003;17:2675–2687. doi: 10.1101/gad.275203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dernburg AF, McDonald K, Moulder G, Barstead R, Dresser M. Villeneuve AM. Meiotic recombination in C. elegans initiates by a conserved mechanism and is dispensable for homologous chromosome synapsis. Cell. 1998;94:387–398. doi: 10.1016/s0092-8674(00)81481-6. [DOI] [PubMed] [Google Scholar]
- McKim KS. Hayashi-Hagihara A. mei-W68 in Drosophila melanogaster encodes a Spo11 homolog: evidence that the mechanism for initiating meiotic recombination is conserved. Genes Dev. 1998;12:2932–2942. doi: 10.1101/gad.12.18.2932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baudat F, Manova K, Yuen JP, Jasin M. Keeney S. Chromosome synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol Cell. 2000;6:989–998. doi: 10.1016/s1097-2765(00)00098-8. [DOI] [PubMed] [Google Scholar]
- Romanienko PJ. Camerini-Otero RD. The mouse Spo11 gene is required for meiotic chromosome synapsis. Mol Cell. 2000;6:975–987. doi: 10.1016/s1097-2765(00)00097-6. [DOI] [PubMed] [Google Scholar]
- Steiner WW, Schreckhise RW. Smith GR. Meiotic DNA breaks at the S. pombe recombination hot spot M26. Mol Cell. 2002;9:847–855. doi: 10.1016/s1097-2765(02)00489-6. [DOI] [PubMed] [Google Scholar]
- Grelon M, Vezon D, Gendrot G. Pelletier G. AtSPO11-1 is necessary for efficient meiotic recombination in plants. EMBO J. 2001;20:589–600. doi: 10.1093/emboj/20.3.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stacey NJ, Kuromori T, Azumi Y, Roberts G, Breuer C, Wada T, Maxwell A, Roberts K. Sugimoto-Shirasu K. Arabidopsis SPO11-2 functions with SPO11-1 in meiotic recombination. Plant J. 2006;48:206–216. doi: 10.1111/j.1365-313X.2006.02867.x. [DOI] [PubMed] [Google Scholar]
- Rasmussen SW. Meiosis in Bombyx mori females. Philos Trans R Soc Lond B Biol Sci. 1977;277:343–350. doi: 10.1098/rstb.1977.0022. [DOI] [PubMed] [Google Scholar]
- Wolf KW. How meiotic cells deal with non-exchange chromosomes. BioEssays. 1994;16:107–114. doi: 10.1002/bies.950160207. [DOI] [PubMed] [Google Scholar]
- Vazquez J, Belmont AS. Sedat JW. The dynamics of homologous chromosome pairing during male Drosophila meiosis. Curr Biol. 2002;12:1473–1483. doi: 10.1016/s0960-9822(02)01090-4. [DOI] [PubMed] [Google Scholar]
- Sturtevant AH. Beadle GW. The Relations of Inversions in the X Chromosome of Drosophila Melanogaster to crossing over and disjunction. Genetics. 1936;21:554–604. doi: 10.1093/genetics/21.5.554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper KW. Normal segregation without Chiasmata in female Drosophila Melanogaster. Genetics. 1945;30:472–484. doi: 10.1093/genetics/30.5.472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carpenter AT. A meiotic mutant defective in distributive disjunction in Drosophila melanogaster. Genetics. 1973;73:393–428. doi: 10.1093/genetics/73.3.393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grell RF. Distributive pairing: the size-dependent mechanism for regular segregation of the fourth chromosomes in Drosophila Melanogaster. Proc Natl Acad Sci U S A. 1964;52:226–232. doi: 10.1073/pnas.52.2.226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann C. Davis RW. Meiotic disjunction of circular minichromosomes in yeast does not require DNA homology. PNAS. 1986;83:6017–6019. doi: 10.1073/pnas.83.16.6017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson DS, Murray AW. Szostak JW. An alternative pathway for meiotic chromosome segregation in yeast. Science. 1986;234:713–717. doi: 10.1126/science.3535068. [DOI] [PubMed] [Google Scholar]
- Guacci V. Kaback DB. Distributive disjunction of authentic chromosomes in Saccharomyces cerevisiae. Genetics. 1991;127:475–488. doi: 10.1093/genetics/127.3.475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Fuente R, Parra MT, Viera A, Calvente A, Gomez R, Suja JA, Rufas JS. Page J. Meiotic pairing and segregation of achiasmate sex chromosomes in eutherian mammals: the role of SYCP3 protein. PLoS Genet. 2007;3:e198. doi: 10.1371/journal.pgen.0030198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page J, Viera A, Parra MT, de la Fuente R, Suja JA, Prieto I, Barbero JL, Rufas JS, Berrios S. Fernandez-Donoso R. Involvement of synaptonemal complex proteins in sex chromosome segregation during marsupial male meiosis. PLoS Genet. 2006;2:e136. doi: 10.1371/journal.pgen.0020136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas SE, Soltani-Bejnood M, Roth P, Dorn R, Logsdon JM., Jr McKee BD. Identification of two proteins required for conjunction and regular segregation of achiasmate homologs in Drosophila male meiosis. Cell. 2005;123:555–568. doi: 10.1016/j.cell.2005.08.043. [DOI] [PubMed] [Google Scholar]
- Dernburg AF, Sedat JW. Hawley RS. Direct evidence of a role for heterochromatin in meiotic chromosome segregation. Cell. 1996;86:135–146. doi: 10.1016/s0092-8674(00)80084-7. [DOI] [PubMed] [Google Scholar]
- Kemp B, Boumil RM, Stewart MN. Dawson DS. A role for centromere pairing in meiotic chromosome segregation. Genes Dev. 2004;18:1946–1951. doi: 10.1101/gad.1227304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Church K. Moens PB. Centromere behavior during interphase and meiotic prophase in Allium fistulosum from 3-D, E.M. reconstruction. Chromosoma. 1976;56:249–263. [Google Scholar]
- Bennett M. Centromere arrangements in Triticum aestivum and their relationship to synapsis. Heredity. 1979;43:157. [Google Scholar]
- Martínez-Pérez E, Shaw P, Reader S, Aragón-Alcaide L, Miller T. Moore G. Homologous chromosome pairing in wheat. J Cell Sci. 1999;112:1761–1769. doi: 10.1242/jcs.112.11.1761. [DOI] [PubMed] [Google Scholar]
- Tsubouchi T. Roeder GS. A synaptonemal complex protein promotes homology-independent centromere coupling. Science. 2005;308:870–873. doi: 10.1126/science.1108283. [DOI] [PubMed] [Google Scholar]
- Obeso D. Dawson DS. Temporal characterization of homology-independent centromere coupling in meiotic prophase. PLoS One. 2010;5:e10336. doi: 10.1371/journal.pone.0010336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prieto P, Santos AP, Moore G. Shaw P. Chromosomes associate premeiotically and in xylem vessel cells via their telomeres and centromeres in diploid rice (Oryza sativa) Chromosoma. 2004;112:300–307. doi: 10.1007/s00412-004-0274-8. [DOI] [PubMed] [Google Scholar]
- Ding D-Q, Yamamoto A, Haraguchi T. Hiraoka Y. Dynamics of homologous chromosome pairing during meiotic prophase in fission yeast. Dev Cell. 2004;6:329–341. doi: 10.1016/s1534-5807(04)00059-0. [DOI] [PubMed] [Google Scholar]
- Zhang J, Pawlowski WP. Han F. Centromere pairing in early meiotic prophase requires active centromeres and precedes installation of the synaptonemal complex in maize. Plant Cell. 2013;25:3900–3909. doi: 10.1105/tpc.113.117846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherthan H, Weich S, Schwegler H, Heyting C, Harle M. Cremer T. Centromere and telomere movements during early meiotic prophase of mouse and man are associated with the onset of chromosome pairing. J Cell Biol. 1996;134:1109–1125. doi: 10.1083/jcb.134.5.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takada Y, Naruse C, Costa Y, Shirakawa T, Tachibana M, Sharif J, Kezuka-Shiotani F, Kakiuchi D, Masumoto H, Shinkai Y, et al. HP1gamma links histone methylation marks to meiotic synapsis in mice. Development. 2011;138:4207–4217. doi: 10.1242/dev.064444. [DOI] [PubMed] [Google Scholar]
- Obeso D, Pezza RJ. Dawson D. Couples, pairs, and clusters: mechanisms and implications of centromere associations in meiosis. Chromosoma. 2014;123:43–55. doi: 10.1007/s00412-013-0439-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klutstein M. Cooper JP. The chromosomal courtship dance-homolog pairing in early meiosis. Curr Opin Cell Biol. 2014;26:123–131. doi: 10.1016/j.ceb.2013.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk JE, Chan AC, Hoffmann E. Hochwagen A. A Mec1- and PP4-dependent checkpoint couples centromere pairing to meiotic recombination. Dev Cell. 2010;19:599–611. doi: 10.1016/j.devcel.2010.09.006. [DOI] [PubMed] [Google Scholar]
- Hawley RS, Irick H, Zitron AE, Haddox DA, Lohe A, New C, Whitley MD, Arbel T, Jang J, Mckim K, et al. There are two mechanisms of Achiasmate segregation in Drosophila females, one of which requires heterochromatic homology. Dev Gen. 1993;13:440–467. doi: 10.1002/dvg.1020130608. [DOI] [PubMed] [Google Scholar]
- Karpen GH, Le M-H. Le H. Centric heterochromatin and the efficiency of Achiasmate disjunction in Drosophila female meiosis. Science. 1996;273:118–122. doi: 10.1126/science.273.5271.118. [DOI] [PubMed] [Google Scholar]
- Sun X, Wahlstrom J. Karpen G. Molecular structure of a functional Drosophila centromere. Cell. 1997;91:1007–1019. doi: 10.1016/s0092-8674(00)80491-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shubochkina EA, Nielsen TL. Nilsson-Tillgren T. Meiotic crossing-over in the regions of homology between homologous chromosomes V. Yeast. 2001;18:1173–1183. doi: 10.1002/yea.759. [DOI] [PubMed] [Google Scholar]
- Maxfield Boumil R, Kemp B, Angelichio M, Nilsson-Tillgren T. Dawson DS. Meiotic segregation of a homeologous chromosome pair. Mol Genet Genomics. 2003;268:750–760. doi: 10.1007/s00438-002-0796-9. [DOI] [PubMed] [Google Scholar]
- Gladstone MN, Obeso D, Chuong H. Dawson DS. The synaptonemal complex protein Zip1 promotes bi-orientation of centromeres at meiosis I. PLoS Genet. 2009;5:e1000771. doi: 10.1371/journal.pgen.1000771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newnham L, Jordan P, Rockmill B, Roeder GS. Hoffmann E. The synaptonemal complex protein, Zip1, promotes the segregation of nonexchange chromosomes at meiosis I. Proc Natl Acad Sci U S A. 2010;107:781–785. doi: 10.1073/pnas.0913435107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers SR, Hunter N, Louis EJ. Borts RH. The mismatch repair system reduces meiotic homeologous recombination and stimulates recombination-dependent chromosome loss. Mol Cell Biol. 1996;16:6110–6120. doi: 10.1128/mcb.16.11.6110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilsson-Tillgren T, Gjermansen C, Holmberg S, Litske Petersen J. Kielland-Brandt M. Analysis of chromosome V and theILV1 gene from Saccharomyces carlsbergensis. Carlsberg Res Commun. 1986;51:309–326. [Google Scholar]
- Storlazzi A, Xu L, Schwacha A. Kleckner N. Synaptonemal complex (SC) component Zipl plays a role in meiotic recombination independent of SC polymerization along the chromosomes. PNAS. 1996;93:9043–9048. doi: 10.1073/pnas.93.17.9043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisig CG, Guiraldelli MF, Kouznetsova A, Scherthan H, Hoog C, Dawson DS. Pezza RJ. Synaptonemal complex components persist at centromeres and are required for homologous centromere pairing in mouse spermatocytes. PLoS Genet. 2012;8:e1002701. doi: 10.1371/journal.pgen.1002701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiao H, Chen JK, Reynolds A, Hoog C, Paddy M. Hunter N. Interplay between synaptonemal complex, homologous recombination, and centromeres during mammalian meiosis. PLoS Genet. 2012;8:e1002790. doi: 10.1371/journal.pgen.1002790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodges CA, LeMaire-Adkins R. Hunt PA. Coordinating the segregation of sister chromatids during the first meiotic division: evidence for sexual dimorphism. J Cell Sci. 2001;114:2417–2426. doi: 10.1242/jcs.114.13.2417. [DOI] [PubMed] [Google Scholar]
- Garcia-Cruz R, Brieno MA, Roig I, Grossmann M, Velilla E, Pujol A, Cabero L, Pessarrodona A, Barbero JL. Garcia Caldes M. Dynamics of cohesin proteins REC8, STAG3, SMC1 beta and SMC3 are consistent with a role in sister chromatid cohesion during meiosis in human oocytes. Hum Reprod. 2010;25:2316–2327. doi: 10.1093/humrep/deq180. [DOI] [PubMed] [Google Scholar]
- Page SL. Hawley RS. The genetics and molecular biology of the synaptonemal complex. Annu Rev Cell Dev Biol. 2004;20:525–558. doi: 10.1146/annurev.cellbio.19.111301.155141. [DOI] [PubMed] [Google Scholar]
- Sym M, Engebrecht JA. Roeder GS. ZIP1 is a synaptonemal complex protein required for meiotic chromosome synapsis. Cell. 1993;72:365–378. doi: 10.1016/0092-8674(93)90114-6. [DOI] [PubMed] [Google Scholar]
- Page SL. Hawley RS. c(3)G encodes a Drosophila synaptonemal complex protein. Genes Dev. 2001;15:3130–3143. doi: 10.1101/gad.935001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meuwissen RL, Offenberg HH, Dietrich AJ, Riesewijk A, van Iersel M. Heyting C. A coiled-coil related protein specific for synapsed regions of meiotic prophase chromosomes. EMBO J. 1992;11:5091–5100. doi: 10.1002/j.1460-2075.1992.tb05616.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page SL, Khetani RS, Lake CM, Nielsen RJ, Jeffress JK, Warren WD, Bickel SE. Hawley RS. Corona is required for higher-order assembly of transverse filaments into full-length synaptonemal complex in Drosophila oocytes. PLoS Genet. 2008;4:e1000194. doi: 10.1371/journal.pgen.1000194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphryes N, Leung WK, Argunhan B, Terentyev Y, Dvorackova M. Tsubouchi H. The Ecm11-Gmc2 complex promotes synaptonemal complex formation through assembly of transverse filaments in budding yeast. PLoS Genet. 2013;9:e1003194. doi: 10.1371/journal.pgen.1003194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voelkel-Meiman K, Taylor LF, Mukherjee P, Humphryes N, Tsubouchi H. Macqueen AJ. SUMO localizes to the central element of synaptonemal complex and is required for the full synapsis of meiotic chromosomes in budding yeast. PLoS Genet. 2013;9:e1003837. doi: 10.1371/journal.pgen.1003837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawley RS. Solving a meiotic LEGO puzzle: transverse filaments and the assembly of the synaptonemal complex in Caenorhabditis elegans. Genetics. 2011;189:405–409. doi: 10.1534/genetics.111.134197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henderson KA. Keeney S. Synaptonemal complex formation: where does it start? BioEssays. 2005;27:995–998. doi: 10.1002/bies.20310. [DOI] [PubMed] [Google Scholar]
- Liu H, Jang JK, Kato N. McKim KS. mei-P22 encodes a chromosome-associated protein required for the initiation of meiotic recombination in Drosophila melanogaster. Genetics. 2002;162:245–258. doi: 10.1093/genetics/162.1.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang JK, Sherizen DE, Bhagat R, Manheim EA. McKim KS. Relationship of DNA double-strand breaks to synapsis in Drosophila. J Cell Sci. 2003;116:3069–3077. doi: 10.1242/jcs.00614. [DOI] [PubMed] [Google Scholar]
- McKim KS, Jang JK. Manheim EA. Meiotic recombination and chromosome segregation in Drosophila females. Annu Rev Genet. 2002;36:205–232. doi: 10.1146/annurev.genet.36.041102.113929. [DOI] [PubMed] [Google Scholar]
- MacQueen AJ, Colaiacovo MP, McDonald K. Villeneuve AM. Synapsis-dependent and -independent mechanisms stabilize homolog pairing during meiotic prophase in C. elegans. Genes Dev. 2002;16:2428–2442. doi: 10.1101/gad.1011602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanneti NS, Landy K, Joyce EF. McKim KS. A pathway for synapsis initiation during zygotene in Drosophila oocytes. Curr Biol. 2011;21:1852–1857. doi: 10.1016/j.cub.2011.10.005. [DOI] [PubMed] [Google Scholar]
- Tsubouchi T, Macqueen AJ. Roeder GS. Initiation of meiotic chromosome synapsis at centromeres in budding yeast. Genes Dev. 2008;22:3217–3226. doi: 10.1101/gad.1709408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khetani RS. Bickel SE. Regulation of meiotic cohesion and chromosome core morphogenesis during pachytene in Drosophila oocytes. J Cell Sci. 2007;120:3123–3137. doi: 10.1242/jcs.009977. [DOI] [PubMed] [Google Scholar]
- Takeo S, Lake CM, Morais-de-Sa E, Sunkel CE. Hawley RS. Synaptonemal complex-dependent centromeric clustering and the initiation of synapsis in Drosophila oocytes. Curr Biol. 2011;21:1845–1851. doi: 10.1016/j.cub.2011.09.044. [DOI] [PubMed] [Google Scholar]
- Christophorou N, Rubin T. Huynh JR. Synaptonemal complex components promote centromere pairing in pre-meiotic germ cells. PLoS Genet. 2013;9:e1004012. doi: 10.1371/journal.pgen.1004012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joyce EF, Apostolopoulos N, Beliveau BJ. Wu CT. Germline progenitors escape the widespread phenomenon of homolog pairing during Drosophila development. PLoS Genet. 2013;9:e1004013. doi: 10.1371/journal.pgen.1004013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian VV. Bickel SE. Aging predisposes oocytes to meiotic nondisjunction when the cohesin subunit SMC1 is reduced. PLoS Genet. 2008;4:e1000263. doi: 10.1371/journal.pgen.1000263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan R. McKee BD. The cohesion protein SOLO associates with SMC1 and is required for synapsis, recombination, homolog bias and cohesion and pairing of centromeres in Drosophila Meiosis. PLoS Genet. 2013;9:e1003637. doi: 10.1371/journal.pgen.1003637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnan B, Thomas SE, Yan R, Yamada H, Zhulin IB. McKee BD. Sisters unbound is required for meiotic centromeric cohesion in Drosophila melanogaster. Genetics. 2014;198:947–965. doi: 10.1534/genetics.114.166009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan PW, Karppinen J. Handel MA. Polo-like kinase is required for synaptonemal complex disassembly and phosphorylation in mouse spermatocytes. J Cell Sci. 2012;125:5061–5072. doi: 10.1242/jcs.105015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sourirajan A. Lichten M. Polo-like kinase Cdc5 drives exit from pachytene during budding yeast meiosis. Genes Dev. 2008;22:2627–2632. doi: 10.1101/gad.1711408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun F. Handel MA. Regulation of the meiotic prophase I to metaphase I transition in mouse spermatocytes. Chromosoma. 2008;117:471–485. doi: 10.1007/s00412-008-0167-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarsounas M, Pearlman RE. Moens PB. Meiotic activation of rat pachytene spermatocytes with okadaic acid: the behaviour of synaptonemal complex components SYN1/SCP1 and COR1/SCP3. J Cell Sci. 1999;112:423–434. doi: 10.1242/jcs.112.4.423. [DOI] [PubMed] [Google Scholar]
- Kitajima TS, Kawashima SA. Watanabe Y. The conserved kinetochore protein shugoshin protects centromeric cohesion during meiosis. Nature. 2004;427:510–517. doi: 10.1038/nature02312. [DOI] [PubMed] [Google Scholar]
- Kitajima TS, Sakuno T, Ishiguro K, Iemura S, Natsume T, Kawashima SA. Watanabe Y. Shugoshin collaborates with protein phosphatase 2A to protect cohesin. Nature. 2006;441:46–52. doi: 10.1038/nature04663. [DOI] [PubMed] [Google Scholar]
- Riedel CG, Katis VL, Katou Y, Mori S, Itoh T, Helmhart W, Galova M, Petronczki M, Gregan J, Cetin B, et al. Protein phosphatase 2A protects centromeric sister chromatid cohesion during meiosis I. Nature. 2006;441:53–61. doi: 10.1038/nature04664. [DOI] [PubMed] [Google Scholar]
- Llano E, Gomez R, Gutierrez-Caballero C, Herran Y, Sanchez-Martin M, Vazquez-Quinones L, Hernandez T, de Alava E, Cuadrado A, Barbero JL, et al. Shugoshin-2 is essential for the completion of meiosis but not for mitotic cell division in mice. Genes Dev. 2008;22:2400–2413. doi: 10.1101/gad.475308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loidl J, Scherthant H. Kaback DB. Physical association between nonhomologous chromosomes precedes distributive disjunction in yeast. PNAS. 1994;91:331–334. doi: 10.1073/pnas.91.1.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grell RF. A new model for secondary nondisjunction: the role of distributive pairing. Genetics. 1962;47:1737–1754. doi: 10.1093/genetics/47.12.1737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun X, Le HD, Wahlstrom JM. Karpen GH. Sequence analysis of a functional Drosophila centromere. Genome Res. 2003;13:182–194. doi: 10.1101/gr.681703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guenatri M, Bailly D, Maison C. Almouzni G. Mouse centric and pericentric satellite repeats form distinct functional heterochromatin. J Cell Biol. 2004;166:493–505. doi: 10.1083/jcb.200403109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis L. Smith GR. Dynein promotes achiasmate segregation in Schizosaccharomyces pombe. Genetics. 2005;170:581–590. doi: 10.1534/genetics.104.040253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lejeune E, Bayne EH. Allshire RC. On the connection between RNAi and heterochromatin at centromeres. Cold Spring Harb Symp Quant Biol. 2010;75:275–283. doi: 10.1101/sqb.2010.75.024. [DOI] [PubMed] [Google Scholar]
- Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI. Martienssen RA. Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science. 2002;297:1833–1837. doi: 10.1126/science.1074973. [DOI] [PubMed] [Google Scholar]
- Ding DQ, Okamasa K, Yamane M, Tsutsumi C, Haraguchi T, Yamamoto M. Hiraoka Y. Meiosis-specific noncoding RNA mediates robust pairing of homologous chromosomes in meiosis. Science. 2012;336:732–736. doi: 10.1126/science.1219518. [DOI] [PubMed] [Google Scholar]
- Gent JI. Dawe RK. RNA as a structural and regulatory component of the centromere. Annu Rev Genet. 2012;46:443–453. doi: 10.1146/annurev-genet-110711-155419. [DOI] [PubMed] [Google Scholar]
- Ding DQ, Haraguchi T. Hiraoka Y. The role of chromosomal retention of noncoding RNA in meiosis. Chromosome Res. 2013;21:665–672. doi: 10.1007/s10577-013-9389-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitajima TS, Ohsugi M. Ellenberg J. Complete kinetochore tracking reveals error-prone homologous chromosome biorientation in mammalian oocytes. Cell. 2011;146:568–581. doi: 10.1016/j.cell.2011.07.031. [DOI] [PubMed] [Google Scholar]
- Meyer RE, Kim S, Obeso D, Straight PD, Winey M. Dawson DS. Mps1 and Ipl1/Aurora B act sequentially to correctly orient chromosomes on the meiotic spindle of budding yeast. Science. 2013;339:1071–1074. doi: 10.1126/science.1232518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stewart MN. Dawson DS. Changing partners: moving from non-homologous to homologous centromere pairing in meiosis. Trends Genet. 2008;24:564–573. doi: 10.1016/j.tig.2008.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes SE, Gilliland WD, Cotitta JL, Takeo S, Collins KA. Hawley RS. Heterochromatic threads connect oscillating chromosomes during prometaphase I in Drosophila oocytes. PLoS Genet. 2009;5:e1000348. doi: 10.1371/journal.pgen.1000348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe Y. Geometry and force behind kinetochore orientation: lessons from meiosis. Nat Rev Mol Cell Biol. 2012;13:370–382. doi: 10.1038/nrm3349. [DOI] [PubMed] [Google Scholar]
- LaFountain JR., Jr Cole RW. Rieder CL. Partner telomeres during anaphase in crane-fly spermatocytes are connected by an elastic tether that exerts a backward force and resists poleward motion. J Cell Sci. 2002;115:1541–1549. doi: 10.1242/jcs.115.7.1541. [DOI] [PubMed] [Google Scholar]
- Chan KL, North PS. Hickson ID. BLM is required for faithful chromosome segregation and its localization defines a class of ultrafine anaphase bridges. EMBO J. 2007;26:3397–3409. doi: 10.1038/sj.emboj.7601777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baumann C, Korner R, Hofmann K. Nigg EA. PICH, a centromere-associated SNF2 family ATPase, is regulated by Plk1 and required for the spindle checkpoint. Cell. 2007;128:101–114. doi: 10.1016/j.cell.2006.11.041. [DOI] [PubMed] [Google Scholar]
- Biebricher A, Hirano S, Enzlin JH, Wiechens N, Streicher WW, Huttner D, Wang LH, Nigg EA, Owen-Hughes T, Liu Y, et al. PICH: a DNA translocase specially adapted for processing anaphase bridge DNA. Mol Cell. 2013;51:691–701. doi: 10.1016/j.molcel.2013.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Germann SM, Schramke V, Pedersen RT, Gallina I, Eckert-Boulet N, Oestergaard VH. Lisby M. TopBP1/Dpb11 binds DNA anaphase bridges to prevent genome instability. J Cell Biol. 2014;204:45–59. doi: 10.1083/jcb.201305157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez R, Viera A, Berenguer I, Llano E, Pendas AM, Barbero JL, Kikuchi A. Suja JA. Cohesin removal precedes topoisomerase IIalpha-dependent decatenation at centromeres in male mammalian meiosis II. Chromosoma. 2014;123:129–146. doi: 10.1007/s00412-013-0434-9. [DOI] [PubMed] [Google Scholar]
- Klein F, Mahr P, Galova M, Buonomo SB, Michaelis C, Nairz K. Nasmyth K. A central role for cohesins in sister chromatid cohesion, formation of axial elements, and recombination during yeast meiosis. Cell. 1999;98:91–103. doi: 10.1016/S0092-8674(00)80609-1. [DOI] [PubMed] [Google Scholar]
- Cheng CH, Lo YH, Liang SS, Ti SC, Lin FM, Yeh CH, Huang HY. Wang TF. SUMO modifications control assembly of synaptonemal complex and polycomplex in meiosis of Saccharomyces cerevisiae. Genes Dev. 2006;20:2067–2081. doi: 10.1101/gad.1430406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng EY, Hunt PA, Naluai-Cecchini TA, Fligner CL, Fujimoto VY, Pasternack TL, Schwartz JM, Steinauer JE, Woodruff TJ, Cherry SM, et al. Meiotic recombination in human oocytes. PLoS Genet. 2009;5:e1000661. doi: 10.1371/journal.pgen.1000661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliver TR, Feingold E, Yu K, Cheung V, Tinker S, Yadav-Shah M, Masse N. Sherman SL. New insights into human nondisjunction of chromosome 21 in oocytes. PLoS Genet. 2008;4:e1000033. doi: 10.1371/journal.pgen.1000033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fledel-Alon A, Wilson DJ, Broman K, Wen X, Ober C, Coop G. Przeworski M. Broad-scale recombination patterns underlying proper disjunction in humans. PLoS Genet. 2009;5:e1000658. doi: 10.1371/journal.pgen.1000658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassold T, Hunt PA. Sherman S. Trisomy in humans: incidence, origin and etiology. Curr Opin Genet Dev. 1993;3:398–403. doi: 10.1016/0959-437x(93)90111-2. [DOI] [PubMed] [Google Scholar]
- Lao JP, Cloud V, Huang CC, Grubb J, Thacker D, Lee CY, Dresser ME, Hunter N. Bishop DK. Meiotic crossover control by concerted action of Rad51-Dmc1 in homolog template bias and robust homeostatic regulation. PLoS Genet. 2013;9:e1003978. doi: 10.1371/journal.pgen.1003978. [DOI] [PMC free article] [PubMed] [Google Scholar]