

Abstract
Computer-aided sperm analysis (CASA) technology was developed in the late 1980s for analyzing sperm movement characteristics or kinematics and has been highly successful in enabling this field of research. CASA has also been used with great success for measuring semen characteristics such as sperm concentration and proportions of progressive motility in many animal species, including wide application in domesticated animal production laboratories and reproductive toxicology. However, attempts to use CASA for human clinical semen analysis have largely met with poor success due to the inherent difficulties presented by many human semen samples caused by sperm clumping and heavy background debris that, until now, have precluded accurate digital image analysis. The authors review the improved capabilities of two modern CASA platforms (Hamilton Thorne CASA-II and Microptic SCA6) and consider their current and future applications with particular reference to directing our focus towards using this technology to assess functional rather than simple descriptive characteristics of spermatozoa. Specific requirements for validating CASA technology as a semi-automated system for human semen analysis are also provided, with particular reference to the accuracy and uncertainty of measurement expected of a robust medical laboratory test for implementation in clinical laboratories operating according to modern accreditation standards.
HISTORICAL INTRODUCTION
Interest in tracking sperm movement, and trying to understand what it can tell us about sperm biology and functional competence, goes back six decades to classic papers published by Lord Rothschild, Gray, Rikmenspoel and their co-workers.1,2,3,4,5 However, the “modern era” of Eutherian sperm kinematics, based originally on manual sperm tracking, has its origins in the 1970s in the work of Prof George David's group in Le Kremlin-Bicêtre (Paris, France), and Hector Dott's laboratory in Cambridge (UK), which were developing ways of using photo- and cine-micrography for quantifying sperm motility.6,7 David Katz and Jim Overstreet (Davis, CA, USA) extended basic work on sperm microcinematography8 and then pioneered the use of videomicrography9 which was rapidly taken up by other workers.10,11 There are a number of major reviews on Eutherian sperm motility and its analysis.12,13,14,15
The basic criteria for sperm kinematics (characteristics that describe sperm movement, as opposed to the proportion of motile cells) that we use today, originated from the work of David's group using 50 fps microcinematography.7 Expanding interest in the field led to confusing terminology and definitions of various kinematic criteria, but these were resolved in 198816 and adopted by the WHO in the 3rd edition of their lab manual in 1992.17 The consensus that was reached by almost everyone working in the field at that time included the three-letter terminology and related definitions (e.g., VCL, LIN, ALH, etc.), as well as the agreed precision of reporting results: measured values such as velocities to 1 decimal place, ratios as integer percentage values.
With the advent of the personal computer in the 1980s, and the development of digitization of video images for automated reconstruction and analysis of individual sperm tracks, there was an explosion in interest in the field, and the first commercial CASA (computer-aided sperm analysis) instruments appeared:16 CellSoft in 1985, followed soon after by the Hamilton-Thorn HTM-2000. In 1992 the proprietary Sperm Motility Quantifier (SMQ) was one of the first systems enabling routine analysis at a 50 Hz frame rate.18
As the field developed further, some scientists wanted to move from using CASA as a research tool that had been designed for analyzing sperm motility and kinematics, to using it for automated human semen analysis in the clinical setting. Unfortunately, due to fundamental limitations of the image analysis technology (largely due to the limited resolution of the then-available digitized images), this was unsuccessful in terms of accuracy compared to standardized assessments made by trained observers.19,20 This major limitation of 1980s–1990s CASA for human semen analysis was addressed in three consensus meetings: at the October 1994 Cairns 7th Symposium on Spermatology,21 and in two ESHRE workshops held in Hamburg and San Miniato, in June 1995 and April 1997 respectively.22,23 The 1998 San Miniato report included the following consensus recommendations in regard to using CASA for human semen analysis:
-
#14.
Determination of sperm concentration must not be a primary reason for acquiring a CASA instrument
-
#15.
If a user wishes to use a CASA instrument to determine sperm concentration then he/she must establish that the intended measurement procedure provides accurate results compared with established, reliable techniques (e.g., WHO haemocytometry method and perhaps flow cytometry; WHO, 199217)
-
#16.
It is considered that the current generation of CASA instruments does not provide accurate, reproducible values for sperm concentration unless the method can differentiate spermatozoa from other cells and debris by a specific staining method, e.g., fluorescent staining of DNA with quantitative determination of nucleus size (Zinaman et al., 1996)24
-
#17.
Determination of the proportion of motile spermatozoa should not be considered a primary reason for acquiring a CASA instrument
-
#18.
If used in the routine analysis of seminal spermatozoa, CASA should be used to determine the concentration of progressively motile spermatozoa. CASA can determine this value accurately if care is taken with specimen preparation, instrument use and appropriate user-defined criteria (Mortimer et al., 1995).21 Current CASA instruments should not be used for the determination of the proportion of motile spermatozoa, since they cannot be relied upon to distinguish between debris and dead spermatozoa while tracking live spermatozoa at the same time
-
#31.
The current generation of CASA instruments is not capable of analysing human sperm morphology in a manner adequate for routine clinical applications. In particular, the inability to include assessment of the midpiece and tail regions (required by the WHO guidelines) is considered to be a major weakness. Consequently, the use of CASA instruments for the clinical assessment of human sperm morphology is not supported at this time (WHO, 1992).17
It must be emphasized that these consensus points reflect the situation as in 1998 – but unless a CASA system employs significantly improved technology and/or software the same concerns and limitations remain valid today. It being almost exactly 20 years after the Cairns meeting, it was believed appropriate for the 2014 12th Spermatology Symposium to include a workshop on the current and near future capabilities of CASA technology, based on the authors’ personal experience with two “next generation” CASA systems that incorporate improved capabilities.
EXPECTATIONS OF ACCURACY AND PRECISION FOR MEDICAL LABORATORY TESTS
Fundamental standards for all medical laboratories are well-established and clearly defined in ISO 15189:2007. As for any medical laboratory test, the results of a semen analysis must be accurate to have any real world value.25,26 Any quantitative result must be within an acceptable range of the “right answer”, i.e., its Uncertainty of Measurement (UoM) must be known relative to established reference standards or methods: an “expert” andrology lab should certainly achieve a UoM of ± 10%.26 As for any medical testing lab, internal quality control (IQC) and external quality assurance (EQA) are essential.26,27 However, for an EQA scheme to have any real value then it must include the capability of quality improvement. A lab must know how close its results are to the correct values for it to be able to apply remedial training and thereby improve if its results are outside the acceptable range of uncertainty.
Therefore, results for semen analysis characteristics obtained using CASA in a clinical andrology laboratory setting need to be within ± 10% of reference values for an expert andrology laboratory (although ± 20% might be adequate for a general diagnostic laboratory). The assumption here is that all clinical andrology laboratories conform to current accepted “Gold Standard” semen analysis methodology, such as the guidelines from the World Health Organization,28 or the ESHRE Andrology SIG,26 with all laboratory staff having been properly trained in all the protocols and quality control systems. Unfortunately, 34 years after the publication of the first WHO laboratory manual for semen analysis, this still remains a major deficit in the majority of andrology laboratories globally. A recent 3-year survey of 40 clinical andrology laboratories in 22 countries revealed that more than 90% had nonconformities in either correct use of the microscope, temperature control, general standardization, analytical quality control, or adherence to current WHO guidelines.29 Poor conformity to international guidelines is well known,30,31 and seems to be especially low in assisted conception centres given that audits carried out over the past 15 years revealed that only 1/47 centres across 14 countries materially conformed to established standards for their andrology laboratory services.32
When using CASA there is also the difficulty that a fixed depth preparation must be made for analysis in the instrument, and that such a preparation really cannot exceed 20 μm for reliable tracking of the spermatozoa under a 10 × objective. Although many commercial slides/chambers exist for making these preparations, all those with a fixed cover glass require loading by capillary action and will therefore be subject to the Segre-Silberberg effect that influences the perceived sperm concentration as a result of laminar flow artefacts, resulting in a viscosity-dependent under-estimation by up to 30% at aqueous viscosity.33,34 However, chambers such as the 2X-CEL and the Cell-VU which use a separate cover glass over drops of sperm suspension (“drop-loading”) are not subject to this error. Consequently, if a CASA instrument evaluation were to compare un-corrected sperm concentration values determined using capillary loaded slides in the instrument against reference values derived using careful haemocytometry and found a good equivalence, it would, in effect, be establishing that the CASA instrument actually had a variable over-estimation, and hence the trial outcome was fortuitous.
A meticulous study comparing the Makler chamber (10 μm) and Leja 20 μm slide to the Improved Neubauer haemocytometer has shown that while the Makler chamber is highly inaccurate for determining sperm concentration, typically giving significantly elevated values, the Leja slide can achieve sufficient comparability to the haemocytometer to allow it to be used clinically.31 However, this equivalence was only achieved by determining the time taken to load the chamber by capillary action and applying the appropriate correction factor for the Segre-Silberberg effect,33,34 and by pre-diluting the semen: neither of which steps are taken when these chambers are used in routine clinical lab practice. Furthermore, there will always remain the issue of robust sampling from even fully-liquefied and well-mixed human ejaculates when taking aliquots of only 4 or 5 μl to load a chamber for CASA analysis.31
It must also be emphasized that comparisons between systems cannot be based on average values or on correlations/linear regression between paired values: both techniques conceal the real differences that exist between specific replicate determinations. Instead, Bland and Altman “limits of agreement” or “discrepancy” plots must be used,19,26,35 which plot the actual differences between the paired values, and can hence be used to establish whether each “new method” (or “trainee”) value is within ± 10% of the reference method or value. This is a “gold standard” comparison technique, and reports using other statistical approaches, many of which typically conceal the true extent of the existing discrepancy, must be viewed with caution. Indeed, the outcome of Bland and Altman plot analysis can always be due to chance, and this possibility should be excluded employing Passing and Bablok analysis,36 using, for example, MedCalc (see www.medcalc.be). Another adjunct to Bland and Altman plot analysis is the Deming regression method (also available in MedCalc) which provides insights in establishing concordance between different methods tested in duplicate, and also gives the coefficient of variation.
WHAT ARE THE FACTORS LIMITING CASA'S FUNCTIONALITY FOR (HUMAN) SEMEN ANALYSIS?
While CASA technology is widely used with excellent results in animal production laboratories and on wildlife species,37,38 the same is not yet true for human clinical laboratories. Those who use it for this purpose have (presumably) accepted that its results are more robust than what might be obtained from inadequately trained semen analysis technicians, and/or their lower determination cost precludes the use of more accurate assessments. However, it must be noted that, since its introduction, CASA has worked extremely well on washed human sperm populations, which typically have very high motility and minimal contamination with other cells and debris. The limitations affecting CASA's ability to provide accurate results for sperm concentration and percentages of motile or progressively motile spermatozoa, fall into two major categories: biological (Table 1) and technical.19,20,21
Table 1.
Biological factors that affect CASA analysis, a comparison of semen samples for humans and domesticated species

Technical limitations centre on issues relating to digital image analysis and the fundamentally different approaches used to define motility. In regard to the latter, human observation defines and classifies sperm motility according to flagellar beating and spatial displacement of the cell (Table 2), but CASA instruments cannot analyze flagellar beating directly, and must rely on tracking the movement of the sperm head. In extreme situations, Brownian motion of immotile objects of similar size and appearance to sperm heads can be mistaken for sperm motility, and even establishing a robust definition for non-progressive sperm motility can be difficult when movement of the flagellum is not able to be used. In terms of image analysis there are several problems that stem from errors in discriminating between spermatozoa and non-sperm objects, and between immotile and motile objects:
Table 2.
Definition and classification of human sperm motility categories26
When several spermatozoa are clumped together (even as few as two) or with debris, the resulting single contiguous object that the instrument “sees” following digitization of the image will be too large to be classified as a sperm head (based on head size gating) and hence will be rejected from the analysis. This means that the overall sperm concentration will be reduced and, since such spermatozoa are typically immotile (or at least non-progressive), the proportions of progressive spermatozoa will be increased. Unfortunately, such “micro-aggregates” are very common in human semen samples.
Especially in instruments employing negative phase contrast optics or dark ground illumination (i.e., a dark or black background), the mis-identification of non-sperm objects (such as large pieces of debris) as “spurious” spermatozoa remains a vexed issue. Obviously this would increase the apparent total number of spermatozoa in the field of view.
Immotile spermatozoa (or even appropriately-sized debris) can be “stirred” by the flagellar beating of nearby motile spermatozoa and erroneously classified as motile spermatozoa. This would increase the apparent motility, as well as the total number of spermatozoa in the case of agitated debris or other cells.
Tracking motile spermatozoa through collisions (real or even “perceived” collisions as spermatozoa swim close by each other within the temporal-spatial discrimination of the instrument) can result in track fragmentation and thereby increase the perceived concentration and proportions of motile and progressively motile spermatozoa. Even if a CASA system has “smart” collision correcting algorithms, there can still be disturbances in the cells’ trajectories, and hence their kinematics.
While the use of positive phase contrast optics (the most common type of such optics) will reduce the mis-classification of debris as sperm heads, it cannot resolve the issue of the contiguous digitization of adjacent spermatozoa (± other cells). The issue of specimen “drift” is also unavoidable, and analyses must not be commenced until all visible “flow” has ceased. The recent innovation of a “drift filter” in the SCA software can counter this artefact (see below).
HOW HAVE MODERN CASA SYSTEMS EVOLVED?
Despite many unresolved problems, the state-of-the-art in CASA has progressed greatly in the last decade so that several modern CASA systems are no longer “black boxes” but embody additional (and often user-adjustable) operational parameter settings that address many of the problems that constrained earlier systems. Progress in resolving many of these image analysis issues, based on the cost-effective availability of higher resolution digitizers and greater computing power combined with software features such as automated and/or interactive illumination control, advanced Brownian motion filtering, drift filtering, tail detection, smart tracking through collisions, adaptive smoothing to derive the average path, and the introduction of fractals as a kinematic measure, have given us CASA instruments that are certainly more user-friendly in both the human clinical and domestic animal fields.
Hamilton-Thorne IVOS-II and CEROS-II systems
Recently we (DM, STM) have been working with the “second generation” CASA platform from Hamilton Thorne (Beverly, MA, USA), the IVOS-II (IVOS = Integrated Visual Optical System) which features improved integrated phase contrast optics; a much greater (5×) image resolution; a 1 ms stroboscopic illumination system that is synchronized to the 60 images/s video camera (which is really the minimum imaging frequency required for reliable human sperm track analysis39) to eliminate “smearing” of the images of moving sperm heads; tail detection for better spermatozoon discrimination; adaptive smoothing for better derivation of the average path (very important for hyperactivation analysis using traditional kinematic definitions; see below); and improved smart tracking through collisions. In addition, the range of kinematic measures has been expanded to include the fractal dimension.40,41,42,43 In addition to the traditional operational settings such as frame rate, number of images to be captured, and specimen chamber type/depth, there is now interactive/automated illumination and focus control to optimize tail detection, as well as user-adjustable settings to control more aspects of cell detection and static/non-progressive/progressive motility differentiation. The long-established Sort function now provides greater user control over the Boolean definitions that can be created to classify individual motile spermatozoa into functional sub-populations based on multiple kinematic characteristics (e.g., hyperactivating spermatozoa, or those that show “good mucus penetrating” characteristics44).
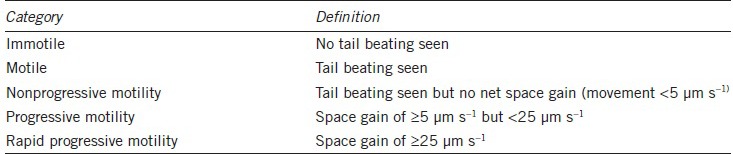
As before, in the Results View mode individual tracks can be selected and zoomed to show the track details and the derived kinematic values, but now the individual frame images of the spermatozoon, including the proximal part of its tail, are also superimposed on the reconstructed track (Figure 1). This is not just an excellent analytical tool but also a powerful teaching tool, allowing users to understand how the beating of the sperm tail moves the sperm head and creates the track that is the basis for the kinematic analysis.
Figure 1.
The upper panel shows the reconstructed sperm tracks in one analysis field of the IVOS-II system (1 s @ 60 Hz), purple tracks are ones that have passed the Boolean sort argument for being hyperactivated. The lower panel shows the zoomed image of one of these tracks including the spermatozoon at the start of the track; note the high curvature wave, characteristic of a hyperactivated spermatozoon, present in the proximal region of the tail. The cell's kinematic values are shown at the bottom of the figure, including the fractal dimension, D, clearly confirming hyperactivation.60
The inbuilt fluorescent imaging options (either LED-based or xenon strobe) allow for sperm detection only via their highly condensed nuclei (eliminating issues with debris and fat droplets) either while the spermatozoa are still motile (“VIADENT”) or after immobilization (“IDENT”). The CASA-II software that runs the IVOS can also be used in conjunction with standard external microscopes as a CEROS-II configured system, although this precludes the stroboscopic illumination system as well as software-based automated illumination control features of the IVOS platform. Options for sperm morphology include the research-orientated “Metrix” system that learns from user interaction for improved classification of sperm head morphology categories, and the Tygerberg Strict Criteria-based “Dimensions” system,45 which is the only human sperm morphology software that has been validated at the Tygerberg Hospital, now available as Dimensions II for the new platform. However, neither of these systems can analyze either the sperm midpiece or tail morphology, and hence only provides a classification of “normal” spermatozoa based on sperm head morphology – determination of the highly useful Teratozoospermia Index19,26,46 is not possible.
While the IVOS-II system has enjoyed widespread success in animal production laboratories, at the time of writing optimization of the IVOS-II “clinical” (human sperm) software is still undergoing refinement to better classify non-progressive and progressive spermatozoa across as wide a gamut of patient ejaculates as possible. Handling the issues stemming from the problem of obtaining truly representative aliquots from a human semen sample when using only 4–5 μl to load a chamber for CASA analysis – likely to be a limiting factor in achieving comparability between WHO/ESHRE and CASA semen analysis results – is also a work in progress.
The Sperm Class Analyzer (“SCA”: Microptic Automatic Diagnostic Systems SL, Barcelona, Spain) is another CASA system that has progressed to solve many of the abovementioned image analysis problems. The SCA 5.4 and SCA 6 Evolution system comprises four modules (concentration and motility, morphology, vitality and fragmentation) that are fully integrated and associated with an extensive database that will allow results import and export in many of the formats used by hospitals. It offers several platforms from human and veterinary to research and several others including automatic stage, datashare and capture stations far away from the analysis system. Rigorous evaluation of the first two modules (by GvdH as well as others) has included various primate species (including man, with generally successful results), all domesticated species and more than 60 other animal species including invertebrates (e.g., mussels, sea urchins), several fishes, amphibians and birds (e.g., ostrich, penguin and zebra finch) and mammals (including rat, mouse, elephant, rhinoceros and lion). It is one of the few systems that have never used video images, rather it is fully digital with distinct advantages such as using any frame rate within the camera's capabilities. With Firewire or Gigabit Ethernet cameras, frame rates up to 100 fps (cost effective) or even higher can be attained. As noted already, hyperactivating human spermatozoa should ideally be studied above 50 fps and ram spermatozoa in culture medium should be studied at no lower than 75 fps but preferably 100 fps.
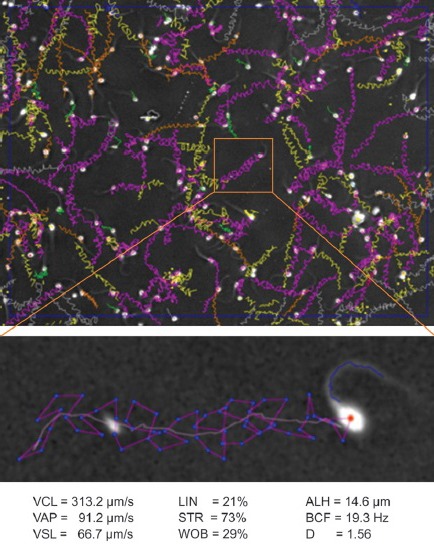
The SCA properties menu allows users to easily change many operational parameters including frame rate, number of images captured, for almost any chamber type/depth, cut-off points for immotile cells and progressive motility, as well as rapid, medium slow swimming sperm, average path smoothing from 3 to 15 track points (important for hyperactivation assessments using ALH), and settings to adapt sperm motility capturing for almost any animal species. No specific modules are required as many different species can be selected for ideal settings from the menu, or settings can be easily adapted and optimized for a new species. In addition, intelligent filters can be selected to further optimize sperm recognition/tail detection, debris rejection and advanced analysis in black and white mode (no greyscale) for semen samples with extremely “noisy” backgrounds (full of debris) as well as a filter system for drifting including Brownian movement and at least most collisions. Together these features assist greatly in the more accurate determination of percentage sperm motility. The SCA can also perform motility and vitality analyses simultaneously in fluorescent mode (only detecting DNA), which eliminates the problem of debris for human semen and in, domestic animals, the fat droplets in egg yolk-base cryopreservation extenders for freeze/thaw quality testing. The motility of spermatozoa in any analyzed field can be replayed with track overlays in a broad selection of colours. For example, rapid spermatozoa in green, or hyperactivated sperm in yellow, or different colours for other motility ratings based on either default (WHO4/528) or tailor-made settings. Finally it is possible to zoom into any analyzed spermatozoon to show the details of the sperm motility track and make an individual report on the selected cell. This is of particular value in establishing the kinematic characteristics of a particular sperm motility pattern (Figure 2). Yet, like all other CASA systems, it is difficult to analyze sperm concentration and motility accurately, if at all, when severe clumping is present.
Figure 2.
Spermatozoa from an Angus bull hyperactivated in capacitating medium with 10 mmol l-1 caffeine analyzed using the SCA (a). The numerous starspin patterns in the upper panel can be easily selected and their individual kinematic values can be recorded (b).29
SCA RatTox Module: the RatTox system is designed to investigate the effect of various substances/toxicants on both sperm function and form. The SCA RatTox module measures rat and mouse sperm motility (percentages and kinematics), morphometry/morphology, vitality and fragmentation in automatic mode. It is the only CASA system that is capable of analyzing rat and mouse sperm morphometry, which is particularly challenging given the peculiar hook-shaped head of most rodent spermatozoa, although it has the advantage of deriving several “new” morphometric parameters such as total curve shaped head length (Arc). The system also has potential for morphometric studies in wildlife rodent species with hooked shaped heads.
Future developments
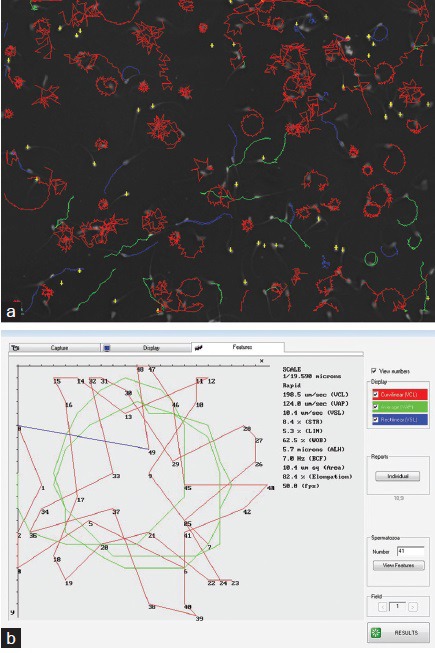
Sperm morphology needs to be better elaborated in humans and different animal species by including the entire cell, especially tail characteristics, and also using polychromatic stains. The SCA6 already employs adaptive thresholding to visualize the entire spermatozoon and then measure all its components, at least in ram spermatozoa (Figure 3). There should also be more uniformity in selecting stains for use in conjunction with CASA, specifically ones that are isosmotic and isotonic to spermatozoa and that produce a homogenous background, such as SpermBlue.47,48 Other approaches could include eliminating staining and looking at the possibilities of more detailed analysis using phase contrast and Nomarski differential interference contrast (DIC) optics.
Figure 3.
(a) SCA analysis of ram spermatozoa showing three columns of paired images of spermatozoa, in each of which the left sub-column is the original image and the right sub-column is the automated analysis of each spermatozoon; yellow = acrosome; blue = rest of head; green = anterior part of midpiece. (b) Ram spermatozoa stained with SpermBlue; (c) screen shot of the entire spermatozoon including its tail as analyzed by the SCA.
Modern CASA systems such as the IVOS-II and SCA6 platforms present new opportunities for CASA in general and will permit breakthroughs via their ability to analyze more parameters and thereby better define the quality of a sperm sample quantitatively and objectively, based on tests of sperm functional ability that will be fully automated. In this context SCA6 has also integrated sperm functional assessment using several sort functions such as sperm cervical mucus penetration and hyperactivation, as well as analyzing the ability to undergo the acrosome reaction. The SCA Vitality module is intended for determining “live/dead” ratios in automated mode in human and almost any animal species, but in the near future will also assess eosin-nigrosin staining automatically, meaning that costly fluorescence imaging will not be required. The SCA system already includes automated reading of the Halosperm® Test (Halotech DNA SL, Madrid, Spain) method for assessing sperm DNA fragmentation analysis, which is also being implemented on the IVOS-II platform. Future developments envisaged by Microptic include analyses involving further tests of sperm function associated with sperm-female reproductive tract interactions.
More intelligent interpretation of the results generated by CASA compared to traditional semen assessments will also be required. For example, the SCA6 system superimposes the actual patient values onto the minimum cut-off parameters (e.g., WHO528) and allocates a relative score. While such a strategy might be very useful, it still assumes that cut-offs and reference values have any real meaning in clinical andrology.25 Nonetheless, by integrating multiple sources of information, and taking a holistic approach to multivariate analysis, we should be able to achieve a far deeper, and pathophysiologically more meaningful, understanding of what parameters might define sperm “quality” in functional terms.
HOW SHOULD WE USE CASA IN HUMAN SEMEN ANALYSIS?
There is still considerable debate as to the clinical usefulness of the traditional descriptive semen analysis, which is held to exhibit very little prognostic value and limited diagnostic value since there is no “disease” of “infertility”.25 While it is clearly impossible to try and resolve such arguments here, it can be concluded that only with the widespread use of robust, accurate methods could we expect to be able to establish real clinical utility for semen analysis, and that such utility would be greatly enhanced by including aspects of sperm functional ability.25,26,46
There are two fundamental approaches to understanding the clinical utility of semen analysis:
As a means of assessing the man's reproductive health; and
Whether the man's ejaculate contains sufficient potentially functional spermatozoa to effectively colonize his partner's reproductive tract and reach the site of fertilization in the Fallopian tube.
In the latter regard, this sperm population is contained within the rapid progressive motility (grade “a”) fraction,46 and we have long employed a kinematic definition of which spermatozoa show “good mucus penetrating” characteristics (VAP >25 μm s−1 AND STR >80% AND ALH >2.5 μm, where the “AND” terms are Boolean arguments).44,49 If a CASA system is able to sub-classify motile spermatozoa according to multi-parametric kinematic definitions, e.g., the IVOS Sort function, a fundamental capability defined by experts more than 17 years ago,23 then this would be a clinically extremely important measure with both discriminative and prognostic value in regard to in vivo fertility endpoints. Moreover, it would provide more biologically important information than the traditional semen analysis characteristics of sperm concentration, total count, and the percentages of motile and even progressively motile, spermatozoa when considering management and ART treatment options for infertile couples (although not necessarily equally powerful in regard to the assessment of a man's overall reproductive health). For some 18 years, we (DM, STM) have used human sperm hyperactivation analysis, as well as determining the fraction of progressively motile spermatozoa in semen, as part of an integrated assessment of sperm functional potential that allows robust determination of the level of assisted conception treatment that a couple might require (i.e., timed intercourse, intrauterine insemination, IVF or ICSI).44
Besides these predictors of sperm fertilizing potential, the percentages of rapid, medium and slow swimming spermatozoa in semen are also considered by many andrologists to be of great importance, but primarily that of rapidly progressively motile spermatozoa.46 Maree and van der Horst50 described a method of determining the VCL cut-off values for these sub-classifications in several species including humans with the SCA, and these cut-off values have revealed differences in this measure of sperm quality among different fractions of spermatozoa obtained following density gradient centrifugation and in their response to caffeine51 (Figure 4).
Figure 4.
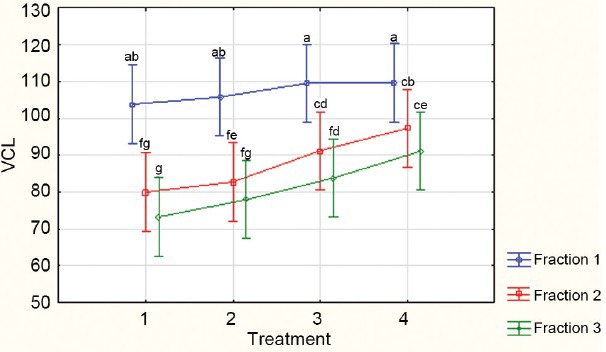
Percentages of rapidly progressive human spermatozoa from three PureSperm fractions (Fractions 1 to 3 = 40%, 60% and 80% respectively) compared when incubated with increasing concentrations of caffeine; Treatment 1 = control; Treatments 2–4 increasing concentrations of caffeine from 2 to 10 mmol l-1.51
THE ANALYSIS OF HUMAN SPERM HYPERACTIVATION
Hyperactivated motility (HA) is a concomitant of capacitation, characterized by the relatively slow development of large flagellar waves which are then propagated very quickly, leading to its description as “whiplash” motility. Although the precise physiological basis for HA has not yet been determined unequivocally,14 it has been suggested that it is related to the ability of the spermatozoon to penetrate the zona pellucida. Consequently, if a man's spermatozoa are unable to hyperactivate it would follow that he would have reduced fertility. Indeed, it has been reported that the proportion of HA spermatozoa following a 3 h incubation could explain up to 50% of the variance in fertilization rates in clinical IVF.52 Therefore, the assessment of HA in a capacitating sperm population (in the presence and absence of an agonist) has been included in the evaluation of sperm function used in the initial investigation for couples presenting with infertility.44
While HA is a flagellar phenomenon, the head trajectory is a good proxy for modelling flagellar movement, so CASA is a practical option for studying HA. Because the amplitude of the flagellar wave can be as high as 12 μm in HA spermatozoa,53 it was recommended that chamber depths of at least 30 μm be used for HA analysis,21 although deeper chambers were used in the initial studies of HA in the human.54 However, experience gained over the past 20 years has shown that 20 μm preparations can be used for reliable CASA analysis of human sperm HA.
At an image sampling frequency of 60 Hz, the Boolean argument for classification of human sperm HA is: VCL >150 μm s−1 AND LIN <50% AND ALH >7.0 μm.55 The potential problem with this argument is that, in CASA assessment, ALH is determined by calculation of the deviation of the average path from the actual, curvilinear, trajectory – but the points that make up the average path are themselves calculated from the average of the points on the curvilinear path. Since CASA instruments use different algorithms to derive the average path, ALH values are not consistent between instruments and so cannot be standardized across CASA platforms. Older CASA systems that still use fixed five-point smoothing to derive the average path, will provide inadequate smoothing and hence widely aberrant ALH values. An alternative measure that describes the degree of deviation of the trajectory from a straight line, and that is not dependent upon ALH calculation, is the fractal dimension (D).56
D is the “quantitative assessment of the ‘space-filling’ properties of curves on a plane”.57 According to Mandelbrot,56 while a straight line has one dimension and a plane has two dimensions, a curved line has a dimension that lies between 1 and 2. Katz and George57 developed algorithms in which the simplest trajectories returned D values close to 1, with more complex trajectories giving values approaching 2, and recursive trajectories having D values between 2 and 3, where the “layering” of the trajectory took into account the involvement of the third dimension. Davis and Siemers13 proposed the use of D to describe sperm trajectories, and it was later shown that in addition to being independent of the average path for its calculation, D was also independent of scale, although still influenced by the image sampling frequency.42,43 While D >1.30 was found to describe HA motility, it was noted that D alone was unlikely to identify HA tracks consistently, and that it would still need to be incorporated into a Boolean argument.40 It was not until recently that D was included in the values calculated by a CASA instrument (Hamilton Thorn IVOS-II), and so it is now possible to determine the practical value of D in the determination of HA motility.
Using the Hamilton Thorn IVOS-II, a total of 6240 trajectories of human spermatozoa incubated under capacitating conditions were followed for 1 s at 60 Hz. Classification of tracks as HA or non-HA were compared using the published Boolean argument for HA (VCL ≥150 μm s−1 AND LIN ≤50% AND ALH ≥7.0 μm) as well as using D >1.30, D >1.20, and VCL ≥150 μm s−1 AND D ≥1.20 (Figure 5). As had been predicted, D alone was not consistent in trajectory classification, with D >1.20 and D >1.30 giving only 63% and 67% agreement, respectively, with the published Boolean argument for HA. However, the definition which included both VCL and D gave a 99% agreement with the published Boolean argument in classification of trajectories.
Figure 5.
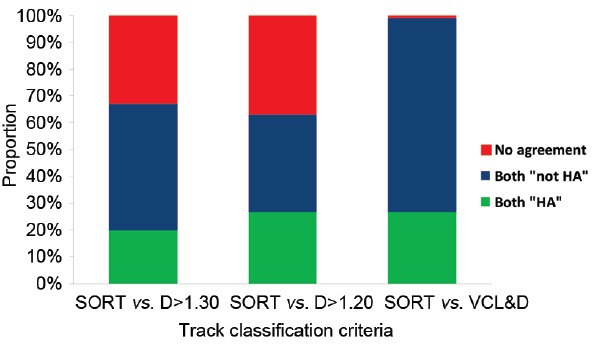
Comparison of methods for classification of capacitating sperm trajectories as HA or non-HA (Mortimer, unpublished data).
This development of a robust definition for HA that is independent of ALH should facilitate the incorporation of HA assessment in the initial work-up of an infertile couple, and promote the creation of personalized treatment plans.
ANALYZING HYPERACTIVATION IN OTHER SPECIES
Since many species of vertebrates, and even invertebrates, do not show a large degree of the “intermediate” hyperactivation pattern as seen in human spermatozoa (previously referred to as a “transitional” pattern, but now established as true HA based on the flagellar beat patterns of such spermatozoa),58 they often exhibit the classical “starspin” hyperactivation as indicated in Figure 2. Visual pattern recognition of these spermatozoa is therefore relatively simple compared to the more linear swimming spermatozoa. Establishing cut-off points for such tracks is relatively simple, involving comparing their kinematic parameters to those of the relatively straight swimming sperm tracks using receiver operating characteristic (ROC) curve analysis: those kinematic parameters that show high levels of specificity and sensitivity (>90%) are then used to construct a Boolean argument. This approach has been successfully used in conjunction with the Sort function of the SCA to establish the percentage HA in many species including bull, ram, elephant and rhinoceros (work currently in progress in the lab of GvdH). Using a similar approach, it was shown that for stallion spermatozoa, HA is required for the in vitro fertilization of equine oocytes.59
CONCLUSIONS
CASA has enormous potential as a research tool, in reproductive toxicology, in animal production, and for human clinical analyses. There is no doubt that the CASA systems of the future will need to be rigorously tested and conform to the differing needs of human clinical andrology laboratories, domesticated animal production laboratories, and reproductive toxicology testing and research laboratories. In addition, they will also need to incorporate functional tests of sperm quality that will assist in quantifying relative fertility, rather than merely measuring descriptive semen parameters per se.
However, especially in the human clinical semen analysis field, we should stop trying to use CASA for applications that are inherently problematic for the underlying technology until the technology has been improved and definitively validated. Currently, we should focus on what we know CASA can do accurately, which in terms of human sperm analysis comprises:
Semen analysis: the concentration of that sub-population of spermatozoa that have the right kinematics to penetrate cervical mucus;
Washed sperm preparation analysis: sperm concentration and motility classes; and
Sperm fertilizing ability: evaluation of sperm hyperactivation, an essential capability for fertilization, both in vivo and in vitro.
Urgent improvements in CASA technology need to address sperm morphology analysis, as well as incorporating tests of sperm function into the platforms. Future CASA systems could provide improved clinical relevance for semen analysis by integrating the automated analysis of semen parameters and sperm functional assessments, and perhaps even incorporating molecular biological aspects of sperm analysis, into a database with specialized statistical analytical capabilities. This type of approach could also be adopted for the domestic animal industry to better predict the relative fertility of sires.
REFERENCES
- 1.Gray J. The movement of sea-urchin spermatozoa. J Exp Biol. 1955;32:775–801. [Google Scholar]
- 2.Gray J. The movement of spermatozoa of the bull. J Exp Biol. 1958;35:96–108. [Google Scholar]
- 3.Rikmenspoel R, van Herpen G. Photoelectric and cinematographic measurements of the motility of bull sperm cells. Phys Med Biol. 1957;2:54–63. doi: 10.1088/0031-9155/2/1/306. [DOI] [PubMed] [Google Scholar]
- 4.Rothschild L, Swann MM. The fertilization reaction in the sea-urchin egg; a propagated response to sperm attachment. J Exp Biol. 1949;26:164–76. doi: 10.1242/jeb.26.2.164. 4 pl. [DOI] [PubMed] [Google Scholar]
- 5.Rothschild L. A new method of measuring sperm speeds. Nature. 1953;171:512–3. doi: 10.1038/171512a0. [DOI] [PubMed] [Google Scholar]
- 6.Katz DF, Dott HM. Methods of measuring swimming speed of spermatozoa. J Reprod Fertil. 1975;45:263–72. doi: 10.1530/jrf.0.0450263. [DOI] [PubMed] [Google Scholar]
- 7.David G, Serres C, Jouannet P. Kinematics of human spermatozoa. Gamete Res. 1981;4:83–95. [Google Scholar]
- 8.Overstreet JW, Katz DF, Hanson FW, Fonseca JR. A simple inexpensive method for objective assessment of human sperm movement characteristics. Fertil Steril. 1979;31:162–72. [PubMed] [Google Scholar]
- 9.Katz DF, Overstreet JW. Sperm motility assessment by videomicrography. Fertil Steril. 1981;35:188–93. doi: 10.1016/s0015-0282(16)45320-3. [DOI] [PubMed] [Google Scholar]
- 10.van der Horst G, Samuels J. A new videographic method with computer analysis for measuring human and bull sperm velocity. S Afr J Sci. 1984;80:144. [Google Scholar]
- 11.Samuels JS, van der Horst G. Sperm motility analysis by means of frame lapse videography. Arch Androl. 1986;17:151–5. doi: 10.3109/01485018608990188. [DOI] [PubMed] [Google Scholar]
- 12.Katz DF, Drobnis EZ, Overstreet JW. Factors regulating mammalian sperm migration through the female reproductive tract and oocyte vestments. Gamete Res. 1989;22:443–69. doi: 10.1002/mrd.1120220410. [DOI] [PubMed] [Google Scholar]
- 13.Davis RO, Siemers RJ. Derivation and reliability of kinematic measures of sperm motion. Reprod Fertil Dev. 1995;7:857–69. doi: 10.1071/rd9950857. [DOI] [PubMed] [Google Scholar]
- 14.Mortimer ST. A critical review of the physiological importance and analysis of sperm movement in mammals. Hum Reprod Update. 1997;3:403–39. doi: 10.1093/humupd/3.5.403. [DOI] [PubMed] [Google Scholar]
- 15.Amann RP, Waberski D. Computer-assisted sperm analysis (CASA): capabilities and potential developments. Theriogenology. 2014;81:5–17. doi: 10.1016/j.theriogenology.2013.09.004. [DOI] [PubMed] [Google Scholar]
- 16.Mortimer D. Objective analysis of sperm motility and kinematics. In: Keel BA, Webster BW, editors. Handbook of the Laboratory Diagnosis and Treatment of Infertility. Boca Raton: CRC Press, Inc; 1990. pp. 97–133. [Google Scholar]
- 17.3rd ed. Cambridge: Cambridge University Press; 1992. World Health Organization. WHO Laboratory Manual for the Examination of Human Semen and Sperm-Cervical Mucus Interaction; p. 107. [Google Scholar]
- 18.Kaskar K, Franken DR, van der Horst G, Kruger TF. The effect of pentoxifylline on sperm movement characteristics and zona pellucida binding potential of teratozoospermic men. Hum Reprod. 1994;9:477–81. doi: 10.1093/oxfordjournals.humrep.a138530. [DOI] [PubMed] [Google Scholar]
- 19.Mortimer D. Practical Laboratory Andrology. New York: Oxford University Press; 1994. p. 393. [Google Scholar]
- 20.Mortimer D, Mortimer ST. Value and reliability of CASA systems. In: Ombelet W, Bosmans E, Vandeput H, Vereecken A, Renier M, et al., editors. Andrology in the Nineties. Modern ART in the 2000's. Carnforth: Parthenon Publishing; 1998. pp. 73–89. [Google Scholar]
- 21.Mortimer D, Aitken RJ, Mortimer ST, Pacey AA. Workshop report: clinical CASA – the quest for consensus. Reprod Fertil Dev. 1995;7:951–9. doi: 10.1071/rd9950951. [DOI] [PubMed] [Google Scholar]
- 22.Consensus workshop on advanced diagnostic andrology techniques. ESHRE (European Society of Human Reproduction and Embryology) Andrology Special Interest Group. Hum Reprod. 1996;11:1463–79. [PubMed] [Google Scholar]
- 23.Guidelines on the application of CASA technology in the analysis of spermatozoa. ESHRE Andrology Special Interest Group. European Society for Human Reproduction and Embryology. Hum Reprod. 1998;13:142–5. [PubMed] [Google Scholar]
- 24.Zinaman MJ, Uhler ML, Vertuno E, Fisher SG, Clegg ED. Evaluation of computer-assisted semen analysis (CASA) with IDENT stain to determine sperm concentration. J Androl. 1996;17:288–92. [PubMed] [Google Scholar]
- 25.Björndahl L. What is normal semen quality. On the use and abuse of reference limits for the interpretation of semen analysis results? Hum Fertil (Camb) 2011;14:179–86. doi: 10.3109/14647273.2011.580823. [DOI] [PubMed] [Google Scholar]
- 26.Björndahl L, Mortimer D, Barratt CL, Castilla JA, Menkveld R, et al. Cambridge: Cambridge University Press; 2010. A Practical Guide to Basic Laboratory Andrology; p. 336. [Google Scholar]
- 27.Palacios ER, Clavero A, Gonzalvo MC, Rosales A, Mozas J, et al. Acceptable variability in external quality assessment programmes for basic semen analysis. Hum Reprod. 2012;27:314–22. doi: 10.1093/humrep/der413. [DOI] [PubMed] [Google Scholar]
- 28.5th ed. Geneva: World Health Organization; 2010. World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; p. 271. [Google Scholar]
- 29.van der Horst G. Unpublished data [Google Scholar]
- 30.Keel BA, Quinn P, Schmidt CF, Jr, Serafy NT, Jr, Serafy NT, Sr, et al. Results of the American Association of Bioanalysts national proficiency testing programme in andrology. Hum Reprod. 2000;15:680–6. doi: 10.1093/humrep/15.3.680. [DOI] [PubMed] [Google Scholar]
- 31.Bailey E, Fenning N, Chamberlain S, Devlin L, Hopkisson J, et al. Validation of sperm counting methods using limits of agreement. J Androl. 2007;28:364–73. doi: 10.2164/jandrol.106.002188. [DOI] [PubMed] [Google Scholar]
- 32.Mortimer D. Unpublished data [Google Scholar]
- 33.Douglas-Hamilton DH, Smith NG, Kuster CE, Vermeiden JP, Althouse GC. Particle distribution in low-volume capillary-loaded chambers. J Androl. 2005a;26:107–14. [PubMed] [Google Scholar]
- 34.Douglas-Hamilton DH, Smith NG, Kuster CE, Vermeiden JP, Althouse GC. Capillary-loaded particle fluid dynamics: effect on estimation of sperm concentration. J Androl. 2005b;26:115–22. [PubMed] [Google Scholar]
- 35.Bland JM, Altman DG. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet. 1986;1:307–10. [PubMed] [Google Scholar]
- 36.Passing H, Bablok W. A new biometrical procedure for testing the equality of measurements from two different analytical methods. Application of linear regression procedures for method comparison studies in clinical chemistry, Part I. J Clin Chem Clin Biochem. 1983;21:709–20. doi: 10.1515/cclm.1983.21.11.709. [DOI] [PubMed] [Google Scholar]
- 37.van der Horst G, Kitchin RM, van der Horst M, Atherton RW. The effect of the breeding season, cryopreservation and physiological extender on selected sperm and semen parameters of four ferret species: implications for captive breeding in the endangered black-footed ferret. Reprod Fertil Dev. 2009;21:351–63. doi: 10.1071/rd08075. [DOI] [PubMed] [Google Scholar]
- 38.Lueders I, Luther I, Scheepers G, van der Horst G. Improved semen collection method for wild felids: urethral catheterization yields high sperm quality in African lions (Panthera leo) Theriogenology. 2012;78:696–701. doi: 10.1016/j.theriogenology.2012.02.026. [DOI] [PubMed] [Google Scholar]
- 39.Mortimer D, Serres C, Mortimer ST, Jouannet P. Influence of image sampling frequency on the perceived movement characteristics of progressively motile human spermatozoa. Gamete Res. 1988;20:313–27. doi: 10.1002/mrd.1120200307. [DOI] [PubMed] [Google Scholar]
- 40.Mortimer ST, Swan MA, Mortimer D. Fractal analysis of capacitating human spermatozoa. Hum Reprod. 1996;11:1049–54. doi: 10.1093/oxfordjournals.humrep.a019295. [DOI] [PubMed] [Google Scholar]
- 41.Mortimer ST. Minimum sperm trajectory length for reliable determination of the fractal dimension. Reprod Fertil Dev. 1998;10:465–9. doi: 10.1071/rd98123. [DOI] [PubMed] [Google Scholar]
- 42.Mortimer ST, Swan MA. The development of smoothing-independent kinematic measures of capacitating human sperm movement. Hum Reprod. 1999a;14:986–96. doi: 10.1093/humrep/14.4.986. [DOI] [PubMed] [Google Scholar]
- 43.Mortimer ST, Swan MA. Effect of image sampling frequency on established and smoothing-independent kinematic values of capacitating human spermatozoa. Hum Reprod. 1999b;14:997–1004. doi: 10.1093/humrep/14.4.997. [DOI] [PubMed] [Google Scholar]
- 44.Mortimer D, Mortimer ST. Laboratory investigation of the infertile male. In: Brinsden PR, editor. A Textbook of In-Vitro Fertilization and Assisted Reproduction. 3rd ed. London: Taylor and Francis Medical Books; 2005. pp. 61–91. [Google Scholar]
- 45.Coetzee K, Kruger TF, Lombard CJ. Repeatability and variance analysis on multiple computer-assisted (IVOS) sperm morphology readings. Andrologia. 1999;31:163–8. doi: 10.1046/j.1439-0272.1999.00257.x. [DOI] [PubMed] [Google Scholar]
- 46.Barratt CL, Björndahl L, Menkveld R, Mortimer D. ESHRE special interest group for andrology basic semen analysis course: a continued focus on accuracy, quality, efficiency and clinical relevance. Hum Reprod. 2011;26:3207–12. doi: 10.1093/humrep/der312. [DOI] [PubMed] [Google Scholar]
- 47.van der Horst G, Maree L. SpermBlue®: a new universal stain for human and animal sperm which is also amenable to automated sperm morphology analysis. Biotechnic Histochem. 2009;84:299–308. doi: 10.3109/10520290902984274. [DOI] [PubMed] [Google Scholar]
- 48.Maree L, du Plessis SS, Menkveld R, van der Horst G. Morphometric dimensions of the human sperm head depend on the staining method used. Hum Reprod. 2010;25:1369–82. doi: 10.1093/humrep/deq075. [DOI] [PubMed] [Google Scholar]
- 49.Mortimer D, Mortimer ST. Computer-aided sperm analysis (CASA) of sperm motility and hyperactivation. In: Carrell DT, Aston KI, editors. Spermatogenesis and Spermiogenesis: Methods and Protocols. Methods in Molecular Biology. Vol. 927. New York: Springer (Humana Press); 2013. pp. 77–87. [DOI] [PubMed] [Google Scholar]
- 50.Maree L, van der Horst G. Quantification and identification of sperm subpopulations using computer-aided sperm analysis and species-specific cut-off values for swimming speed. Biotech Histochem. 2013;88:181–93. doi: 10.3109/10520295.2012.757366. [DOI] [PubMed] [Google Scholar]
- 51.Ntanjana N. MS. Thesis. South Africa: Department of Medical Bioscience, University of the Western Cape; 2015. Hyperactivation in Human Semen and Sperm Subpopulations by Selected Calcium Modulators. [Google Scholar]
- 52.Sukcharoen N, Keith J, Irvine DS, Aitken RJ. Definition of the optimal criteria for identifying hyperactivated human spermatozoa at 25 Hz using in-vitro fertilization as a functional end-point. Hum Reprod. 1995;10:2928–37. doi: 10.1093/oxfordjournals.humrep.a135822. [DOI] [PubMed] [Google Scholar]
- 53.Mortimer ST, Schëväert D, Swan MA, Mortimer D. Quantitative observations of flagellar motility of capacitating human spermatozoa. Hum Reprod. 1997;12:1006–12. doi: 10.1093/humrep/12.5.1006. [DOI] [PubMed] [Google Scholar]
- 54.Burkman LJ. Characterization of hyperactivated motility by human spermatozoa during capacitation: comparison of fertile and oligozoospermic sperm populations. Arch Androl. 1984;13:153–65. doi: 10.3109/01485018408987514. [DOI] [PubMed] [Google Scholar]
- 55.Mortimer ST, Swan MA, Mortimer D. Effect of seminal plasma on capacitation and hyperactivation in human spermatozoa. Hum Reprod. 1998;13:2139–46. doi: 10.1093/humrep/13.8.2139. [DOI] [PubMed] [Google Scholar]
- 56.Mandelbrot BB. New York: WH Freeman and Company; 1983. The Fractal Geometry of Nature. [Google Scholar]
- 57.Katz MJ, George EB. Fractals and the analysis of growth paths. Bull Math Biol. 1985;47:273–86. doi: 10.1007/BF02460036. [DOI] [PubMed] [Google Scholar]
- 58.Mortimer ST, Mortimer D. Kinematics of human spermatozoa incubated under capacitating conditions. J Androl. 1990;11:195–203. [PubMed] [Google Scholar]
- 59.McPartlin LA, Suarez SS, Czaya CA, Hinrichs K, Bedford-Guaus SJ. Hyperactivation of stallion sperm is required for successful in vitro fertilization of equine oocytes. Biol Reprod. 2009;81:199–206. doi: 10.1095/biolreprod.108.074880. [DOI] [PubMed] [Google Scholar]