

Abstract
AIM: To investigate the molecular mechanisms of berberine inhibition of hepatic gluconeogenesis in a diabetic rat model.
METHODS: The 40 rats were randomly divided into five groups. One group was selected as the normal group. In the remaining groups (n = 8 each), the rats were fed on a high-fat diet for 1 mo and received intravenous injection of streptozotocin for induction of the diabetic models. Berberine (156 mg/kg per day) (berberine group) or metformin (184 mg/kg per day) (metformin group) was intragastrically administered to the diabetic rats and 5-aminoimidazole-4-carboxamide1-β-D-ribofuranoside (AICAR) (0.5 mg/kg per day) (AICAR group) was subcutaneously injected to the diabetic rats for 12 wk. The remaining eight diabetic rats served as the model group. Fasting plasma glucose and insulin levels as well as lipid profile were tested. The expressions of proteins were examined by western blotting. The nuclear translocation of CREB-regulated transcription co-activator (TORC)2 was observed by immunohistochemical staining.
RESULTS: Berberine improved impaired glucose tolerance and decreased plasma hyperlipidemia. Moreover, berberine decreased fasting plasma insulin and homeostasis model assessment of insulin resistance (HOMA-IR). Berberine upregulated protein expression of liver kinase (LK)B1, AMP-activated protein kinase (AMPK) and phosphorylated AMPK (p-AMPK). The level of phophorylated TORC2 (p-TORC2) protein in the cytoplasm was higher in the berberine group than in the model group, and no significant difference in total TORC2 protein level was observed. Immunohistochemical staining revealed that more TORC2 was localized in the cytoplasm of the berberine group than in the model group. Moreover, berberine treatment downregulated protein expression of the key gluconeogenic enzymes (phosphoenolpyruvate carboxykinase and glucose-6-phosphatase) in the liver tissues.
CONCLUSION: Our findings revealed that berberine inhibited hepatic gluconeogenesis via the regulation of the LKB1-AMPK-TORC2 signaling pathway.
Keywords: Berberine, Diabetes, AMPK, LKB1, Hepatic gluconeogenesis, TORC2
Core tip: We showed that liver kinase (LK)B1 acts as the upstream regulator of AMP-activated protein kinase (AMPK) and participates in gluconeogenesis. AMPK phosphorylation triggers CREB-regulated transcription co-activator (TORC)2 phosphorylation, which results in the inhibition of the nuclear translocation of TORC2. Thus, gluconeogenesis is restrained. No previous studies have reported the molecular mechanisms of berberine reducing hyperglycemia via the inhibition of hepatic gluconeogenesis. We found that berberine upregulated protein expression of LKB1, AMPK, p-AMPK and p-TORC2. Moreover, we observed that berberine inhibited the translocation of TOCR2 into the cell nucleus.
INTRODUCTION
The liver plays a crucial role in the maintenance of systemic glucose homeostasis. In the absorptive state, the liver increases glucose uptake via the absorption of glucose by hepatocytes and subsequent transformation into glycogen and lipids. In the fasting state, hepatocytes provide glucose via glycogenolysis and gluconeogenesis to maintain glucose homeostasis. However, abnormal hepatic gluconeogenesis results in the elevation of glucose levels. Gluconeogenesis in the liver is regulated through the transcriptional modulation of gluconeogenic enzymes such as glucose-6-phosphatase (G-6-Pase) and phosphoenolpyruvate carboxykinase (PEPCK)[1].
AMP-activated protein kinase (AMPK) plays a vital role in gluconeogenesis in the liver. AMPK is a conserved sensor and regulator of cellar energy balance that is activated when the cellular AMP: ATP ratio exhibits a large increase due to conditions of nutrient deprivation or pathological stress[2]. Liver kinase (LK)B1 is a serine/threonine protein kinase that was originally identified as a tumor suppressor gene. The LKB1 mutation is responsible for the familiar Peutz-Jeghers syndrome[3]. The deletion of hepatic LKB1 in adult mice results in the nearly complete loss of AMPK activity, which in turn, results in hyperglycemia due to increased gluconeogenic gene expression[4]. Previous research has indicated that LKB1 acts as the upstream regulator of AMPK and participates in gluconeogenesis. Koo et al[5] illustrated that CREB-regulated transcription co-activator (TORC)2 is a key regulator of glucose output that acts through the cAMP responsive element binding protein (CREB) and found that TORC2-deficient mice exhibit fasting hypoglycemia. Subsequently, CREB stimulates hepatic gluconeogenesis to drive the expression of the nuclear receptor coactivator peroxisome proliferator-activated receptor co-activator (PGC)-1α[4,5]. PGC-1α is a transcriptional coactivator of nuclear receptors and plays a vital role in activating the expression of the genes for key gluconeogenic enzymes such as PEPCK and G-6-P[6,7]. The research of Koo et al[5] showed that AMPK phosphorylation due to ATP depletion triggers TORC2 phosphorylation, which results in the inhibition of the nuclear translocation of TORC2; in turn, the cytoplasmic localization of TORC2 prevents its combination with CREB elements. Thus, gluconeogenesis is restrained. In the future, the LKB1-AMPK-TORC2 signaling pathway will probably be a target for the treatment of type 2 diabetes.
Berberine is an isoquinoline alkaloid extracted from Rhizoma Coptidis. The hypoglycemic effect of berberine was first identified in 1988 via the treatment of diarrhea in diabetic patients[8]. Since that time, many studies about the influence of berberine on hyperglycemia-reducing and insulin resistance-improving have been reported. Recently, berberine was proven to be capable of reducing hyperglycemia via the inhibition of hepatic gluconeogenesis[1,9,10]. Based on the inhibition of gluconeogenesis by the LKB1-AMPK-TORC2 signaling pathway, we hypothesized that berberine reduces hyperglycemia via the LKB1-AMPK-TORC2 signaling pathway to control gluconeogenesis.
MATERIALS AND METHODS
Animal care and use statement
Male Wistar rats, weighing 160 g, supplied by the Centers for Disease Control and Prevention (Wuhan, China) were fed adaptively for 1 wk in an ambient temperature of 22 ± 1 °C and on a 12-h light/dark cycle with free access to water and the standard rat diet (containing 35% flour, 20% soy meal, 20% corn meal, 15.5% bran, 0.5% bean oil, 5% fish meal, 2.5% bone meal, 1% dusty yeast, and 0.5% salt). All experimental procedures were performed in accordance with the guide principle for experimental animals (MSTPRC Directive of 1988, No. 88-2).
Chemicals and experimental drugs
Streptozotocin (STZ) was produce by Sigma (St Louis, MO, United States). The assay kits used for blood lipid determinations were purchased from Jiancheng Bio-engineering Institute (Nanjing, China). Berberine was provided by the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China). Metformin was purchased from Shenzhen Vanda Pharmaceuticals (Shenzhen, China), and AICAR was procured from the Beyotime Institute of Biotechnology (Jiangsu, China).
Experimental design
The rats were randomly assigned to a normal control group that received the standard rat diet (normal) or the remaining four groups that received a high-fat diet (containing 67.5% standard laboratory rat chow, 15% lard, 15% sugar, 2% cholesterol, and 0.5% bile salts) for 4 wk. Next, the rats received tail vein injections of STZ (30 mg/kg) dissolved in 0.05mol/L sodium citrate (pH 4.5) after 12-h fast for induction of the diabetic models[11]. One week later, oral glucose tolerance test (OGTT) was performed. The 95% confidence intervals were calculated based on the plasma glucose levels of normal rats. The rats with diabetes (i.e., rats with plasma glucose levels that were above the normal upper limit at two time points or 20% greater than the normal upper limit at one time point) were selected. Next, the diabetic rats were randomized into the following four groups (n = 8 per group): an untreated diabetic group (model), a berberine-treated group (berberine), a metformin-treated group (metformin) and an AICAR-treated group (AICAR). Berberine (156 mg/kg per day) and metformin (184 mg/kg per day) were dissolved in sodium carboxymethylcellulose and intragastrically administered to the rats daily for 12 wk. AICAR (0.5 mg/kg per day) was dissolved in normal saline, and the rats in the AICAR-treated group were given daily subcutaneous injections of AICAR for 12 wk. The doses were adjusted according to the body weight, which was recorded once per week. The day before the rats were sacrificed, the rats were anesthetized with diethyl ether after fasting for 12 h, and orbital venous blood was obtained. Next, the rats were given glucose by gavage (2 g/kg), and additional blood samples were collected at regular intervals (t = 60 and 120 min) for glucose and insulin measurements. The rats were deeply anesthetized with pentobarbital in the fasting (12-h) condition. Blood samples were collected from the abdominal aorta and allowed to clot for 30 min at 4 °C. After centrifuging at 3000 r/min for 15 min at 4 °C, the serum was separated and stored at -80 °C until examination. The liver was removed and flushed with saline. Next the liver was collected and stored at -80 °C until use.
OGTT and fasting insulin
Blood glucose levels were examined with the glucose oxidase method using a glucose monitor (LifeScan Milpitas, CA, United States). Serum fasting insulin concentrations were measured with radioimmunoassay.
Analysis of blood lipids
The serum total cholesterol (TC), triglycerides (TG), low-density lipoprotein cholesterol (LDL-C) and high-density lipoprotein cholesterol (HDL-C) concentrations were estimated via the oxidase method using commercial reagents.
Western blot analysis
Liver total protein was extracted, and the concentrations of total protein were measured by the BCA method. The liver extractions (100 μg) were mixed with sample buffer (25 μg), boiled for 10 min, and separated on 10% SDS-PAGE. The separated proteins were electrophoretically transferred to nitrocellulose membranes. The membranes were blocked with 5% nonfat dry milk dissolved in phosphate-buffered saline with Tween-20 (PBST) or 0.5% bovine serum albumin for 2 h at room temperature. The membranes were then washed in PBST and incubated overnight with primary antibodies (LKB1, AMPK, p-AMPK, TORC2, p-TORC2, G-6-P, PEPCK, and β-actin) at 4 °C. After three washes in PBST, the membranes were incubated with the Dylight 800-labeled antibody to rabbit IgG (KPL, Hongkong, China) for 2 h. Immunoreactive proteins were visualized with a near-infrared double color laser imaging system (Odyssey, Lincoln, NE, United States). Quantity one 4.6.2 was used for assaying the protein quantification.
Immunohistochemical staining for TORC2
The liver tissues were fixed with 4% paraformaldehyde for paraffin embedding. The paraffin-embedded sections were subjected to immunohistochemical staining for TORC2 in the liver. The tissue sections were incubated with rabbit anti-TORC2 primary antibody (1:50). After washing with PBST, the sections were incubated with secondary antibody, and the diaminobenzidine method was used. Next, the TORC2 protein expressions were observed under an optical microscope.
Statistical analysis
The data are presented as the means ± SD and were assayed with SPSS version 19.0 statistical software. All experience data were analyzed with one-way analysis of variance (ANOVA). Data with equal variances were evaluated with Tukey’s test. A P value below 0.05 was considered significant. The statistical methods of the study were reviewed by Sheng Wei from the school of Public Health of Tongji Medical College.
RESULTS
Effect of berberine on glucose tolerance in type 2 diabetic rats
As shown in Figure 1A, the plasma glucose levels in the model group were significantly higher than those in the normal control group at 0, 1 and 2 h (P < 0.01). Glucose tolerances were improved in the berberine, AICAR and metformin groups compared to the model group (P < 0.01). In the berberine, AICAR and metformin groups, the areas under the curves (AUCs) constructed from the plasma glucose levels at the three time points were decreased by 52%, 64% and 62%, respectively, compared to the model group (Figure 1B).
Figure 1.
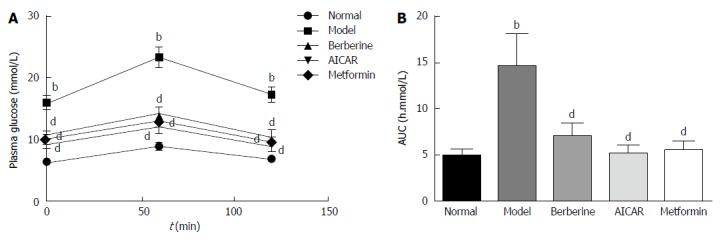
Effects of berberine on plasma glucose levels in the oral glucose tolerance test and the areas under the curves for plasma glucose. bP < 0.01 vs the normal control group at the corresponding time point; dP < 0.01 vs the model group at the corresponding time point (by ANOVA). AUC: Areas under the curve.
Effect of berberine on fasting plasma insulin and insulin resistance index
Blood insulin was monitored to assay pancreatic beta cell function. As shown in Figure 2, fasting insulin level was significantly higher in the model group than in the normal control group (P < 0.01), and berberine significantly lowered fasting insulin level compared to the model group (P < 0.01) (Figure 2A). Moreover, the fasting plasma insulin and homeostasis model assessment of insulin resistance (HOMA-IR) in the model group was higher than in the normal control group (P < 0.01), and berberine notably decreased HOMA-IR compared to the model group (P < 0.01) (Figure 2B).
Figure 2.
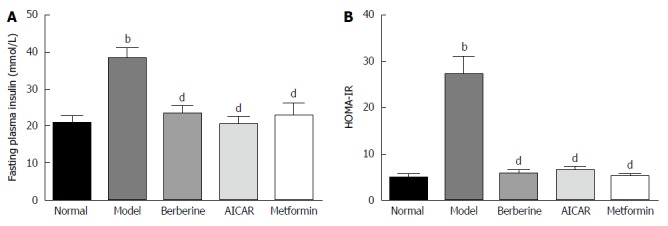
Effects of berberine on fasting plasma insulin level and homeostasis model assessment of insulin resistance in diabetic rats. Each bar represents the mean ± SD (n = 8). bP < 0.01 vs the normal control group; dP < 0.01 vs the model group (by ANOVA). HOMA-IR: Homeostasis model assessment of insulin resistance.
Berberine improved hyperlipidemia in type 2 diabetic rats
As shown in Table 1, the model rats exhibited severe dyslipidemia. The serum TG, TC, and LDL-C levels were higher in the model group than in the normal control group (P < 0.01). Treatments with berberine, AICAR and metformin markedly ameliorated the increases in the TG, TC and LDL-C levels in the diabetic rats compared to the model rats (P < 0.01). The HDL-C levels of the model group were lower than those of the normal control group, and the HDL-C levels of the treatment groups were increased compared to those of the model rats (P < 0.01).
Table 1.
Effects of berberine on plasma lipid profiles of diabetic rats (mean ± SD, n = 8)
| Group | TG (mmol/L) | TC (mmol/L) | LDL-C (mmol/L) | HDL-C (mmol/L) |
| Normal | 0.98 ± 0.15 | 3.74 ± 0.56 | 1.48 ± 0.18 | 2.68 ± 0.48 |
| Model | 2.7 ± 0.57b | 6.66 ± 1.14b | 4.26 ± 0.63b | 1.14 ± 0.15b |
| Berberine | 1.44 ± 0.23d | 4.88 ± 0.96d | 1.46 ± 0.32d | 2.12 ± 0.63d |
| AICAR | 1.28 ± 0.31d | 4.54 ± 0.55d | 1.82 ± 0.22d | 2.34 ± 0.40d |
| Metformin | 1.26 ± 0.37d | 4.72 ± 0.56d | 1.60 ± 0.27d | 2.52 ± 0.59d |
P < 0.01 vs the normal control group,
P < 0.01 vs the model group (by ANOVA). TC: Total cholesterol; TG: Triglycerides; LDL-C: Low-density lipoprotein cholesterol; HDL-C: High-density lipoprotein cholesterol.
Berberine regulated expression of LKB1 protein in livers of type 2 diabetic rats
As shown in Figure 3, the expression of LKB1 protein in the model rats decreased compared to the normal control group (P < 0.05). However, treatments of berberine, AICAR and metformin increased the expression of LKB1 protein compared to the model rats (P < 0.05).
Figure 3.
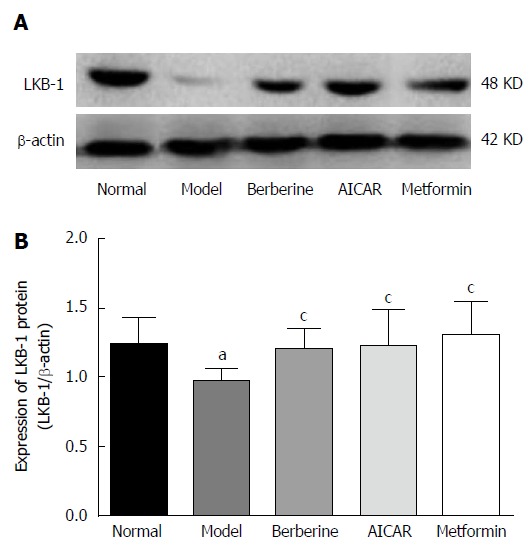
Effect of berberine on hepatic LKB-1 protein expression. Western blot analyses of LKB-1 levels in liver tissues of normal control rats, model rats and diabetic rats treated with berberine, AICAR and metformin. A: Representative blots for each group are shown; B: Each bar is expressed as LKB-1/β-actin and represents the mean ± SD (n = 8). aP < 0.05 vs the normal control group; cP < 0.05 vs the model group (by ANOVA).
Berberine regulated expression of AMPK and p-AMPK proteins in the livers of type 2 diabetic rats
AMPK is an energy sensor, and phosphorylation of AMPK is increased when it is activated. As shown in Figure 4, the liver AMPK and P-AMPK protein levels were lower in the model group than in the normal control group, and berberine, AICAR and metformin treatments considerably increased the expressions of AMPK and P-AMPK proteins compared to the model rats (P < 0.01).
Figure 4.
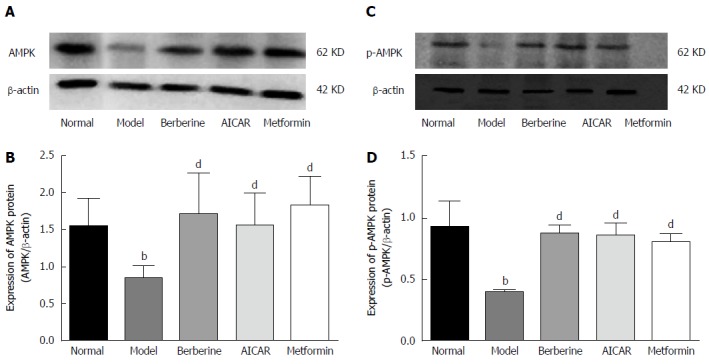
Effect of berberine on hepatic AMPK and p-AMPK protein expression. Western blot analyses of AMPK and p-AMPK protein in liver tissues of normal control rats, model rats and diabetic rats treated with berberine, AICAR or metformin. A, C: Representative blots for each group are shown; B: Each bar is expressed as AMPKD/β-actin and represents the mean ± SD (n = 8); D: Each bar is expressed as p-AMPK/β-actin and represents the mean ± SD (n = 8). bP < 0.01 vs the normal control group; dP < 0.01 vs the model group (by ANOVA).
Berberine regulated TORC2 nuclear translocation in livers of type 2 diabetic rats
When TORC2 is phosphorylated in the liver, it is located in the cytoplasm and gluconeogenesis does not occur. As shown in Figure 5, the p-TORC2 levels of the model group was lower than that of the normal control group (P < 0.01), and the p-TORC2 levels were significantly increased in groups treated with berberine, AICAR or metformin compared to the model group (P < 0.01). However, there was no significant difference in the expression of total TORC2 protein across the five groups (P > 0.01). As shown in Figure 6, we also verified that berberine inhibited TORC2 nuclear translocation in the liver tissues via immunohistochemical staining. The nuclear expression of TORC2 protein was obviously increased in the model group compared to the normal group; however, the treatments with berberine, AICAR and metformin inhibited the nuclear translocation of the TORC2 protein.
Figure 5.
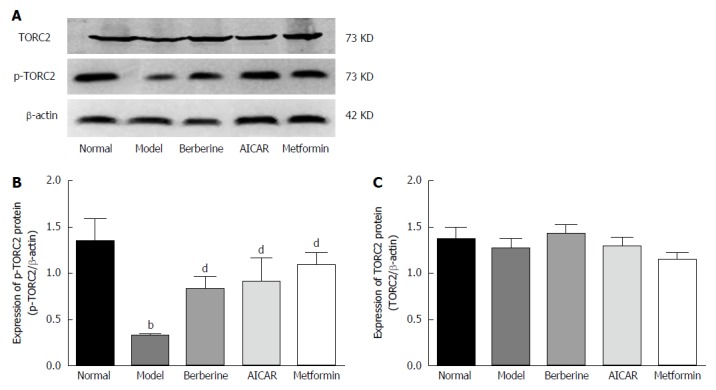
Effect of berberine on hepatic p-TORC2 and total TORC2 protein expression. Western blot analyses of p-TORC2 and TORC2 proteins from liver tissues of normal rats, model rats and diabetic rats treated with Berberine, AICAR and Metformin. A: Representative blots for each group are shown; B: Each bar is expressed as p-TORC2/β-actin and represents the mean ± SD (n = 8); C: Each bar is expressed as total TORC2/β-actin and represents the mean ± SD (n = 8). bP < 0.01 vs the normal group; dP < 0.01 vs the model group (by ANOVA).There was no significant difference in the expression of total TORC2 protein across the five groups.
Figure 6.
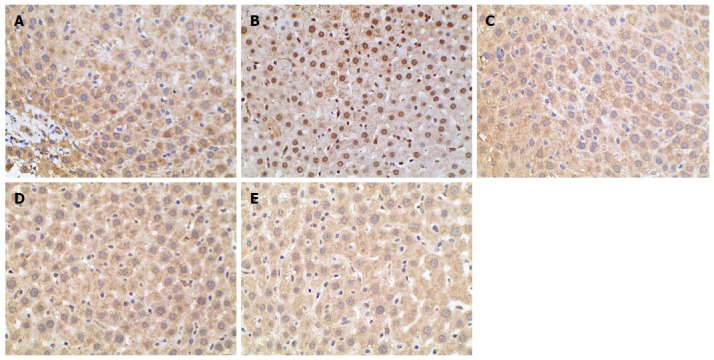
Immunohistochemical staining for TORC2 in the liver tissues. Optical microscopy image of TORC2 is shown in brown. The normal group (A) exhibited little TORC2 in the nuclei. However, more TORC2 was present in the nuclei of the model group (B). The groups treated with Berberine (C), AICAR (D) and Metformin (E) exhibited lower levels of TORC2 compared to the model group (magnification × 400).
Berberine regulated expression of PEPCK and G-6-P proteins in livers of type 2 diabetic rats
PEPCK and G-6-P are key gluconeogenesis enzymes and can affect plasma glucose. Expression of PEPCK and G-6-P proteins was increased in the model rats compared to the normal control group (P < 0.01), and treatment with berberine, AICAR and metformin decreased expression of PEPCK and G-6-P protein compared to the model rats (P < 0.05) (Figure 7).
Figure 7.
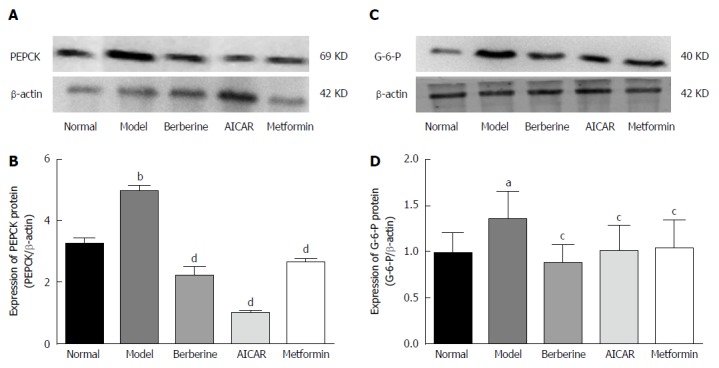
Berberine inhibited expression of key gluconeogenic enzyme proteins. Western blot analyses of PEPCK and G-6-P proteins in liver tissues of normal rats, model rats and diabetic rats treated with berberine, AICAR or metformin. A: PEPCK blots for each group are shown; C: G-6-P blots for each group are shown; B: Each bar is expressed as the total PEPCK/β-actin and represents the mean ± SD (n = 8); D: Each bar is expressed as the total G-6-P/β-actin and represents the mean ± SD (n = 8). bP < 0.01 vs normal control group; dP < 0.01 vs model group; aP < 0.05 vs normal control group; cP < 0.05 vs model group (by ANOVA).
DISCUSSION
Berberine was first found to exhibit hypoglycemic actions in 1988, and numerous studies related to the ability of berberine to attenuate diabetes have been reported in the last 25 years. Previous evidence has shown that berberine can decrease blood glucose, regulate lipids, and improve insulin resistance via many different molecular mechanisms[12-14]; however, little research has focused on whether berberine inhibits hepatic gluconeogenesis via AMPK. Previous studies have illustrated that the regulation of gluconeogenesis is involved in the insulin signaling pathway. In re-feeding mice, insulin inhibits gluconeogenic gene expression via the promotion of the phosphorylation of TOCR2[15]. In the models of insulin signaling deficiency, the expression of PGC-1 which plays a role in liver gluconeogenesis is elevated. Thus, insulin is a primary suppressor of gluconeogenesis[16]. However, the current study revealed that glucose metabolism was regulated independently of insulin action. The loss of LKB1 in the mouse liver resulted in an increase in TOCR2 gene expression and drove gluconeogenesis via the AMPK signaling pathway[4]. In a clinical trial, Keshavarz et al[17] examined identification of single nucleotide polymorphisms in LKB1 and TOCR2 genes, and the results suggested a probable association between the LKB1-AMPK-TOCR2 signaling pathway and glucose homeostasis in the liver. These studies provided more insight to consider whether berberine suppresses gluconeogenesis to attenuate hyperglycemia via the AMPK signaling pathway.
In this study, we showed that berberine restrained protein expression of the key gluconeogenic enzymes PEPCK and G-6-Pase in model rats (Figure 7). These results agree with those of previous reports[9,10]. Berberine inhibited PEPCK and G-6-Pase protein expression via the suppression of mitochondrial function[10]. The glucose-lowering effect of berberine is related to the suppression of the expression of the key hepatic gluconeogenic enzymes PEPCK and G-6-Pase via the AMPK signaling pathway[9]. AMPK is a potential target for balancing glucose and lipid metabolism in the treatment of type 2 diabetes. Berberine treatment increases AMPK activity and contributes to the elevations in the level of AMPK phosphorylation in the liver[9,10,18,19]. In the present study, we examined the protein expression of AMPK and p-AMPK in the liver tissues (Figure 4). We observed that berberine increased the amount of total AMPK and phosphorylation of AMPK. Treatment with berberine restored the AMPK activity observed in the diabetic condition to the level observed in the non-diabetic condition (Figure 5). This increase in AMPK activity was accompanied by reductions in PEPCK and G-6-Pase expression. These results are consistent with previous data. The research of Shaw et al[4] provided us with inspiration to explore further the hypoglycemic actions of berberine. In their study, LKB1 deletion in the liver led to a reduction in AMPK phosphorylation; thus, the activation of AMPK depends on LKB1. We considered whether LKB1 acts as a critical upstream target of AMPK when berberine treatment is accompanied by a change in AMPK. In our study, we measured the expression of LKB1 in the diabetic liver. Intriguingly, we found that LKB1 protein expression in treated groups was increased compared to the levels observed in the diabetic rats (Figure 3). Next, we sought to understand how AMPK affects the expression of the gluconeogenic enzymes PEPCK and G-6-Pase. Koo et al[5] reported that the activation of AMPK promotes TOCR2 phosphorylation and blocks its nuclear accumulation. Consequently, gluconeogenic enzyme expression is interrupted[4,20]. In the current research, we detected no significant difference in the total amount of TOCR2 between the normal and diabetic rats, but TOCR2 phosphorylation in the cytoplasm was increased by the berberine treatment relative to model rats (Figure 5). Berberine treatment inhibited the translocation of TOCR2 into the cell nucleus, and the TORC2 nuclear accumulation observed in the berberine group was lower than that observed in the model group (Figure 6). Thus, the transcription of gluconeogenic genes was reduced, and the liver glucose output was decreased. In our study, we observed lower blood glucose levels in the treated group than in the model group (Figure 1). High blood glucose levels stimulate the pancreas to secrete insulin and result in hyperinsulinemia. Our results revealed that berberine treatment reduced fasting insulin level compared to those observed in the model group (Figure 2).
To research the therapeutic effects of berberine, we chose to use AICAR and metformin as positive control groups. Some studies have shown that AICAR and metformin are AMPK agonists, and that they inhibit gluconeogenesis to regulate glucose metabolism through the AMPK signaling pathway[4,21-23]. In our research, we found no significant differences between these treatment groups.
In conclusion, our study revealed that berberine inhibited expression of the gluconeogenic proteins PEPCK and G-6-Pase in the liver. Consequently, reductions in blood glucose levels were accompanied by reductions in blood insulin levels reduction due to the inhibition of gluconeogenesis. Moreover, blood lipid levels simultaneously improved (Table 1). The mechanisms responsible for the effects of berberine treatment might be related to the suppression of gluconeogenesis through the LKB1-AMPK-TOCR2 signaling pathway.
COMMENTS
Background
Numerous studies related to the ability of berberine to attenuate diabetes have been reported. Previous evidence has shown that berberine can decrease blood glucose, regulate lipids, and improve insulin resistance via many different molecular mechanisms. However, little research has focused on whether berberine inhibits hepatic gluconeogenesis via AMP-activated protein kinase (AMPK).
Research frontiers
Animal experiments showed that the loss of liver kinase (LK)B1 in the mouse liver resulted in an increase in CREB-regulated transcription co-activator (TORC)2 gene expression and drove gluconeogenesis via the AMPK signaling pathway. Moreover, a clinical trial suggested a probable association between the LKB1-AMPK-TOCR2 signaling pathway and glucose homeostasis in the liver. Recently, berberine was proven to be capable of reducing hyperglycemia via the inhibition of hepatic gluconeogenesis. Therefore, we hypothesized that berberine reduces hyperglycemia via the LKB1-AMPK-TORC2 signaling pathway to control gluconeogenesis.
Innovations and breakthroughs
This is the first study to show that berberine reduces hyperglycemia via the LKB1-AMPK-TORC2 signaling pathway to control gluconeogenesis.
Applications
In the future, the LKB1-AMPK-TORC2 signaling pathway will probably be a target for berberine treating type 2 diabetes.
Terminology
Hepatic gluconeogenesis is strongly stimulated in the fasting state and converts glycogen into glucose to increase glucose output. AMPK is a conserved sensor and regulator of cellar energy balance that is activated when the cellular AMP: ATP ratio exhibits a large increase.
Peer-review
In this paper, the authors identified the association between the LKB1- AMPK-TOCR2 signaling pathway and glucose homeostasis in the liver. At the time, this study proved the molecular mechanisms of berberine inhibiting hepatic gluconeogenesis. The research is important for further research of berberine.
Footnotes
Supported by National Natural Science Foundation of China, No. 30973836.
Ethics approval: The various ethics statements related to scientific conduct are detailed below.
Institutional animal care and use committee: All procedures involving animals were reviewed and approved by the Institutional Animal Care and Use Committee at Tongji Medical College, Huazhong University of Science and Technology (IACUC number: 372).
Conflict-of-interest statement: The authors declare that there is no conflict of interests regarding the publication of this paper.
Data sharing statement: Technical appendix, statistical code, and dataset available from the corresponding author at pyi219@163.com. Participants gave informed consent for data sharing. No additional data are available.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: January 6, 2015
First decision: January 22, 2015
Article in press: March 31, 2015
P- Reviewer: Xu Z, Wang L S- Editor: Qi Y L- Editor: Kerr C E- Editor: Wang CH
References
- 1.Radziuk J, Pye S. Hepatic glucose uptake, gluconeogenesis and the regulation of glycogen synthesis. Diabetes Metab Res Rev. 2001;17:250–272. doi: 10.1002/dmrr.217. [DOI] [PubMed] [Google Scholar]
- 2.Kahn BB, Alquier T, Carling D, Hardie DG. AMP-activated protein kinase: ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005;1:15–25. doi: 10.1016/j.cmet.2004.12.003. [DOI] [PubMed] [Google Scholar]
- 3.Hemminki A, Markie D, Tomlinson I, Avizienyte E, Roth S, Loukola A, Bignell G, Warren W, Aminoff M, Höglund P, et al. A serine/threonine kinase gene defective in Peutz-Jeghers syndrome. Nature. 1998;391:184–187. doi: 10.1038/34432. [DOI] [PubMed] [Google Scholar]
- 4.Shaw RJ, Lamia KA, Vasquez D, Koo SH, Bardeesy N, Depinho RA, Montminy M, Cantley LC. The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science. 2005;310:1642–1646. doi: 10.1126/science.1120781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Koo SH, Flechner L, Qi L, Zhang X, Screaton RA, Jeffries S, Hedrick S, Xu W, Boussouar F, Brindle P, et al. The CREB coactivator TORC2 is a key regulator of fasting glucose metabolism. Nature. 2005;437:1109–1111. doi: 10.1038/nature03967. [DOI] [PubMed] [Google Scholar]
- 6.Lin J, Wu PH, Tarr PT, Lindenberg KS, St-Pierre J, Zhang CY, Mootha VK, Jäger S, Vianna CR, Reznick RM, et al. Defects in adaptive energy metabolism with CNS-linked hyperactivity in PGC-1alpha null mice. Cell. 2004;119:121–135. doi: 10.1016/j.cell.2004.09.013. [DOI] [PubMed] [Google Scholar]
- 7.Yoon JC, Puigserver P, Chen G, Donovan J, Wu Z, Rhee J, Adelmant G, Stafford J, Kahn CR, Granner DK, et al. Control of hepatic gluconeogenesis through the transcriptional coactivator PGC-1. Nature. 2001;413:131–138. doi: 10.1038/35093050. [DOI] [PubMed] [Google Scholar]
- 8.Ni YX. [Therapeutic effect of berberine on 60 patients with type II diabetes mellitus and experimental research] Zhong Xi Yi Jie He Za Zhi. 1988;8:711–713, 707. [PubMed] [Google Scholar]
- 9.Zhang M, Lv X, Li J, Meng Z, Wang Q, Chang W, Li W, Chen L, Liu Y. Sodium caprate augments the hypoglycemic effect of berberine via AMPK in inhibiting hepatic gluconeogenesis. Mol Cell Endocrinol. 2012;363:122–130. doi: 10.1016/j.mce.2012.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xia X, Yan J, Shen Y, Tang K, Yin J, Zhang Y, Yang D, Liang H, Ye J, Weng J. Berberine improves glucose metabolism in diabetic rats by inhibition of hepatic gluconeogenesis. PLoS One. 2011;6:e16556. doi: 10.1371/journal.pone.0016556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li JB, Xu LJ, Dong H, Huang ZY, Zhao Y, Chen G, Lu FE. Effects of Chinese Fructus Mume formula and its separated prescription extract on insulin resistance in type 2 diabetic rats. J Huazhong Univ Sci Technolog Med Sci. 2013;33:877–885. doi: 10.1007/s11596-013-1215-7. [DOI] [PubMed] [Google Scholar]
- 12.Liu X, Li G, Zhu H, Huang L, Liu Y, Ma C, Qin C. Beneficial effect of berberine on hepatic insulin resistance in diabetic hamsters possibly involves in SREBPs, LXRα and PPARα transcriptional programs. Endocr J. 2010;57:881–893. doi: 10.1507/endocrj.k10e-043. [DOI] [PubMed] [Google Scholar]
- 13.Kong WJ, Zhang H, Song DQ, Xue R, Zhao W, Wei J, Wang YM, Shan N, Zhou ZX, Yang P, et al. Berberine reduces insulin resistance through protein kinase C-dependent up-regulation of insulin receptor expression. Metabolism. 2009;58:109–119. doi: 10.1016/j.metabol.2008.08.013. [DOI] [PubMed] [Google Scholar]
- 14.Zhou L, Yang Y, Wang X, Liu S, Shang W, Yuan G, Li F, Tang J, Chen M, Chen J. Berberine stimulates glucose transport through a mechanism distinct from insulin. Metabolism. 2007;56:405–412. doi: 10.1016/j.metabol.2006.10.025. [DOI] [PubMed] [Google Scholar]
- 15.Dentin R, Liu Y, Koo SH, Hedrick S, Vargas T, Heredia J, Yates J, Montminy M. Insulin modulates gluconeogenesis by inhibition of the coactivator TORC2. Nature. 2007;449:366–369. doi: 10.1038/nature06128. [DOI] [PubMed] [Google Scholar]
- 16.Puigserver P, Rhee J, Donovan J, Walkey CJ, Yoon JC, Oriente F, Kitamura Y, Altomonte J, Dong H, Accili D, et al. Insulin-regulated hepatic gluconeogenesis through FOXO1-PGC-1alpha interaction. Nature. 2003;423:550–555. doi: 10.1038/nature01667. [DOI] [PubMed] [Google Scholar]
- 17.Keshavarz P, Inoue H, Nakamura N, Yoshikawa T, Tanahashi T, Itakura M. Single nucleotide polymorphisms in genes encoding LKB1 (STK11), TORC2 (CRTC2) and AMPK alpha2-subunit (PRKAA2) and risk of type 2 diabetes. Mol Genet Metab. 2008;93:200–209. doi: 10.1016/j.ymgme.2007.08.125. [DOI] [PubMed] [Google Scholar]
- 18.Cao S, Zhou Y, Xu P, Wang Y, Yan J, Bin W, Qiu F, Kang N. Berberine metabolites exhibit triglyceride-lowering effects via activation of AMP-activated protein kinase in Hep G2 cells. J Ethnopharmacol. 2013;149:576–582. doi: 10.1016/j.jep.2013.07.025. [DOI] [PubMed] [Google Scholar]
- 19.Ge Y, Zhang Y, Li R, Chen W, Li Y, Chen G. Berberine regulated Gck, G6pc, Pck1 and Srebp-1c expression and activated AMP-activated protein kinase in primary rat hepatocytes. Int J Biol Sci. 2011;7:673–684. doi: 10.7150/ijbs.7.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Screaton RA, Conkright MD, Katoh Y, Best JL, Canettieri G, Jeffries S, Guzman E, Niessen S, Yates JR, Takemori H, et al. The CREB coactivator TORC2 functions as a calcium- and cAMP-sensitive coincidence detector. Cell. 2004;119:61–74. doi: 10.1016/j.cell.2004.09.015. [DOI] [PubMed] [Google Scholar]
- 21.Aatsinki SM, Buler M, Salomäki H, Koulu M, Pavek P, Hakkola J. Metformin induces PGC-1α expression and selectively affects hepatic PGC-1α functions. Br J Pharmacol. 2014;171:2351–2363. doi: 10.1111/bph.12585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.He L, Sabet A, Djedjos S, Miller R, Sun X, Hussain MA, Radovick S, Wondisford FE. Metformin and insulin suppress hepatic gluconeogenesis through phosphorylation of CREB binding protein. Cell. 2009;137:635–646. doi: 10.1016/j.cell.2009.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shirai T, Inoue E, Ishimi Y, Yamauchi J. AICAR response element binding protein (AREBP), a key modulator of hepatic glucose production regulated by AMPK in vivo. Biochem Biophys Res Commun. 2011;414:287–291. doi: 10.1016/j.bbrc.2011.08.120. [DOI] [PubMed] [Google Scholar]