

Abstract
We address adaptive vs. maladaptive responses to hypoxemia in healthy humans and hypoxic-tolerant species during wakefulness, sleep, and exercise. Types of hypoxemia discussed include short-term and life-long residence at high altitudes, the intermittent hypoxemia attending sleep apnea, or training regimens prescribed for endurance athletes. We propose that hypoxia presents an insult to O2 transport, which is poorly tolerated in most humans because of the physiological cost.
This brief essay questions whether we humans have begun to take hypoxic exposure for granted, at least in a biological sense. For example, more than 25 million humans now live above an altitude of 3,000 m, a point at which arterial O2 saturations even at rest approach the steep portion of the HbO2 dissociation curve (below ∼90% SaO2), and recently over 50,000 persons have relocated to reside between 2,500 and 3,500 m in the beautiful mountains of Colorado. More severe, albeit shorter duration, exposures to hypoxia are popularly available for $50,000+ for Everest expeditions or over the Internet in the form of “hypoxicators,” which promise to simulate up to 6,500-m altitude in your living room to treat asthma and hypertension and promote general well being. Literally thousands of athletes throughout the world are exposing themselves nightly to hypoxia for extended periods in the hopes of acquiring even small gains in blood O2 carrying capacity to improve endurance performance at sea level. Finally, although not entirely intentional, our addiction to high-calorie diets and a sedentary lifestyle in Western cultures brings with it an obesity epidemic, which in turn has contributed to the doubling of the prevalence of serious sleep apnea and its accompanying cyclical, intermittent hypoxemia (IH) (88). Our question is: Do humans truly “adapt” or even benefit from hypoxic exposures of various durations and magnitudes or do the biological “costs” of these apparent adaptations approach or even exceed the benefits? Similar questions of biological cost vs. benefit were posed 50 years ago when premature infants with severely compromised lung function were treated with supplemental O2, a practice that relieved much of their arterial hypoxemia, while at the same time introducing the toxic long-term consequences of excessive oxygenation (122). Concern over the appropriateness of departures-plus or minus–from normoxic levels of arterial oxygenation continues.
Positive Outcomes
There is considerable evidence in support of positive “adaptations” to hypoxia–even “conquering” hypoxia–in humans, including the following examples.
The sigmoid shape of the HbO2 disassociation curve permits substantial (up to one-third) reductions from the normal arterial Po2 before serious reductions occur in arterial O2 saturation and content and therefore in O2 transport. Accordingly, at no discernable extra biological cost, healthy humans (at rest, at least) easily and efficiently undergo brief exposures to the hypoxemia attending either the compression of commercial airliner cabins to ∼2,500-m altitude or the hypoventilation normally attending sleep.
Powerful hypoxic-induced gene transcription factors (HIF-1/HIF-2 alpha) are activated even very early upon hypoxic exposure, guaranteeing time-dependent upregulation of cardiorespiratory and hematological responses aimed at limiting deficits in O2 transport (O2 transport = arterial O2 content × blood flow) (93).
Hundreds of humans–even a few octogenarians–have summited mountains where the barometric pressure and therefore inspired Po2 is less than one-third that at sea level, usually with the help of supplemental O2 but even without it in some rare cases.
Certain types of experimentally imposed IH of low-frequency, moderate severity and short duration have been shown to illicit synaptic plasticity, with potential clinical benefits (see IH section below).
With lifelong hypoxia, remodeling and angiogenesis in many organs brings improved efficiency in gas transport.
On the other hand, we believe a significant case can be made that the biological costs of many types of hypoxic adaptations can sometimes outweigh their benefits. We now explore this premise under various types of hypoxic scenarios.
Sojourn to Hypoxia in the Native Lowlander
The Pros and Cons of Chemoreceptor Adaptation
The primary oxygen sensor of the body, i.e., the carotid chemoreceptor, is ideally located anatomically between the lung and the brain at the bifurcation of the carotid artery (see FIGURE 1A). It responds within a few seconds to the onset of arterial hypoxemia, eliciting hyperventilation and increased sympathetic nerve activity. Within a few hours, and even more so over several days, the carotid chemoreceptors undergo a gradual rise in sensitivity achieved via increased protein expression in the form of newly generated type 1 glomus cells (see FIGURE 1B). This increased sensory input from the hypoxemic carotid body is further amplified within the central nervous system at several potential locations in the chemoreceptor pathway, which may include the cells of the central CO2-sensitive chemoreceptors, the hypothalamus, and the ventral lateral medulla (28, 42). This time-dependent increase in chemosensitivity appears to be specific to hypoxemia, per se, as shown by the absence of further sensitization over time when carotid chemoreceptors were stimulated with hypercapnia (16). The resultant, time-dependent increase in ventilation is critical to acclimatization in the sojourner because it minimizes the reduction in alveolar Po2 as inspired Po2 falls with increasing altitude. For example, at 4,300-m altitude, time-dependent hyperventilation over a 2-wk period accounts for a rise of a full 10% in resting arterial O2 saturation, and at the summit of Everest (PiO2 ∼ one-third of sea-level), the extreme hyperventilation is essential to maintaining alveolar Po2 above 20 Torr and therefore to survival. Enhanced time-dependent sympathetic activation via the sensitized carotid chemoreceptor (45) (FIGURE 2) may also assist cardiac contractility and contribute to heart rate elevation in hypoxia, with both helping to increase blood flow and minimize the reduction in O2 transport in the face of a falling CaO2.
FIGURE 1.

Human carotid chemoreceptor and immunofluorescence staining of bromodeoxyuridine
A: human carotid chemoreceptor, located at the bifurcation of the common carotid artery. In health and in normoxia, the carotid body is usually 1–7 mm in length and 1–2 mm in diameter. The carotid chemoreceptor is located ∼10- to 15-s circulation time from the lung (under resting conditions), has sensory output via the carotid sinus nerve to the petrosal ganglion, with central projections via the nucleus of the solitary tract to the medullary pattern generation neurons, central chemoreceptor cells of the retrotrapezoid nucleus, and the paraventricular nucleus. Figure reproduced from Ref. 53 with permission from BMJ Publishing Group, Ltd. B: immunofluorescence staining of bromodeoxyuridine (BrdU), a uridine analog that is stably incorporated into cells undergoing DNA synthesis, in carotid body type I cells from rats maintained in normoxia, exposed to sustained normobaric hypoxia (FiO2, 0.12) for 1 (A) and 2 days, and rats exposed to hypoxia for 7 days, then returned to normoxia for another 30 days. Sustained hypoxia caused glomus cell proliferation that occurred mainly in the first 3 days of exposure. The new glomus cells survived for at least 30 days after return to normoxia. Scale bar, 50 μm. Figure reproduced from Ref. 121 with permission.
FIGURE 2.

Hypoxic effects on muscle sympathetic nerve activity and on exercise ventilation and work of breathing
A: time-dependent increase in muscle sympathetic nerve activity (MSNA) in a healthy human during sojourn at 4,100-m altitude. MSNA frequency remained elevated for up to 5 days following return to sea level. Figure reproduced with permission from M. Sander [also see Hansen et al. (45)]. B: effects on ventilation, expiratory flow limitation, and inspiratory and expiratory esophageal pressures (Pes) during constant load, heavy-intensity cycle exercise at 273 W in normoxia (95% SaO2, 69% of peak work rate) vs. acute hypoxia (FiO2, 15; SaO2, ∼81%; 99% of peak work rate) in trained cyclists. At right, note the increased ventilation in hypoxia vs. normoxia at 3 min and at termination (9 min) of the same 273-W work rate. The maximum volitional flow-volume loop (obtained at rest) is the largest loop shown (inspiration down), and the three loops within this are average tidal breaths at rest and during exercise at 3 min and exercise termination. Note the occurrence of expiratory flow limitation in hypoxia near end-exercise. At left, negative Pes occurs during inspiration and positive Pes during expiration. These values for Pes were obtained at end-exercise in both normoxia and hypoxia. Ratings of perceived dyspnea on a 10-point scale at end exercise of equal durations averaged 4.4 ± 0.5 in normoxia and 8.7 ± 0.3 in acute hypoxia. Figure reproduced from Ref. 4 with permission.
So what's not to like about carotid chemoreceptor sensitization in a hypoxic environment, including its central amplification effects and its after-effects on ventilation and sympathetic nerve activity, which are sustained for several days upon restoration of normoxia? Consider the following negative consequences of carotid chemoreceptor sensitization.
The hyperventilatory response to exercise in hypoxia brings with it expiratory flow limitation and excessive levels of negative inspiratory and positive expiratory intrathoracic pressures and respiratory muscle work, leading to extreme dyspnea (see FIGURE 2B) (4, 118). Studies utilizing respiratory muscle unloading or expiratory loading during exercise in normoxia or acute hypoxia have demonstrated that such increases in intrathoracic pressures will compromise stroke volume and limb locomotor muscle blood flow leading to exacerbation of limb fatigue and compromised exercise performance (4, 70, 118). Reinhold Messner, during his ascent (without supplemental O2) to the summit of Everest, commented on his overwhelming pre-occupation with the labor of breathing: “I am nothing more than a single, narrow gasping lung floating over the mists and the summits!” (69).
The vasoconstrictor effects of supernormal levels of sympathoexcitation would be expected to oppose the vasodilatory effects of local hypoxemia in the systemic circulation (see Local Blood Flow Regulation in the Systemic and Pulmonary Circulation below), thereby contributing to systemic hypertension and increased vascular resistance at rest and especially during exercise accompanying prolonged sojourn to high altitude (126–128). These hypertensive effects persist for several days upon returning to sea level. A compromised endothelial responsiveness to local vasodilators (ATP and adenosine) and increased arterial stiffness have also been observed in sojourners and likely contribute significantly to the systemic hypertension and raised vascular resistance at rest and exercise (21, 59).
Hypoxia also increases carotid chemoreceptor sensitivity to CO2 both above and below eupnea, which steepens the slope of the ventilatory response to CO2 and moves the hypocapnic-induced apneic threshold very close to steady-state eupneic PaCO2, thereby precipitating unstable ventilatory overshoots and undershoots, especially during sleep (30, 130) (see FIGURE 3). The accompanying periodic breathing during sleep causes severe transient IH (superimposed on the existing constant hypoxemia), transient arousals and sleep disruption, and also likely exacerbates further sympathetic activation following the termination of each apneic cycle. The severity of sleep-induced reductions in SaO2 in the sojourner has been associated with symptoms of acute mountain sickness (20). Moreover, the elevated MSNA, even with short-term IH, persists for several days upon restoration of normoxia (131). Accordingly, these nocturnal events likely contribute significantly to the systemic hypertension during both waking and sleeping hours in hypoxia as well as to the now well documented effects of IH on reducing reactivity of the vascular endothelium (also see IH below).
FIGURE 3.

Periodic breathing with apneas
Periodic breathing with apneas interspersed with three to four augmented tidal volumes occurring in 15- to 25-s cycles thought NREM sleep in a healthy sojourner to 4,100-m altitude. The “average” SaO2 is substantially reduced, and PaCO2 increased from wakefulness to NREM sleep in hypoxia (not shown), although little difference or even slight increases in the average SaO2 values between periodic and non-periodic patterns have been reported during sleep in hypoxia (14, 20, 43, 124). As explained in Adaptive IH, it is the oscillatory periods of HbO2 desaturation and resaturation that enhance the rate of free-radical production and their cardiovascular sequelae “. . . consistent with the notion that relevant enzymes are generating superoxide in response to the rate at which the electron receptor (O2) is introduced into the cell” (60). Reproduced from Ref. 14 with permission from J Physiol.
A carotid chemoreceptor-dependent diuresis and reduction in plasma volume by as much as 20% occurs over the initial weeks at moderate altitudes (8, 96) and likely contributes to the reduced left-ventricular filling pressure and stroke volume and cardiac output during exercise in hypoxia (2, 8).
Local Blood Flow Regulation in the Systemic and Pulmonary Circulation
Acute hypoxia causes vasodilation in most systemic and cerebral arterioles. This is an important adaptive mechanism to minimize reductions in O2 transport (CaO2·Q̇) at high altitudes. On the other hand, this adaptive hyperemia in the brain has been linked to increases in intracranial pressure, symptoms of severe headache, and even cerebral edema in extreme cases (125). Similarly, high blood flows to vasodilated retinal vessels likely contribute to the not uncommon occurrence of retinal hemorrhage at high attitudes (68).
Local regulation of vascular resistance in the lung is likely the most costly “adaptation” to hypoxia. The lung vasculature receives all of the cardiac output; accordingly, it is a thin-walled, highly compliant vasculature not subject to the same degree of autonomic control as the systemic circulation and with a resistance to flow that is normally <1/20th of the systemic circulation at rest. In contrast to arterioles of the systemic circulation, pulmonary arterioles constrict in response to local hypoxia. This is clearly “adaptive” to pulmonary gas transport if a relatively small portion of the lung is underventilated and hypoxic (for example with heterogeneous airway narrowing/closure), thereby allowing blood flow to be redirected to a better ventilated, non-hypoxic area of the lung. Unfortunately, at high altitudes, “global” alveolar hypoxia prevails, promoting widespread pulmonary arteriolar vasoconstriction. After as little as only 24–48 h of hypoxic exposure, pulmonary vascular remodeling begins to occur; thus pulmonary hypertension is not completely reversible upon restoration of normoxia (33, 64). This combination of vasoconstriction and especially vascular remodeling reduces the distensibility of pulmonary resistance vessels. Accordingly, as pulmonary blood flow increases with exercise, the normal fall in pulmonary vascular resistance is prevented, and a sharp rise in pulmonary artery pressure occurs (78, 85). The consequences to the exercising sojourner at high altitude are a substantial rise in afterload on the right ventricle and even pulmonary edema in some cases. Importantly, the inhomogeneous nature of global hypoxic pulmonary vasoconstriction leads to high blood flow-induced increased capillary pressures in much of the lung (47). Since this hydrostatic pressure in the pulmonary capillaries rises with high flow rates, the movement of plasma water into the lung interstitium increases, sometimes to the point where it exceeds the drainage capability of the thoracic lymphatics (i.e., the lung's “sump pump”), and alveolar flooding and high altitude pulmonary edema (HAPE) ensue. This scenario is perhaps the prime example of what is the most serious and widespread consequence in hypoxia of a “well intentioned” adaptive mechanism, i.e., local hypoxic-induced pulmonary vasoconstriction, gone awry in a globally hypoxic environment.
This paradoxical effect in the hypoxic lung is reinforced by another time-dependent “adaptive” mechanism in the form of increased red blood cell production, which enhances O2-carrying capacities but also increases blood viscosity, thereby further raising pulmonary vascular resistance, especially during exercise (78)1. In addition to high capillary hydrostatic pressures, HAPE might also be precipitated by hypoxic impairment of sodium transport in the lung (65), an organ which, in normoxia, has a very high capacity for epithelial sodium reabsorption. As with most cardiorespiratory responses to hypoxia, susceptibility to hypoxic pulmonary vasoconstriction and to HAPE varies markedly among healthy sea-level natives and is exacerbated by exercise (78).
Exercise Performance
V̇o2 max and especially endurance exercise performance are highly sensitive to even very small alterations in O2 transport to locomotor muscles and/or to brain. At altitudes as low as 800-1,000 m above sea level, performance is impaired, especially in many highly trained endurance athletes who appear to be especially susceptible (44). These negative effects became widely known as early as the 1968 Mexico City Summer Olympics (Pb 580 Torr, PiO2 112 Torr), during which world record holders in many distance running events (i.e., >800 m) failed to medal and even in some cases to qualify for event finals, whereas high-altitude natives from Kenya and Ethiopia made their Olympic debuts by medaling in all events over 800 m. As outlined in FIGURE 4 (also see FIGURE 4 legend), there appear to be three types of limitations that hypoxia, especially chronic hypoxia, can inflict on systemic and cerebral O2 transport in the sojourner to the detriment of exercise performance.
FIGURE 4.

Three potential routes to exacerbation of exercise-induced central and peripheral (locomotor muscle) fatigue and reduction in exercise performance in hypoxia
1: reduced O2 transport to locomotor muscles via a) exercise-induced arterial hypoxemia (a); reductions in locomotor muscle blood flow secondary to an augmented sympathetic vasoconstriction of locomotor muscle vasculature elicited by high levels of respiratory muscle work and fatigue (b) (29); and/or stroke volume and cardiac output may be compromised via high left ventricular afterload induced by positive intrathoracic pressure on expiration in the face of airway expiratory flow limitation (70) (see also FIGURE 2A) or by increased pulmonary vascular resistance causing interventricular septal deviation toward the left ventricle, thereby altering left ventricular geometry and delaying filling (c) (56, 79). 2: CNS hypoxia, especially at extreme levels of arterial O2 desaturation (<75% SaO2), will inhibit central motor command (likely linked to reduced neurotransmitter turnover) and limit performance even in the face of limited amounts of peripheral muscle fatigue (6). 3: peripheral and central fatigue are causally linked via feedback inhibition of central motor output through group III–IV muscle afferents. When these afferents were partially blocked during a cycling time trial, the cyclist “chose” a much higher power output (vs. placebo control) in the initial half of the trial, leading to excessive muscle metabolite accumulation, reduced power output, and excessive locomotor muscle fatigue at end-exercise (5).
Why do the highly trained show greater deficits in V̇o2 max and performance in hypoxia? And why sometimes even at altitudes <1,000 m above sea level? One explanation is to be found in the mild to moderate exercise-induced arterial hypoxemia even at sea level that occurs in some endurance trained runners, especially in female runners (25, 31, 46, 123). With even mild elevations in altitude, these athletes are most likely to exacerbate their exercise-induced arterial hypoxemia, thus driving arterial HbO2 saturation down the steep portion of the HbO2 dissociation curve (27, 34, 39, 57). Additional potential limitations imposed via hypoxic exposure, especially in the highly fit, include 1) increased pulmonary arterial pressure and right heart wall stress in the presence of high pulmonary blood flows in excess of 20–30 l/min (56, 80), combined with hypoxic-induced pulmonary vasoconstriction and arteriolar remodeling (see above); and 2) the excessive work of breathing with positive expiratory intrathoracic pressures secondary to expiratory flow limitation at high ventilatory demand (51, 70) (also see FIGURE 2B). Accordingly, in acute hypoxia in healthy athletes, reducing pulmonary vascular resistance via endothelin receptor blockade has been shown to improve endurance exercise performance (78, 79), and reducing intrathoracic pressures and the work of breathing (with mechanical ventilation) slowed the rate of development of quadriceps fatigue, reduced symptoms of both dyspnea and limb discomfort, and improved exercise performance (4).
In summary, we propose that, while the sojourner to high altitudes–even the highly fit–will likely improve O2 transport as alveolar ventilation, HbO2 saturation as well as red cell mass all improve over time, the biological cost of many “adaptations” is substantial, thereby eroding and sometimes exceeding the benefits. Not only are maladaptations such as super-enhanced sympathetic nerve activity and its negative cardiovascular sequelae intensified with duration of hypoxic exposure, they are carried over to the post-hypoxic period, with absolutely no apparent “adaptive” benefit! Furthermore, enhanced chemoreceptor sensitivity (along with reduced baroreceptor sensitivity) and its negative cardiovascular consequences appear similar in many respects to the “adaptations” to the low blood flow-induced “stagnant hypoxia” of the carotid chemoreceptors reported in patients with chronic heart failure (102)2 or to the IH of severe sleep apnea (see IH section below). Apart from our focus on cardiorespiratory maladaptations in the sojourner, we need to also mention the well documented impairment of cognitive function in sojourners to high altitudes, especially above 3,000 m, often with residual after-effects upon descent (125), as well as the significant reduction in immune function with increased probability of infection in the high-altitude sojourner, induced by downregulation of dendritic or lymphocyte cell functions that lead to impairment in immune memory development (83).
Adaptation/Maladaptation in the Native or Long-Term Resident of High Altitudes
Sufficient evidence has accumulated to demonstrate that most natives and long-term residents of altitudes >2,500 m show unique levels of organ system angiogenesis and remodeling; but like the sojourner, this “remodeling” has both adaptive and maladaptive consequences. For example, on the adaptive side, substantial hypoxic-induced angiogenesis in the coronary circulation enhances myocardial O2 transport in a hypoxic environment. Furthermore, in the lung of natives and long-term residents of high altitudes (22) and in young animals exposed to long-term hypoxia (49), alveolar-capillary proliferation occurs, inducing large increases in the pulmonary diffusion surface as manifested in substantial increases in diffusion capacity at both rest and exercise. At the same time, ventilatory chemosensitivity is substantially blunted in the altitude resident both at rest and during exercise. This combination of remodeling means that the altitude resident shows little or no hyperventilation or dyspnea during exercise relative to that experienced by the sojourner at high altitude, and yet arterial Po2 is maintained similar to that in the sojourner because of enhanced pulmonary O2 transport and a greatly narrowed alveolar-to-arterial O2 difference (19, 26, 28). This highly efficient gas exchange during exercise in hypoxia does not necessarily translate into a higher peak exercise performance, which is dependent on many factors but likely allows the highlander a more “comfortable” active, dyspnea-free daily lifestyle at high altitude without sacrificing arterial O2 saturation. At the same time, remodeling in the lung also has significant maladaptive side effects because high-altitude residents retain their hypoxic pulmonary vasoconstriction, which over time results in persistent pulmonary hypertension at rest and especially as cardiac output increases during even mild-intensity exercise. This pulmonary hypertension is resistant to correction to normal upon acute inhalation of hyperoxia or upon inhalation of the potent vasodilator nitric oxide and often requires up to 2 years residence at sea level for normalization of pulmonary vascular resistance (7, 8, 86, 103, 113, 114, 120) (see FIGURE 5).
FIGURE 5.
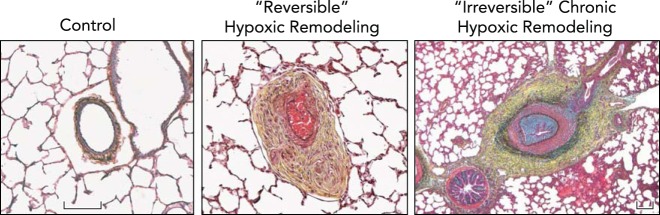
Chronic hypoxic exposure leads to severe pulmonary vascular remodeling, which is often reversible upon acute return to normoxia, but in extreme cases may be irreversible
Distal pulmonary arteries of neonatal calves raised at ambient altitude (Control) are thin-walled and highly compliant, with minimal surrounding adventitia, whereas neonatal calves, exposed to 2 wk of hypobaric hypoxia, develop severe pulmonary hypertension, with mean PAP often exceeding 80 mmHg, and have pulmonary arteries that demonstrate marked fibro-proliferative changes in media and adventitia accompanied by formation of vasa vasorum (Reversible Hypoxic Remodeling). This profound remodeling, however, is reversible, and spontaneously gradually resolves when animals are returned to normobaric conditions. In older (5- to 9-mo-old) calves that were raised at moderately high altitudes (3,000-3,500 m), hypoxia-associated vascular remodeling and pulmonary hypertensive process can become severe and not regress in response to normoxia (Irreversible Chronic Hypoxic Remodeling). These changes are observed in animals exhibiting signs and symptoms of severe right heart failure. Remodeling of pulmonary arteries in these animals include severe neointimal, often occlusive, lesions as well as profound adventitial and medial thickening and fibrosis. Figure and legend were provided by Prof. Kurt Stenmark, University of Colorado, and used with his permission.
Overproduction of red blood cells in high-altitude residents, a condition labeled “chronic mountain sickness” (or Monge's disease), is an adaptation that enhances O2 transport in the face of HbO2 desaturation. Unfortunately, it also increases viscosity in the systemic and pulmonary circulation and exacerbates the high-altitude resident's pulmonary hypertension at rest and especially during exercise (78, 113).
A fundamental manifestation of the consequences of cardiovascular maladaptation in the high-altitude resident is found in the reproductive process. Birth weight is reduced ∼100 g for every 1,000-m increase in altitude above 1,500-m altitude and with a more pronounced reduction in birth weight above 2,500-m altitude due to a slowing of fetal growth, in turn attributable to hypoxic effects on retarding uterine artery development and reducing placental blood flow. Furthermore, the prevalence of maternal hypertension and preeclampsia is increased threefold above 3,000 m vs. that at sea level, exerting detrimental effects on intrauterine and fetal morality. In addition, marked pulmonary hypertension in the human neonate often occurs because, at birth when vasodilation is needed (to support the onset of blood flow through the lung), instead pulmonary vasoconstriction occurs in the face of global alveolar hypoxia. (50, 71).
So, for at least much of the high-altitude resident population with up to a few thousand years of ancestry, such adaptive mechanisms as erythropoiesis and lung and carotid chemoreceptor remodeling, which favor minimizing reductions and maximizing efficiency in O2 transport, also have maladaptive consequences to the highlander fetus, newborn, and adult. Interestingly, these characteristics are prevalent in native highlanders with widely divergent ancestries of hypoxic exposure from 3 to 4 generations in the natives of Leadville, Colorado to 40 to 50 generations in Andean natives.
On the other hand, recent evidence suggests that genetic selection in Tibetan natives with >30,000 years of exposure to altitudes above 3,000 m may have avoided at least some of these maladaptive responses. That is, there is solid evidence that excessive erythropoiesis is not present in the Tibetan, and to date there are suggestions, but not yet conclusive evidence in large numbers of subjects, of higher arterial O2 saturation, less pulmonary vasoconstriction, greater upregulation of the production of the vasodilator NO, decreased wall-to-lumen ratio in the carotid artery, and higher utero-placental blood flow with lower infant mortality (9–11, 59, 72, 111). Genomic surveys comparing native Tibetans with Han Chinese have revealed population-specific allele frequency changes (9) underlying several important loci in the genetic adaptation to high altitude, including the EPAS1 locus, which encodes the antioxidant HIF-2α (132).
True Hypoxic Tolerance
Unlike humans, truly hypoxia-tolerant species demonstrate adaptive responses to hypoxic exposure and shun most of those responses with high biological cost. A few examples are given below.
Bar headed geese migrating over the Himalayas face the challenge of sustaining metabolic rates during wing flapping in the range of 10–15 times resting levels and near maximum heart rates while exposed to altitudes in excess of 5,000 m and occasionally even above 6,000 m. These geese utilize a highly efficient cross-current gas-exchange system in the lung such that Po2 in arterial blood can even exceed that in expired air, and they do not experience hypoxic pulmonary vasoconstriction. They also experience a marked cerebral vasodilation without the counteracting effects of hypocapnic cerebral vasoconstriction or the devastating consequences of an increased intracranial pressure (104, 105).
Reproduction in the high-altitude dwelling llama is exquisitely protected via enhanced placental O2 transport through a very thin maternal:fetal membrane with fenestrated capillaries in the placenta. In the llama neonate at birth, the pulmonary vasculature immediately undergoes hypoxic vasodilation to accommodate the onset of blood flow through the lung without hypertensive consequences (61).
The Crucian Carp, a relative of the lowly goldfish, remains physically active and alert throughout the winter in frozen lakes that are completely devoid of oxygen. While anaerobic metabolism prevails under these anoxic conditions, this species employs a unique enzymatic conversion of lactic acid into ethanol, which then diffuses out over the gills (15).
The human fetus has a strong pulmonary vasoconstriction in response to hypoxia, which is adaptive in utero as venous return is shunted through the heart. The human fetus, like birds and llamas, also has a markedly left-shifted HbO2 dissociation curve to enhance O2 loading in a relatively hypoxic environment in utero. Unfortunately in the human, at birth, hypoxic pulmonary vasoconstriction remains and persists throughout life, whereas the HbO2 dissociation curve gradually reduces its affinity for O2.
Upon exposure to acute hypoxia, the laboratory rodent, unlike the adult human but like other small mammals, quickly lowers metabolic rate and core body temperature, thereby reducing much of the associated high costs of cardioventilatory responses to hypoxia without sacrificing blood gas transport (73, 77). Paradoxically, one important mediator of this hypoxic-induced hypometabolism in small mammals may be found in a carotid chemoreceptor-induced reduction in sympathetic outflow to brown adipose tissue (75).
Intermittent Hypoxia
The biological responses to IH may be adaptive, even “therapeutic,” or maladaptive, depending on the severity of the hypoxemia, its frequency of occurrence, its duration, and, importantly, the “pattern” and timing of each of the HbO2 desaturation/resaturation cycles (3, 60, 67, 92, 106).
Adaptive IH
On the adaptive side, short-term exposures (via manipulation of FiO2) in rodents and humans to a few weeks of daily sessions consisting of 10–15 1- to 2-min bouts of moderate isocapnic hypoxemia (SaO2 75–80%) alternating with equal durations of normoxia have been shown to yield several benefits without maladaptive cardiovascular sequelae. Most notably, this noncyclical pattern, short duration, and mild severity type of IH induces respiratory motoneuron plasticity characterized by a progressive increase in phrenic and hypoglossal nerve outputs following IH application, with enhancement of ventilation in rodents and even in humans with spinal cord injury (3, 24, 37, 67, 117). Apparently, these forms of short-term, low-frequency moderate IH alter the expression of hypoxic-sensitive growth and trophic factors within respiratory motoneurons (3, 24, 37). These forms of IH treatment have also been shown to improve cardiac function in both healthy and transgenic mice in heart failure (81, 82), to protect against myocardial infarction (13), to enhance innate immunity, and to lower blood pressure in hypertensive patients (107), although these latter studies in humans were not placebo controlled. A potential role for reactive oxygen species (ROS) in the response to different forms and severities of IH has been suggested, with low levels of ROS emerging as protective of specific cells, tissues, and organs but higher levels being detrimental (see below) (3, 100).
Maladaptive Cyclical IH
The maladaptive side of IH may be found in the cardiovascular, metabolic, and cognitive consequences of obstructive sleep apnea, including excessive oxidative stress and inflammation, increased cerebral and systemic vascular resistance, systemic and pulmonary hypertension, and increased insulin resistance (3, 12, 30, 60, 66, 97, 129). It is important to point out that, in addition to severe, long-standing cyclical IH, the patient with OSA often presents with several additional confounding causes of cardiovascular morbidity such as obesity, diabetes, as well as intermittent sleep disruption, and exaggerated swings in intrathoracic pressure occurring repeatedly throughout the night. Nevertheless, animal studies that simulate the severity, frequency, and duty cycle of the OSA patient's nightly cyclical patterns of IH have shown key maladaptive contributions from IH alone (3, 17, 30, 60, 66, 92).
The underlying theory contends that, while continuous, sustained hypoxia promotes the increased expression of both the pro-oxidant HIF1-α-dependent enzymes as well as the antioxidant HIF2-α-dependent enzymes, with high-frequency, oscillatory forms of IH akin to those attending periodic breathing, the antioxidant effect of HIF2-α is downregulated within the O2-sensitive glomus cells of the carotid chemoreceptor (3, 94, 133). This imbalance in the HIF isoforms results in extraordinarily high levels of ROS in the carotid chemoreceptor glomus cells, which increases carotid sinus nerve activity, leading over time to enhanced activity of neurons in downstream elements of the carotid chemoreflex pathway, i.e., in the NTS, RVLM, and paraventricular nucleus of the hypothalamus (55, 87, 102, 106, 108). The enhanced chemoreceptor sensitivity and sympathoexcitation that occur during IH persist upon reoxygenation well beyond the nocturnal IH exposure. This persistent vasoconstrictor sympathetic outflow combined with the impaired endothelial vascular responsiveness induced by IH-mediated ROS (32, 52, 74, 89) is an important mediator of daytime IH-induced hypertension. The consequences of this imbalance between oxidant vs. antioxidant HIF isoforms embodies the theme expressed throughout our essay of hypoxic-induced adaptations and maladaptations emanating from a common source.
Most recently, cardiovascular, metabolic, and cognitive effects of nocturnal cyclical IH attending OSA have been tested experimentally (using dynamic manipulation of FiO2) in healthy young adult humans over daily periods of 4–6 h and a range of 4 days to 4 wk. In general, the after-effects are similar in direction, but with less severe and persistent consequences, to those obtained in rodents with more severe and longer periods of IH. Persistent after-effects include increased markers of oxidative stress and chemosensitivity, impaired insulin sensitivity, elevated daytime MSNA combined with reductions in baroreceptor sensitivity, impaired vascular endothelial function, increased peripheral and cerebral vascular resistance with modest increases in daytime SBP and DBP (see FIGURE 6) (18, 35, 38, 62, 91, 115, 116). The increased oxidative stress and BP resulting from a few nights of oscillatory IH were prevented via angiotensin receptor blockade (90).
FIGURE 6.

Effects on ambulatory systemic blood pressure of nocturnal intermittent, oscillating hypoxemia
Hour-by-hour effects on ambulatory systemic blood pressure of nocturnal intermittent, oscillating hypoxemia consisting of 8 h of daily of oscillatory poikilocapnic hypoxia (FiO2 of 0.13 every 105 s alternated with 15 s of FiO2 of 1.0, producing periodic swings of SpO2 of ∼88–98%) in healthy young adults. Note that increases in blood pressure resulting from IH occurred 1) just before waking (5–7 AM), then returned to normal until increasing again over the late afternoon and evening; 2) during the daytime hours after 1 night of IH, and increased further after 13 IH nights; and 3) returned to normal after 5 nights in normoxia. The IH effects on daytime BP were accompanied by significant increases in MSNA burst frequency and reductions in baroreceptor sensitivity. Figure was reproduced from Ref. 116 with permission from the European Respiratory Society.
Hypoxic Training, IH, and Exercise Performance
Given the negative nocturnal and daytime consequences of the oscillatory sleep apnea form of IH, it would seem imprudent to impose this insult intentionally on healthy subjects. However, this may well be the case in training practices in which hundreds of elite athletes throughout the world live and train at high altitudes (live high:train high) or sleep in normobaric hypoxic environments and train each day in normoxia (live high:train low). The goal here is “legal” blood doping in which hypoxic exposure is used to stimulate erythropoiesis, increase Hb mass, and thereby enhance O2-carrying capacity and O2 transport to muscle. There is little doubt that even 1 or 2 g of added hemoglobin (which adds ∼1.2–2.5 ml O2/100 ml or ∼10–15% to arterial O2 content) will significantly and consistently increase V̇o2 max and performance (19a, 63, 119). However, the effects of these hypoxic training practices have produced highly heterogeneous results, even among elite endurance athletes in both their influence on Hb mass (especially in the live high:train low regimen) and on sea-level endurance performance, compared with living and training in normoxia (23, 44, 48, 58, 99, 101, 110) (see FIGURE 7). Recent evidence shows that even in sustained (24 h/day) hypoxia, a 3-wk sojourn at 3,454-m altitude was required to elicit consistent, significant increases in Hb mass among healthy subjects, which returned to control values after 2 wk back at sea level (109).
FIGURE 7.
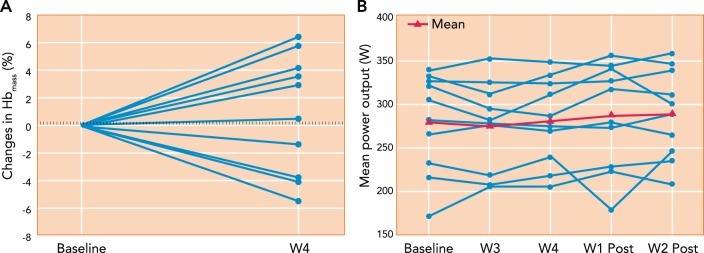
Effects of “live high:train low” training regimen in 10 endurance-trained cyclists on Hb mass and mean power output during a 26-km time trial
The cyclists were exposed to daily normobaric hypoxia simulating 3,000-m altitude for 16 h/day for 28 nights and trained daily in normoxia. Note the high individual variability in the changes in Hb mass and mean power output during the time trials. The dashed lines represent the random error of the CO rebreathe test used for Hb mass. The group mean changes in Hb mass, time trial mean power output or performance, and V̇o2 max (not shown) were not significantly different from those in a placebo group (n = 6) exposed to normoxia. Figure was reproduced from Ref. 110 with permission.
A multitude of factors may contribute significantly to this striking individual variability in responsiveness to hypoxic training regimens. These include such variables as ferritin levels, beginning RBC mass, severity and duration of hypoxic exposure, the degree to which exercising at high altitudes will reduce training intensity, and the variable placebo effects among the athletic subjects of “expected” improvements in performance (58, 99, 101, 110). Might the cardiovascular consequences of sustained hypoxia or IH also contribute? On the one hand, as might be expected, exposure of athletes for 15 days to 2 h/day of moderately severe continuous hypoxia (at >4,000-m simulated altitude) with return to normoxia for the remainder of each day elicited no measurable cardiovascular sequelae, at least when measured on day 3 following the cessation of hypoxic exposure (36). However, some of the maladaptive responses to continuous (24 h) short-term hypoxic exposure, such as elevated sympathetic vasoconstrictor outflow and pulmonary vascular remodeling, reduced plasma volume, poor sleep quality, and high ventilatory drive, may conceivably impose variable, carryover consequences on return to sea level (see Sojourn to Hypoxia in the Native Lowlander above), which may negatively impact exercise performance for several days (101).
Perhaps more importantly, since both live high:train high and live high:train low hypoxic training regimens require sleeping in hypoxic environments of between 2,500 and 3,500 m there is a high probability of the occurrence of nocturnal periodic breathing with accompanying cyclical IH (14, 54, 98). Thus, although the reduced atmospheric Po2 is “constant hypoxia,” the sleeping athlete would experience varying severities of cyclical, intermittent arterial hypoxemia (see FIGURE 3). This form of IH attending periodic breathing in hypoxic environments contains many of the features of maladaptive IH in OSA. These include ROS producing cyclical HbO2 desaturation/resaturation at high frequency cycles that persist throughout sleep and over many nights, transient arousals, and repeated substantial swings in cerebral vascular resistance and blood flow and intra-thoracic pressures coinciding with apneic/hyperpneic phases of the periodic cycles (1, 30).
We acknowledge that the cardiovascular consequences of only a few days of nocturnal IH in healthy subjects appear to be short-lived once the IH is terminated (see above and FIGURE 6). However, a consistent finding across these studies was that the maladaptive effects of the nocturnal IH did carry over to at least one or more of the following days and therefore would be expected to persist during the conduct of the athlete's daily training sessions in normoxia. Accordingly, although not yet verified, available evidence predicts that these maladaptive responses to cyclical IH over time would oppose and even erode some of the key adaptive, performance-enhancing mechanisms elicited via physical training per se, namely reduced MSNA, enhanced vascular endothelial responses, reduced postjunctional alpha-adrenergic responsiveness in locomotor muscle, increased ventricular compliance, and reduced insulin resistance (40, 41, 76, 95, 112). This postulate deserves thorough investigation across the range of duration and severity of hypoxia currently used in hypoxic training regimens.
In summary, we believe it difficult to justify the continued widespread use of hypoxic training regimens without more thorough understanding of how the maladaptive effects of sustained and/or nocturnal hypoxia with cyclical IH impact both exercise performance and possibly even the cardiovascular health of the athlete. Remarkably, to date, even the placebo effects (on performance) of these hypoxic training regimens have not been adequately quantified (110), nor have blood pressure or HbO2 saturation been monitored systematically in these athletes throughout these training regimens. We would predict that titrating the severity and duration of reduced inspired Po2 in individual athletes to achieve an optimal combination of a positive erythropoietic effect without significantly counteracting maladaptive consequences to performance will present a substantial challenge!
Overall Summary
We have considered the evidence to date concerning intermittent, short-duration, and life-long hypoxic exposures and their effects on chemosensitivity, cardiovascular and respiratory physiology and health, exercise performance, sleep quality, reproduction, and the overall quality of life. The adult human's diverse, energy-consuming, biological adaptation to all forms of constant and/or intermittent hypoxia–even with relatively moderate degrees of hypoxia, especially if prolonged–stands in sharp contrast to the highly selective and efficient mechanisms available to hypoxic-tolerant species. It needs to be emphasized that substantial individual differences do exist in the balance struck between adaptive vs. maladaptive responses to hypoxia and that the Tibetan native's unique genetic endowment may prove to permit a truly unqualified biological adaptation to hypoxia. We propose that hypoxemia presents a complex, multifaceted insult to O2 transport to all organs that is poorly tolerated by most humans at substantial physiological cost.
Footnotes
We are indebted to Kurt Stenmark for FIGURE 5; to Mikael Sander for FIGURE 2A; and to Carsten Lundby, Danny Green, Kevin Shoemaker, and Marc Poulin for scientific input; and to Anthony Jacques for preparation of the manuscript.
Original research reported here was supported by National Institutes of Health and American Heart Association grants to the authors.
No conflicts of interest, financial or otherwise, are declared by the author(s).
Author contributions: J.A.D. and B.J.M. prepared figures; J.A.D. and B.J.M. drafted manuscript; J.A.D. and B.J.M. edited and revised manuscript; J.A.D. and B.J.M. approved final version of manuscript.
1Poiseuille's equation: Resistance to flow = 8ln/πr4, where l is tube length, r is the radius, and n is a viscosity coefficient.
2High sympathetic vasoconstrictor outflow has also been documented in many forms of chronic hypertension and associated with enhanced carotid chemoreceptor sensitivity. Accordingly, it has been recently proposed that denervation of the carotid chemoreceptors be used to treat select patients whose hypertension is unresponsive to pharmacological treatments (84).
References
- 1.Ainslie PN, Lucas SJ, Burgess KR. Breathing and sleep at high altitude. Respir Physiol Neurobiol 188: 233–256, 2013. [DOI] [PubMed] [Google Scholar]
- 2.Alexander JK, Grover RF. Mechanism of reduced cardiac stroke volume at high altitude. Clin Cardiol 6: 301–303, 1983. [DOI] [PubMed] [Google Scholar]
- 3.Almendros I, Wang Y, Gozal D. The polymorphic and contradictory aspects of intermittent hypoxia. Am J Physiol Lung Cell Mol Physiol 307: L129–L140, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Amann M, Pegelow DF, Jacques AJ, Dempsey JA. Inspiratory muscle work in acute hypoxia influences locomotor muscle fatigue and exercise performance of healthy humans. Am J Physiol Regul Integr Comp Physiol 293: R2036–R2045, 2007. [DOI] [PubMed] [Google Scholar]
- 5.Amann M, Proctor LT, Sebranek JJ, Pegelow DF, Dempsey JA. Opioid-mediated muscle afferents inhibit central motor drive and limit peripheral muscle fatigue development in humans. J Physiol 587: 271–283, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Amann M, Romer LM, Subudhi AW, Pegelow DF, Dempsey JA. Severity of arterial hypoxaemia affects the relative contributions of peripheral muscle fatigue to exercise performance in healthy humans. J Physiol 581: 389–403, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Arias-Stella J, Saldana M. The terminal portion of the pulmonary arterial tree in people native to high altitudes. Circulation 28: 915–925, 1963. [DOI] [PubMed] [Google Scholar]
- 8.Bartsch P, Gibbs JS. Effect of altitude on the heart and the lungs. Circulation 116: 2191–2202, 2007. [DOI] [PubMed] [Google Scholar]
- 9.Beall CM, Cavalleri GL, Deng L, Elston RC, Gao Y, Knight J, Li C, Li JC, Liang Y, McCormack M, Montgomery HE, Pan H, Robbins PA, Shianna KV, Tam SC, Tsering N, Veeramah KR, Wang W, Wangdui P, Weale ME, Xu Y, Xu Z, Yang L, Zaman MJ, Zeng C, Zhang L, Zhang X, Zhaxi P, Zheng YT. Natural selection on EPAS1 (HIF2alpha) associated with low hemoglobin concentration in Tibetan highlanders. Proc Natl Acad Sci USA 107: 11459–11464, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Beall CM, Laskowski D, Strohl KP, Soria R, Villena M, Vargas E, Alarcon AM, Gonzales C, Erzurum SC. Pulmonary nitric oxide in mountain dwellers. Nature 414: 411–412, 2001. [DOI] [PubMed] [Google Scholar]
- 11.Beall CM, Song K, Elston RC, Goldstein MC. Higher offspring survival among Tibetan women with high oxygen saturation genotypes residing at 4,000 m. Proc Natl Acad Sci USA 101: 14300–14304, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Becker HF, Jerrentrup A, Ploch T, Grote L, Penzel T, Sullivan CE, Peter JH. Effect of nasal continuous positive airway pressure treatment on blood pressure in patients with obstructive sleep apnea. Circulation 107: 68–73, 2003. [DOI] [PubMed] [Google Scholar]
- 13.Beguin PC, Joyeux-Faure M, Godin-Ribuot D, Levy P, Ribuot C. Acute intermittent hypoxia improves rat myocardium tolerance to ischemia. J Appl Physiol 99: 1064–1069, 2005. [DOI] [PubMed] [Google Scholar]
- 14.Berssenbrugge A, Dempsey J, Iber C, Skatrud J, Wilson P. Mechanisms of hypoxia-induced periodic breathing during sleep in humans. J Physiol 343: 507–526, 1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bickler PE, Buck LT. Hypoxia tolerance in reptiles, amphibians, and fishes: life with variable oxygen availability. Annu Rev Physiol 69: 145–170, 2007. [DOI] [PubMed] [Google Scholar]
- 16.Bisgard GE, Busch MA, Daristotle L, Berssenbrugge AD, Forster HV. Carotid body hypercapnia does not elicit ventilatory acclimatization in goats. Respir Physiol 65: 113–125, 1986. [DOI] [PubMed] [Google Scholar]
- 17.Brooks D, Horner RL, Kimoff RJ, Kozar LF, Render-Teixeira CL, Phillipson EA. Effect of obstructive sleep apnea versus sleep fragmentation on responses to airway occlusion. Am J Respir Crit Care Med 155: 1609–1617, 1997. [DOI] [PubMed] [Google Scholar]
- 18.Brugniaux JV, Pialoux V, Foster GE, Duggan CT, Eliasziw M, Hanly PJ, Poulin MJ. Effects of intermittent hypoxia on erythropoietin, soluble erythropoietin receptor and ventilation in humans. Eur Respir J 37: 880–887, 2011. [DOI] [PubMed] [Google Scholar]
- 19.Brutsaert TD, Parra EJ, Shriver MD, Gamboa A, Rivera-Ch M, Leon-Velarde F. Ancestry explains the blunted ventilatory response to sustained hypoxia and lower exercise ventilation of Quechua altitude natives. Am J Physiol Regul Integr Comp Physiol 289: R225–R234, 2005. [DOI] [PubMed] [Google Scholar]
- 19a.Buick FJ, Gledhill N, Froese AB, Spriet L, Meyers EC. Effect of induced erythrocythemia on aerobic work capacity. J Appl Physiol Respir Environ Exerc Physiol 48: 636–642, 1980. [DOI] [PubMed] [Google Scholar]
- 20.Burgess KR, Johnson PL, Edwards N. Central and obstructive sleep apnoea during ascent to high altitude. Respirology 9: 222–229, 2004. [DOI] [PubMed] [Google Scholar]
- 21.Calbet JA, Boushel R, Robach P, Hellsten Y, Saltin B, Lundby C. Chronic hypoxia increases arterial blood pressure and reduces adenosine and ATP induced vasodilatation in skeletal muscle in healthy humans. Acta Physiol (Oxf) 211: 574–584, 2014. [DOI] [PubMed] [Google Scholar]
- 22.Cerny FC, Dempsey JA, Reddan WG. Pulmonary gas exchange in nonnative residents of high altitude. J Clin Invest 52: 2993–2999, 1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chapman RF, Stray-Gundersen J, Levine BD. Individual variation in response to altitude training. J Appl Physiol 85: 1448–1456, 1998. [DOI] [PubMed] [Google Scholar]
- 24.Dale EA, Ben MF, Mitchell GS. Unexpected benefits of intermittent hypoxia: enhanced respiratory and nonrespiratory motor function. Physiology 29: 39–48, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dempsey JA, Amann M, Harms CA, Wetter TJ. Respiratory system limitations to performance in the healthy athlete: some answers, more questions! Dtsch Z Sportmed 63: 157–162, 2012. [Google Scholar]
- 26.Dempsey JA, Forster HV, Birnbaum ML, Reddan WG, Thoden J, Grover RF, Rankin J. Control of exercise hyperpnea under varying durations of exposure to moderate hypoxia. Respir Physiol 16: 213–231, 1972. [DOI] [PubMed] [Google Scholar]
- 27.Dempsey JA, Hanson PG, Henderson KS. Exercise-induced arterial hypoxaemia in healthy human subjects at sea level. J Physiol 355: 161–175, 1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dempsey JA, Powell FL, Bisgard GE, Blain GM, Poulin MJ, Smith CA. Role of chemoreception in cardiorespiratory acclimatization to, and deacclimatization from, hypoxia. J Appl Physiol 116: 858–866, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dempsey JA, Romer L, Rodman J, Miller J, Smith C. Consequences of exercise-induced respiratory muscle work. Respir Physiol Neurobiol 151: 242–250, 2006. [DOI] [PubMed] [Google Scholar]
- 30.Dempsey JA, Veasey SC, Morgan BJ, O'Donnell CP. Pathophysiology of sleep apnea. Physiol Rev 90: 47–112, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dempsey JA, Wagner PD. Exercise-induced arterial hypoxemia. J Appl Physiol 87: 1997–2006, 1999. [DOI] [PubMed] [Google Scholar]
- 32.Dopp JM, Philippi NR, Marcus NJ, Olson EB, Bird CE, Moran JJ, Mueller SW, Morgan BJ. Xanthine oxidase inhibition attenuates endothelial dysfunction caused by chronic intermittent hypoxia in rats. Respiration 82: 458–467, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Dorrington KL, Clar C, Young JD, Jonas M, Tansley JG, Robbins PA. Time course of the human pulmonary vascular response to 8 hours of isocapnic hypoxia. Am J Physiol Heart Circ Physiol 273: H1126–H1134, 1997. [DOI] [PubMed] [Google Scholar]
- 34.Durand F, Mucci P, Prefaut C. Evidence for an inadequate hyperventilation inducing arterial hypoxemia at submaximal exercise in all highly trained endurance athletes. Med Sci Sports Exerc 32: 926–932, 2000. [DOI] [PubMed] [Google Scholar]
- 35.Foster GE, Brugniaux JV, Pialoux V, Duggan CT, Hanly PJ, Ahmed SB, Poulin MJ. Cardiovascular and cerebrovascular responses to acute hypoxia following exposure to intermittent hypoxia in healthy humans. J Physiol 587: 3287–3299, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fu Q, Townsend NE, Shiller SM, Martini ER, Okazaki K, Shibata S, Truijens MJ, Rodriguez FA, Gore CJ, Stray-Gundersen J, Levine BD. Intermittent hypobaric hypoxia exposure does not cause sustained alterations in autonomic control of blood pressure in young athletes. Am J Physiol Regul Integr Comp Physiol 292: R1977–R1984, 2007. [DOI] [PubMed] [Google Scholar]
- 37.Fuller DD, Johnson SM, Olson EB Jr., Mitchell GS. Synaptic pathways to phrenic motoneurons are enhanced by chronic intermittent hypoxia after cervical spinal cord injury. J Neurosci 23: 2993–3000, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gilmartin GS, Lynch M, Tamisier R, Weiss JW. Chronic intermittent hypoxia in humans during 28 nights results in blood pressure elevation and increased muscle sympathetic nerve activity. Am J Physiol Heart Circ Physiol 299: H925–H931, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gore CJ, Hahn AG, Scroop GC, Watson DB, Norton KI, Wood RJ, Campbell DP, Emonson DL. Increased arterial desaturation in trained cyclists during maximal exercise at 580 m altitude. J Appl Physiol 80: 2204–2210, 1996. [DOI] [PubMed] [Google Scholar]
- 40.Green DJ, Maiorana A, O'Driscoll G, Taylor R. Effect of exercise training on endothelium-derived nitric oxide function in humans. J Physiol 561: 1–25, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Green DJ, Spence A, Halliwill JR, Cable NT, Thijssen DH. Exercise and vascular adaptation in asymptomatic humans. Exp Physiol 96: 57–70, 2011. [DOI] [PubMed] [Google Scholar]
- 42.Guyenet PG. Regulation of breathing and autonomic outflows by chemoreceptors. Compr Physiol 4: 1511–1562, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hackett PH, Roach RC, Harrison GL, Schoene RB, Mills WJ Jr. Respiratory stimulants and sleep periodic breathing at high altitude. Almitrine versus acetazolamide. Am Rev Respir Dis 135: 896–898, 1987. [DOI] [PubMed] [Google Scholar]
- 44.Hahn AG, Gore CJ. The effect of altitude on cycling performance: a challenge to traditional concepts. Sports Med 31: 533–557, 2001. [DOI] [PubMed] [Google Scholar]
- 45.Hansen J, Sander M. Sympathetic neural overactivity in healthy humans after prolonged exposure to hypobaric hypoxia. J Physiol 546: 921–929, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Harms CA, McClaran SR, Nickele GA, Pegelow DF, Nelson WB, Dempsey JA. Exercise-induced arterial hypoxaemia in healthy young women. J Physiol 507: 619–628, 1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hlastala MP, Lamm WJ, Karp A, Polissar NL, Starr IR, Glenny RW. Spatial distribution of hypoxic pulmonary vasoconstriction in the supine pig. J Appl Physiol 96: 1589–1599, 2004. [DOI] [PubMed] [Google Scholar]
- 48.Holliss BA, Fulford J, Vanhatalo A, Pedlar CR, Jones AM. Influence of intermittent hypoxic training on muscle energetics and exercise tolerance. J Appl Physiol 114: 611–619, 2013. [DOI] [PubMed] [Google Scholar]
- 49.Hsia CC, Johnson RL Jr., McDonough P, Dane DM, Hurst MD, Fehmel JL, Wagner HE, Wagner PD. Residence at 3,800-m altitude for 5 mo in growing dogs enhances lung diffusing capacity for oxygen that persists at least 25 years. J Appl Physiol 102: 1448–1455, 2007. [DOI] [PubMed] [Google Scholar]
- 50.Jean D, Moore LG. Travel to high altitude during pregnancy: frequently asked questions and recommendations for clinicians. High Alt Med Biol 13: 73–81, 2012. [DOI] [PubMed] [Google Scholar]
- 51.Johnson BD, Saupe KW, Dempsey JA. Mechanical constraints on exercise hyperpnea in endurance athletes. J Appl Physiol 73: 874–886, 1992. [DOI] [PubMed] [Google Scholar]
- 52.Kaczmarek E, Bakker JP, Clarke DN, Csizmadia E, Kocher O, Veves A, Tecilazich F, O'Donnell CP, Ferran C, Malhotra A. Molecular biomarkers of vascular dysfunction in obstructive sleep apnea. PLos One 8: e70559, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Khan Q, Heath D, Smith P. Anatomical variations in human carotid bodies. J Clin Pathol 41: 1196–1199, 1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kinsman TA, Hahn AG, Gore CJ, Wilsmore BR, Martin DT, Chow CM. Respiratory events and periodic breathing in cyclists sleeping at 2,650-m simulated altitude. J Appl Physiol 92: 2114–2118, 2002. [DOI] [PubMed] [Google Scholar]
- 55.Knight WD, Little JT, Carreno FR, Toney GM, Mifflin SW, Cunningham JT. Chronic intermittent hypoxia increases blood pressure and expression of FosB/DeltaFosB in central autonomic regions. Am J Physiol Regul Integr Comp Physiol 301: R131–R139, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.La Gerche A, MacIsaac AI, Burns AT, Mooney DJ, Inder WJ, Voigt JU, Heidbuchel H, Prior DL. Pulmonary transit of agitated contrast is associated with enhanced pulmonary vascular reserve and right ventricular function during exercise. J Appl Physiol 109: 1307–1317, 2010. [DOI] [PubMed] [Google Scholar]
- 57.Lawler J, Powers SK, Thompson D. Linear relationship between V̇o2 max and V̇o2 max decrement during exposure to acute hypoxia. J Appl Physiol 64: 1486–1492, 1988. [DOI] [PubMed] [Google Scholar]
- 58.Levine BD, Stray-Gundersen J. “Living high-training low”: effect of moderate-altitude acclimatization with low-altitude training on performance. J Appl Physiol 83: 102–112, 1997. [DOI] [PubMed] [Google Scholar]
- 59.Lewis NC, Bailey DM, Dumanoir GR, Messinger L, Lucas SJ, Cotter JD, Donnelly J, McEneny J, Young IS, Stembridge M, Burgess KR, Basnet AS, Ainslie PN. Conduit artery structure and function in lowlanders and native highlanders: relationships with oxidative stress and role of sympathoexcitation. J Physiol 592: 1009–1024, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lim DC, Brady DC, Po P, Chuang LP, Marcondes L, Kim EY, Keenan BT, Guo X, Maislin G, Galante RJ, Pack AI. Simulating obstructive sleep apnea patients' oxygenation characteristics into a mouse model of cyclical intermittent hypoxia. J Appl Physiol 118: 544–557, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Llanos AJ, Ebensperger G, Herrera EA, Reyes RV, Moraga FA, Parer JT, Giussani DA. Counterpoint: high altitude is not for the birds! J Appl Physiol 111: 1515–1518, 2011. [DOI] [PubMed] [Google Scholar]
- 62.Louis M, Punjabi NM. Effects of acute intermittent hypoxia on glucose metabolism in awake healthy volunteers. J Appl Physiol 106: 1538–1544, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lundby C, Robach P, Boushel R, Thomsen JJ, Rasmussen P, Koskolou M, Calbet JA. Does recombinant human Epo increase exercise capacity by means other than augmenting oxygen transport? J Appl Physiol 105: 581–587, 2008. [DOI] [PubMed] [Google Scholar]
- 64.Maggiorini M, Melot C, Pierre S, Pfeiffer F, Greve I, Sartori C, Lepori M, Hauser M, Scherrer U, Naeije R. High-altitude pulmonary edema is initially caused by an increase in capillary pressure. Circulation 103: 2078–2083, 2001. [DOI] [PubMed] [Google Scholar]
- 65.Mairbaurl H, Weymann J, Mohrlein A, Swenson ER, Maggiorini M, Gibbs JS, Bartsch P. Nasal epithelium potential difference at high altitude (4,559 m): evidence for secretion. Am J Respir Crit Care Med 167: 862–867, 2003. [DOI] [PubMed] [Google Scholar]
- 66.Marcus NJ, Li YL, Bird CE, Schultz HD, Morgan BJ. Chronic intermittent hypoxia augments chemoreflex control of sympathetic activity: role of the angiotensin II type 1 receptor. Respir Physiol Neurobiol 171: 36–45, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mateika JH, El-Chami M, Shaheen D, Ivers B. Intermittent hypoxia: a low risk research tool with therapeutic value in humans. J Appl Physiol 118: 520–532, 2014. [DOI] [PubMed] [Google Scholar]
- 68.McFadden DM, Houston CS, Sutton JR, Powles AC, Gray GW, Roberts RS. High-altitude retinopathy. JAMA 245: 581–586, 1981. [PubMed] [Google Scholar]
- 69.Messner R. Everest: Expedition to the Ultimate. London: Mountaineers Books, 1979. [Google Scholar]
- 70.Miller JD, Hemauer SJ, Smith CA, Stickland MK, Dempsey JA. Expiratory threshold loading impairs cardiovascular function in health and chronic heart failure during submaximal exercise. J Appl Physiol 101: 213–227, 2006. [DOI] [PubMed] [Google Scholar]
- 71.Moore LG, Charles SM, Julian CG. Humans at high altitude: hypoxia and fetal growth. Respir Physiol Neurobiol 178: 181–190, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Moore LG, Zamudio S, Zhuang J, Sun S, Droma T. Oxygen transport in tibetan women during pregnancy at 3,658 m. Am J Phys Anthropol 114: 42–53, 2001. [DOI] [PubMed] [Google Scholar]
- 73.Morgan BJ, Adrian R, Bates ML, Dopp JM, Dempsey JA. Quantifying hypoxia-induced chemoreceptor sensitivity in the awake rodent. J Appl Physiol 117: 816–824, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Morgan BJ, Crabtree DC, Palta M, Skatrud JB. Combined hypoxia and hypercapnia evokes long-lasting sympathetic activation in humans. J Appl Physiol 79: 205–213, 1995. [DOI] [PubMed] [Google Scholar]
- 75.Morrison SF. 2010 Carl Ludwig Distinguished Lectureship of the APS Neural Control and Autonomic Regulation Section: Central neural pathways for thermoregulatory cold defense. J Appl Physiol 110: 1137–1149, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mortensen SP, Nyberg M, Gliemann L, Thaning P, Saltin B, Hellsten Y. Exercise training modulates functional sympatholysis and alpha-adrenergic vasoconstrictor responsiveness in hypertensive and normotensive individuals. J Physiol 592: 3063–3073, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mortola JP, Rezzonico R, Lanthier C. Ventilation and oxygen consumption during acute hypoxia in newborn mammals: a comparative analysis. Respir Physiol 78: 31–43, 1989. [DOI] [PubMed] [Google Scholar]
- 78.Naeije R, Dedobbeleer C. Pulmonary hypertension and the right ventricle in hypoxia. Exp Physiol 98: 1247–1256, 2013. [DOI] [PubMed] [Google Scholar]
- 79.Naeije R, Huez S, Lamotte M, Retailleau K, Neupane S, Abramowicz D, Faoro V. Pulmonary artery pressure limits exercise capacity at high altitude. Eur Respir J 36: 1049–1055, 2010. [DOI] [PubMed] [Google Scholar]
- 80.Naeije R, Vanderpool R, Dhakal BP, Saggar R, Saggar R, Vachiery JL, Lewis GD. Exercise-induced pulmonary hypertension: physiological basis and methodological concerns. Am J Respir Crit Care Med 187: 576–583, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Naghshin J, McGaffin KR, Witham WG, Mathier MA, Romano LC, Smith SH, Janczewski AM, Kirk JA, Shroff SG, O'Donnell CP. Chronic intermittent hypoxia increases left ventricular contractility in C57BL/6J mice. J Appl Physiol 107: 787–793, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Naghshin J, Rodriguez RH, Davis EM, Romano LC, McGaffin KR, O'Donnell CP. Chronic intermittent hypoxia exposure improves left ventricular contractility in transgenic mice with heart failure. J Appl Physiol 113: 791–798, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Oliver SJ, Macdonald JH, Harper Smith AD, Lawley JS, Gallagher CA, Di FU, Walsh NP. High altitude impairs in vivo immunity in humans. High Alt Med Biol 14: 144–149, 2013. [DOI] [PubMed] [Google Scholar]
- 84.Paton JF, Sobotka PA, Fudim M, Engelman ZJ, Hart EC, McBryde FD, Abdala AP, Marina N, Gourine AV, Lobo M, Patel N, Burchell A, Ratcliffe L, Nightingale A. The carotid body as a therapeutic target for the treatment of sympathetically mediated diseases. Hypertension 61: 5–13, 2013. [DOI] [PubMed] [Google Scholar]
- 85.Penaloza D, Arias-Stella J. The heart and pulmonary circulation at high altitudes: healthy highlanders and chronic mountain sickness. Circulation 115: 1132–1146, 2007. [DOI] [PubMed] [Google Scholar]
- 86.Penaloza D, Sime F. Chronic cor pulmonale due to loss of altitude acclimatization (chronic mountain sickness). Am J Med 50: 728–743, 1971. [DOI] [PubMed] [Google Scholar]
- 87.Peng YJ, Yuan G, Khan S, Nanduri J, Makarenko VV, Reddy VD, Vasavda C, Kumar GK, Semenza GL, Prabhakar NR. Regulation of hypoxia-inducible factor-alpha isoforms and redox state by carotid body neural activity in rats. J Physiol 592: 3841–3858, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Peppard PE, Young T, Palta M, Dempsey J, Skatrud J. Longitudinal study of moderate weight change and sleep-disordered breathing. JAMA 284: 3015–3021, 2000. [DOI] [PubMed] [Google Scholar]
- 89.Phillips SA, Olson EB, Morgan BJ, Lombard JH. Chronic intermittent hypoxia impairs endothelium-dependent dilation in rat cerebral and skeletal muscle resistance arteries. Am J Physiol Heart Circ Physiol 286: H388–H393, 2004. [DOI] [PubMed] [Google Scholar]
- 90.Pialoux V, Foster GE, Ahmed SB, Beaudin AE, Hanly PJ, Poulin MJ. Losartan abolishes oxidative stress induced by intermittent hypoxia in humans. J Physiol 589: 5529–5537, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Pialoux V, Hanly PJ, Foster GE, Brugniaux JV, Beaudin AE, Hartmann SE, Pun M, Duggan CT, Poulin MJ. Effects of exposure to intermittent hypoxia on oxidative stress and acute hypoxic ventilatory response in humans. Am J Respir Crit Care Med 180: 1002–1009, 2009. [DOI] [PubMed] [Google Scholar]
- 92.Prabhakar NR, Peng YJ, Kumar GK, Nanduri J. Peripheral chemoreception and arterial pressure responses to intermittent hypoxia. Compr Physiol 5: 561–577, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Prabhakar NR, Semenza GL. Adaptive and maladaptive cardiorespiratory responses to continuous and intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2. Physiol Rev 92: 967–1003, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Prabhakar NR, Semenza GL. Adaptive and maladaptive cardiorespiratory responses to continuous and intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2. Physiol Rev 92: 967–1003, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Ray CA. Sympathetic adaptations to one-legged training. J Appl Physiol 86: 1583–1587, 1999. [DOI] [PubMed] [Google Scholar]
- 96.Reeves JT, Groves BM, Sutton JR, Wagner PD, Cymerman A, Malconian MK, Rock PB, Young PM, Houston CS. Operation Everest II: preservation of cardiac function at extreme altitude. J Appl Physiol 63: 531–539, 1987. [DOI] [PubMed] [Google Scholar]
- 97.Reichmuth KJ, Dopp JM, Barczi SR, Skatrud JB, Wojdyla P, Hayes D Jr., Morgan BJ. Impaired vascular regulation in patients with obstructive sleep apnea: effects of continuous positive airway pressure treatment. Am J Respir Crit Care Med 180: 1143–1150, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Reite M, Jackson D, Cahoon RL, Weil JV. Sleep physiology at high altitude. Electroencephalogr Clin Neurophysiol 38: 463–471, 1975. [DOI] [PubMed] [Google Scholar]
- 99.Roels B, Bentley DJ, Coste O, Mercier J, Millet GP. Effects of intermittent hypoxic training on cycling performance in well-trained athletes. Eur J Appl Physiol 101: 359–368, 2007. [DOI] [PubMed] [Google Scholar]
- 100.Sadoshima J. Redox regulation of growth and death in cardiac myocytes. Antioxid Redox Signal 8: 1621–1624, 2006. [DOI] [PubMed] [Google Scholar]
- 101.Saunders PU, Pyne DB, Gore CJ. Endurance training at altitude. High Alt Med Biol 10: 135–148, 2009. [DOI] [PubMed] [Google Scholar]
- 102.Schultz HD, Li YL. Carotid body function in heart failure. Respir Physiol Neurobiol 157: 171–185, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Schwab M, Jayet PY, Stuber T, Salinas CE, Bloch J, Spielvogel H, Villena M, Allemann Y, Sartori C, Scherrer U. Pulmonary-artery pressure and exhaled nitric oxide in Bolivian and Caucasian high altitude dwellers. High Alt Med Biol 9: 295–299, 2008. [DOI] [PubMed] [Google Scholar]
- 104.Scott GR, Hawkes LA, Frappell PB, Butler PJ, Bishop CM, Milsom WK. How bar-headed geese fly over the Himalayas. Physiology (Bethesda) 30: 107–115, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Scott GR, Meir JU, Hawkes LA, Frappell PB, Milsom WK. Point: high altitude is for the birds! J Appl Physiol 111: 1514–1515, 2011. [DOI] [PubMed] [Google Scholar]
- 106.Semenza GL, Prabhakar NR. Neural regulation of hypoxia-inducible factors and redox state drives the pathogenesis of hypertension in a rodent model of sleep apnea. J Appl Physiol. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Serebrovskaya TV, Nikolsky IS, Nikolska VV, Mallet RT, Ishchuk VA. Intermittent hypoxia mobilizes hematopoietic progenitors and augments cellular and humoral elements of innate immunity in adult men. High Alt Med Biol 12: 243–252, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Sharpe AL, Calderon AS, Andrade MA, Cunningham JT, Mifflin SW, Toney GM. Chronic intermittent hypoxia increases sympathetic control of blood pressure: role of neuronal activity in the hypothalamic paraventricular nucleus. Am J Physiol Heart Circ Physiol 305: H1772–H1780, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Siebenmann C, Cathomen A, Hug M, Keiser S, Lundby AK, Hilty MP, Goetze JP, Rasmussen P, Lundby C. Hemoglobin mass and intravascular volume kinetics during and after exposure to 3454 m altitude. J Appl Physiol. In press. [DOI] [PubMed] [Google Scholar]
- 110.Siebenmann C, Robach P, Jacobs RA, Rasmussen P, Nordsborg N, Diaz V, Christ A, Olsen NV, Maggiorini M, Lundby C. “Live high-train low” using normobaric hypoxia: a double-blinded, placebo-controlled study. J Appl Physiol 112: 106–117, 2012. [DOI] [PubMed] [Google Scholar]
- 111.Simonson TS, Yang Y, Huff CD, Yun H, Qin G, Witherspoon DJ, Bai Z, Lorenzo FR, Xing J, Jorde LB, Prchal JT, Ge R. Genetic evidence for high-altitude adaptation in Tibet. Science 329: 72–75, 2010. [DOI] [PubMed] [Google Scholar]
- 112.Sinoway L, Shenberger J, Leaman G, Zelis R, Gray K, Baily R, Leuenberger U. Forearm training attenuates sympathetic responses to prolonged rhythmic forearm exercise. J Appl Physiol 81: 1778–1784, 1996. [DOI] [PubMed] [Google Scholar]
- 113.Stuber T, Sartori C, Schwab M, Jayet PY, Rimoldi SF, Garcin S, Thalmann S, Spielvogel H, Salmon CS, Villena M, Scherrer U, Allemann Y. Exaggerated pulmonary hypertension during mild exercise in chronic mountain sickness. Chest 137: 388–392, 2010. [DOI] [PubMed] [Google Scholar]
- 114.Stuber T, Scherrer U. Circulatory adaptation to long-term high altitude exposure in Aymaras and Caucasians. Prog Cardiovasc Dis 52: 534–539, 2010. [DOI] [PubMed] [Google Scholar]
- 115.Tamisier R, Gilmartin GS, Launois SH, Pepin JL, Nespoulet H, Thomas R, Levy P, Weiss JW. A new model of chronic intermittent hypoxia in humans: effect on ventilation, sleep, and blood pressure. J Appl Physiol 107: 17–24, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Tamisier R, Pepin JL, Remy J, Baguet JP, Taylor JA, Weiss JW, Levy P. 14 nights of intermittent hypoxia elevate daytime blood pressure and sympathetic activity in healthy humans. Eur Respir J 37: 119–128, 2011. [DOI] [PubMed] [Google Scholar]
- 117.Tester NJ, Fuller DD, Fromm JS, Spiess MR, Behrman AL, Mateika JH. Long-term facilitation of ventilation in humans with chronic spinal cord injury. Am J Respir Crit Care Med 189: 57–65, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Thoden JS, Dempsey JA, Reddan WG, Birnbaum ML, Forster HV, Grover RF, Rankin J. Ventilatory work during steady-state response to exercise. Fed Proc 28: 1316–1321, 1969. [PubMed] [Google Scholar]
- 119.Thomson JM, Stone JA, Ginsburg AD, Hamilton P. O2 transport during exercise following blood reinfusion. J Appl Physiol Respir Environ Exercise Physiol 53: 1213–1219, 1982. [DOI] [PubMed] [Google Scholar]
- 120.Vogel JH, Weaver WF, Rose RL, Blount SG Jr., Grover RF. Pulmonary hypertension on exertion in normal man living at 10,150 feet (Leadville, Colorado). Med Thorac 19: 461–477, 1962. [DOI] [PubMed] [Google Scholar]
- 121.Wang ZY, Olson EB Jr., Bjorling DE, Mitchell GSand Bisgard GE. Sustained hypoxia-induced proliferation of carotid body type I cells in rats. J Appl Physiol 104: 803–808, 2008. [DOI] [PubMed] [Google Scholar]
- 122.Weinberger B, Laskin DL, Heck DE, Laskin JD. Oxygen toxicity in premature infants. Toxicol Appl Pharmacol 181: 60–67, 2002. [DOI] [PubMed] [Google Scholar]
- 123.Wetter TJ, Xiang Z, Sonetti DA, Haverkamp HC, Rice AJ, Abbasi AA, Meyer KC, Dempsey JA. Role of lung inflammatory mediators as a cause of exercise-induced arterial hypoxemia in young athletes. J Appl Physiol 93: 116–126, 2002. [DOI] [PubMed] [Google Scholar]
- 124.White DP, Gleeson K, Pickett CK, Rannels AM, Cymerman A, Weil JV. Altitude acclimatization: influence on periodic breathing and chemoresponsiveness during sleep. J Appl Physiol 63: 401–412, 1987. [DOI] [PubMed] [Google Scholar]
- 125.Wilson MH, Newman S, Imray CH. The cerebral effects of ascent to high altitudes. Lancet Neurol 8: 175–191, 2009. [DOI] [PubMed] [Google Scholar]
- 126.Wolfel EE, Groves BM, Brooks GA, Butterfield GE, Mazzeo RS, Moore LG, Sutton JR, Bender PR, Dahms TE, McCullough RE. Oxygen transport during steady-state submaximal exercise in chronic hypoxia. J Appl Physiol 70: 1129–1136, 1991. [DOI] [PubMed] [Google Scholar]
- 127.Wolfel EE, Levine BD. The cardiovascular system at high altitude: heart and systemic circulation. In:High Altitude edited by Hornbein TF, Schoene RB. New York: Marcel Dekker, 2001. p. 235–292. [Google Scholar]
- 128.Wolfel EE, Selland MA, Mazzeo RS, Reeves JT. Systemic hypertension at 4,300 m is related to sympathoadrenal activity. J Appl Physiol 76: 1643–1650, 1994. [DOI] [PubMed] [Google Scholar]
- 129.Xie A, Skatrud JB, Crabtree DC, Puleo DS, Goodman BM, Morgan BJ. Neurocirculatory consequences of intermittent asphyxia in humans. J Appl Physiol 89: 1333–1339, 2000. [DOI] [PubMed] [Google Scholar]
- 130.Xie A, Skatrud JB, Dempsey JA. Effect of hypoxia on the hypopnoeic and apnoeic threshold for CO2 in sleeping humans. J Physiol 535: 269–278, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Xie A, Skatrud JB, Puleo DS, Morgan BJ. Exposure to hypoxia produces long-lasting sympathetic activation in humans. J Appl Physiol 91: 1555–1562, 2001. [DOI] [PubMed] [Google Scholar]
- 132.Yi X, Liang Y, Huerta-Sanchez E, Jin X, Cuo ZX, Pool JE, Xu X, Jiang H, Vinckenbosch N, Korneliussen TS, Zheng H, Liu T, He W, Li K, Luo R, Nie X, Wu H, Zhao M, Cao H, Zou J, Shan Y, Li S, Yang Q, Asan Ni P, Tian G, Xu J, Liu X, Jiang T, Wu R, Zhou G, Tang M, Qin J, Wang T, Feng S, Li G, Huasang Luosang J, Wang W, Chen F, Wang Y, Zheng X, Li Z, Bianba Z, Yang G, Wang X, Tang S, Gao G, Chen Y, Luo Z, Gusang L, Cao Z, Zhang Q, Ouyang W, Ren X, Liang H, Zheng H, Huang Y, Li J, Bolund L, Kristiansen K, Li Y, Zhang Y, Zhang X, Li R, Li S, Yang H, Nielsen R, Wang J, Wang J. Sequencing of 50 human exomes reveals adaptation to high altitude. Science 329: 75–78, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Yuan G, Nanduri J, Khan S, Semenza GL, Prabhakar NR. Induction of HIF-1alpha expression by intermittent hypoxia: involvement of NADPH oxidase, Ca2+ signaling, prolyl hydroxylases, and mTOR. J Cell Physiol 217: 674–685, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]